
Comparative Analysis of Sectional Anatomy, Computed Tomography and Magnetic Resonance of the Cadaveric Six-Banded Armadillo (Euphractus sexcintus) Head

 ,
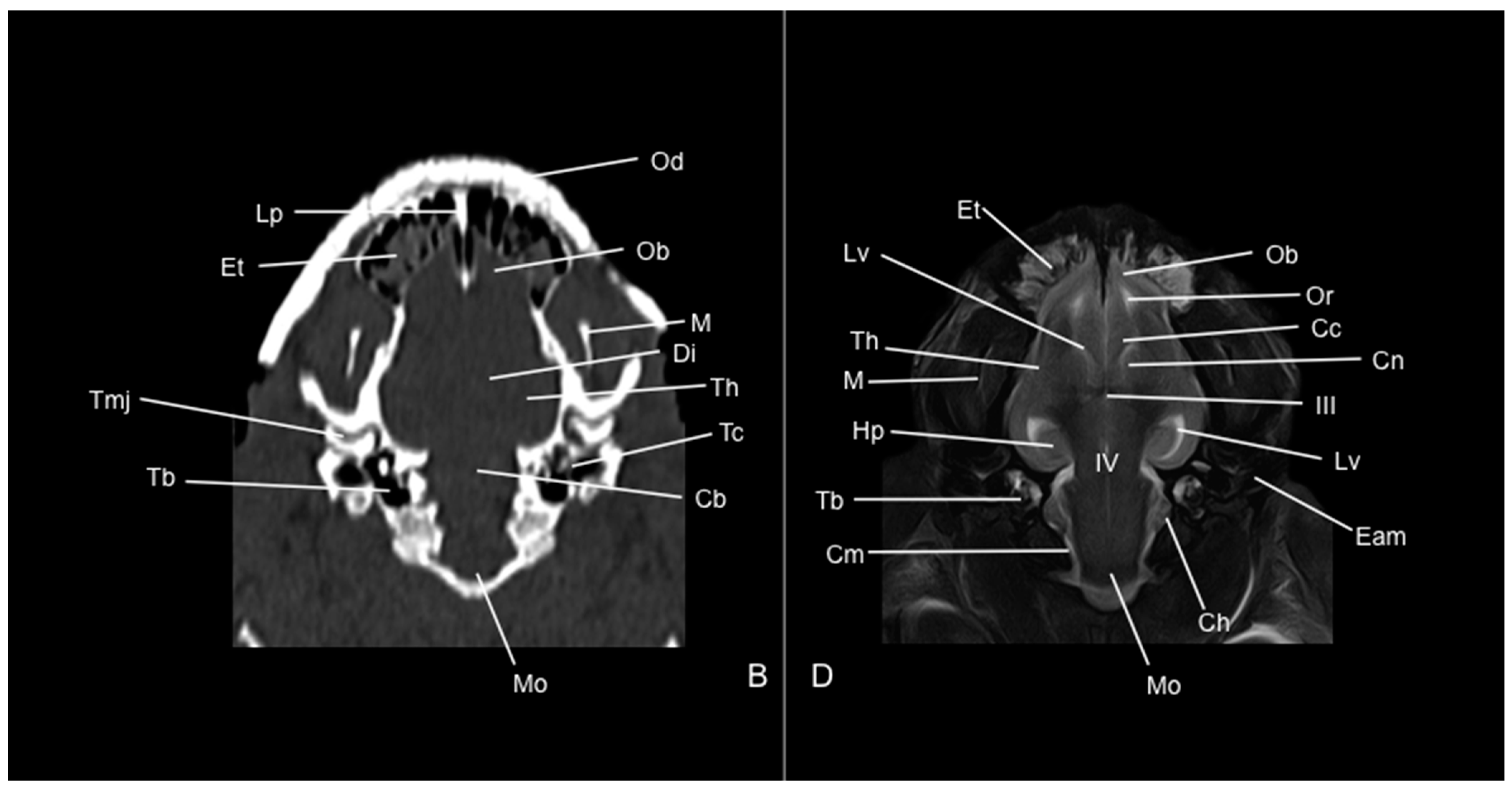
,  , , ,
, , ,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. CT Technique
2.3. MRI Technique
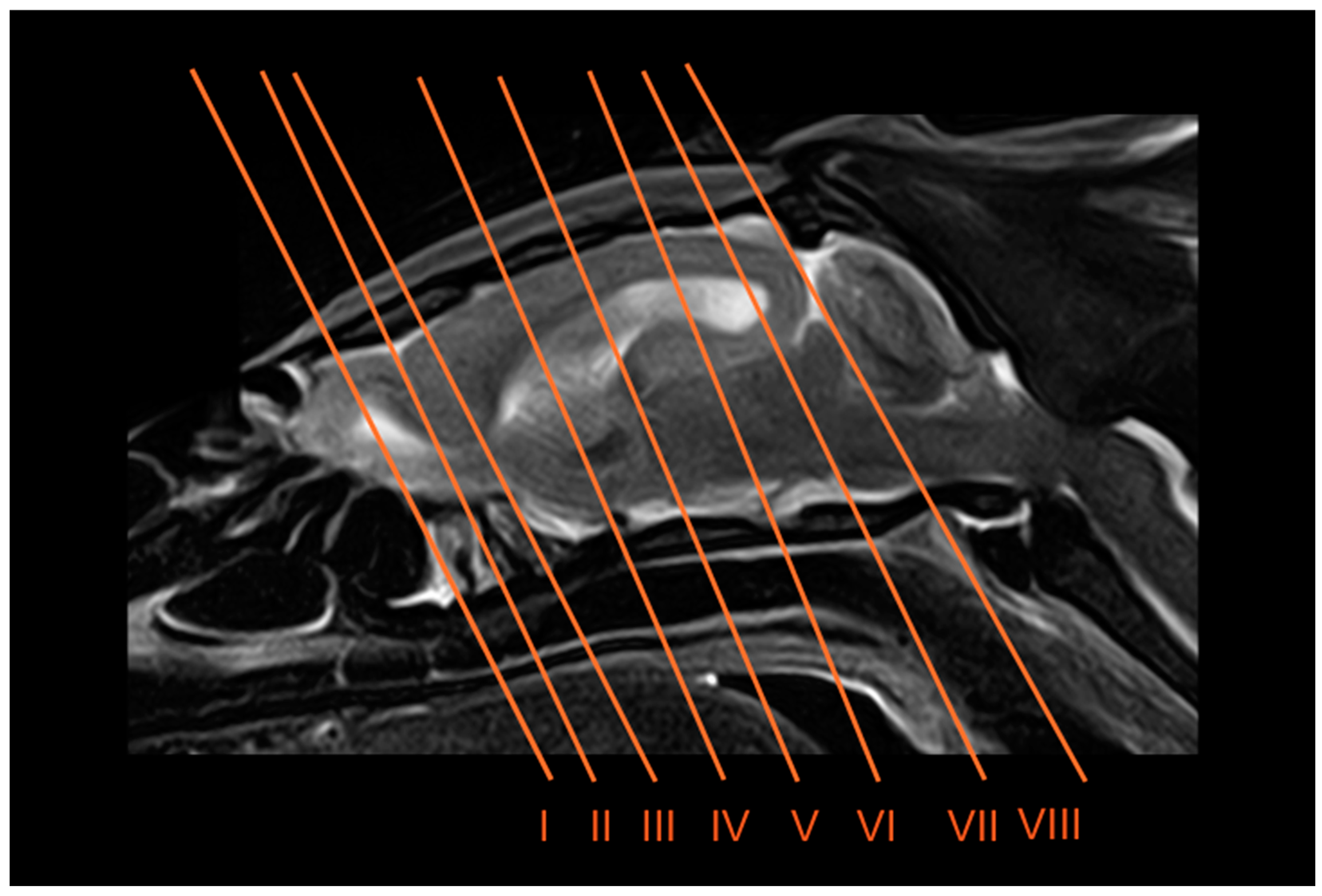
2.4. Anatomical Sections
2.5. Anatomic Evaluation
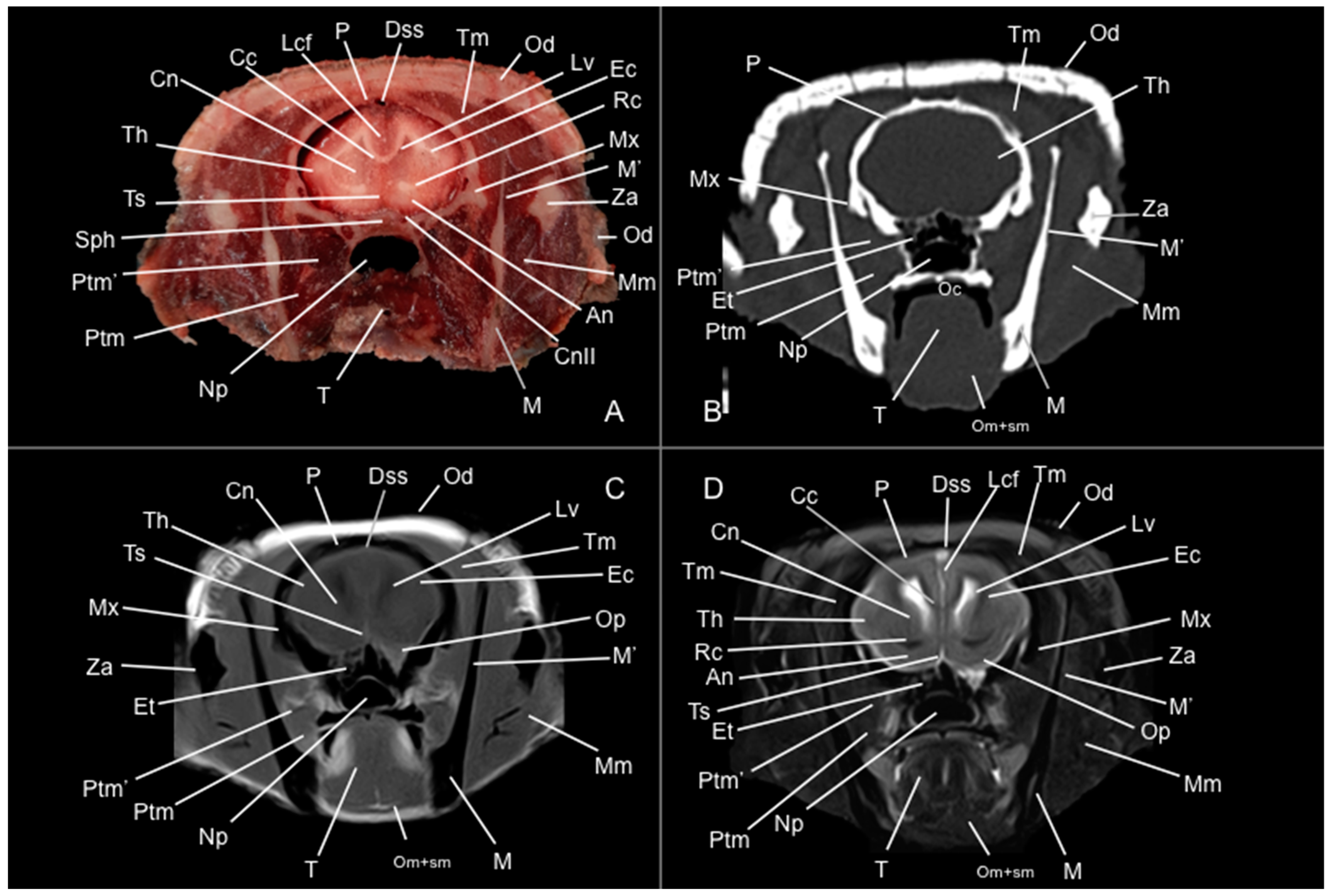
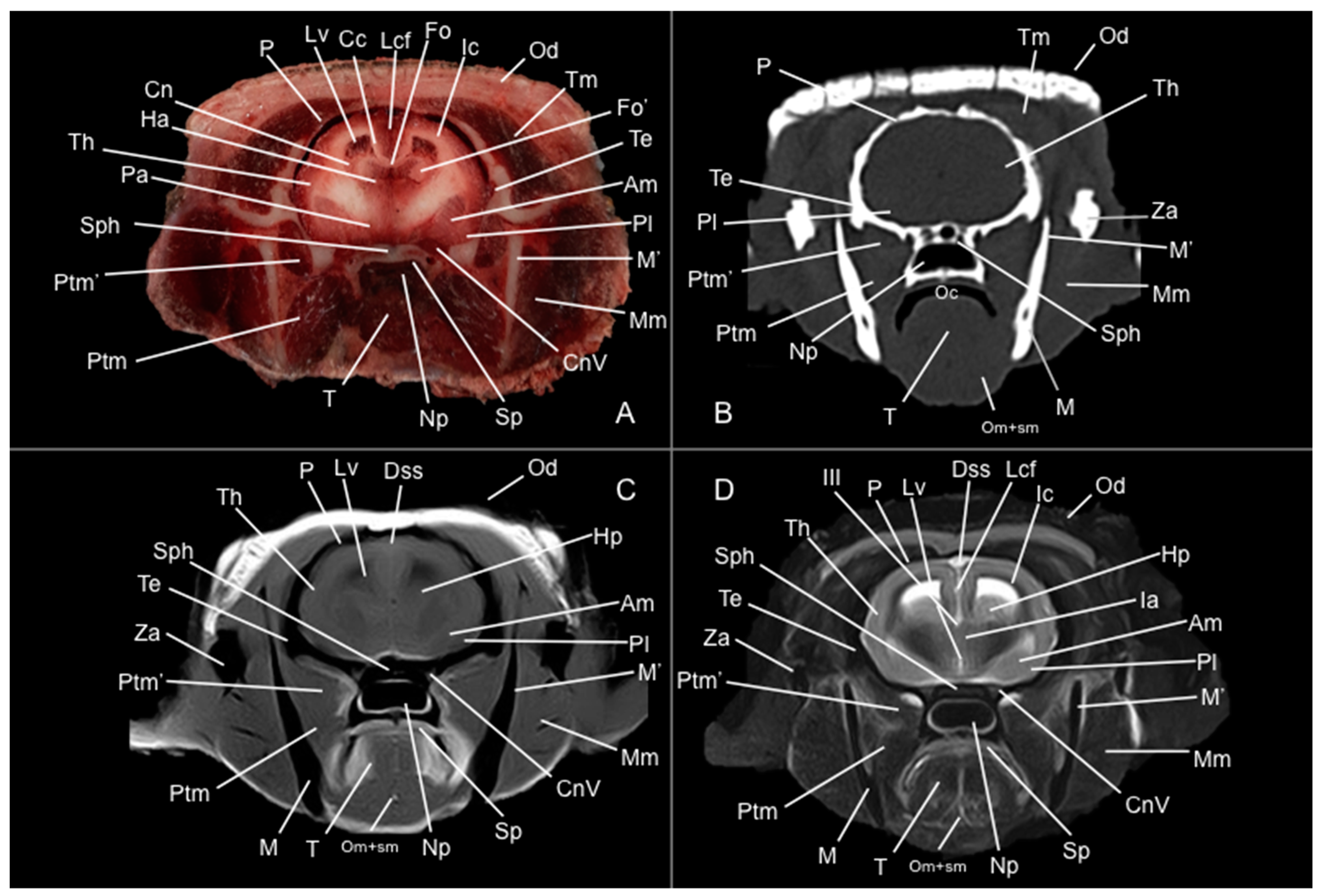
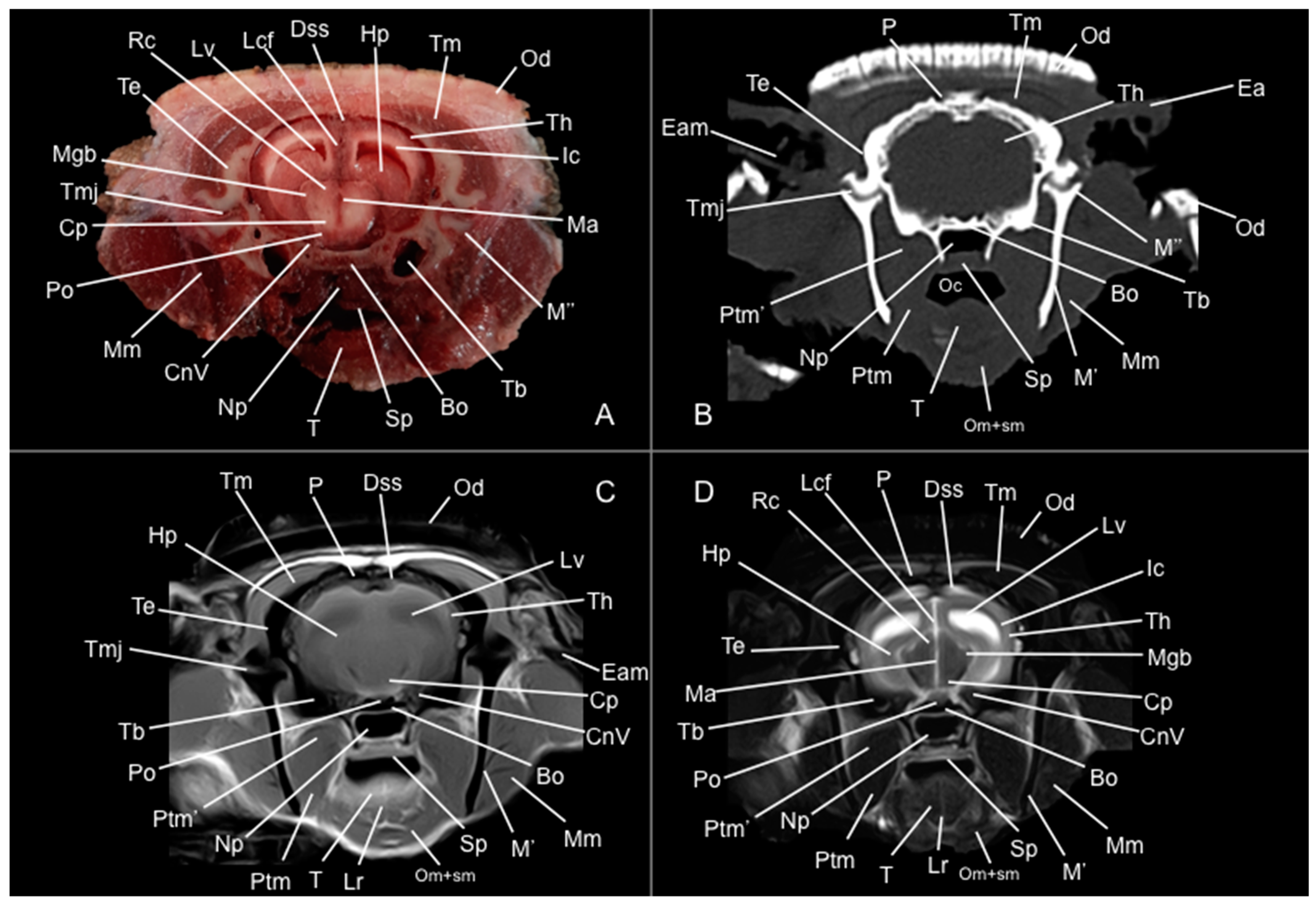
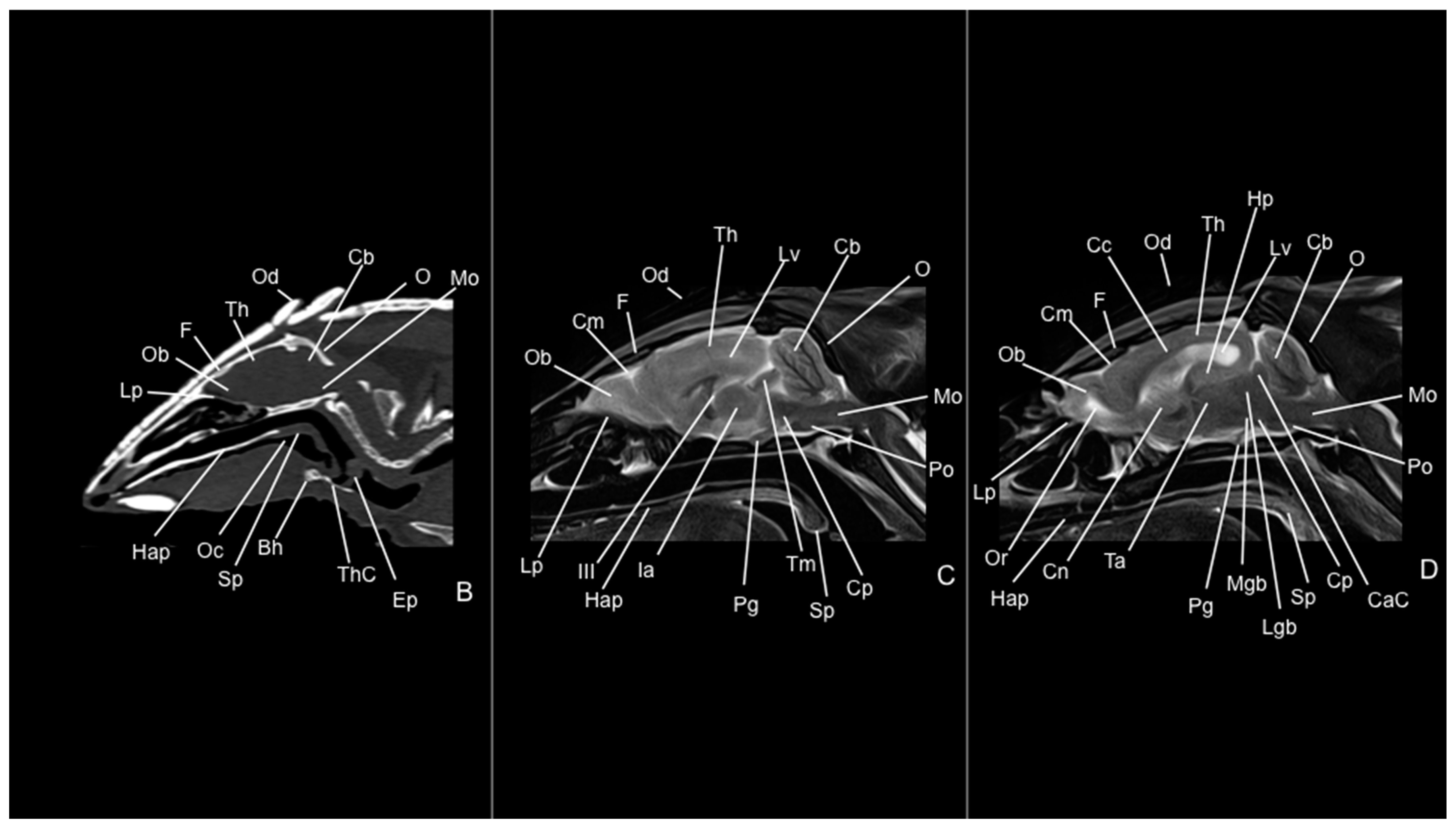
3. Results
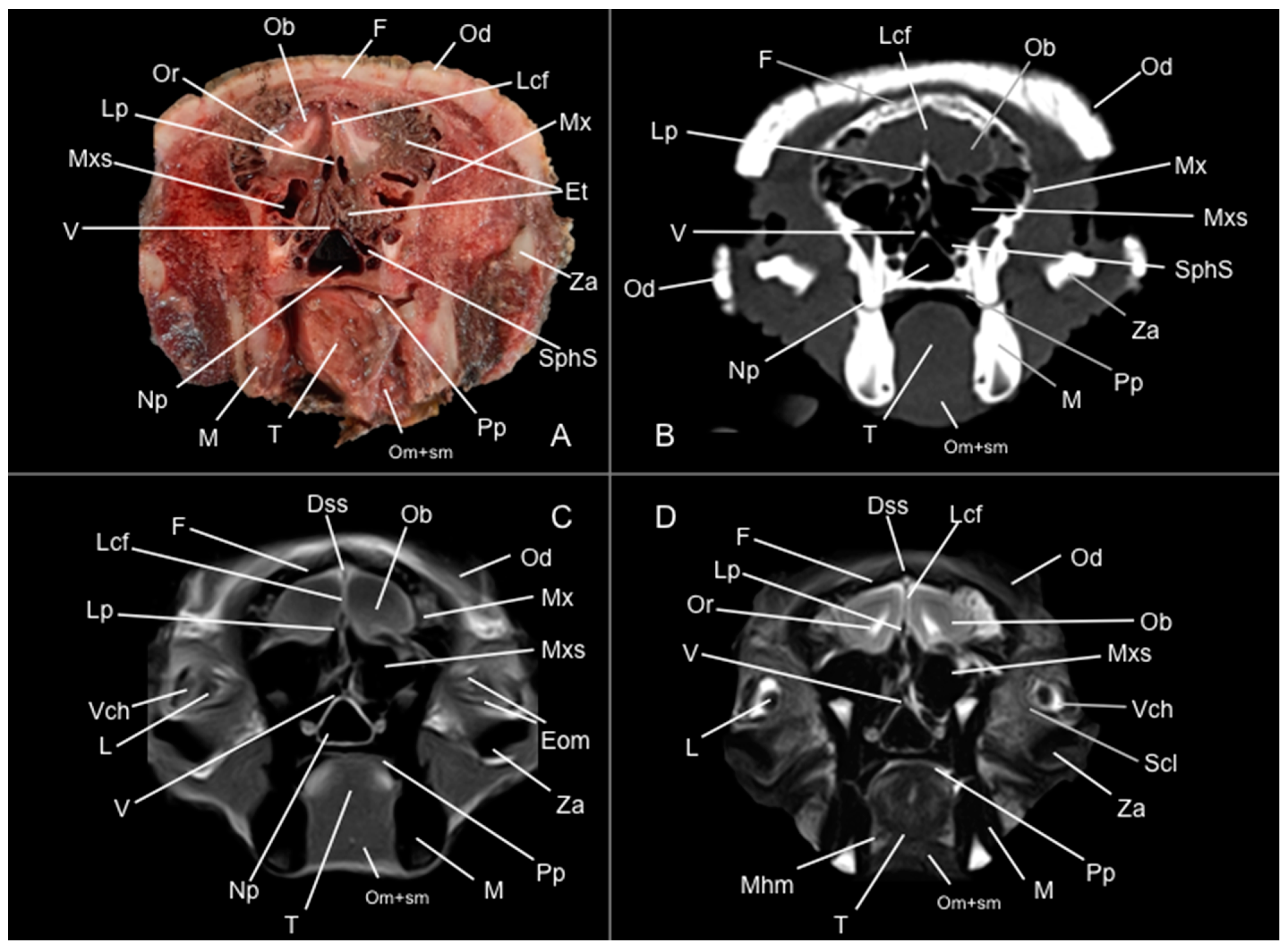
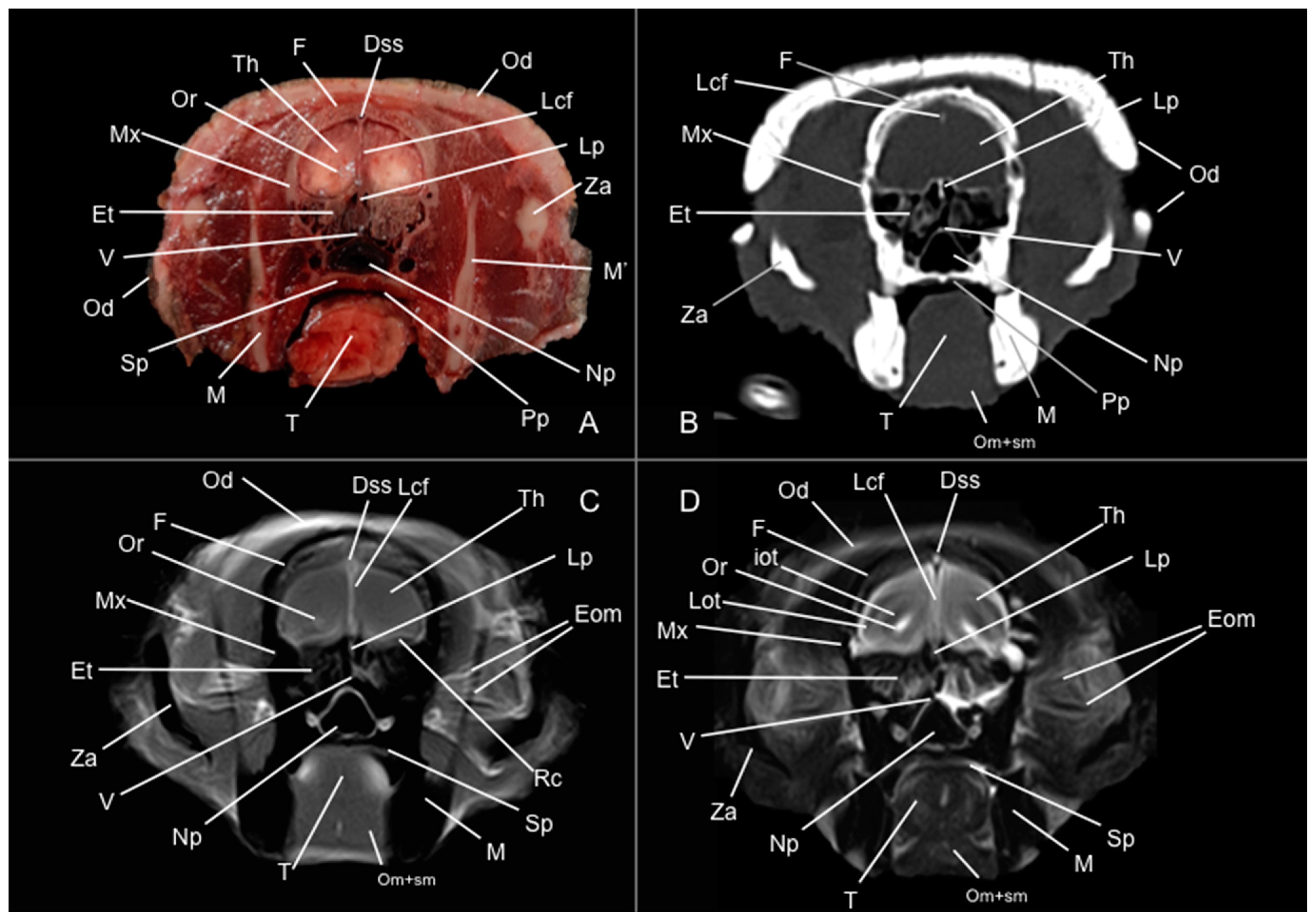
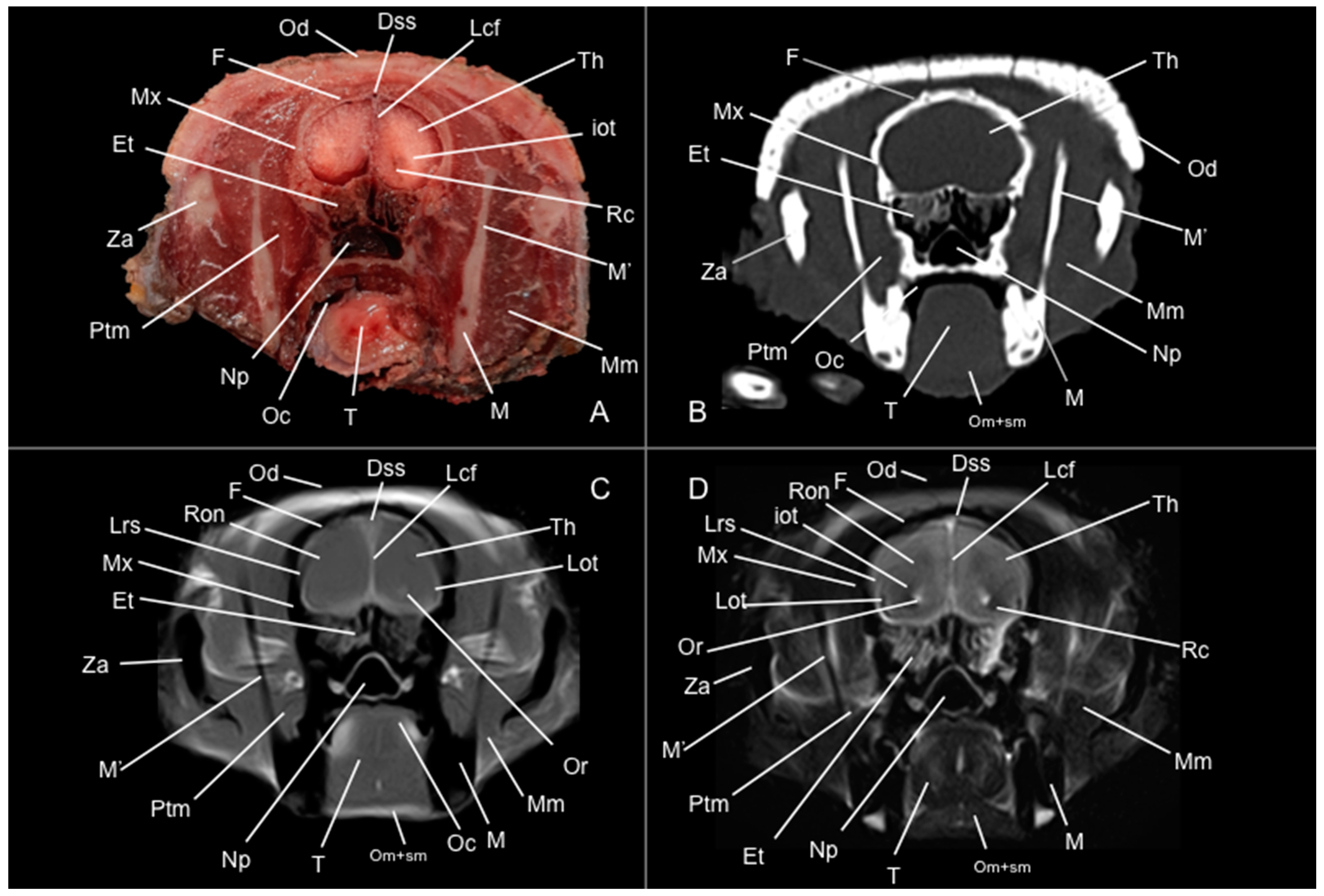
3.1. Anatomical Study
3.2. Computed Tomography Study
3.3. Magnetic Resonance Imaging (MRI)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gardner, A.L. Order Cingulata. In Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; Wilson, D.E., Reeder, D.M., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2005; p. 97. ISBN 978-0-8018-8221-0. [Google Scholar]
- Redford, K.H.; Wetzel, R.M. Euphractus sexcinctus. Mamm. Species 1985, 252, 1–4. [Google Scholar] [CrossRef]
- Brittany, B. Euphractus sexcinctus. Available online: https://animaldiversity.org/accounts/Euphractus_sexcinctus/ (accessed on 20 November 2024).
- Farrow, C.S. Veterinary Diagnostic Imaging: Birds, Exotic Pets and Wildlife; Mosby Elsevier: St. Louis, MO, USA, 2009. [Google Scholar]
- Lauridsen, H.; Hansen, K.; Wang, T.; Agger, P.; Andersen, J.L.; Knudsen, P.S.; Rasmussen, A.S.; Uhrenholt, L.; Pedersen, M. Inside out: Modern Imaging Techniques to Reveal Animal Anatomy. PLoS ONE 2011, 6, e17879. [Google Scholar] [CrossRef]
- Behroozi, M.; Billings, B.K.; Helluy, X.; Manger, P.R.; Güntürkün, O.; Ströckens, F. Functional MRI in the Nile Crocodile: A New Avenue for Evolutionary Neurobiology. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180178. [Google Scholar] [CrossRef]
- Knipe, M.F. Principles of Neurological Imaging of Exotic Animal Species. Vet. Clin. N. Am. Exot. Anim. Pract. 2007, 10, 893–907. [Google Scholar] [CrossRef] [PubMed]
- Banzato, T.; Hellebuyck, T.; Van Caelenberg, A.; Saunders, J.H.; Zotti, A. A Review of Diagnostic Imaging of Snakes and Lizards. Vet. Rec. 2013, 173, 43–49. [Google Scholar] [CrossRef]
- Morales Bordon, D.; Suárez-Cabrera, F.; Ramírez, G.; Paz-Oliva, P.; Morales-Espino, A.; Arencibia, A.; Encinoso, M.; Ventura, M.R.; Jaber, J.R. Study of the Normal Crested Porcupine (Hystrix cristata) Nasal Cavity and Paranasal Sinuses by Cross-Sectional Anatomy and Computed Tomography. Vet. Sci. 2024, 11, 611. [Google Scholar] [CrossRef] [PubMed]
- Morales-Bordon, D.; Encinoso, M.; Arencibia, A.; Jaber, J.R. Cranial Investigations of Crested Porcupine (Hystrix cristata) by Anatomical Cross-Sections and Magnetic Resonance Imaging. Animals 2023, 13, 2551. [Google Scholar] [CrossRef]
- Fumero-Hernández, M.; Encinoso, M.; Melian, A.; Nuez, H.A.; Salman, D.; Jaber, J.R. Cross Sectional Anatomy and Magnetic Resonance Imaging of the Juvenile Atlantic Puffin Head (Aves, Alcidae, Fratercula arctica). Animals 2023, 13, 3434. [Google Scholar] [CrossRef]
- Gaudin, T.J.; Biewener, A.A. The functional morphology of xenarthrous vertebrae in the armadillo Dasypus novemcinctus (Mammalia, Xenarthra). J. Morphol. 1992, 214, 63–81. [Google Scholar] [CrossRef]
- Oliver, J.D.; Jones, K.E.; Pierce, S.E.; Hautier, L. Size and shape regional differentiation during the development of the spine in the nine-banded armadillo (Dasypus novemcinctus). Evol. Dev. 2021, 23, 496–512. [Google Scholar] [CrossRef]
- Alves, L.S.; Babicsak, V.R.; Charlier, M.G.S.; Vulcano, L.C. Radiography, computed tomography and 3D reconstruction of the pelvis in the nine-banded armadillo, Dasypus novemcinctus. In Proceedings of the 40th World Small Animal Veterinary Association Congress, Bangkok, Thailand, 15–18 May 2015; Volume 40, pp. 66–67. [Google Scholar]
- Vizcaıno, S.F.; Milne, N. Structure and function in armadillo limbs (Mammalia: Xenarthra: Dasypodidae). J. Zool. 2002, 257, 117–127. [Google Scholar] [CrossRef]
- Wible, J.R. Petrosal anatomy of the nine-banded armadillo, Dasypus novemcintus Linnaeus, 1758 (Mammalia, Xenarthra, Dasypodidae). Ann. Carnegie Mus. 2010, 79, 1–28. [Google Scholar] [CrossRef]
- Alves, L.S.; Midon, M.; Filadelpho, A.L.; Vulcano, L.C.; Knotek, Z. Gross Osteology, Radiographic and Computed Tomographic Morphology of the Axial Skeleton of the Nine-Banded Armadillo (Dasypus novemcinctus). Anat. Histol. Embryol. 2017, 46, 162–177. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.B.; Sousa, M.M.; Silva, J.V.; Oliveira, I.M.; Barbosa, C.M.; Santos, M.; Conde, A.M. Anatomy of the nasal cavity of nine-banded armadillo (Dasypus novemcinctus, Linnaeus, 1758). J. Interdiscip. Biocienc. 2016, 1, 1–4. [Google Scholar] [CrossRef]
- Billet, G.; Hautier, L.; de Thoisy, B.; Delsuc, F. The hidden anatomy of paranasal sinuses reveals biogeographically distinct morphotypes in the nine-banded armadillo (Dasypus novemcinctus). PeerJ 2017, 5, e3593. [Google Scholar] [CrossRef]
- Fonseca, C.M.B.; da Silva, A.B.S.; Cavalcante, M.M.A.S.; de Oliveira, I.M.; Moura, S.M.S.; Cunha, B.M.; Leite, C.M.C.; de Carvalho, M.A.M.; Moura, W.R.A.; Rizzo, M.D.S.; et al. Morphology of laryngeal cartilage of the nine-banded armadillo (Dasypus novemcinctus) Linnaeus, 1758. Microsc. Res. Tech. 2017, 80, 1089–1095. [Google Scholar] [CrossRef]
- Vizcaíno, S.F.; Fariña, R.A.; Bargo, M.S.; De Iuliis, G. Functional and phylogenetic assessment of the masticatory adaptations in Cingulata (Mammalia, Xenarthra). Rev. Asos. Paleontol. Argent. 2004, 41, 651–664. [Google Scholar]
- Alves, L.S.; Oliva, L.R.; Charlier, M.G.S.; Bonatelli, S.P.; Inamassu, L.R.; Vulcano, L.C.; Teixeira, C.R. Fratura em seguimento lombar da coluna vertebral em um tatugalinha (Dasypus novemcinctus). An. Simp. Intern. Diag. Imag. SINDIV 2013, 3, 57–58. [Google Scholar]
- Boyde, A.; Mills, D.; Abba, A.M.; Ezquiaga, M.C. Fleas and lesions in armadillo osteoderms. J. Anat. 2023, 242, 1029–1036. [Google Scholar] [CrossRef]
- Jaber, J.R.; Morales Bordon, D.; Arencibia, A.; Corbera, J.A.; Conde-Felipe, M.; Ayala, M.D.; Encinoso, M. Correlation between Cross-Sectional Anatomy and Computed Tomography of the Normal Six-Banded Armadillo (Euphractus sexcintus) Nasal Cavity and Paranasal Sinuses. Animals 2024, 14, 1135. [Google Scholar] [CrossRef]
- Scholl, B.; Rylee, J.; Luci, J.J.; Priebe, N.J.; Padberg, J. Orientation selectivity in the visual cortex of the nine-banded armadillo. J. Neurophysiol. 2017, 117, 1395–1406. [Google Scholar] [CrossRef] [PubMed]
- Jacqmot, O.; Van Thielen, B.; Hespel, A.-M.; Luijten, P.R.; de Mey, J.; Van Binst, A.; Provyn, S.; Tresignie, J. T2-Weighted Turbo Spin-Echo Magnetic Resonance Imaging of Canine Brain Anatomy at 1.5T, 3T, and 7T Field Strengths. Anat. Rec. 2022, 305, 222–233. [Google Scholar] [CrossRef]
- Araújo, J.V.S.; de Sousa Cavalcante, M.M.A.; de Jesus Gonçalves, P.C.; Guerra, S.P.L.; Da Silva, A.B.S.; Conde Júnior, A.M. Descriptive macroscopic anatomy of the central nervous system six-banded armadillo (Euphractus sexcintus, Linnaeus, 1758) and nine-banded armadillo (Dasypus novemcinctus, Linnaeus, 1758). J. Interdiscip. Biociênc. 2015, 1, 13. [Google Scholar] [CrossRef]
- González Rodríguez, E.; Encinoso Quintana, M.; Morales Bordon, D.; Garcés, J.G.; Artiles Nuez, H.; Jaber, J.R. Anatomical Description of Rhinoceros Iguana (Cyclura cornuta cornuta) Head by Computed Tomography, Magnetic Resonance Imaging and Gross-Sections. Animals 2023, 13, 955. [Google Scholar] [CrossRef] [PubMed]
- Arencibia, A.; Vazquez, J.M.; Rivero, M.; Latorre, R.; Sandoval, J.A.; Vilar, J.M.; Ramirez, J.A. Computed tomography of normal cranioencephalic structures in two horses. Anat. Histol. Embryol. 2000, 29, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Mahdy, M. Correlation between computed tomography, magnetic resonance imaging and cross-sectional anatomy of the head of the guinea pig (Cavia porcellus, Linnaeus 1758). Anat. Histol. Embryol. 2022, 51, 51–61. [Google Scholar] [CrossRef]
- De Rycke, L.M.; Saunders, J.H.; Gielen, I.M.; van Bree, H.J.; Simoens, P.J. Magnetic resonance imaging, computed tomography, and cross-sectional views of the anatomy of normal nasal cavities and paranasal sinuses in mesaticephalic dogs. Am. J. Vet. Res. 2003, 64, 1093–1098. [Google Scholar] [CrossRef]
- Van Caelenberg, A.I.; De Rycke, L.M.; Hermans, K.; Verhaert, L.; Van Bree, H.J.; Gielen, I.M. Low-field magnetic resonance imaging and cross-sectional anatomy of the rabbit head. Vet. J. 2011, 188, 83–91. [Google Scholar] [CrossRef]
- Ferrari, C.C.; Carmanchahi, P.D.; Aldana Marcos, H.J.; Affanni, J.M. Ultrastructural Characterisation of the Olfactory Mucosa of the Armadillo Dasypus hybridus (Dasypodidae, Xenarthra). J. Anat. 2000, 196, 269–278. [Google Scholar] [CrossRef]
- Carmanchahi, P.D.; Aldana Marcos, H.J.; Ferrari, C.C.; Affanni, J.M. The Vomeronasal Organ of the South American Armadillo Chaetophractus Villosus (Xenarthra, Mammalia): Anatomy, Histology and Ultrastructure. J. Anat. 1999, 195, 587–604. [Google Scholar] [CrossRef]
- Basso, A.P.; Sidorkewicj, N.S.; Casanave, E.B.; Mason, M.J. The Middle Ear of the Pink Fairy Armadillo Chlamyphorus truncatus (Xenarthra, Cingulata, Chlamyphoridae): Comparison with Armadillo Relatives Using Computed Tomography. J. Anat. 2020, 236, 809–826. [Google Scholar] [CrossRef]
- Ruby, J.R.; Allen, E.R. Ultrastructure of the Salivary Bladder of the Nine-Banded Armadillo. Cell Tissue Res. 1976, 169, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Le Verger, K.; González Ruiz, L.R.; Billet, G. Comparative Anatomy and Phylogenetic Contribution of Intracranial Osseous Canals and Cavities in Armadillos and Glyptodonts (Xenarthra, Cingulata). J. Anat. 2021, 239, 1473–1502. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.A.; Harlow, H.J.; McArthur, N.H.; Ralph, C.L. Epithalamus of the Nine-Banded Armadillo, Dasypus Novemcinctus. Comp. Biochem. Physiol. A Comp. Physiol. 1986, 85, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Squarcia, S.M.; Sidorkewicj, N.S.; Camina, R.; Casanave, E.B. Sexual dimorphism in the mandible of the armadillo Chaetophractus villosus (Desmarest, 1804) (Dasypodidae) from northern Patagonia, Argentina. Braz. J. Biol. 2009, 69, 347–352. [Google Scholar] [CrossRef]
- Sigurdsson, B.; Hauglund, N.L.; Lilius, T.O.; Mogensen, F.L.-H.; Mortensen, K.N.; Beschorner, N.; Klinger, L.; Bærentzen, S.L.; Rosenholm, M.P.; Shalgunov, V.; et al. A SPECT-Based Method for Dynamic Imaging of the Glymphatic System in Rats. J. Cereb. Blood Flow Metab. 2023, 43, 1153–1165. [Google Scholar] [CrossRef]
- Mrzílková, J.; Patzelt, M.; Gallina, P.; Wurst, Z.; Šeremeta, M.; Dudák, J.; Krejčí, F.; Žemlička, J.; Musil, V.; Karch, J.; et al. Imaging of Mouse Brain Fixated in Ethanol in Micro-CT. Biomed Res. Int. 2019, 2019, 2054262. [Google Scholar] [CrossRef]
- Boscaini, A.; Iurino, D.A.; Billet, G.; Hautier, L.; Sardella, R.; Tirao, G.; Gaudin, T.J.; Pujos, F. Phylogenetic and Functional Implications of the Ear Region Anatomy of Glossotherium robustum (Xenarthra, Mylodontidae) from the Late Pleistocene of Argentina. Sci. Nat. 2018, 105, 28. [Google Scholar] [CrossRef]
- Hautier, L.; Billet, G.; de Thoisy, B.; Delsuc, F. Beyond the Carapace: Skull Shape Variation and Morphological Systematics of Long-Nosed Armadillos (Genus Dasypus). PeerJ 2017, 5, e3650. [Google Scholar] [CrossRef]
- Feijó, A.; Patterson, B.D.; Cordeiro-Estrela, P. Taxonomic Revision of the Long-Nosed Armadillos, Genus Dasypus Linnaeus, 1758 (Mammalia, Cingulata). PLoS ONE 2018, 13, e0195084. [Google Scholar] [CrossRef]
- Craven, B.A.; Paterson, E.G.; Settles, G.S. The fluid dynamics of canine olfaction: Unique nasal airflow patterns as an explanation of macrosmia. J. Roy Soc. Interface 2010, 7, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Eiting, T.P.; Smith, T.D.; Perot, J.B.; Dumont, E.R. The role of the olfactory recess in olfactory airflow. J. Exp. Biol. 2014, 217, 1799–1803. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, C.; Aldana, H.; Carmanchahi, P.D.; Benítez, I.; Affanni, J.M. The brain of the armadillo Dasypus hybridus. A general view of its most salient features. Biocell 1998, 22, 123–140. [Google Scholar]
- Sisson, S.; Grossman, J.D. Anatomía de los Animales Domésticos; Salvat Editores: Madrid, Spain, 1972. [Google Scholar]
- Köning, J.F.; Klippel, R.A. The Rat Brain: An Stereotaxic Atlas; R. Krieger Publishing Co. Inc: New York, NY, USA, 1967. [Google Scholar]
- Ariëns Kappers, C.U.; Huber, G.C.; Crosby, E.C. The Comparative Anatomy of the Nervous System of Vertebrates Including the Man Vol II; Hafner Publishing Company: New York, NY, USA, 1960. [Google Scholar]
- Herrera, A.; Pereira, J.; Bonadia, S.; Mamprim, M.J.; Schimming, B.C. Computed tomography and cross-sectional anatomy of the head in the giant anteater (Myrmecophaga tridactyla). Anat. Histol. Embryol. 2021, 50, 614–624. [Google Scholar]
- Santos, I.B.; Fonseca, G.A.; Rigueira, S.E.; Machado, R.B. The rediscovery of Brazilian three banded armadillos and notes on its conservation status. In: Da Fonseca, G. A.; Leitte, Y.L.; Santos, I. Edentata. A news letter of the IUCN Edentate Specialist Group. Edentata 1994, 1, 11–14. [Google Scholar] [CrossRef]
- Kot, B.C.W.; Tsui, H.C.L.; Chung, T.Y.T.; Lau, A.P.Y. Postmortem Neuroimaging of Cetacean Brains Using Computed Tomography and Magnetic Resonance Imaging. Front. Mar. Sci. 2020, 7, 544037. [Google Scholar] [CrossRef]
- Diniz, L.S.; Costa, E.O.; Oliveira, P.M. Clinical disorders in armadillos (Dasypodidae, Edentata) in captivity. J. Vet. Med. Ser. B 1997, 44, 577–582. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jaber, J.R.; Morales-Bordon, D.; Morales, M.; Paz-Oliva, P.; Encinoso, M.; Morales, I.; Roldan-Medina, N.; Ramírez Zarzosa, G.; Morales-Espino, A.; Ros, A.; et al. Comparative Analysis of Sectional Anatomy, Computed Tomography and Magnetic Resonance of the Cadaveric Six-Banded Armadillo (Euphractus sexcintus) Head. Vet. Sci. 2025, 12, 433. https://doi.org/10.3390/vetsci12050433
Jaber JR, Morales-Bordon D, Morales M, Paz-Oliva P, Encinoso M, Morales I, Roldan-Medina N, Ramírez Zarzosa G, Morales-Espino A, Ros A, et al. Comparative Analysis of Sectional Anatomy, Computed Tomography and Magnetic Resonance of the Cadaveric Six-Banded Armadillo (Euphractus sexcintus) Head. Veterinary Sciences. 2025; 12(5):433. https://doi.org/10.3390/vetsci12050433
Chicago/Turabian StyleJaber, José Raduan, Daniel Morales-Bordon, Manuel Morales, Pablo Paz-Oliva, Mario Encinoso, Inmaculada Morales, Natalia Roldan-Medina, Gregorio Ramírez Zarzosa, Alejandro Morales-Espino, Alvaro Ros, and et al. 2025. "Comparative Analysis of Sectional Anatomy, Computed Tomography and Magnetic Resonance of the Cadaveric Six-Banded Armadillo (Euphractus sexcintus) Head" Veterinary Sciences 12, no. 5: 433. https://doi.org/10.3390/vetsci12050433
APA StyleJaber, J. R., Morales-Bordon, D., Morales, M., Paz-Oliva, P., Encinoso, M., Morales, I., Roldan-Medina, N., Ramírez Zarzosa, G., Morales-Espino, A., Ros, A., & Conde-Felipe, M. M. (2025). Comparative Analysis of Sectional Anatomy, Computed Tomography and Magnetic Resonance of the Cadaveric Six-Banded Armadillo (Euphractus sexcintus) Head. Veterinary Sciences, 12(5), 433. https://doi.org/10.3390/vetsci12050433