
Lipid Dysmetabolism in Canine Chronic Liver Disease: Relationship Between Clinical, Histological and Immunohistochemical Features

 , , ,
, , , 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals: Enrollment and Subgrouping
2.2. Histology and Immunohistochemistry
2.3. Statistical Analysis
3. Results
3.1. Animals
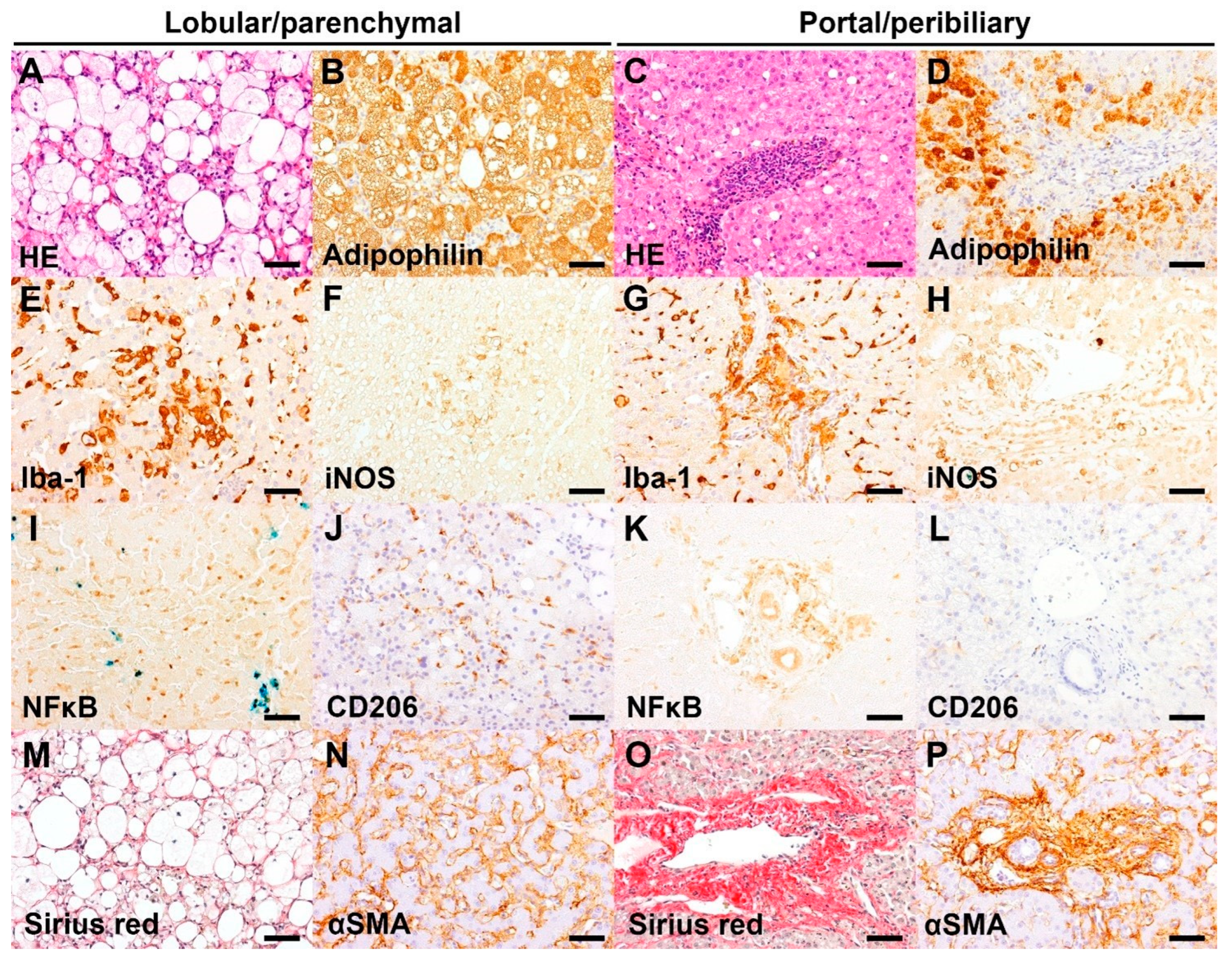
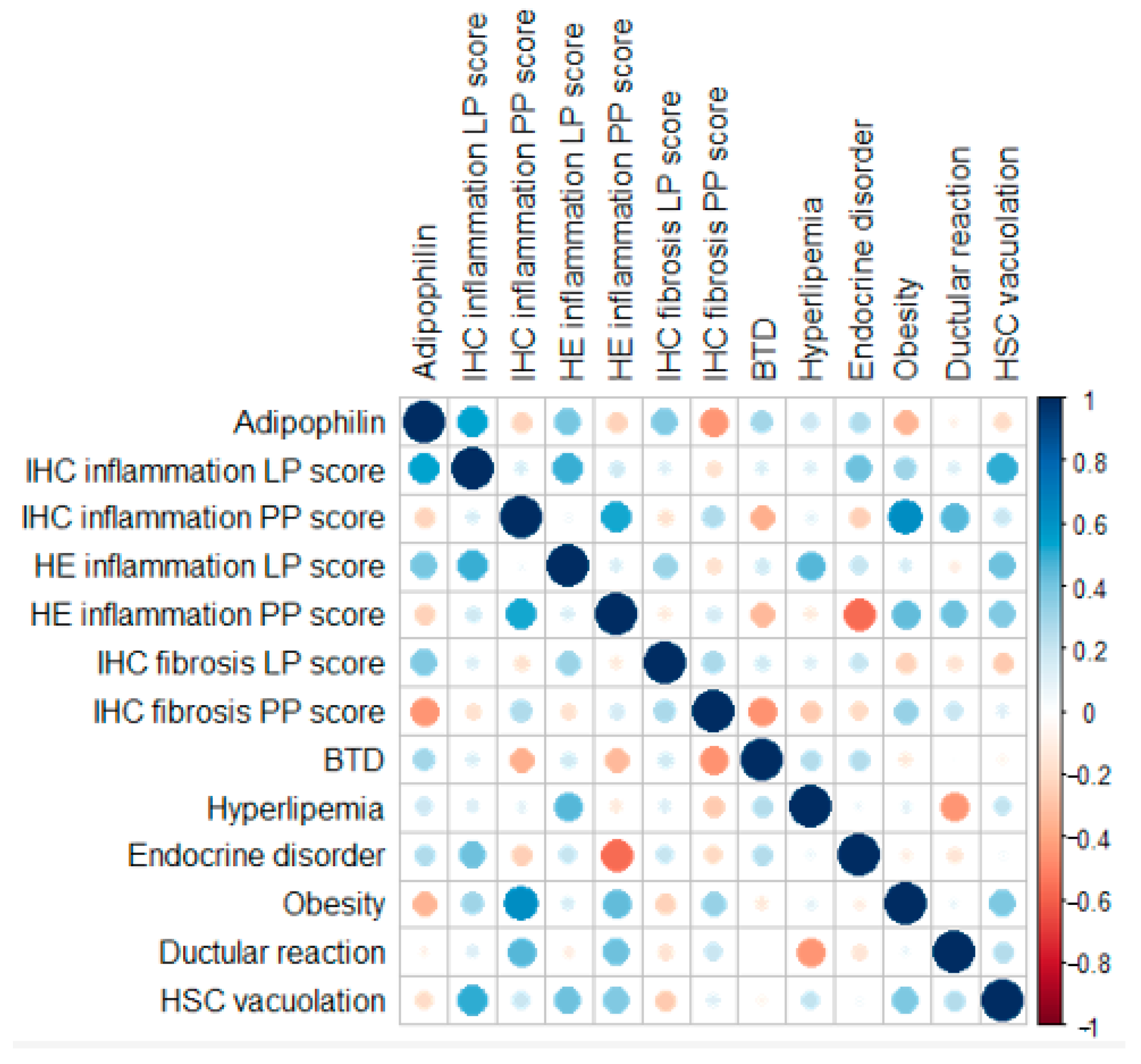
3.2. Histological and Immunohistochemical Features
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ALP | Alkaline phosphatase |
| ALT | alanine aminotransferase |
| AST | aspartate aminotransferase |
| BCS | Body Condition Score |
| BTD | Biliary Tract Disease |
| CD206 | Macrophage Mannose Receptor |
| CLD | Chronic Liver Disease |
| FFPE | Formalin-Fixed Paraffin-Embedded |
| G | General |
| GGT | gamma-glutamyl transferase |
| HE | Hematoxylin-eosin |
| HSC | Hepatic Stellate Cell |
| Iba-1 | Ionized Calcium Binding Adaptor Molecule 1 |
| IHC | Immunohystochemistry |
| iNOS | Inducible Nitric Oxide Synthase |
| LP | Lobular-parenchymal |
| LPS | lipopolysaccharides |
| MASH | Metabolic Dysfunction Associated Steatohepatitis |
| MASLD | Metabolic Dysfunction Associated Steatotic Liver Disease |
| NAFLD | Non-alcoholic Fatty Lived Disease |
| NASH | Non-alcoholic Steatohepatitis |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| PP | Portal-peribiliary |
| α-SMA | Alpha Smooth Muscle Actin |
References
- Washabau, R.J. Chapter 61: Liver in Canine and Feline Gastroenterology, 1st ed.; Washabau, R.J., Day, M.J., Eds.; Elsevier: St. Louis, MO, USA, 2013; pp. 849–939. [Google Scholar]
- Ettinger, S.J.; Feldman, E.C.; Côté, E. Section 19—Hepatobiliary disease. In Textbook of Veterinary Internal Medicine: Diseases of the Dog and the Cat, 8th ed.; Elsevier: St. Louis, MO, USA, 2017; pp. 3933–4088. [Google Scholar]
- Cullen, J.M.; van den Ingh, T.S.; Bunch, S.E.; Rothuizen, J.; Washabau, R.J.; Desmet, V.J. Morphological classification of circulatory disorders of the canine and feline liver. In WSAVA Standards for Clinical and Histological Diagnosis of Canine and Feline Liver Diseases, 1st ed.; Saunders: Edinburgh, UK; Elsevier: New York, NY, USA, 2006; pp. 41–59. [Google Scholar]
- Webster, C.R.L.; Center, S.A.; Cullen, J.M.; Penninck, D.G.; Richter, K.P.; Twedt, D.C.; Watson, P.J. ACVIM consensus statement on the diagnosis and treatment of chronic hepatitis in dogs. J. Vet. Intern. Med. 2019, 33, 1173–1200. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, S.A.; Chen, Y.X.P.; Williams, J.E.; Kendziorski, J.A.; Smedley, R.C. Concurrent hepatopathy in dogs with gallbladder mucocele: Prevalence, predictors, and impact on long-term outcome. J. Vet. Intern. Med. 2024, 38, 176–186. [Google Scholar] [CrossRef]
- Cullen, J.M.; van den Ingh, T.S.; Winkle, T.J.; Charles, J.A.; Desmet, V.J. Morphological classification of parenchymal disorders of the canine and feline liver: 1. Normal histology, reversible hepatocytic injury, and hepatic amyloidosis. In WSAVA Standards for Clinical and Histological Diagnosis of Canine and Feline Liver Diseases, 1st ed.; Saunders: Edinburgh, UK; Elsevier: New York, NY, USA, 2006; pp. 77–83. [Google Scholar]
- Kuchay, M.S.; Choudhary, N.S.; Mishra, S.K. Pathophysiological mechanisms underlying MAFLD. Diabetes Metab. Syndr. 2020, 14, 1875–1887. [Google Scholar] [CrossRef]
- Yamate, J.; Izawa, T.; Kuwamura, M. Histopathological Analysis of Rat Hepatotoxicity Based on Macrophage Functions: In Particular, an Analysis for Thioacetamide-induced Hepatic Lesions. Food Saf. 2016, 4, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Zhao, M.; Miao, C.; Shi, X.; Lu, J. Identification and validation of key biomarkers associated with macrophages in nonalcoholic fatty liver disease based on hdWGCNA and machine learning. Aging 2023, 15, 15451–15472. [Google Scholar] [CrossRef]
- Tacke, F.; Horn, P.; Wong, W.-S.; Ratziu, V.; Bugianesi, E.; Francque, S.; Zelber-Sagi, S.; Yki-Järvinen, H.; Gastaldelli, A.; Vettor, R.; et al. Clinical Practice Guidelines on the management of metabolic dysfunction-associated steatotic liver disease (MASLD). J. Hepatol. 2024, 81, 492–542. [Google Scholar] [CrossRef]
- Xenoulis, P.G.; Steiner, J.M. Canine hyperlipidaemia. J. Small Anim. Pract. 2015, 56, 595–605. [Google Scholar] [CrossRef]
- Xenoulis, P.G.; Steiner, J.M. Lipid metabolism and hyperlipidemia in dogs. Vet. J. 2010, 183, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Carloni, A.; Paninarova, M.; Cavina, D.; Romanucci, M.; Salda, L.D.; Teodori, S.; Vignoli, M. Negative hepatic computed tomographic attenuation pattern in a dog with vacuolar hepatopathy and hepatic fat accumulation secondary to cushing’s syndrome. Vet. Radiol. Ultrasound. 2019, 60, E54–E57. [Google Scholar] [CrossRef]
- Habermaass, V.; Bartoli, F.; Gori, E.; Dini, R.; Cogozzo, A.; Puccinelli, C.; Pierini, A.; Marchetti, V. Fecal Bile Acids in Canine Chronic Liver Disease: Results from 46 Dogs. Animals 2024, 14, 3051. [Google Scholar] [CrossRef]
- Krenkel, O.; Tacke, F. Liver macrophages in tissue homeostasis and disease. Nat. Rev. Immunol. 2017, 17, 306–321. [Google Scholar] [CrossRef] [PubMed]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Lee, J.H.; Li, J.; Yu, R.; Tan, L.; Xia, Y.; Zheng, Y.; Bian, X.L.; Lorenzi, P.L.; Chen, Q.; et al. Choline kinase alpha 2 acts as a protein kinase to promote lipolysis of lipid droplets. Mol. Cell 2021, 81, 2722–2735. [Google Scholar] [CrossRef] [PubMed]
- Meyerholz, D.K.; Beck, A.P. Principles and approaches for reproducible scoring of tissue stains in research. Lab. Investig. 2018, 98, 844–855. [Google Scholar] [CrossRef]
- Treuting, P.M.; Boyd, K.L. Histopathological Scoring. Vet. Pathol. 2019, 56, 17–18. [Google Scholar] [CrossRef]
- Hoogland, I.C.; Houbolt, C.; van Westerloo, D.J.; van Gool, W.A.; van de Beek, D. Systemic inflammation and microglial activation: Systematic review of animal experiments. J. Neuroinflamm. 2015, 12, 114. [Google Scholar] [CrossRef]
- Yamate, J.; Izawa, T.; Kuwamura, M. Macrophage pathology in hepatotoxicity. J. Toxicol. Pathol. 2023, 36, 51–68. [Google Scholar] [CrossRef]
- Fiaschini, N.; Mancuso, M.; Tanori, M.; Colantoni, E.; Vitali, R.; Diretto, G.; Lorenzo Rebenaque, L.; Stronati, L.; Negroni, A. Liver Steatosis and Steatohepatitis Alter Bile Acid Receptors in Brain and Induce Neuroinflammation: A Contribution of Circulating Bile Acids and Blood-Brain Barrier. Int. J. Mol. Sci. 2022, 23, 14254. [Google Scholar] [CrossRef]
- Pazdzior-Czapula, K.; Mikiewicz, M.; Gesek, M.; Zwolinski, C.; Otrocka-Domagala, I. Diagnostic immunohistochemistry for canine cutaneous round cell tumours—Retrospective analysis of 60 cases. Folia Histochem. Cytobiol. 2019, 57, 146–154. [Google Scholar] [CrossRef]
- Iwakiri, Y. Nitric oxide in liver fibrosis: The role of inducible nitric oxide synthase. Clin. Mol. Hepatol. 2015, 21, 319–325. [Google Scholar] [CrossRef]
- Mitaka, C.; Hirata, Y.; Masaki, Y.; Takei, T.; Yokoyama, K.; Imai, T. S-Methylisothiourea sulfate improves renal, but not hepatic dysfunction in canine endotoxic shock model. Intensive Care Med. 2000, 26, 117–124. [Google Scholar] [CrossRef]
- Preiser, J.C.; Zhang, H.; Vray, B.; Hrabak, A.; Vincent, J.L. Time course of inducible nitric oxide synthase activity following endotoxin administration in dogs. Nitric Oxide 2001, 5, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Zafra, R.; Jaber, J.R.; Pérez-Ecija, R.A.; Barragán, A.; Martínez-Moreno, A.; Pérez, J. High iNOS expression in macrophages in canine leishmaniasis is associated with low intracellular parasite burden. Vet. Immunol. Immunopathol. 2010, 143, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.U.; Huang, Y.; Zhu, L.; Chu, X.; Junejo, S.A.; Zhang, Y.; Khan, I.M.; Li, Y.; Feng, S.; Wu, J.; et al. Tea polyphenols attenuate liver inflammation by modulating obesity-related genes and down-regulating COX-2 and iNOS expression in high fat-fed dogs. BMC Vet. Res. 2020, 16, 234. [Google Scholar] [CrossRef]
- Vince, A.R.; Hayes, M.A.; Jefferson, B.J.; Stalker, M.J. Hepatic injury correlates with apoptosis, regeneration, and nitric oxide synthase expression in canine chronic liver disease. Vet. Pathol. 2014, 51, 932–945. [Google Scholar] [CrossRef] [PubMed]
- Papa, S.; Bubici, C.; Zazzeroni, F.; Franzoso, G. Mechanisms of liver disease: Cross-talk between the NF-kappaB and JNK pathways. Biol. Chem. 2009, 390, 965–976. [Google Scholar] [CrossRef]
- Schlein, L.J.; Thamm, D.H. Review: NF-kB activation in canine cancer. Vet. Pathol. 2022, 59, 724–732. [Google Scholar] [CrossRef]
- Huo, H.; Wu, H.; Ma, F.; Li, X.; Liao, J.; Hu, L.; Han, Q.; Li, Y.; Pan, J.; Zhang, H.; et al. N-acetyl-L-cysteine ameliorates hepatocyte pyroptosis of dog type 1 diabetes mellitus via suppression of NLRP3/NF-κB pathway. Life Sci. 2022, 306, 120802. [Google Scholar] [CrossRef]
- Wang, C.; Ma, C.; Gong, L.; Guo, Y.; Fu, K.; Zhang, Y.; Zhou, H.; Li, Y. Macrophage Polarization and Its Role in Liver Disease. Front. Immunol. 2021, 12, 803037. [Google Scholar] [CrossRef]
- Wynn, T.A.; Vannella, K.M. Macrophages in Tissue Repair, Regeneration, and Fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef]
- Belluco, S.; Sammarco, A.; Sapin, P.; Lurier, T.; Marchal, T. FOXP3, CD208, and CD206 Expression in Canine Cutaneous Histiocytoma. Vet. Pathol. 2020, 57, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Kerboeuf, M.; Haugeberg, D.A.; Olsen, T.; Sørling, L.K.; Koppang, E.O.; Moe, L.; Haaland, A.H. Tumor-associated macrophages in canine visceral hemangiosarcoma. Vet. Pathol. 2024, 61, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Ijzer, J.; Roskams, T.; Molenbeek, R.F.; Ultee, T.; Penning, L.C.; Rothuizen, J.; van den Ingh, T.S. Morphological characterisation of portal myofibroblasts and hepatic stellate cells in the normal dog liver. Comp. Hepatol. 2006, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Vince, A.R.; Hayes, M.A.; Jefferson, B.J.; Stalker, M.J. Sinusoidal endothelial cell and hepatic stellate cell phenotype correlates with stage of fibrosis in chronic liver disease in dogs. J. Vet. Diagn. Investig. 2016, 28, 498–505. [Google Scholar] [CrossRef]
- Burgos-Morón, E.; Abad-Jiménez, Z.; Marañón, A.M.; Iannantuoni, F.; Escribano-López, I.; López-Domènech, S.; Salom, C.; Jover, A.; Mora, V.; Roldan, I.; et al. Relationship Between Oxidative Stress, ER Stress, and Inflammation in Type 2 Diabetes: The Battle Continues. J. Clin. Med. 2019, 8, 1385. [Google Scholar] [CrossRef]
- Tilg, H.; Moschen, A.R. Evolution of inflammation in nonalcoholic fatty liver disease: The multiple parallel hits hypothesis. Hepatology 2010, 52, 1836–1846. [Google Scholar] [CrossRef]
- Seki, E.; Schwabe, R.F. Hepatic inflammation and fibrosis: Functional links and key pathways. Hepatology 2015, 61, 1066–1079. [Google Scholar] [CrossRef]
- Demetris, A.J.; Bellamy, C.O.C.; Gandhi, C.R.; Prost, S.; Nakanuma, Y.; Stolz, D.B. Functional Immune Anatomy of the Liver—As an Allograft. Am. J. Transplant. 2016, 16, 1653–1680. [Google Scholar] [CrossRef]
- Zhang, P.; Zheng, L.; Duan, Y.; Gao, Y.; Gao, H.; Mao, D.; Luo, Y. Gut microbiota exaggerates triclosan-induced liver injury via gut-liver axis. J. Hazard. Mater. 2022, 421, 126707. [Google Scholar] [CrossRef]
- Sato, K.; Marzioni, M.; Meng, F.; Francis, H.; Glaser, S.; Alpini, G. Ductular Reaction in Liver Diseases: Pathological Mechanisms and Translational Significances. Hepatology 2018, 69, 420–430. [Google Scholar] [CrossRef]
- Mavila, N.; Siraganahalli, E.; Kennedy, J. Ductular Reactions in Liver Injury, Regeneration, and Disease Progression—An Overview. Cells 2024, 13, 579. [Google Scholar] [CrossRef] [PubMed]
- van den Ingh, T.S.; Winkle, T.J.; Cullen, J.M.; Charles, J.A.; Desmet, V.J. Chapter 7: Morphological classification of parenchymal disorders of the canine and feline liver: 2. Hepatocellular death, hepatitis, and cirrhosis. In WSAVA Standards for Clinical and Histological Diagnosis of Canine and Feline Liver Diseases, 1st ed.; Saunders: Edinburgh, UK; Elsevier: New York, NY, USA, 2006; pp. 85–101. [Google Scholar]
- Delgado, M.E.; Cárdenas, B.I.; Farran, N.; Fernandez, M. Metabolic Reprogramming of Liver Fibrosis. Cells 2021, 10, 3604. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.; Anstee, Q.M.; Marietti, M.; Hardy, T.; Henry, L.; Eslam, M.; George, J.; Bugianesi, E. Global burden of NAFLD and NASH: Trends, predictions, risk factors and prevention. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Fukusato, T. Histopathology of nonalcoholic fatty liver disease/nonalcoholic steatohepatitis. World J. Gastroenterol. 2014, 20, 15539–15548. [Google Scholar] [CrossRef]
- Park, C.S.; Shastri, N. The Role of T Cells in Obesity-Associated Inflammation and Metabolic Disease. Immune Netw. 2022, 22, e13. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Significance | Host/Clone | Dilution | Pretreatment | CatalogID | Source |
|---|---|---|---|---|---|---|
| CD206 | Alternatively activated (M2) or reparative macrophage (collagen deposition) | Rabbit/polyclonal | 1:1000 | AC/Citrate | Ab64693 | Abcam, Cambridge, MA, USA |
| iNOS | Classically activated (M1) or proinflammatory macrophage | Rabbit/polyclonal | 1:500 | AC/Citrate | AHP303 | Bio-Rad, Hercules, CA, USA |
| αSMA | Activated myofibroblast/ activated HSC | Mouse/1A4 | 1:500 | No pretreatment | M0851 | Dako, Carpinteria, CA, USA |
| Iba-1 | Kupffer cell/macrophage (pan-macrophage marker) | Rabbit/polyclonal | 1:1000 | AC/Citrate | 019-19741 | FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan |
| NF-κB (p65/RelA) | Transcription factor for pro-inflammatory factors (cytokines, chemokines) | Rabbit /1D8 | 1:500 | AC/Citrate | ZRB1498 | Sigma-Aldrich, St Louis, MO, USA |
| Adipophilin (perilipin 2) | Structural component of lipid droplets, localized in their outer surface | Mouse/AP125 | 1:200 | AC/Citrate | 690102S | Progen, Heidelberg, Germany |
| N | Breed | Sex | Age | Endocrinopathy | BCS | Cholesterol (150–280 mg/dL) | Triglicerids (25–90 mg/dL) | Hyperlipemia | BTD |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Border Collie | F | 7 | diabetes mellitus | 3 | 297.5 | 370 | yes | yes |
| 2 | Dachshund | M | 8.1 | diabetes mellitus | 4 | 459.1 | 65 | yes | yes |
| 3 | Golden Retriever | F | 1.8 | / | 5 | 127.2 | 63 | no | no |
| 4 | WHWT | F | 10 | diabetes mellitus | 5 | 248.4 | 48 | no | no |
| 5 | Mix-breed | FN | 10 | / | 5 | 270.6 | 143 | yes | no |
| 6 | Mix-Breed | M | 2.7 | / | 5 | 307.1 | 56 | yes | yes |
| 7 | Zwergpinscher | M | 10.4 | / | 6 | 250.5 | 59 | no | no |
| 8 | Mix-Breed | FS | 9.9 | hypothyroidism | 6 | 266.7 | 69 | no | no |
| 9 | Mix-breed | FN | 9.9 | / | 6 | 353.5 | 58 | yes | no |
| 10 | Toy Poodle | M | 6.4 | / | 6 | 250 | 172 | yes | no |
| 11 | Mix-breed | F | 9.7 | / | 6 | 167.9 | 83 | no | yes |
| 12 | Mix-breed | FN | 11.4 | / | 6 | 345.2 | 77 | yes | yes |
| 13 | Mix-breed | FN | 7.02 | / | 7 | 338 | 187 | yes | no |
| 14 | Mix-Breed | M | 9.3 | / | 7 | 808.8 | 717 | yes | yes |
| 15 | Beagle | F | 8.5 | / | 4 | 251 | 80 | no | yes |
| 16 | Toy Poodle | F | 14.8 | hyperadrenocorticism | 6 | 354 | 66 | yes | yes |
| N | US Hepatobiliary Findings | Histological Features |
|---|---|---|
| 1 | Moderate diffuse liver disease with increased liver size and diffuse increase in echogenicity. Mild cholecystopathy with slightly thickened and hyperechoic walls. Common bile duct moderately dilated along its entire course (4 mm), filled with corpuscular echogenic contents, and with slightly thickened walls. Gallbladder with biliary sludge slightly corpuscular contents in suspension. | Degenerative hepatopathy (steatosis), multifocal steatohepatitis |
| 2 | Mild/moderate diffuse liver disease with slightly increased in size, increased echogenicity. Mild/moderate cholecystopathy with slightly enlarged walls thick and hyperechoic, irregular mucosal profile with the presence of a cystic mucosal lesion. | Degenerative hepatopathy (glicogenosis) |
| 3 | Moderate/severe diffuse liver disease with slightly reduced in size with irregular shape and slightly lumpy profile, heterogeneous echogenicity and altered echo structure due to the presence of multiple hyperechoic bands and striae delimiting multiple hypoechoic nodular lesions of variable dimensions deforming the capsule. Peritoneal effusion. Aberrant venous vessels compatible with shunts acquired. | Lobular dissecting hepatitis |
| 4 | Moderate diffuse liver disease with increased in size with rounded margins, diffusely increased echogenicity. Mild cholecystopathy. Gallbladder with biliary sludge and aggregated hyperechoic content suspended. | Chronic hepatitis, degenerative hepatopathy (glycogenosis) |
| 5 | Moderate diffuse liver disease with moderately increased in size, with rounded margins, moderately increased echogenicity in diffuse manner and slightly heterogeneous echo structure with the presence of multiple nodular lesions. Mineralizations of the bile/parenchymal ducts. Mild diffuse cholecystopathy. Hyperechoic structure in the lumen of the gallbladder (cholelitis, aggregated bile sludge, mucosal proliferation). | Cholangiohepatitis, intrahepatic cholestasis |
| 6 | Moderate diffuse liver disease with moderately enlarged and hyperechoic, finely dissimilar echo structure. Severe diffuse cholecystopathy with gallbladder mucocele stage VI, suspected parietal rupture. | Colangitis, portal inflammation, degenerative hepatopathy (glycogenosis), pigmentary granulomas |
| 7 | Moderate diffuse liver disease with liver slightly increased in size, slightly increased echogenicity and finely inhomogeneous echo structure with multiple lesions. Gallbladder with slight hyperechoic content in suspension | Degenerative hepatopathy (glycogenosis), portal inflammation, pigmentary granulomas |
| 8 | Moderate diffuse liver disease with liver moderately enlarged in size, with increased parenchymal echogenicity and inhomogeneous echo structure (stiae, multiple lesions). Gallbladder with biliary sludge, deposited. | Chronic hepatitis |
| 9 | Moderate diffuse liver disease, with liver with generally hyperechoic parenchyma. Mild interlobar hepatic effusion. | Portal inflammation, Intrahepatic cholestasis, degenerative hepatopathy (glycogenosis) |
| 10 | Moderate cholecistopathy, with gallbladder with abundant hyperechoic corpuscular content, partially organized, thickened and hypoechoic walls. | Chronic hepatitis, degenerative hepatopathy (glycogenosis), pigmentary granulomas |
| 11 | Mild diffuse liver disease with moderately increased in size with slight increase in echogenicity. Severe diffuse cholecystopathy with gallbladder mucocele stage VI, suspected parietal rupture. Moderate/severe multifocal peritoneal reactivity. | Cholangiohepatitis, portal inflammation, intrahepatic cholestasis |
| 12 | Moderate cholecystopathy with intraluminal cholelitis and suboccluding/occluding the common bile duct. | Colangitis with concentric periductural fibrosis, intra and extra-hepatic cholestasis, pigmentary granulomas |
| 13 | Mild diffuse liver disease, slightly increased in size. | Portal inflammation, intrahepatic cholestasis |
| 14 | Moderate diffuse liver disease, with liver slightly increased in size and diffusely hyperechoic. Severe diffuse cholecystopathy with gallbladder mucocele stage VI, suspected parietal rupture. | Chronic hepatitis, concentric periductular fibrosis, degenerative hepatopathy (steatosis) |
| 15 | Moderate diffuse liver disease, with liver with slightly hypoechoic parenchyma alteration of the echo structure for nodular lesions. Gallbladder with gravity-dependent biliary sludge and gallstones. | Portal inflammation, hepatocellular cholestasis, pigmentary granulomas |
| 16 | Moderate cholecystopathy with thickened walls and hyperechoic, presence of mild amount of peripheral anechoic fluid. Slightly dilated common bile duct and presence of a small layer of liquid near the duodenal papilla. | Cholangiohepatitis, intrahepatic cholestasis, pigmentary granulomas |
| N. | Inflammation HE PL (0–3) | Inflammation HE PP (0–3) | SUM Inflammation HE (0–6) | Ductular Reaction (A/P) | HSC Vacuolation (0–3) |
|---|---|---|---|---|---|
| 1 | 3 | 0 | 3 | A | 0 |
| 2 | 0 | 0 | 0 | A | 0 |
| 3 | 0 | 2 | 2 | P | 0 |
| 4 | 1 | 0 | 1 | P | 0 |
| 5 | 1 | 1 | 2 | P | 0 |
| 6 | 2 | 2 | 4 | P | 0 |
| 7 | 1 | 2 | 3 | P | 1 |
| 8 | 1 | 2 | 3 | P | 0 |
| 9 | 1 | 1 | 2 | A | 0 |
| 10 | 2 | 3 | 5 | P | 3 |
| 11 | 1 | 2 | 3 | P | 0 |
| 12 | 1 | 2 | 3 | P | 0 |
| 13 | 1 | 1 | 2 | A | 0 |
| 14 | 1 | 0 | 1 | P | 0 |
| 15 | 0 | 0 | 0 | P | 0 |
| 16 | 2 | 1 | 3 | P | 3 |
| N. | Iba-1 (0–3) | iNOS (0–3) | NFκB (0–3) | αSMA (0–3) | CD206 (0–3) | SiriusRed (0–3) | Adipophilin (0–3) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LP | PP | G | LP | PP | G | LP | PP | G | LP | PP | G | LP | PP | G | LP | PP | G | G | |
| 1 | 3 | 0 | 3 | 1 | 0 | 1 | 2 | 0 | 2 | 2 | 2 | 4 | 2 | 0 | 2 | 1 | 2 | 3 | 2 |
| 2 | 2 | 0 | 2 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 1 | 2 | 0 | 0 | 0 | 0 | 1 | 1 | 1 |
| 3 | 2 | 2 | 4 | 2 | 1 | 3 | 2 | 0 | 2 | 2 | 3 | 5 | 1 | 0 | 1 | 0 | 2 | 2 | 1 |
| 4 | 1 | 1 | 2 | 1 | 1 | 2 | 2 | 1 | 3 | 0 | 2 | 2 | 1 | 0 | 1 | 0 | 2 | 1 | 1 |
| 5 | 1 | 1 | 2 | 1 | 1 | 2 | 1 | 2 | 3 | 1 | 3 | 4 | 0 | 0 | 0 | 1 | 2 | 4 | 1 |
| 6 | 2 | 2 | 4 | 2 | 1 | 3 | 2 | 1 | 3 | 1 | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| 7 | 2 | 2 | 4 | 2 | 2 | 4 | 1 | 1 | 2 | 0 | 2 | 2 | 1 | 0 | 1 | 0 | 2 | 2 | 1 |
| 8 | 3 | 2 | 5 | 1 | 1 | 2 | 2 | 2 | 4 | 1 | 3 | 5 | 0 | 0 | 0 | 1 | 2 | 3 | 1 |
| 9 | 2 | 1 | 3 | 1 | 1 | 2 | 2 | 2 | 4 | 1 | 3 | 4 | 1 | 0 | 1 | 0 | 2 | 2 | 1 |
| 10 | 2 | 2 | 4 | 2 | 1 | 3 | 2 | 1 | 3 | 1 | 3 | 4 | 0 | 0 | 0 | 0 | 2 | 2 | 1 |
| 11 | 2 | 1 | 3 | 1 | 0 | 1 | 2 | 1 | 3 | 0 | 2 | 2 | 1 | 0 | 1 | 0 | 2 | 2 | 0 |
| 12 | 1 | 3 | 4 | 1 | 1 | 2 | 1 | 2 | 3 | 2 | 3 | 5 | 0 | 0 | 0 | 1 | 2 | 3 | 3 |
| 13 | 1 | 1 | 1 | 1 | 1 | 2 | 2 | 2 | 4 | 0 | 2 | 2 | 0 | 0 | 0 | 0 | 2 | 2 | 3 |
| 14 | 3 | 2 | 5 | 2 | 2 | 4 | 2 | 1 | 2 | 1 | 2 | 3 | 0 | 0 | 0 | 1 | 2 | 3 | 1 |
| 15 | 1 | 1 | 2 | 1 | 0 | 1 | 1 | 0 | 1 | 1 | 3 | 4 | 0 | 0 | 0 | 0 | 2 | 1 | 3 |
| 16 | 3 | 1 | 4 | 2 | 2 | 4 | 3 | 1 | 4 | 1 | 2 | 3 | 0 | 0 | 0 | 0 | 2 | 1 | 1 |
| N. | SUM IHC Inflammation LP (0–9) | SUM IHC Inflammation PP (0–9) | SUM IHC Inflammation G (0–9) | SUM IHC Fibrosis LP (0–9) | SUM IHC Fibrosis PP (0–9) | SUM IHC Fibrosis G (0–9) |
|---|---|---|---|---|---|---|
| 1 | 6 | 0 | 6 | 5 | 4 | 9 |
| 2 | 4 | 0 | 4 | 1 | 2 | 3 |
| 3 | 6 | 3 | 9 | 3 | 5 | 8 |
| 4 | 4 | 3 | 7 | 1 | 4 | 5 |
| 5 | 3 | 4 | 7 | 2 | 5 | 7 |
| 6 | 6 | 4 | 10 | 1 | 1 | 2 |
| 7 | 5 | 5 | 10 | 1 | 4 | 5 |
| 8 | 6 | 5 | 11 | 2 | 5 | 7 |
| 9 | 5 | 4 | 9 | 2 | 5 | 7 |
| 10 | 6 | 4 | 10 | 1 | 5 | 6 |
| 11 | 5 | 2 | 7 | 1 | 4 | 5 |
| 12 | 3 | 6 | 9 | 3 | 5 | 8 |
| 13 | 4 | 4 | 8 | 0 | 4 | 4 |
| 14 | 7 | 5 | 12 | 2 | 4 | 6 |
| 15 | 3 | 1 | 4 | 1 | 5 | 6 |
| 16 | 8 | 4 | 12 | 1 | 4 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habermaass, V.; Takami, Y.; Izawa, T.; Abramo, F.; Biolatti, C.; Marchetti, V. Lipid Dysmetabolism in Canine Chronic Liver Disease: Relationship Between Clinical, Histological and Immunohistochemical Features. Vet. Sci. 2025, 12, 220. https://doi.org/10.3390/vetsci12030220
Habermaass V, Takami Y, Izawa T, Abramo F, Biolatti C, Marchetti V. Lipid Dysmetabolism in Canine Chronic Liver Disease: Relationship Between Clinical, Histological and Immunohistochemical Features. Veterinary Sciences. 2025; 12(3):220. https://doi.org/10.3390/vetsci12030220
Chicago/Turabian StyleHabermaass, Verena, Yuki Takami, Takeshi Izawa, Francesca Abramo, Corrado Biolatti, and Veronica Marchetti. 2025. "Lipid Dysmetabolism in Canine Chronic Liver Disease: Relationship Between Clinical, Histological and Immunohistochemical Features" Veterinary Sciences 12, no. 3: 220. https://doi.org/10.3390/vetsci12030220
APA StyleHabermaass, V., Takami, Y., Izawa, T., Abramo, F., Biolatti, C., & Marchetti, V. (2025). Lipid Dysmetabolism in Canine Chronic Liver Disease: Relationship Between Clinical, Histological and Immunohistochemical Features. Veterinary Sciences, 12(3), 220. https://doi.org/10.3390/vetsci12030220