
Characterization of a Potential Therapeutic Anti-Canine PD-1 Single Domain Antibody Produced in Yeast

 , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Construction and Selection of Canine PD-1 SDAb Library
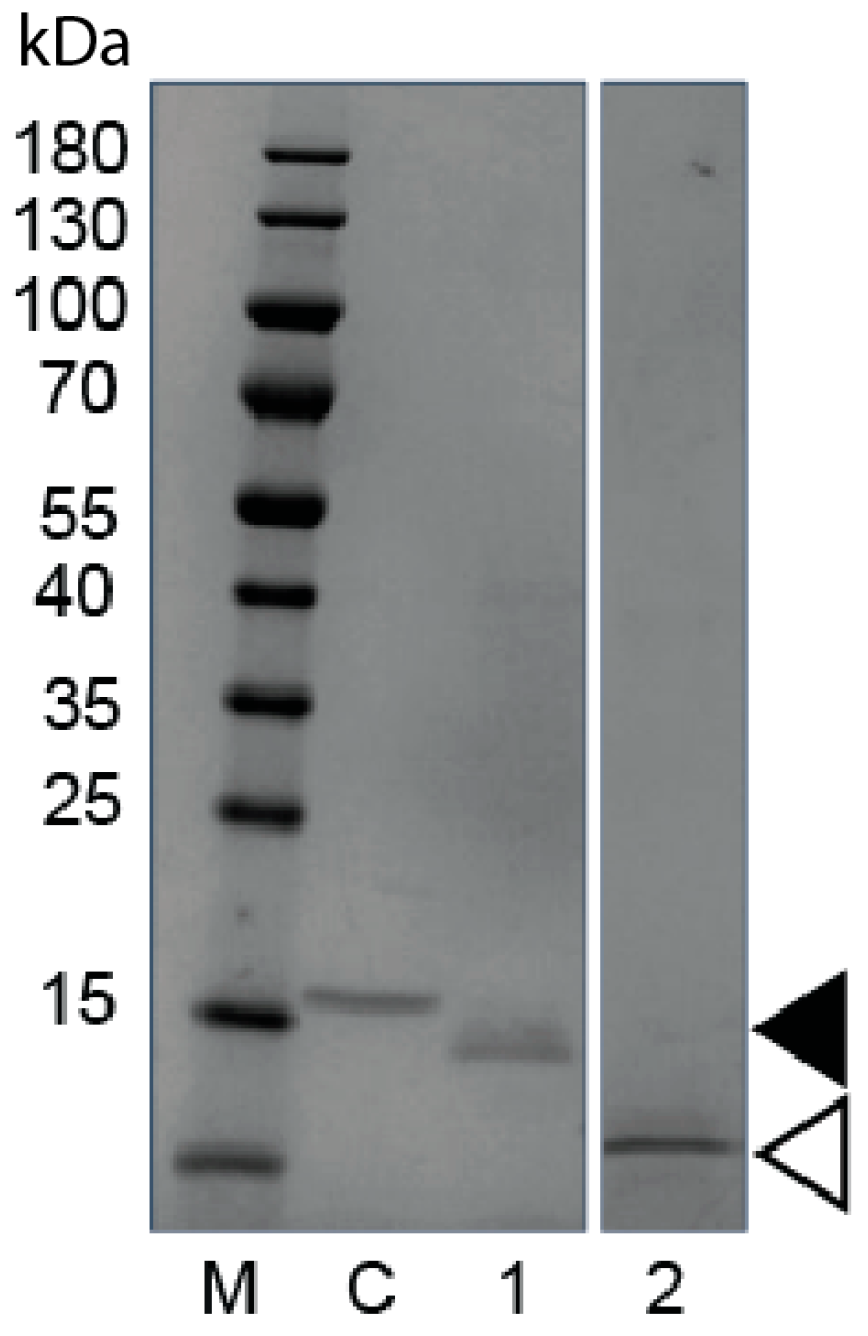
3.2. Alternative Host Expression of an Anti-Canine PD-1 SDAb
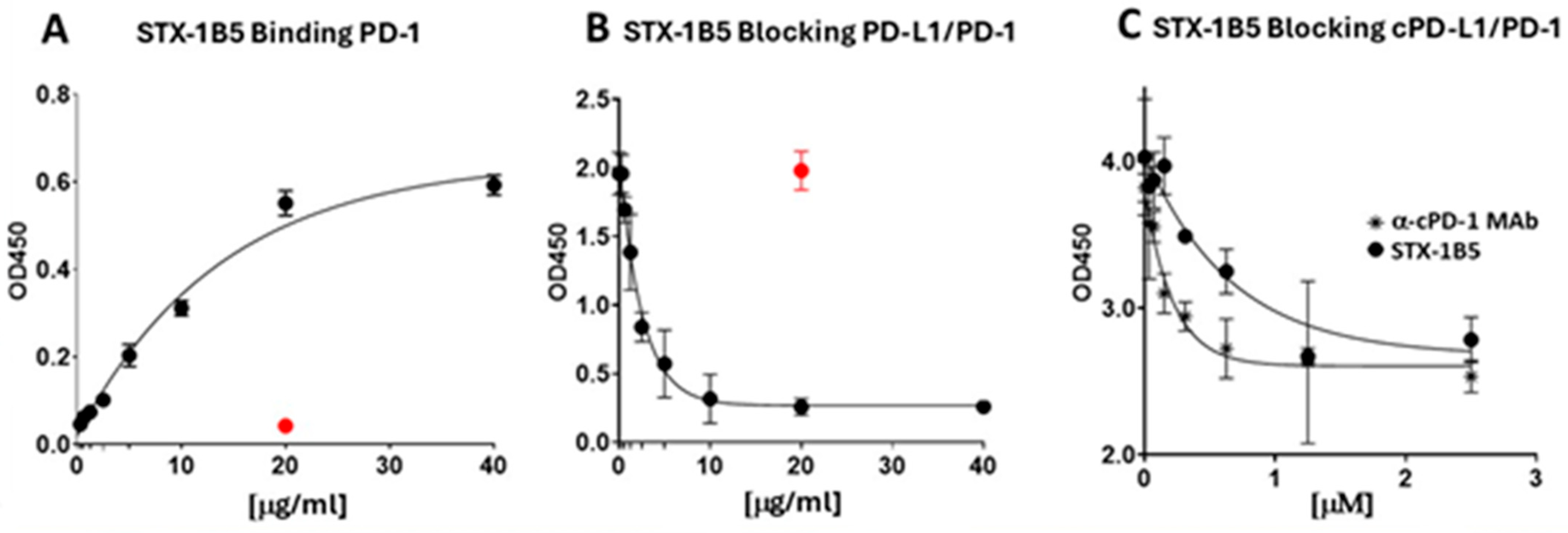
3.3. Affinity and PDL-1 Blocking Binding of STX-1B5 to PD-1 Validated via ELISA
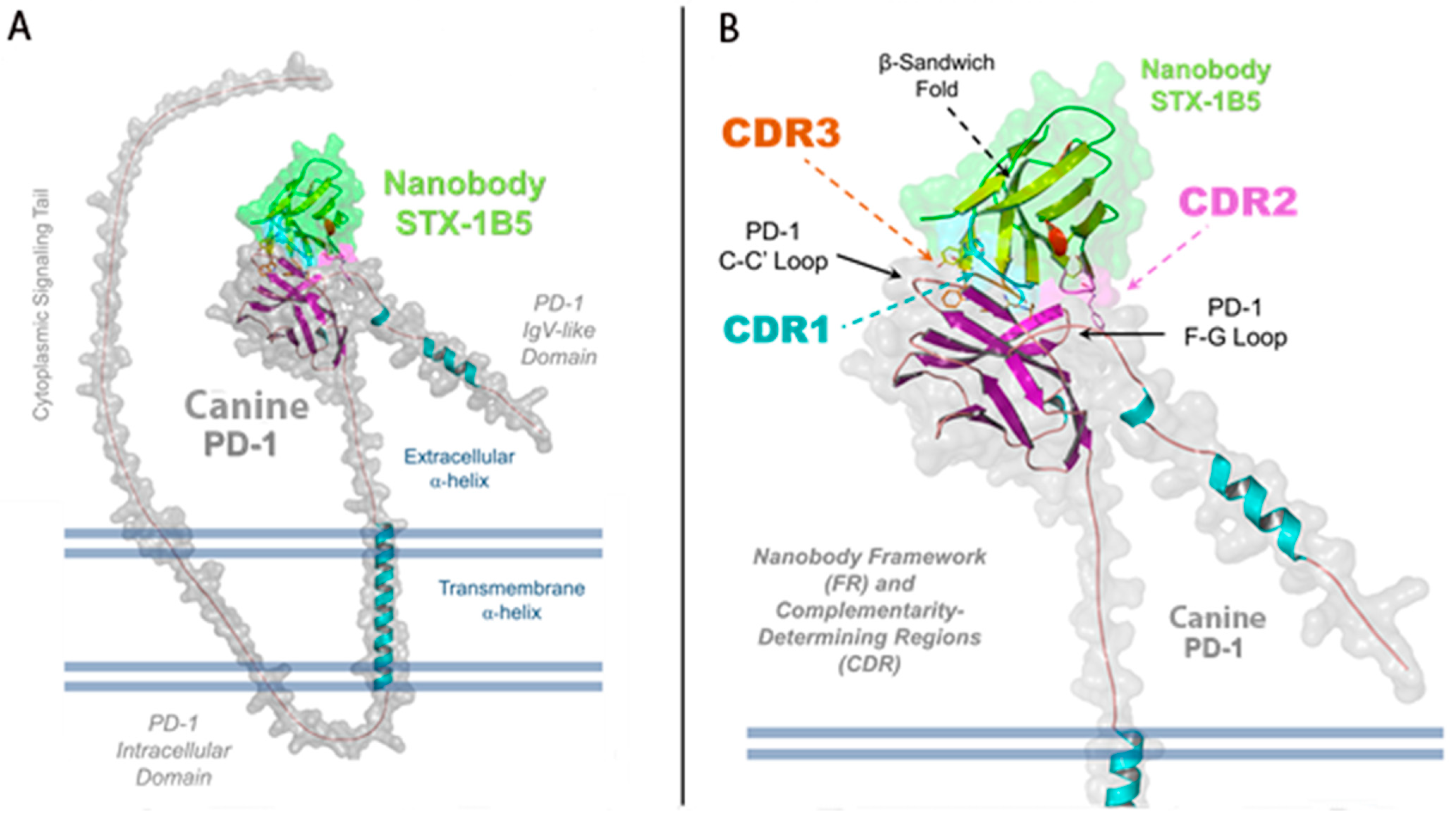
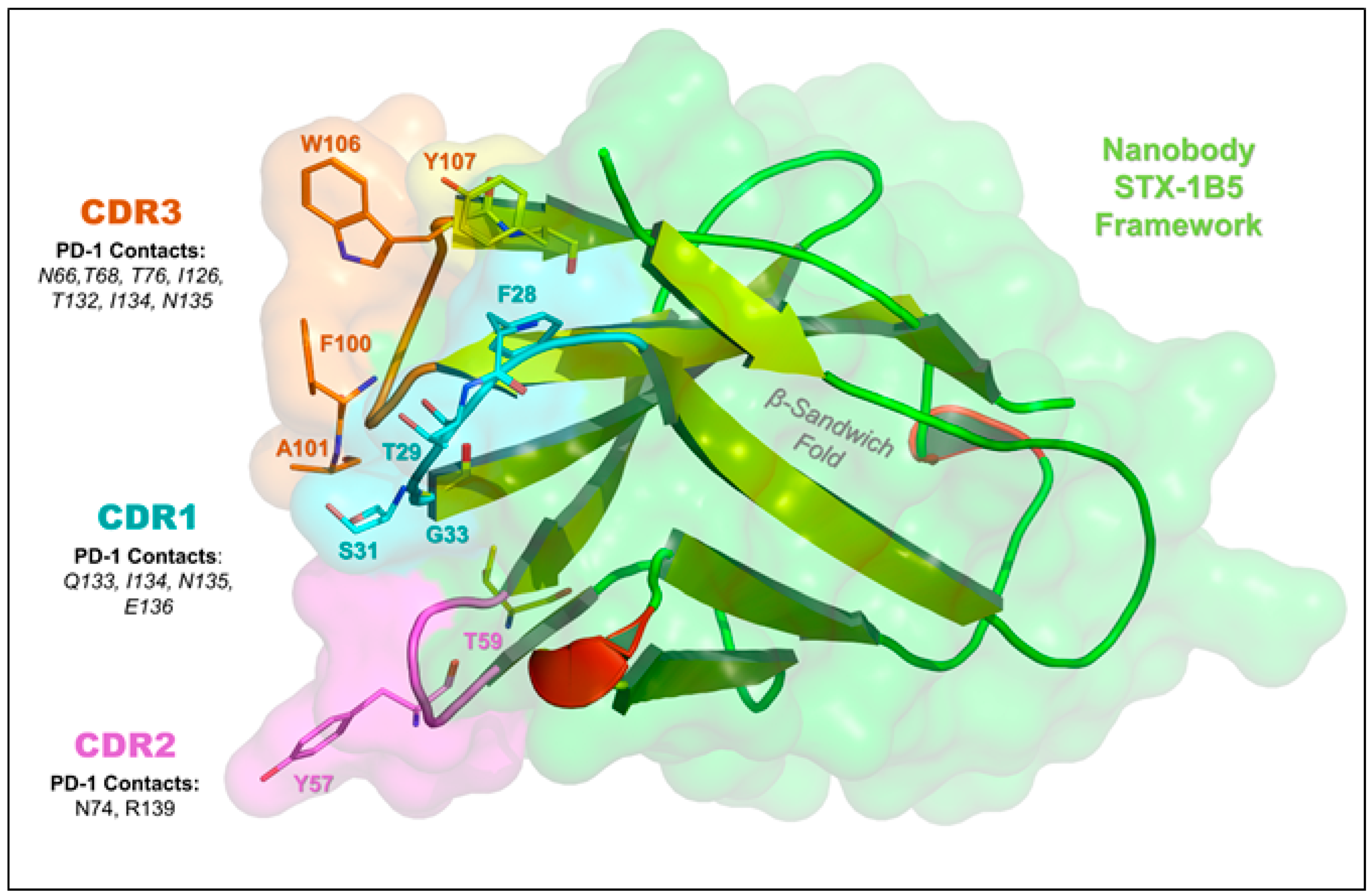
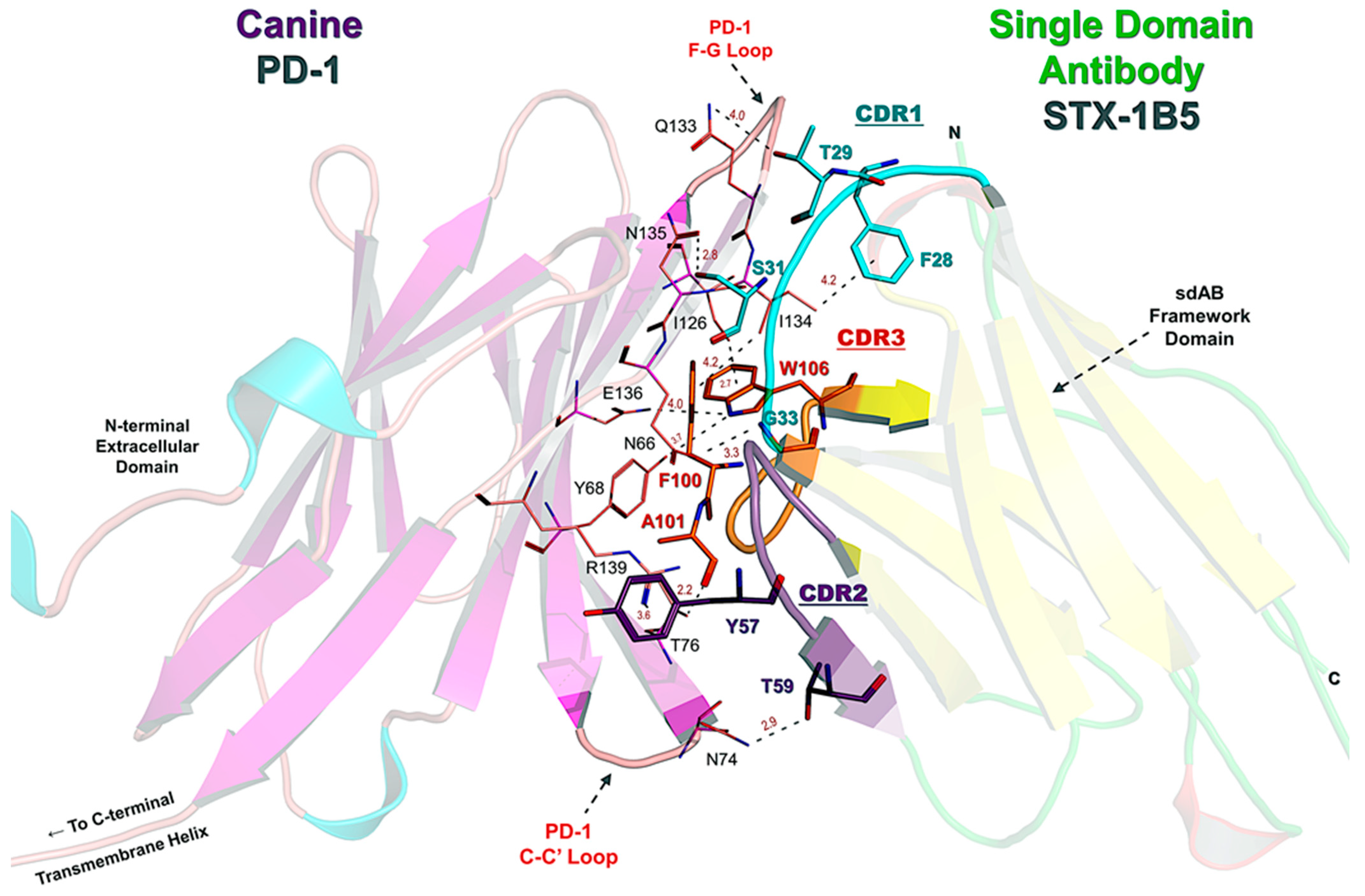
3.4. Nanobody Binding Interface and Overlap with PD-L1/PD-L2 Sites
3.5. Structural Comparisons with Pembrolizumab (Keytruda)
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sarver, A.L.; Makielski, K.M.; DePauw, T.A.; Schulte, A.J.; Modiano, J.F. Increased risk of cancer in dogs and humans: A consequence of recent extension of lifespan beyond evolutionarily-determined limitations? Aging Cancer 2022, 3, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Fleming, J.M.; Creevy, K.E.; Promislow, D.E. Mortality in north American dogs from 1984 to 2004: An investigation into age-, size-, and breed-related causes of death. J. Vet. Intern. Med. 2011, 25, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Rafalko, J.M.; Kruglyak, K.M.; McCleary-Wheeler, A.L.; Goyal, V.; Phelps-Dunn, A.; Wong, L.K.; Warren, C.D.; Brandstetter, G.; Rosentel, M.C.; DiMarzio, L.; et al. Age at cancer diagnosis by breed, weight, sex, and cancer type in a cohort of more than 3000 dogs: Determining the optimal age to initiate cancer screening in canine patients. PLoS ONE 2023, 18, e0280795. [Google Scholar] [CrossRef]
- Johannes, C.M. What You Need to Know About New Cancer Treatments for Dogs. Today’s Vet. Pract. 2022, 12, 42–48. [Google Scholar]
- Hsu, C.Y.; Pallathadka, H.; Jasim, S.A.; Rizaev, J.; Olegovich, B.D.; Hjazi, A.; Mahajan, S.; Mustafa, Y.F.; Husseen, B.; Jawad, M.A. Innovations in cancer immunotherapy: A comprehensive overview of recent breakthroughs and future directions. Crit. Rev. Oncol. Hematol. 2025, 206, 104588. [Google Scholar] [CrossRef] [PubMed]
- Lamba, N.; Ott, P.A.; Iorgulescu, J.B. Use of First-Line Immune Checkpoint Inhibitors and Association with Overall Survival Among Patients with Metastatic Melanoma in the Anti-PD-1 Era. JAMA Netw. Open 2022, 5, e2225459. [Google Scholar] [CrossRef]
- Miller, S.R.; Schipper, M.; Fritsche, L.G.; Jiang, R.; Strohbehn, G.; Ötleş, E.; McMahon, B.H.; Crivelli, S.; Zamora-Resendiz, R.; Ramnath, N.; et al. Pan-Cancer Survival Impact of Immune Checkpoint Inhibitors in a National Healthcare System. Cancer Med. 2024, 13, e70379. [Google Scholar] [CrossRef]
- Yoo, S.-K.; Fitzgerald, C.W.; Cho, B.A.; Fitzgerald, B.G.; Han, C.; Koh, E.S.; Pandey, A.; Sfreddo, H.; Crowley, F.; Korostin, M.R.; et al. Prediction of checkpoint inhibitor immunotherapy efficacy for cancer using routine blood tests and clinical data. Nat. Med. 2025, 31, 869–880. [Google Scholar] [CrossRef]
- Raskov, H.; Orhan, A.; Christensen, J.P.; Gögenur, I. Cytotoxic CD8+ T cells in cancer and cancer immunotherapy. Br. J. Cancer 2021, 124, 359–367. [Google Scholar] [CrossRef]
- Diskin, B.; Adam, S.; Cassini, M.F.; Sanchez, G.; Liria, M.; Aykut, B.; Buttar, C.; Li, E.; Sundberg, B.; Salas, R.D.; et al. PD-L1 engagement on T cells promotes self-tolerance and suppression of neighboring macrophages and effector T cells in cancer. Nat. Immunol. 2020, 21, 442–454. [Google Scholar] [CrossRef]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar] [PubMed]
- Cha, J.-H.; Chan, L.-C.; Li, C.-W.; Hsu, J.L.; Hung, M.-C. Mechanisms Controlling PD-L1 Expression in Cancer. Mol. Cell 2019, 76, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Zheng, P. Tumor cells versus host immune cells: Whose PD-L1 contributes to PD-1/PD-L1 blockade mediated cancer immunotherapy? Cell Biosci. 2018, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Vathiotis, I.A.; Gomatou, G.; Stravopodis, D.J.; Syrigos, N. Programmed Death-Ligand 1 as a Regulator of Tumor Progression and Metastasis. Int. J. Mol. Sci. 2021, 22, 5383. [Google Scholar] [CrossRef]
- Lurain, K.; Ramaswami, R.; Yarchoan, R.; Uldrick, T.S. Anti-PD-1 and Anti-PD-L1 Monoclonal Antibodies in People Living with HIV and Cancer. Curr. HIV/AIDS Rep. 2020, 17, 547–556. [Google Scholar] [CrossRef]
- Choi, J.W.; Withers, S.S.; Chang, H.; Spanier, J.A.; De La Trinidad, V.L.; Panesar, H.; Fife, B.T.; Sciammas, R.; Sparger, E.E.; Moore, P.F.; et al. Development of canine PD-1/PD-L1 specific monoclonal antibodies and amplification of canine T cell function. PLoS ONE 2020, 15, e0235518. [Google Scholar] [CrossRef]
- Maekawa, N.; Konnai, S.; Nishimura, M.; Kagawa, Y.; Takagi, S.; Hosoya, K.; Ohta, H.; Kim, S.; Okagawa, T.; Izumi, Y.; et al. PD-L1 immunohistochemistry for canine cancers and clinical benefit of anti-PD-L1 antibody in dogs with pulmonary metastatic oral malignant melanoma. npj Precis. Oncol. 2021, 5, 10. [Google Scholar] [CrossRef]
- Minoli, L.; Licenziato, L.; Kocikowski, M.; Cino, M.; Dziubek, K.; Iussich, S.; Fanelli, A.; Morello, E.; Martano, M.; Hupp, T.; et al. Development of Monoclonal Antibodies Targeting Canine PD-L1 and PD-1 and Their Clinical Relevance in Canine Apocrine Gland Anal Sac Adenocarcinoma. Cancers 2022, 14, 6188. [Google Scholar] [CrossRef]
- Song, M.-Y.; Cho, J.; Park, H.; Song, Y.; Kim, K.; Ahn, J.-H.; Lee, C.-M.; Kim, D.H.; Ko, H.-J. Discovery and functional characterization of canine PD-L1-targeted antibodies for evaluating antitumor efficacy in a canine osteosarcoma xenograft model. Sci. Rep. 2025, 15, 7574. [Google Scholar] [CrossRef]
- Giuliano, A.; Pimentel, P.A.B.; Horta, R.S. Checkpoint Inhibitors in Dogs: Are We There Yet? Cancers 2024, 16, 2003. [Google Scholar] [CrossRef]
- Igase, M.; Nemoto, Y.; Itamoto, K.; Tani, K.; Nakaichi, M.; Sakurai, M.; Sakai, Y.; Noguchi, S.; Kato, M.; Tsukui, T.; et al. A pilot clinical study of the therapeutic antibody against canine PD-1 for advanced spontaneous cancers in dogs. Sci. Rep. 2020, 10, 18311. [Google Scholar] [CrossRef] [PubMed]
- Kocikowski, M.; Dziubek, K.; Węgrzyn, K.; Hrabal, V.; Zavadil-Kokas, F.; Vojtesek, B.; Alfaro, J.A.; Hupp, T.; Parys, M. Comparative characterization of two monoclonal antibodies targeting canine PD-1. Front. Immunol. 2024, 15, 1382576. [Google Scholar] [CrossRef] [PubMed]
- Alejandra, W.-P.; Irene, J.-P.M.; Antonio, G.-S.F.; Patricia, R.-G.R.; Elizabeth, T.-A.; Aleman-Aguilar, J.P.; Rebeca, G.-V. Production of monoclonal antibodies for therapeutic purposes: A review. Int. Immunopharmacol. 2023, 120, 110376. [Google Scholar] [CrossRef]
- Schlör, A.; Hirschberg, S.; Amor, G.B.; Meister, T.L.; Arora, P.; Pöhlmann, S.; Hoffmann, M.; Pfaender, S.; Eddin, O.K.; Kamhieh-Milz, J.; et al. SARS-CoV-2 neutralizing camelid heavy-chain-only antibodies as powerful tools for diagnostic and therapeutic applications. Front. Immunol. 2022, 13, 930975. [Google Scholar] [CrossRef]
- Arbabi-Ghahroudi, M. Camelid Single-Domain Antibodies: Promises and Challenges as Lifesaving Treatments. Int. J. Mol. Sci. 2022, 23, 5009. [Google Scholar] [CrossRef]
- Hoey, R.J.; Eom, H.; Horn, J.R. Structure and development of single domain antibodies as modules for therapeutics and diagnostics. Exp. Biol. Med. 2019, 244, 1568–1576. [Google Scholar] [CrossRef]
- Nessler, I.; Khera, E.; Vance, S.; Kopp, A.; Qiu, Q.; Keating, T.A.; Abu-Yousif, A.O.; Sandal, T.; Legg, J.; Thompson, L.; et al. Increased Tumor Penetration of Single-Domain Antibody-Drug Conjugates Improves In Vivo Efficacy in Prostate Cancer Models. Cancer Res. 2020, 80, 1268–1278. [Google Scholar] [CrossRef]
- Rossotti, M.A.; Bélanger, K.; Henry, K.A.; Tanha, J. Immunogenicity and humanization of single-domain antibodies. FEBS J. 2022, 289, 4304–4327. [Google Scholar] [CrossRef]
- de Marco, A. Recombinant expression of nanobodies and nanobody-derived immunoreagents. Protein Expr. Purif. 2020, 172, 105645. [Google Scholar] [CrossRef]
- Love, K.R.; Dalvie, N.C.; Love, J.C. The yeast stands alone: The future of protein biologic production. Curr. Opin. Biotechnol. 2018, 53, 50–58. [Google Scholar] [CrossRef]
- Bustamante-Cordova, L.; Melgoza-Gonzalez, E.A.; Hernandez, J. Recombinant Antibodies in Veterinary Medicine: An Update. Front. Vet. Sci. 2018, 5, 175. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Huang, H. Expression of single-domain antibody in different systems. Appl. Microbiol. Biotechnol. 2018, 102, 539–551. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.N.; Rekhadevi, P.V.; Ramesh, A. Comparative evaluation of different cell lysis and extraction methods for studying benzo(a)pyrene metabolism in HT-29 colon cancer cell cultures. Cell. Physiol. Biochem. 2011, 28, 209–218. [Google Scholar] [CrossRef]
- Sabir, J.S.; Atef, A.; El-Domyati, F.M.; Edris, S.; Hajrah, N.; Alzohairy, A.M.; Bahieldin, A. Construction of naive camelids VHH repertoire in phage display-based library. Comptes Rendus Biol. 2014, 337, 244–249. [Google Scholar] [CrossRef]
- Pardon, E.; Laeremans, T.; Triest, S.; Rasmussen, S.G.F.; Wohlkönig, A.; Ruf, A.; Muyldermans, S.; Hol, W.G.J.; Kobilka, B.K.; Steyaert, J. A general protocol for the generation of Nanobodies for structural biology. Nat. Protoc. 2014, 9, 674–693. [Google Scholar] [CrossRef]
- Bobrovnik, S.A. Determination of antibody affinity by ELISA. Theory. J. Biochem. Biophys. Methods 2003, 57, 213–236. [Google Scholar] [CrossRef]
- Crowell, L.E.; Goodwine, C.; Holt, C.S.; Rocha, L.; Vega, C.; Rodriguez, S.A.; Dalvie, N.C.; Tracey, M.K.; Puntel, M.; Wigdorovitz, A.; et al. Development of a platform process for the production and purification of single-domain antibodies. Biotechnol. Bioeng. 2021, 118, 3348–3358. [Google Scholar] [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef]
- Goddard, T.D.; Huang, C.C.; Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Sci. 2018, 27, 14–25. [Google Scholar] [CrossRef]
- Schrodinger, LLC. The PyMOL Molecular Graphics System; Version 3.1.0; Schrödinger, LLC.: New York, NY, USA, 2025. [Google Scholar]
- Mattanovich, D.; Branduardi, P.; Dato, L.; Gasser, B.; Sauer, M.; Porro, D. Recombinant protein production in yeasts. Methods Mol. Biol. 2012, 824, 329–358. [Google Scholar]
- Baghban, R.; Farajnia, S.; Ghasemi, Y.; Mortazavi, M.; Zarghami, N.; Samadi, N. New Developments in Pichia pastoris Expression System, Review and Update. Curr. Pharm. Biotechnol. 2018, 19, 451–467. [Google Scholar] [CrossRef] [PubMed]
- Mahal, H.; Branton, H.; Farid, S.S. End-to-end continuous bioprocessing: Impact on facility design, cost of goods, and cost of development for monoclonal antibodies. Biotechnol. Bioeng. 2021, 118, 3468–3485. [Google Scholar] [CrossRef] [PubMed]
- Matthews, C.B.; Wright, C.; Kuo, A.; Colant, N.; Westoby, M.; Love, J.C. Reexamining opportunities for therapeutic protein production in eukaryotic microorganisms. Biotechnol. Bioeng. 2017, 114, 2432–2444. [Google Scholar] [CrossRef] [PubMed]
- Brady, J.R.; Love, J.C. Alternative hosts as the missing link for equitable therapeutic protein production. Nat. Biotechnol. 2021, 39, 404–407. [Google Scholar] [CrossRef]
- Lin, D.Y.W.; Tanaka, Y.; Iwasaki, M.; Gittis, A.G.; Su, H.P.; Mikami, B.; Okazaki, T.; Honjo, T.; Minato, N.; Garboczi, D.N. The PD-1/PD-L1 complex resembles the antigen-binding Fv domains of antibodies and T cell receptors. Proc. Natl. Acad. Sci. USA 2008, 105, 3011–3016. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Candidate SDAb | MW (kDa) | Overall pI | Variable Region pI | Overall GRAVY |
|---|---|---|---|---|
| 1B5 | 12.3 | 7.94 | 3.67 | −0.42 |
| 5A5 | 12.2 | 9.04 | 9.96 | −0.31 |
| 4B2 | 12.4 | 6.80 | 3.80 | −0.27 |
| 5A1 | 12.7 | 4.44 | 3.17 | −0.40 |
| 4B4 | 12.8 | 5.78 | 4.50 | −0.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vijayasimha, K.; Annalora, A.J.; Mourich, D.V.; Ruby, C.E.; Dolan, B.P.; Crowell, L.; Le, V.H.M.; Larson, M.K.; Bracha, S.; Cebra, C.K. Characterization of a Potential Therapeutic Anti-Canine PD-1 Single Domain Antibody Produced in Yeast. Vet. Sci. 2025, 12, 649. https://doi.org/10.3390/vetsci12070649
Vijayasimha K, Annalora AJ, Mourich DV, Ruby CE, Dolan BP, Crowell L, Le VHM, Larson MK, Bracha S, Cebra CK. Characterization of a Potential Therapeutic Anti-Canine PD-1 Single Domain Antibody Produced in Yeast. Veterinary Sciences. 2025; 12(7):649. https://doi.org/10.3390/vetsci12070649
Chicago/Turabian StyleVijayasimha, Kartikeya, Andrew J. Annalora, Dan V. Mourich, Carl E. Ruby, Brian P. Dolan, Laura Crowell, Vu Ha Minh Le, Maureen K. Larson, Shay Bracha, and Christopher K. Cebra. 2025. "Characterization of a Potential Therapeutic Anti-Canine PD-1 Single Domain Antibody Produced in Yeast" Veterinary Sciences 12, no. 7: 649. https://doi.org/10.3390/vetsci12070649
APA StyleVijayasimha, K., Annalora, A. J., Mourich, D. V., Ruby, C. E., Dolan, B. P., Crowell, L., Le, V. H. M., Larson, M. K., Bracha, S., & Cebra, C. K. (2025). Characterization of a Potential Therapeutic Anti-Canine PD-1 Single Domain Antibody Produced in Yeast. Veterinary Sciences, 12(7), 649. https://doi.org/10.3390/vetsci12070649