

Effects of Designed Herbal Formula on Growth Performance, Blood Indices, Organ Traits, and Cecum Microbiology in Broilers


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Herbal Formula Preparation
2.2. Animals, Diets, and Experimental Design
2.3. Growth Performance
2.4. Organ Traits
2.5. Routine Blood Test
2.6. Microbial Diversity of the Cecum
2.7. Statistical Analysis
3. Results
3.1. Pre-Experimental Growth Performance
3.2. Growth Performance
3.3. Organ Traits
3.4. Routine Blood Test
3.5. Microbial Diversity of the Cecum
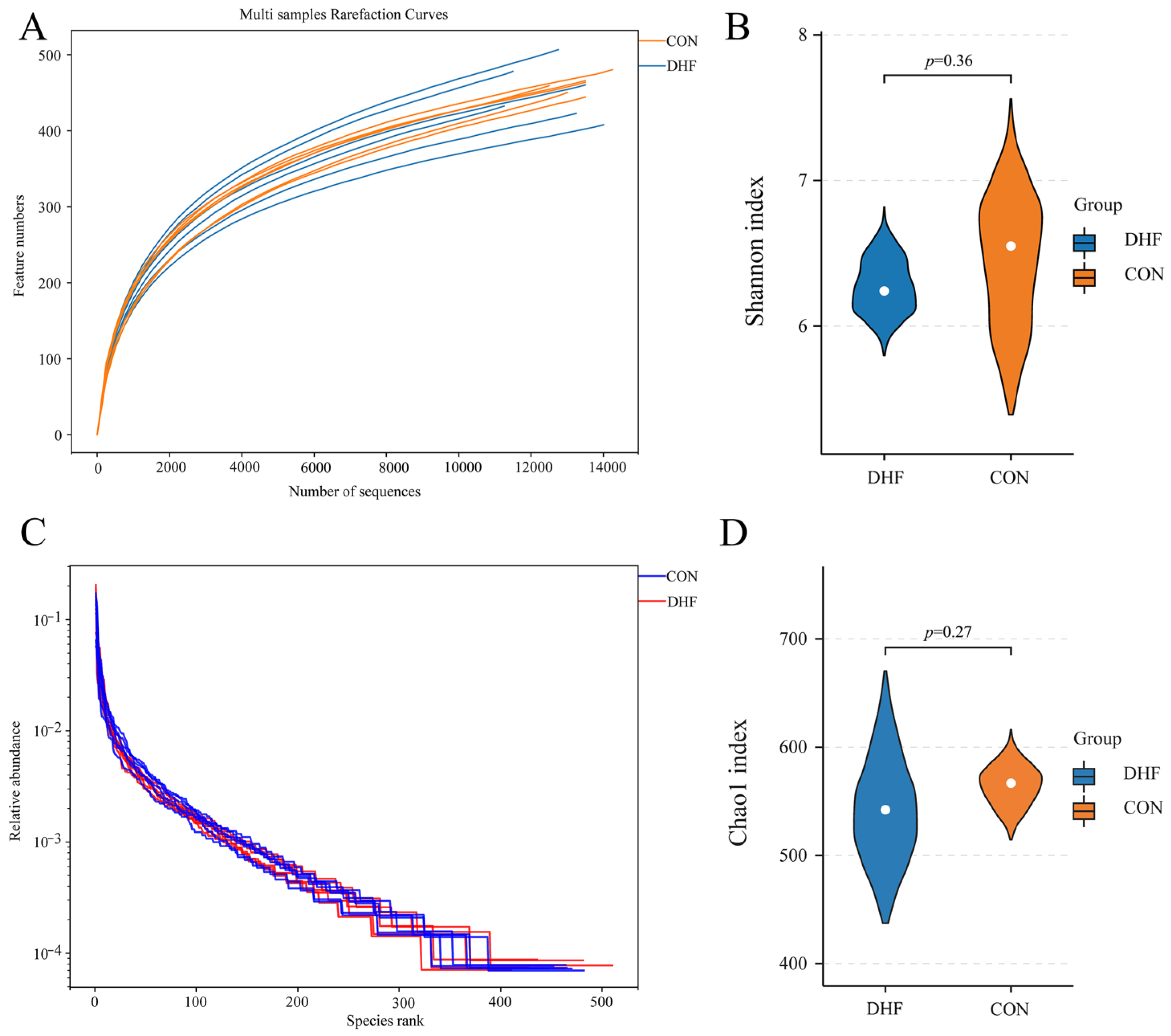
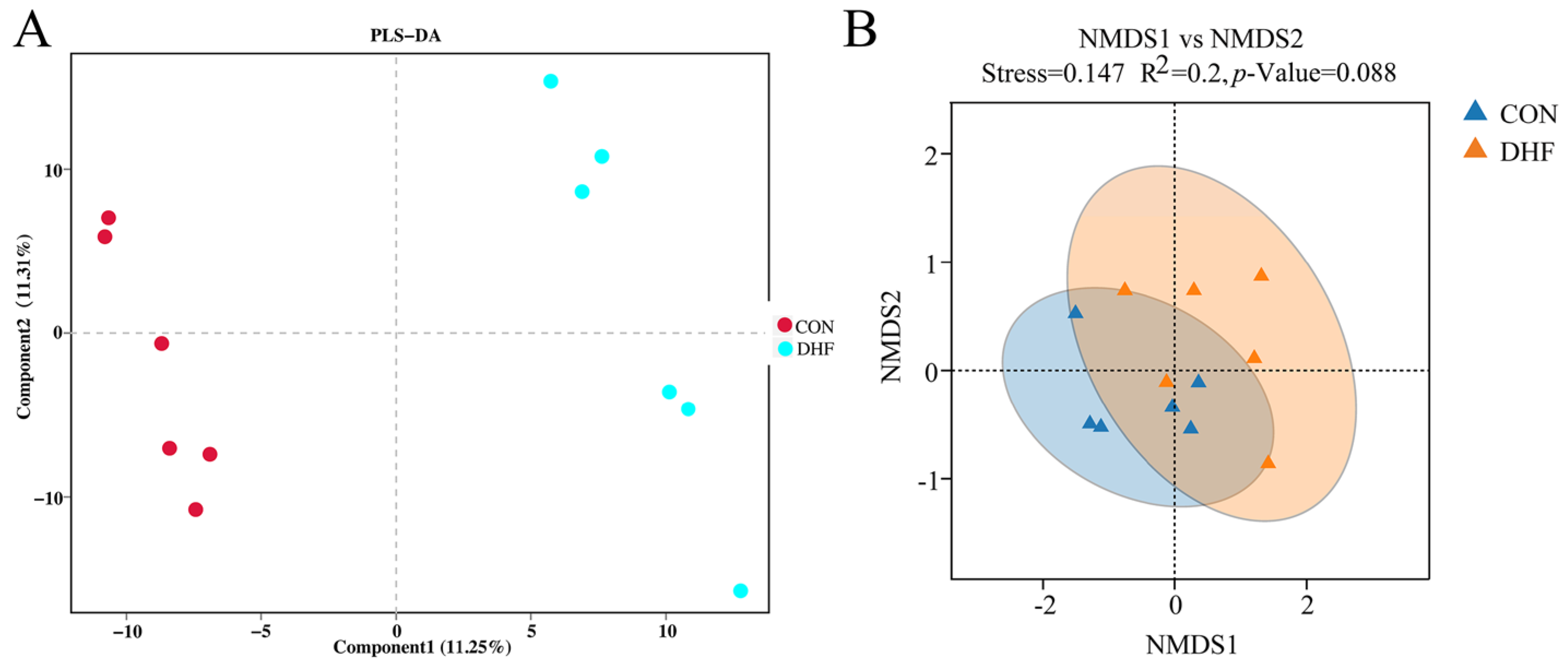
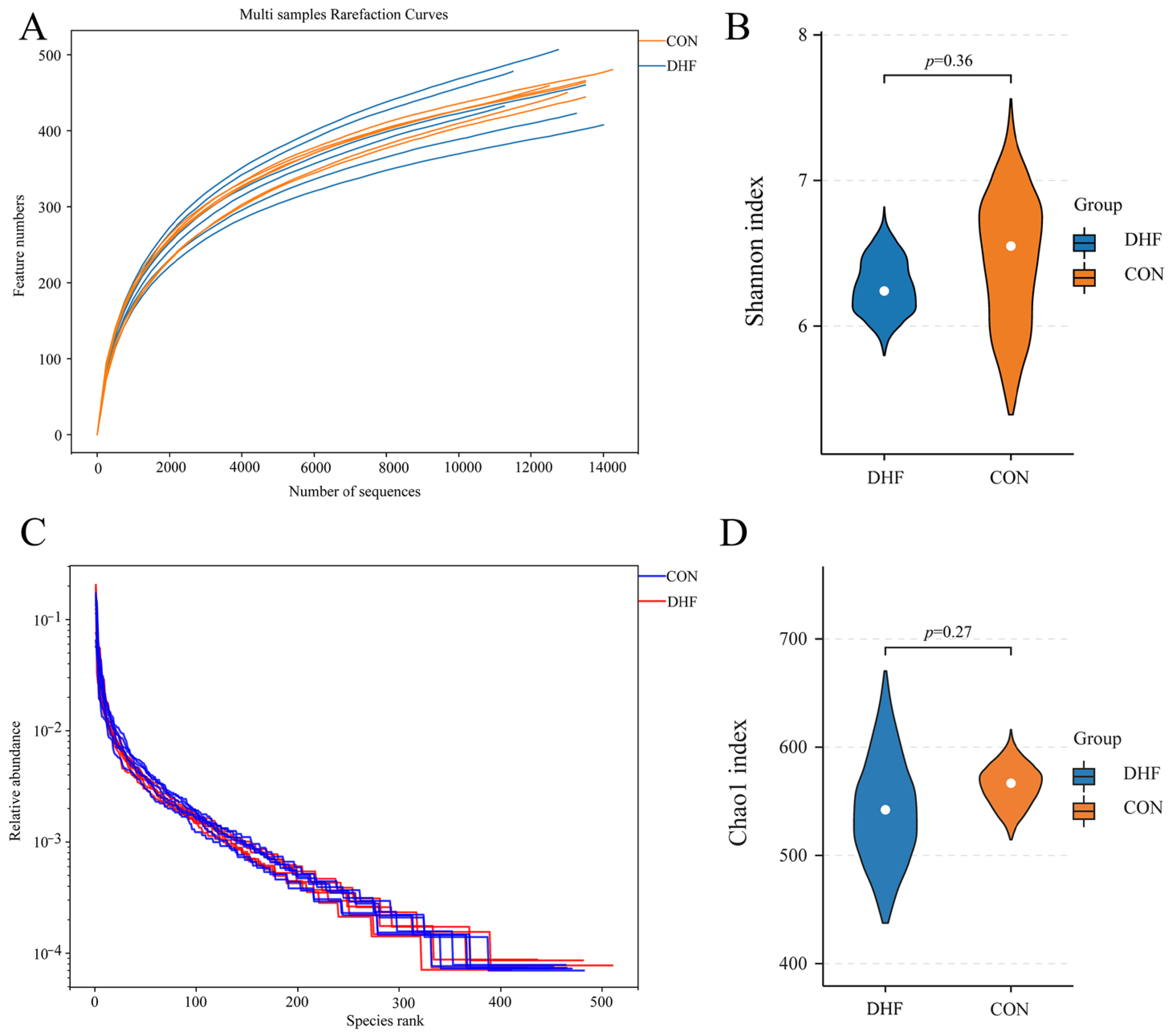
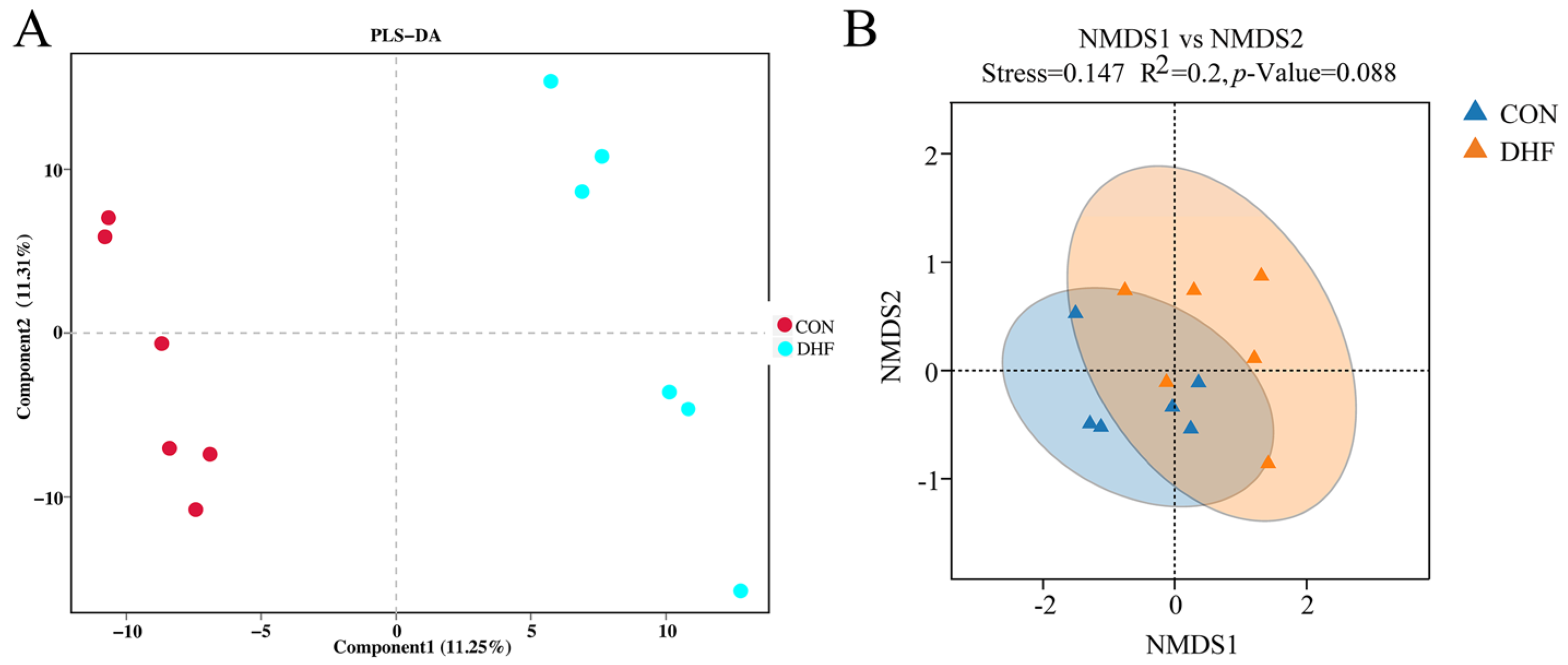
3.5.1. Alpha and Beta Diversity Analysis
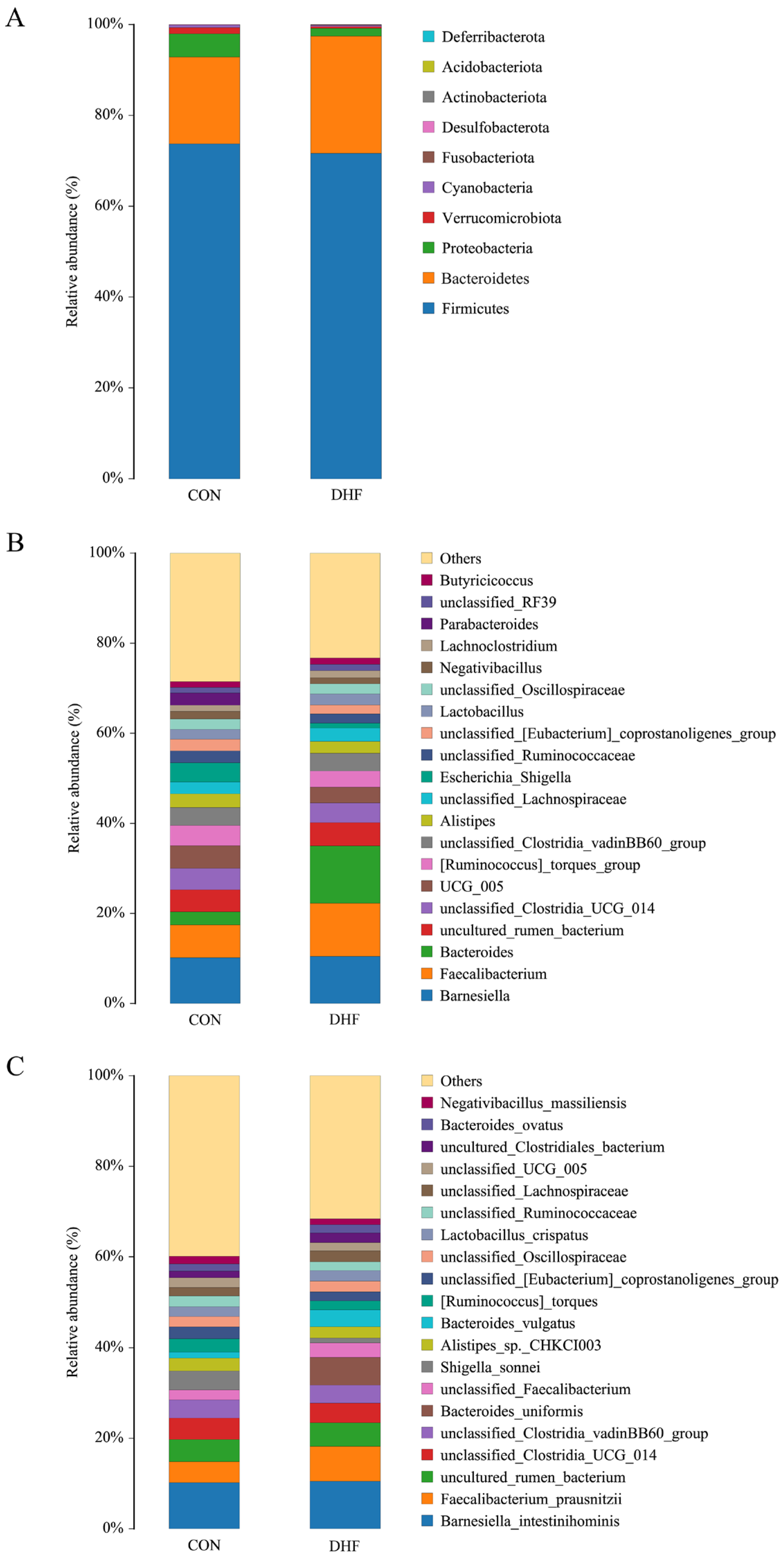
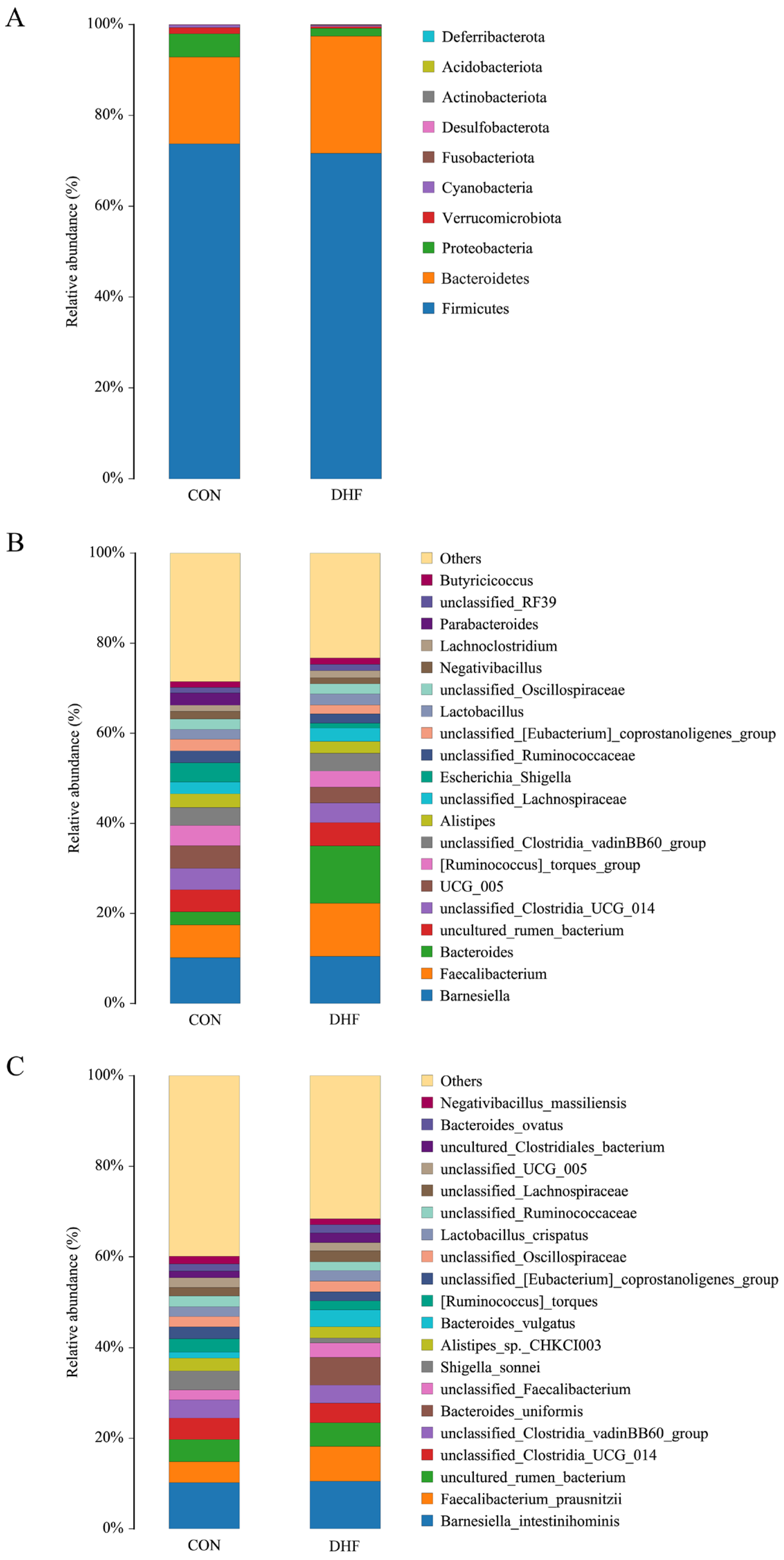
3.5.2. Analysis of Community Composition
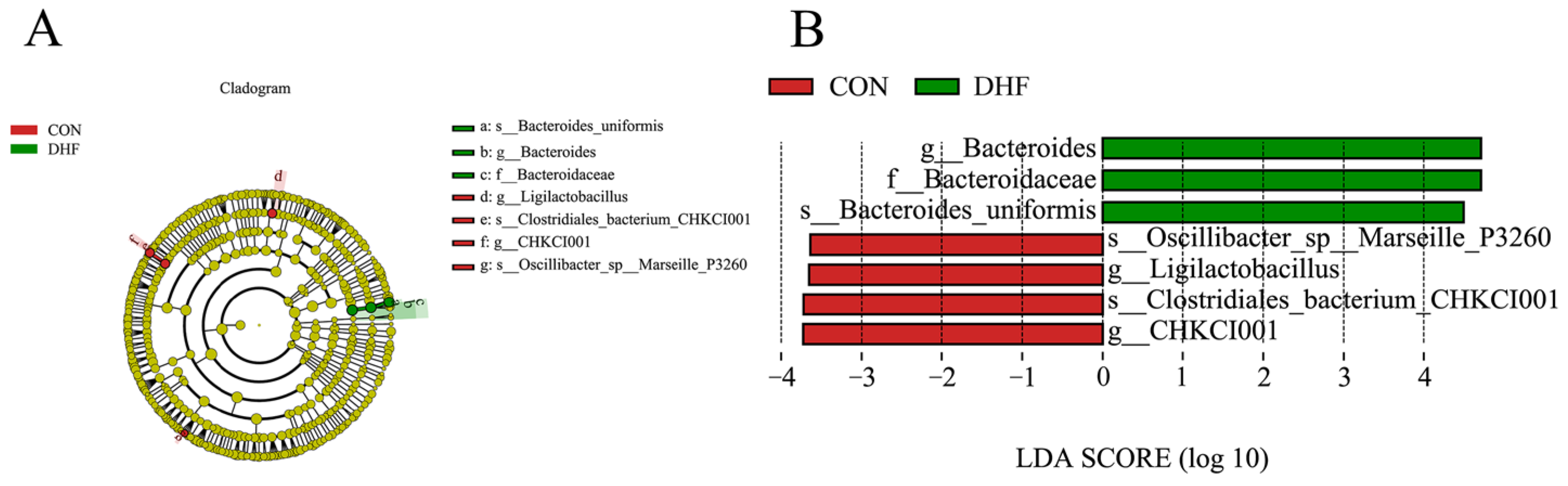
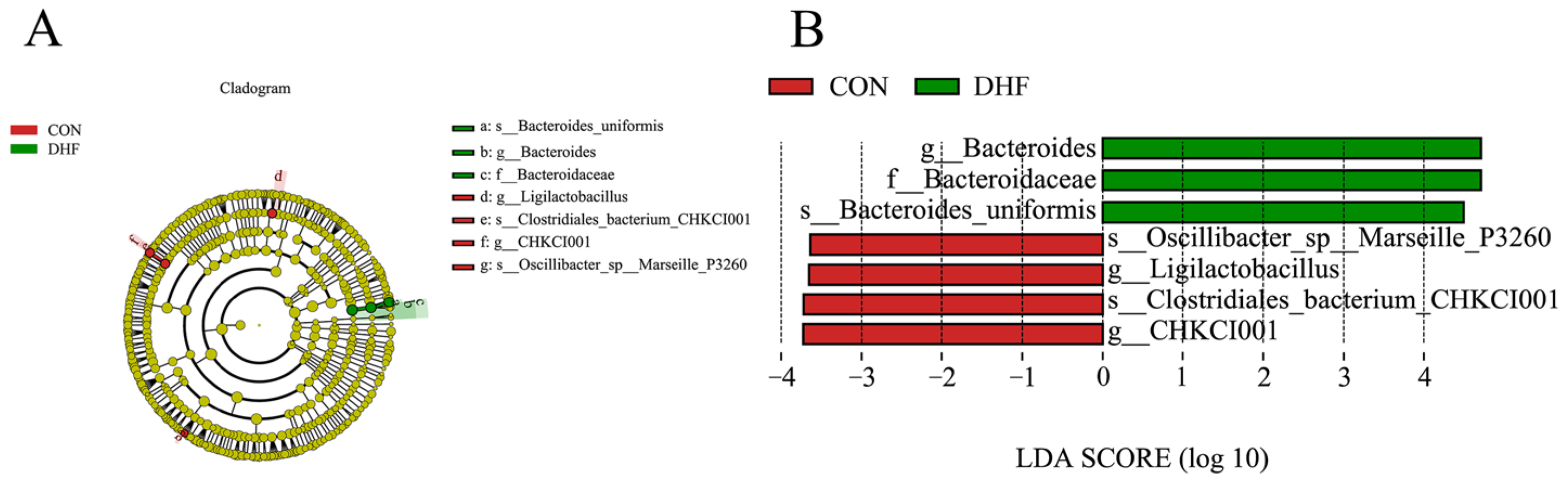
3.5.3. Analysis of Significance of Differences between Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ayalew, H.; Zhang, H.; Wang, J.; Wu, S.; Qiu, K.; Qi, G.; Tekeste, A.; Wassie, T.; Chanie, D. Potential feed additives as antibiotic alternatives in broiler production. Front. Vet. Sci. 2022, 9, 916473. [Google Scholar] [CrossRef]
- Zhu, Q.; Sun, P.; Zhang, B.; Kong, L.; Xiao, C.; Song, Z. Progress on gut health maintenance and antibiotic alternatives in broiler chicken production. Front. Nutr. 2021, 8, 692839. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, M.K. Use of antibiotics as feed additives: A burning question. Front. Microbiol. 2014, 5, 334. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, Y.; Wu, J.; Li, X.; Yu, L.; Xie, K.; Zhang, M.; Ren, L.; Ji, Y.; Li, Y. Exposure to veterinary antibiotics via food chain disrupts gut microbiota and drives increased Escherichia coli virulence and drug resistance in young adults. Pathogens 2022, 11, 1062. [Google Scholar] [CrossRef] [PubMed]
- Puvača, N.; Ljubojević Pelić, D.; Pelić, M.; Bursić, V.; Tufarelli, V.; Piemontese, L.; Vuković, G. Microbial resistance to antibiotics and biofilm formation of bacterial isolates from different carp species and risk assessment for public health. Antibiotics 2023, 12, 143. [Google Scholar] [CrossRef]
- Tan, M.F.; Li, H.Q.; Yang, Q.; Zhang, F.F.; Tan, J.; Zeng, Y.B.; Wei, Q.P.; Huang, J.N.; Wu, C.C.; Li, N.; et al. Prevalence and antimicrobial resistance profile of bacterial pathogens isolated from poultry in Jiangxi Province, China from 2020 to 2022. Poult. Sci. 2023, 102, 102830. [Google Scholar] [CrossRef] [PubMed]
- Mohammadagheri, N.; Najafi, R.; Najafi, G. Effects of dietary supplementation of organic acids and phytase on performance and intestinal histomorphology of broilers. Vet. Res. Forum. 2016, 7, 189–195. [Google Scholar]
- Giannenas, I.; Papadopoulos, E.; Tsalie, E.; Triantafillou, E.; Henikl, S.; Teichmann, K.; Tontis, D. Assessment of dietary supplementation with probiotics on performance, intestinal morphology and microflora of chickens infected with Eimeria tenella. Vet. Parasitol. 2012, 188, 31–40. [Google Scholar] [CrossRef]
- Reda, F.M.; El-Saadony, M.T.; El-Rayes, T.K.; Farahat, M.; Attia, G.; Alagawany, M. Dietary effect of licorice (Glycyrrhiza glabra) on quail performance, carcass, blood metabolites and intestinal microbiota. Poult. Sci. 2021, 100, 101266. [Google Scholar] [CrossRef]
- Kiarie, E.G.; Leung, H.; Akbari Moghaddam Kakhki, R.; Patterson, R.; Barta, J.R. Utility of feed enzymes and yeast derivatives in ameliorating deleterious effects of coccidiosis on intestinal health and function in broiler chickens. Front. Vet. Sci. 2019, 6, 473. [Google Scholar] [CrossRef]
- Mahmoud, M.M.; Al-Hejin, A.M.; Abujamel, T.S.; Ghetas, A.M.; Yacoub, H.A. Chicken β-defensin-1 peptide as a candidate anticoccidial agent in broiler chickens. Anim. Biotechnol. 2023, 34, 3108–3125. [Google Scholar] [CrossRef]
- Ogbuewu, I.P.; Okoro, V.M.; Mbajiorgu, C.A. Meta-analysis of the influence of phytobiotic (pepper) supplementation in broiler chicken performance. Trop. Anim. Health Prod. 2020, 52, 17–30. [Google Scholar] [CrossRef]
- Chen, Z.; Liu, L.; Gao, C.; Chen, W.; Vong, C.T.; Yao, P.; Yang, Y.; Li, X.; Tang, X.; Wang, S.; et al. Astragali radix (Huangqi): A promising edible immunomodulatory herbal medicine. J. Ethnopharmacol. 2020, 258, 112895. [Google Scholar] [CrossRef]
- Zhao, X.; Sun, W.; Zhang, S.; Meng, G.; Qi, C.; Fan, W.; Wang, Y.; Liu, J. The immune adjuvant response of polysaccharides from Atractylodis macrocephalae Koidz in chickens vaccinated against Newcastle disease (ND). Carbohydr. Polym. 2016, 141, 190–196. [Google Scholar] [CrossRef]
- Wang, H.; Zheng, X.; Lin, Y.; Zheng, X.; Yan, M.; Li, Y.; Shi, D.; Guo, S.; Liu, C. The mixture of Radix isatidis, Forsythiae, and Gypsum alleviates lipopolysaccharide-induced fever in broilers by inhibition of TLR4/NF-κB signaling pathway. Poult. Sci. 2023, 102, 103032. [Google Scholar] [CrossRef]
- Jiang, X.-R.; Zhang, H.-J.; Wang, J.; Wu, S.-G.; Yue, H.-Y.; Lü, H.-Y.; Cui, H.; Bontempo, V.; Qi, G.-H. Effect of dried Tangerine peel extract supplementation on the growth performance and antioxidant status of broiler chicks. Ital. J. Anim. Sci. 2016, 15, 642–648. [Google Scholar] [CrossRef]
- Xiao, Y.Q.; Shao, D.; Sheng, Z.W.; Wang, Q.; Shi, S.R. A mixture of daidzein and Chinese herbs increases egg production and eggshell strength as well as blood plasma Ca, P, antioxidative enzymes, and luteinizing hormone levels in post-peak, brown laying hens. Poult. Sci. 2019, 98, 3298–3303. [Google Scholar] [CrossRef]
- Li, X.L.; He, W.L.; Wang, Z.B.; Xu, T.S. Effects of Chinese herbal mixture on performance, egg quality and blood biochemical parameters of laying hens. J. Anim. Physiol. Anim. Nutr. 2016, 100, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Zhou, Y.; Song, Y.; Liu, C.; Hu, M.; Xie, Q.; Wang, J.; Zhang, Y.; Shi, Y.; Chen, S.; et al. The effect of combined dietary supplementation of herbal additives on carcass traits, meat quality, immunity and cecal microbiota composition in Hungarian white geese. PeerJ 2023, 11, e15316. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, T.; Cui, X.; Li, S.; Zhao, X.; Zhong, X. Hepatoprotective effects of a chinese herbal formula, longyin decoction, on carbon-tetrachloride-induced liver injury in chickens. Evid. Based Complement. Alternat. Med. 2013, 2013, 392743. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Geng, Z.; Li, Y.; Song, X.; Li, L.; Wen, A.; Yin, Z. Effects of “Shi Ying Zi” powder and osthole on immune and antioxidant function of Eimeria tenella-infected broilers. Exp. Parasitol. 2023, 246, 108451. [Google Scholar] [CrossRef] [PubMed]
- Dale, N. National Research Council Nutrient Requirements of Poultry-Ninth Revised Edition (1994). J. Appl. Poult. Res. 1994, 3, 101. [Google Scholar] [CrossRef]
- Yang, L.; Chen, L.; Zheng, K.; Ma, Y.J.; He, R.X.; Arowolo, M.A.; Zhou, Y.J.; Xiao, D.F.; He, J.H. Effects of fenugreek seed extracts on growth performance and intestinal health of broilers. Poult. Sci. 2022, 101, 101939. [Google Scholar] [CrossRef]
- GB/T 194728-2018; Operating Procedure of Livestock and Poultry Slaughtering—Chicken. State Administration for Market Regulation; Standardization Administration of the People’s Republic of China; Standards Press of China: Beijing, China, 2019.
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Ka, L.C. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Cheng, G.; Hao, H.; Xie, S.; Wang, X.; Dai, M.; Huang, L.; Yuan, Z. Antibiotic alternatives: The substitution of antibiotics in animal husbandry? Front. Microbiol. 2014, 5, 217. [Google Scholar] [CrossRef]
- Lee, M.; Shin, H.; Park, M.; Kim, A.; Cha, S.; Lee, H. Systems pharmacology approaches in herbal medicine research: A brief review. BMB Rep. 2022, 55, 417–428. [Google Scholar] [CrossRef]
- Liang, X.; Yamazaki, K.; Kamruzzaman, M.; Bi, X.; Panthee, A.; Sano, H. Effects of Chinese herbal medicine on plasma glucose, protein and energy metabolism in sheep. J. Anim. Sci. Biotechnol. 2013, 4, 51. [Google Scholar] [CrossRef]
- Lipiński, K.; Antoszkiewicz, Z.; Kotlarczyk, S.; Mazur-Kuśnirek, M.; Kaliniewicz, J.; Makowski, Z. The effect of herbal feed additive on the growth performance, carcass characteristics and meat quality of broiler chickens fed low-energy diets. Arch. Anim. Breed. 2019, 62, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Oloruntola, O.D.; Agbede, J.O.; Ayodele, S.O.; Oloruntola, D.A. Neem, pawpaw and bamboo leaf meal dietary supplementation in broiler chickens: Effect on performance and health status. J. Food Biochem. 2019, 43, e12723. [Google Scholar] [CrossRef] [PubMed]
- Behboodi, H.R.; Hosseini, D.; Salarieh, A.; Gholampour, M.; Panahi, M.; Alemi, M.; Baradaran, A.; Nazarpak, H.H. Impact of drinking water supplementation of a blend of peppermint, coneflower (Echinacea purpurea), thyme, propolis, and prebiotic on performance, serum constituents, and immunocompetence of broiler chickens. Trop. Anim. Health Prod. 2022, 54, 289. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Elbestawy, A.R.; Gado, A.R.; Nader, M.M.; Saad, A.M.; El-Tahan, A.M.; Taha, A.E.; Salem, H.M.; El-Tarabily, K.A. Hot red pepper powder as a safe alternative to antibiotics in organic poultry feed: An updated review. Poult. Sci. 2022, 101, 101684. [Google Scholar] [CrossRef]
- Tufarelli, V.; Ghavami, N.; Nosrati, M.; Rasouli, B.; Kadim, I.T.; Suárez Ramírez, L.; Gorlov, I.; Slozhenkina, M.; Mosolov, A.; Seidavi, A.; et al. The effects of peppermint (Mentha piperita L.) and chicory (Cichorium intybus L.) in comparison with a prebiotic on productive performance, blood constituents, immunity and intestinal microflora in broiler chickens. Anim. Biotechnol. 2023, 34, 3046–3052. [Google Scholar] [CrossRef]
- Lin, P.-H.; Chen, Z.-W.; Liu, J.-Y.; Ye, J.-C. Dietary supplementation of Ocimum gratissimum improves growth performance and immune response in broilers under high ambient temperature. J. Anim. Sci. 2023, 101, skad212. [Google Scholar] [CrossRef]
- Alagawany, M.; Elnesr, S.S.; Farag, M.R.; Abd El-Hack, M.E.; Khafaga, A.F.; Taha, A.E.; Tiwari, R.; Yatoo, M.I.; Bhatt, P.; Marappan, G.; et al. Use of Licorice (Glycyrrhiza glabra) herb as a feed additive in poultry: Current knowledge and prospects. Animals 2019, 9, 536. [Google Scholar] [CrossRef]
- Yuan, S.; Wang, Q.; Li, J.; Xue, J.C.; Li, Y.; Meng, H.; Hou, X.T.; Nan, J.X.; Zhang, Q.G. Inflammatory bowel disease: An overview of Chinese herbal medicine formula-based treatment. Chin. Med. 2022, 17, 74. [Google Scholar] [CrossRef]
- Liu, Y.; Liang, S.; Zi, X.; Yan, S.; Liu, M.; Li, M.; Zhao, Y.; Dou, T.; Ge, C.; Wang, K.; et al. Influence of Chinese herbal formula on bone characteristics of cobb broiler chickens. Genes 2022, 13, 1865. [Google Scholar] [CrossRef]
- Ghafouri, S.A.; Ghaniei, A.; Tamannaei, A.E.T.; Sadr, S.; Charbgoo, A.; Ghiassi, S.; Abuali, M. Evaluation of therapeutic effects of an herbal mixture (Echinacea purpurea and Glycyrrhiza glabra) for treatment of clinical coccidiosis in broilers. Vet. Med. Sci. 2023, 9, 829–836. [Google Scholar] [CrossRef]
- Travel, A.; Petit, A.; Barat, P.; Collin, A.; Bourrier-Clairat, C.; Pertusa, M.; Skiba, F.; Crochet, S.; Cailleau-Audouin, E.; Chartrin, P.; et al. Methodologies to assess the bioactivity of an herbal extract on immunity, health, welfare and production performance in the chicken: The case of Melissa officinalis L. extract. Front. Vet. Sci. 2021, 8, 759456. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, R.; Jia, H.; Zhu, Z.; Li, H.; Ma, Y. Supplementation of probiotics in water beneficial growth performance, carcass traits, immune function, and antioxidant capacity in broiler chickens. Open Life Sci. 2021, 16, 311–322. [Google Scholar] [CrossRef]
- Sławińska, A.; Siwek, M.; Zylińska, J.; Bardowski, J.; Brzezińska, J.; Gulewicz, K.A.; Nowak, M.; Urbanowski, M.; Płowiec, A.; Bednarczyk, M. Influence of synbiotics delivered in ovo on immune organs development and structure. Folia Biol. 2014, 62, 277–285. [Google Scholar] [CrossRef]
- Sjofjan, O.; Adli, D.N.; Harahap, R.P.; Jayanegara, A.; Utama, D.T.; Seruni, A.P. The effects of lactic acid bacteria and yeasts as probiotics on the growth performance, relative organ weight, blood parameters, and immune responses of broiler: A meta-analysis. F1000Research 2021, 10, 183. [Google Scholar] [CrossRef]
- Li, S.; Ren, L.; Zhu, X.; Li, J.; Zhang, L.; Wang, X.; Gao, F.; Zhou, G. Immunomodulatory effect of γ-irradiated Astragalus polysaccharides on immunosuppressed broilers. Anim. Sci. J. 2019, 90, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Mao, S.; Zhou, M. Effect of the flavonoid baicalein as a feed additive on the growth performance, immunity, and antioxidant capacity of broiler chickens. Poult. Sci. 2019, 98, 2790–2799. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Xie, K.; Zhang, Y.; Xie, Q.; He, X.; Zhang, H. Effects of dietary ginsenoside Rg1 supplementation on growth performance, gut health, and serum immunity in broiler chickens. Front. Nutr. 2021, 8, 705279. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Maksoud, E.M.; Daha, A.; Taha, N.M.; Lebda, M.A.; Sadek, K.M.; Alshahrani, M.Y.; Ahmed, A.E.; Shukry, M.; Fadl, S.E.; Elfeky, M. Effects of ginger extract and/or propolis extract on immune system parameters of vaccinated broilers. Poult. Sci. 2023, 102, 102903. [Google Scholar] [CrossRef] [PubMed]
- Diaz Carrasco, J.M.; Casanova, N.A.; Fernández Miyakawa, M.E. Microbiota, gut health and chicken productivity: What is the connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef] [PubMed]
- Wickramasuriya, S.S.; Park, I.; Lee, K.; Lee, Y.; Kim, W.H.; Nam, H.; Lillehoj, H.S. Role of physiology, immunity, microbiota, and infectious diseases in the gut health of poultry. Vaccines 2022, 10, 172. [Google Scholar] [CrossRef] [PubMed]
- Alharthi, A.S.; Alruwaili, N.W.; Al-Baadani, H.H.; Al-Garadi, M.A.; Shamlan, G.; Alhidary, I.A. Investigating the effect of Pulicaria jaubertii as a natural feed additive on the growth performance, blood biochemistry, immunological response, and cecal microbiota of broiler chickens. Animals 2023, 13, 1116. [Google Scholar] [CrossRef]
- Khan, S.; Moore, R.J.; Stanley, D.; Chousalkar, K.K. The gut microbiota of laying hens and its manipulation with prebiotics and probiotics to enhance gut health and food safety. Appl. Environ. Microbiol. 2020, 86, e00600-20. [Google Scholar] [CrossRef]
- Zafar, H.; Saier, M.H., Jr. Gut Bacteroides species in health and disease. Gut Microbes 2021, 13, 1848158. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar Reddy, D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Luo, Y.H.; Peng, H.W.; Wright, A.D.; Bai, S.P.; Ding, X.M.; Zeng, Q.F.; Li, H.; Zheng, P.; Su, Z.W.; Cui, R.Y.; et al. Broilers fed dietary vitamins harbor higher diversity of cecal bacteria and higher ratio of Clostridium, Faecalibacterium, and Lactobacillus than broilers with no dietary vitamins revealed by 16S rRNA gene clone libraries. Poult. Sci. 2013, 92, 2358–2366. [Google Scholar] [CrossRef]
- Braz, V.S.; Melchior, K.; Moreira, C.G. Escherichia coli as a multifaceted pathogenic and versatile bacterium. Front. Cell Infect. Microbiol. 2020, 10, 548492. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, M.S.; Kim, J.H. Recent updates on outbreaks of shiga toxin-producing Escherichia coli and its potential reservoirs. Front. Cell Infect. Microbiol. 2020, 10, 273. [Google Scholar] [CrossRef] [PubMed]
- Mirsepasi-Lauridsen, H.C.; Vallance, B.A.; Krogfelt, K.A.; Petersen, A.M. Escherichia coli pathobionts associated with inflammatory bowel disease. Clin. Microbiol. Rev. 2019, 32, e00060-18. [Google Scholar] [CrossRef]
- Song, B.; Li, P.; Yan, S.; Liu, Y.; Gao, M.; Lv, H.; Lv, Z.; Guo, Y. Effects of dietary Astragalus polysaccharide supplementation on the Th17/Treg balance and the gut microbiota of broiler chickens challenged with necrotic enteritis. Front. Immunol. 2022, 13, 781934. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, Q.; Ye, F.; Tang, H.; Xiong, Y.; Wu, Y.; Wang, L.; Feng, X.; Zhang, S.; Wan, Y.; et al. Dietary Purslane (Portulaca oleracea L.) promotes the growth performance of broilers by modulation of gut microbiota. AMB Express 2021, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; He, J.; Zhang, J.; Zhang, X.; Zhang, D.; Zhou, L.; Yuan, Y.; Fu, S.; Qiu, Y.; Ye, C.; et al. Baicalin-aluminum modulates the broiler gut microbiome. DNA Cell Biol. 2021, 40, 881–894. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Li, T.; Niu, H.; Zhu, Y.; Liu, Y.; Duan, Y.; Sun, Q.; Yang, X. Effects of glucose oxidase on growth performance, gut function, and cecal microbiota of broiler chickens. Poult. Sci. 2019, 98, 828–841. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Gong, L.; Zhou, Y.; Tang, L.; Zeng, Z.; Wang, Q.; Zou, P.; Yu, D.; Li, W. Probiotic Paenibacillus polymyxa 10 and Lactobacillus plantarum 16 enhance growth performance of broilers by improving the intestinal health. Anim. Nutr. 2021, 7, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Grondin, J.M.; Déjean, G.; Van Petegem, F.; Brumer, H. Cell surface xyloglucan recognition and hydrolysis by the human gut commensal Bacteroides uniformis. Appl. Environ. Microbiol. 2022, 88, e0156621. [Google Scholar] [CrossRef] [PubMed]
- Medina-Larqué, A.S.; Rodríguez-Daza, M.C.; Roquim, M.; Dudonné, S.; Pilon, G.; Levy, É.; Marette, A.; Roy, D.; Jacques, H.; Desjardins, Y. Cranberry polyphenols and agave agavins impact gut immune response and microbiota composition while improving gut barrier function, inflammation, and glucose metabolism in mice fed an obesogenic diet. Front. Immunol. 2022, 13, 871080. [Google Scholar] [CrossRef] [PubMed]
- Vallianou, N.G.; Kounatidis, D.; Tsilingiris, D.; Panagopoulos, F.; Christodoulatos, G.S.; Evangelopoulos, A.; Karampela, I.; Dalamaga, M. The role of next-generation probiotics in obesity and obesity-associated disorders: Current knowledge and future perspectives. Int. J. Mol. Sci. 2023, 24, 6755. [Google Scholar] [CrossRef]
- Ferreira-Halder, C.V.; Faria, A.V.S.; Andrade, S.S. Action and function of Faecalibacterium prausnitzii in health and disease. Best Pract. Res. Clin. Gastroenterol. 2017, 31, 643–648. [Google Scholar] [CrossRef]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef] [PubMed]
- Miquel, S.; Leclerc, M.; Martin, R.; Chain, F.; Lenoir, M.; Raguideau, S.; Hudault, S.; Bridonneau, C.; Northen, T.; Bowen, B.; et al. Identification of metabolic signatures linked to anti-inflammatory effects of Faecalibacterium prausnitzii. mBio 2015, 6, e00300-15. [Google Scholar] [CrossRef] [PubMed]
- De Cesare, A.; Sala, C.; Castellani, G.; Astolfi, A.; Indio, V.; Giardini, A.; Manfreda, G. Effect of Lactobacillus acidophilus D2/CSL (CECT 4529) supplementation in drinking water on chicken crop and caeca microbiome. PLoS ONE 2020, 15, e0228338. [Google Scholar] [CrossRef] [PubMed]
- Shad, A.A.; Shad, W.A. Shigella sonnei: Virulence and antibiotic resistance. Arch. Microbiol. 2021, 203, 45–58. [Google Scholar] [CrossRef]
- Anokwuru, C.P.; Tankeu, S.; van Vuuren, S.; Viljoen, A.; Ramaite, I.D.I.; Taglialatela-Scafati, O.; Combrinck, S. Unravelling the antibacterial activity of Terminalia sericea Root bark through a metabolomic approach. Molecules 2020, 25, 3683. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | 1~21 d | 22~35 d | ||
|---|---|---|---|---|
| CON | DHF | CON | DHF | |
| Corn (%) | 54.75 | 53.75 | 57.68 | 56.68 |
| Soybean meal (%) | 37.00 | 37.00 | 33.30 | 33.30 |
| Soya bean oil (%) | 4.10 | 4.10 | 5.20 | 5.20 |
| Calcium hydrogen phosphate (%) | 1.35 | 1.35 | 1.40 | 1.40 |
| Stone powder (%) | 1.15 | 1.15 | 0.90 | 0.90 |
| DHF (%) | - | 1.00 | - | 1.00 |
| Salt (%) | 0.30 | 0.30 | 0.30 | 0.30 |
| Methionine (%) | 0.20 | 0.20 | 0.12 | 0.12 |
| Lysine (%) | 0.15 | 0.15 | 0.10 | 0.10 |
| Premix 1 (%) | 1.00 | 1.00 | 1.00 | 1.00 |
| Total (%) | 100.00 | 100 | 100 | 100 |
| Metabolizable energy (kcal/kg) | 3002.00 | 2971.36 | 3100.00 | 3069.36 |
| Crude protein (%) | 21.50 | 21.49 | 20.00 | 19.99 |
| Lysine (%) | 1.27 | 1.27 | 1.13 | 1.13 |
| Methionine (%) | 0.51 | 0.51 | 0.42 | 0.42 |
| Tryptophan (%) | 0.25 | 0.25 | 0.23 | 0.23 |
| Arginine (%) | 1.46 | 1.46 | 1.35 | 1.35 |
| Threonine (%) | 0.80 | 0.80 | 0.74 | 0.74 |
| Valine (%) | 0.98 | 0.98 | 0.92 | 0.92 |
| Isoleucine (%) | 0.87 | 0.87 | 0.81 | 0.81 |
| Leucine (%) | 1.75 | 1.74 | 1.65 | 1.64 |
| Calcium (%) | 1.00 | 1.00 | 0.90 | 0.90 |
| Total phosphorus (%) | 0.68 | 0.68 | 0.66 | 0.66 |
| Non-phytate phosphorus (%) | 0.45 | 0.45 | 0.40 | 0.40 |
| Items | CON | 0.5%DHF | 1.0%DHF | 1.5%DHF | SEM | p-Value |
|---|---|---|---|---|---|---|
| Initial weight (g) | 39.58 ± 0.35 | 39.60 ± 0.18 | 39.75 ± 0.21 | 39.65 ± 0.39 | 0.07 | 0.843 |
| 21 day weight (g) | 617.5 ± 47.05 b | 605 ± 15.56 b | 725.3 ± 50.61 a | 618 ± 76.17 b | 17.13 | 0.022 |
| ADG (g) | 27.51 ± 2.23 b | 26.92 ± 0.75 b | 32.98 ± 2.49 a | 27.54 ± 3.63 b | 0.85 | 0.016 |
| ADFI (g) | 43.93 ± 2.99 | 45.02 ± 1.88 | 47.09 ± 1.51 | 44.90 ± 2.93 | 0.62 | 0.349 |
| FCR | 1.60 ± 0.10 b | 1.62 ± 0.13 b | 1.43 ± 0.07 a | 1.64 ± 0.01 b | 0.05 | 0.049 |
| Items | Groups | p-Value | |||
|---|---|---|---|---|---|
| CON | DHF | Time | Diet | Time × Diet | |
| Body weight (g) | <0.001 | <0.001 | 0.029 | ||
| 21 day | 617.60 ± 16.40 | 673.28 ± 13.67 ** | 0.002 | ||
| 35 day | 1864.90 ± 61.80 | 1995.60 ± 19.04 ** | 0.004 | ||
| ADG (g) | <0.001 | <0.001 | 0.541 | ||
| 21 day | 27.35 ± 0.77 | 30.39 ± 1.10 ** | 0.006 | ||
| 35 day | 52.05 ± 1.76 | 55.78 ± 0.54 ** | 0.004 | ||
| ADFI (g) | <0.001 | 0.118 | 0.892 | ||
| 21 day | 42.23 ± 1.82 | 43.33 ± 0.92 | 0.288 | ||
| 35 day | 95.18 ± 0.47 | 96.11 ± 1.21 | 0.263 | ||
| FCR | <0.001 | <0.001 | 0.092 | ||
| 21 day | 1.54 ± 0.03 | 1.43 ± 0.02 ** | 0.001 | ||
| 35 day | 1.78 ± 0.03 | 1.70 ± 0.02 ** | 0.004 | ||
| Items | Groups | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| CON | DHF | Time | Diet | Time × Diet | ||
| Heart | <0.001 | 0.040 | 0.253 | |||
| 21 day | 5.64 ± 0.67 | 5.38 ± 0.76 | 0.20 | 0.543 | ||
| 35 day | 4.26 ± 0.72 | 3.40 ± 0.18 * | 0.19 | 0.017 | ||
| Liver | <0.001 | 0.783 | 0.721 | |||
| 21 day | 23.59 ± 2.34 | 23.16 ± 1.81 | 0.58 | 0.730 | ||
| 35 day | 18.69 ± 1.21 | 18.74 ± 0.71 | 0.27 | 0.924 | ||
| Spleen | <0.001 | 0.210 | 0.129 | |||
| 21 day | 0.70 ± 0.13 | 1.00 ± 0.28 * | 0.08 | 0.041 | ||
| 35 day | 1.48 ± 0.27 | 1.45 ± 0.30 | 0.08 | 0.858 | ||
| Bursa of fabricius | <0.001 | 0.037 | 0.055 | |||
| 21 day | 4.07 ± 1.36 | 2.78 ± 0.52 | 0.34 | 0.056 | ||
| 35 day | 0.73 ± 0.12 | 0.67 ± 0.19 | 0.04 | 0.511 | ||
| Pancreas | <0.001 | 0.001 | 0.033 | |||
| 21 day | 4.56 ± 0.45 | 3.67 ± 0.37 ** | 0.18 | 0.004 | ||
| 35 day | 2.58 ± 0.24 | 2.29 ± 0.13 * | 0.07 | 0.027 | ||
| Thymus | <0.001 | 0.041 | 0.006 | |||
| 21 day | 2.71 ± 0.44 | 4.08 ± 0.93 ** | 0.38 | 0.009 | ||
| 35 day | 2.03 ± 0.58 | 1.80 ± 0.47 | 0.15 | 0.753 | ||
| Stomach | <0.001 | 0.044 | 0.533 | |||
| 21 day | 26.82 ± 3.79 | 23.01 ± 4.58 | 1.29 | 0.147 | ||
| 35 day | 15.90 ± 1.25 | 13.83 ± 2.87 | 0.69 | 0.151 | ||
| Duodenum | 0.518 | 0.004 | 0.074 | |||
| 21 day | 11.20 ± 1.88 | 8.46 ± 1.26 * | 0.60 | 0.014 | ||
| 35 day | 9.85 ± 1.04 | 9.11 ± 0.74 | 0.27 | 0.188 | ||
| Cecum | 0.365 | 0.067 | 0.964 | |||
| 21 day | 6.96 ± 1.96 | 5.92 ± 0.51 | 0.42 | 0.234 | ||
| 35 day | 7.50 ± 1.56 | 6.41 ± 0.92 | 0.39 | 0.168 | ||
| Items | Groups | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| CON | DHF | Time | Diet | Time × Diet | ||
| White blood cells (109/L) | <0.001 | 0.015 | 0.020 | |||
| 21 day | 123.57 ± 2.22 | 123.32 ± 2.06 | 0.59 | 0.847 | ||
| 35 day | 140.61 ± 6.59 | 132.12 ± 3.41 * | 1.93 | 0.019 | ||
| Lymphocytes (109/L) | 0.152 | <0.001 | 0.056 | |||
| 21 day | 60.20 ± 0.86 | 62.75 ± 1.13 ** | 0.47 | <0.001 | ||
| 35 day | 59.66 ± 4.30 | 66.30 ± 1.98 ** | 1.36 | 0.006 | ||
| Middle cells (109/L) | <0.001 | 0.353 | 0.724 | |||
| 21 day | 17.79 ± 0.53 | 17.65 ± 0.70 | 0.17 | 0.713 | ||
| 35 day | 19.67 ± 0.39 | 19.37 ± 0.57 | 0.14 | 0.317 | ||
| Heterophils (109/L) | <0.001 | 0.002 | 0.020 | |||
| 21 day | 45.58 ± 2.16 | 42.92 ± 1.58 * | 0.66 | 0.036 | ||
| 35 d | 61.28 ± 10.60 | 46.45 ± 4.38 * | 3.16 | 0.010 | ||
| Lymphocytes (%) | 0.009 | 0.001 | 0.112 | |||
| 21 day | 48.37 ± 1.77 | 51.87 ± 1.60 ** | 0.70 | 0.005 | ||
| 35 d | 42.73 ± 4.99 | 50.33 ± 2.46 ** | 1.58 | 0.007 | ||
| Middle cells (%) | 0.701 | 0.039 | 0.052 | |||
| 21 day | 14.22 ± 0.26 | 14.23 ± 0.39 | 0.09 | 0.932 | ||
| 35 day | 13.95 ± 0.58 | 14.62 ± 0.19 * | 0.16 | 0.023 | ||
| Heterophils (%) | 0.015 | <0.001 | 0.089 | |||
| 21 day | 37.41 ± 1.90 | 33.90 ± 1.32 ** | 0.70 | 0.004 | ||
| 35 day | 43.32 ± 5.54 | 35.05 ± 2.55 | 1.72 | 0.013 | ||
| Red blood cells (1012/L) | 0.397 | 0.633 | 0.139 | |||
| 21 day | 2.46 ± 0.22 | 2.54 ± 0.10 | 0.05 | 0.421 | ||
| 35 day | 2.65 ± 0.25 | 2.49 ± 0.16 | 0.06 | 0.222 | ||
| Hemoglobin (g/L) | 0.914 | 0.052 | 0.107 | |||
| 21 day | 95.17 ± 8.84 | 94.00 ± 5.02 | 1.99 | 0.784 | ||
| 35 day | 100.00 ± 8.72 | 88.50 ± 6.72 * | 2.76 | 0.028 | ||
| Hematokrit (L/L) | 0.541 | 0.136 | 0.188 | |||
| 21 day | 0.24 ± 0.02 | 0.24 ± 0.01 | 0.01 | 0.879 | ||
| 35 day | 0.26 ± 0.02 | 0.24 ± 0.02 | 0.01 | 0.095 | ||
| MCV (fL) | 0.167 | 0.127 | 0.984 | |||
| 21 day | 98.97 ± 1.63 | 97.00 ± 3.18 | 0.76 | 0.207 | ||
| 35 day | 97.20 ± 4.00 | 95.18 ± 2.96 | 1.01 | 0.344 | ||
| MCH (pg) | 0.003 | 0.001 | 0.302 | |||
| 21 day | 38.53 ± 0.43 | 37.15 ± 0.97 * | 0.29 | 0.010 | ||
| 35 day | 37.62 ± 1.04 | 35.42 ± 1.16 ** | 0.45 | 0.006 | ||
| MCHC (g/L) | 0.098 | 0.011 | 0.231 | |||
| 21 day | 369.84 ± 7.20 | 364.17 ± 8.04 | 2.27 | 0.227 | ||
| 35 day | 368.00 ± 12.74 | 353.33 ± 3.31 * | 3.54 | 0.030 | ||
| RDW-SD (fL) | 0.154 | 0.067 | 0.233 | |||
| 21 day | 12.76 ± 3.16 | 10.92 ± 0.13 | 0.62 | 0.183 | ||
| 35 day | 19.88 ± 12.31 | 11.58 ± 2.39 | 2.74 | 0.136 | ||
| RDW-CV (%) | 0.860 | 0.756 | 0.953 | |||
| 21 day | 30.68 ± 2.95 | 30.32 ± 0.79 | 0.60 | 0.774 | ||
| 35 day | 30.45 ± 3.35 | 30.20 ± 1.59 | 0.72 | 0.872 | ||
| Thrombocytes (109/L) | <0.001 | 0.894 | 0.131 | |||
| 21 day | 7.00 ± 2.10 | 10.17 ± 1.60 * | 0.70 | 0.015 | ||
| 35 day | 28.00 ± 6.29 | 25.33 ± 5.99 | 1.74 | 0.469 | ||
| PCT (L/L) | <0.001 | 0.357 | 0.741 | |||
| 21 day | 0.012 ± 0.008 | 0.011 ± 0.005 | 0.002 | 0.666 | ||
| 35 day | 0.032 ± 0.007 | 0.029 ± 0.007 | 0.002 | 0.402 | ||
| MPV (fL) | 0.003 | 0.062 | 0.999 | |||
| 21 day | 11.25 ± 0.10 | 11.08 ± 0.23 | 0.06 | 0.139 | ||
| 35 day | 11.53 ± 0.23 | 11.37 ± 0.23 | 0.07 | 0.237 | ||
| PDW (%) | 0.081 | 0.491 | 0.589 | |||
| 21 day | 70.02 ± 14.66 | 76.83 ± 14.91 | 4.20 | 0.443 | ||
| 35 day | 63.00 ± 11.46 | 63.83 ± 11.98 | 3.22 | 0.904 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Zhang, M.; Shi, D.; Dai, X.; Li, X. Effects of Designed Herbal Formula on Growth Performance, Blood Indices, Organ Traits, and Cecum Microbiology in Broilers. Vet. Sci. 2024, 11, 107. https://doi.org/10.3390/vetsci11030107
Sun Y, Zhang M, Shi D, Dai X, Li X. Effects of Designed Herbal Formula on Growth Performance, Blood Indices, Organ Traits, and Cecum Microbiology in Broilers. Veterinary Sciences. 2024; 11(3):107. https://doi.org/10.3390/vetsci11030107
Chicago/Turabian StyleSun, Yuelong, Mengjie Zhang, Dongdong Shi, Xiaofeng Dai, and Xiumei Li. 2024. "Effects of Designed Herbal Formula on Growth Performance, Blood Indices, Organ Traits, and Cecum Microbiology in Broilers" Veterinary Sciences 11, no. 3: 107. https://doi.org/10.3390/vetsci11030107
APA StyleSun, Y., Zhang, M., Shi, D., Dai, X., & Li, X. (2024). Effects of Designed Herbal Formula on Growth Performance, Blood Indices, Organ Traits, and Cecum Microbiology in Broilers. Veterinary Sciences, 11(3), 107. https://doi.org/10.3390/vetsci11030107