

The Effect of Tumor Necrosis Factor-α and Interleu-Kin-1β on the Restorative Properties of Human Oligodendrocyte Precursor Cells In Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. hOPC Culture and Differentiation
2.2. Immunocytochemistry
2.3. Flow Cytometry
2.4. Cell Counting Kit-8
2.5. Migration Assay
2.6. Cell Proliferation Assay
2.7. RNA-Sequencing Analysis
2.8. Statistical Analyses
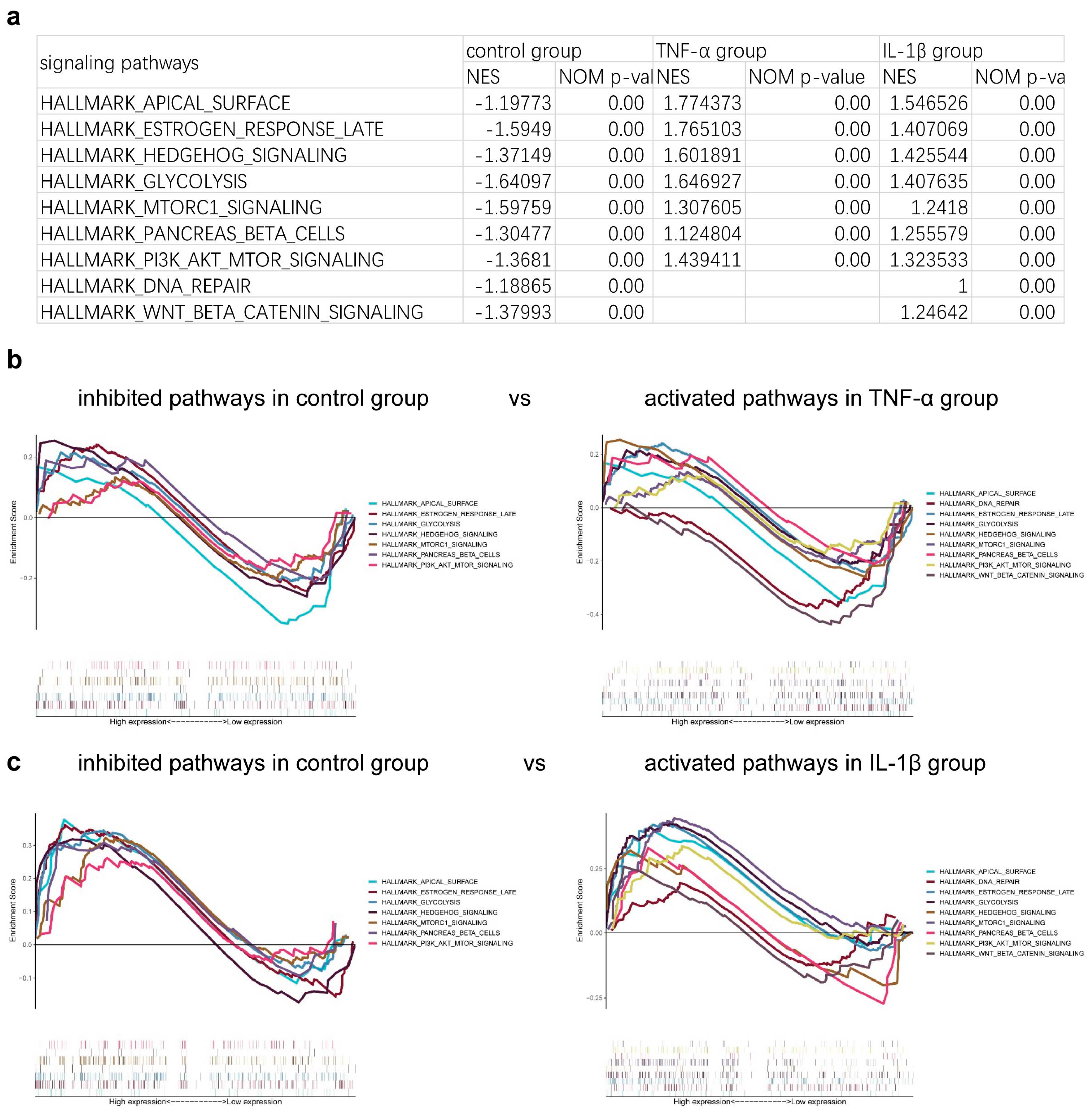
3. Results
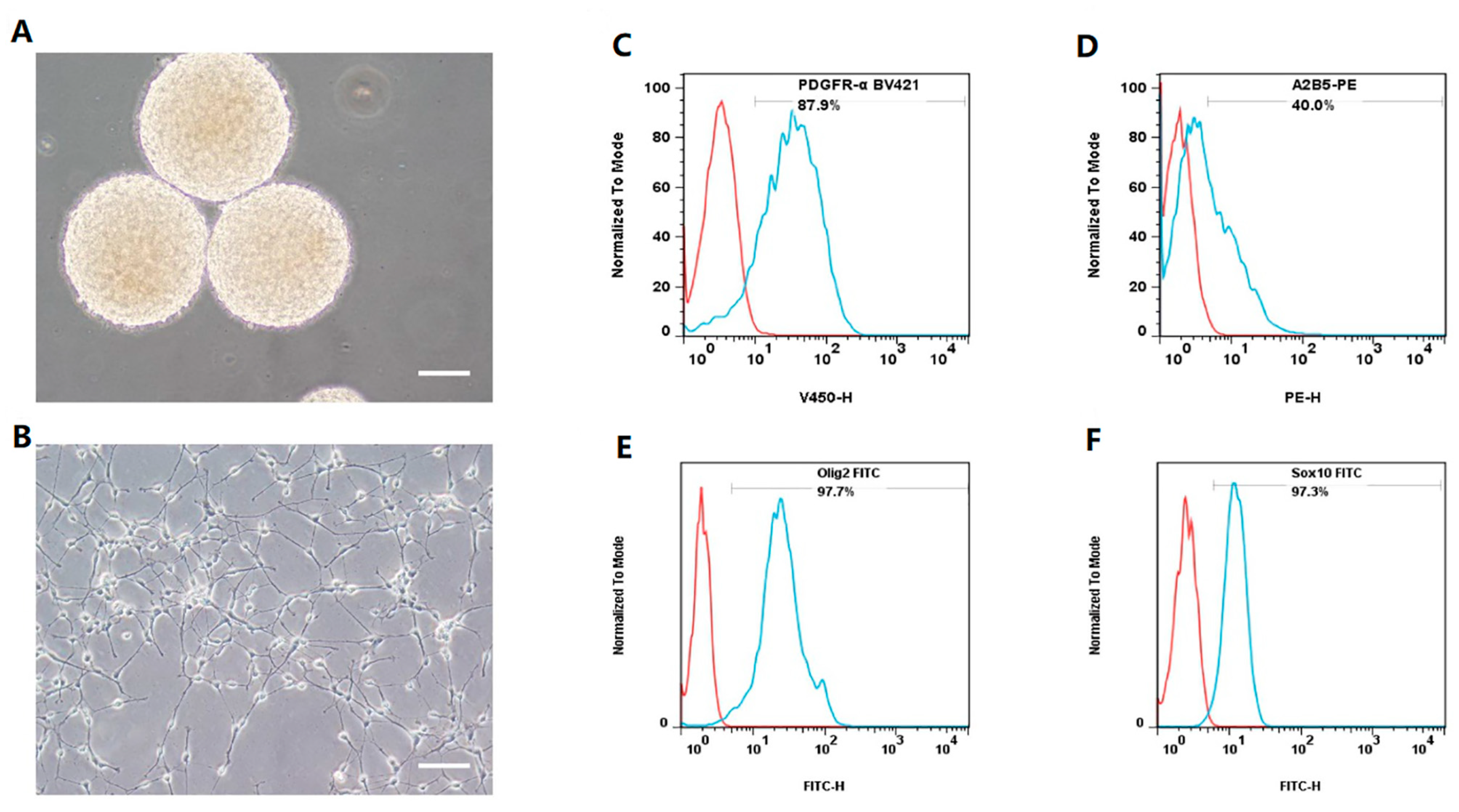
3.1. OLs Are Efficiently Established from hNSC-Derived hOPCs
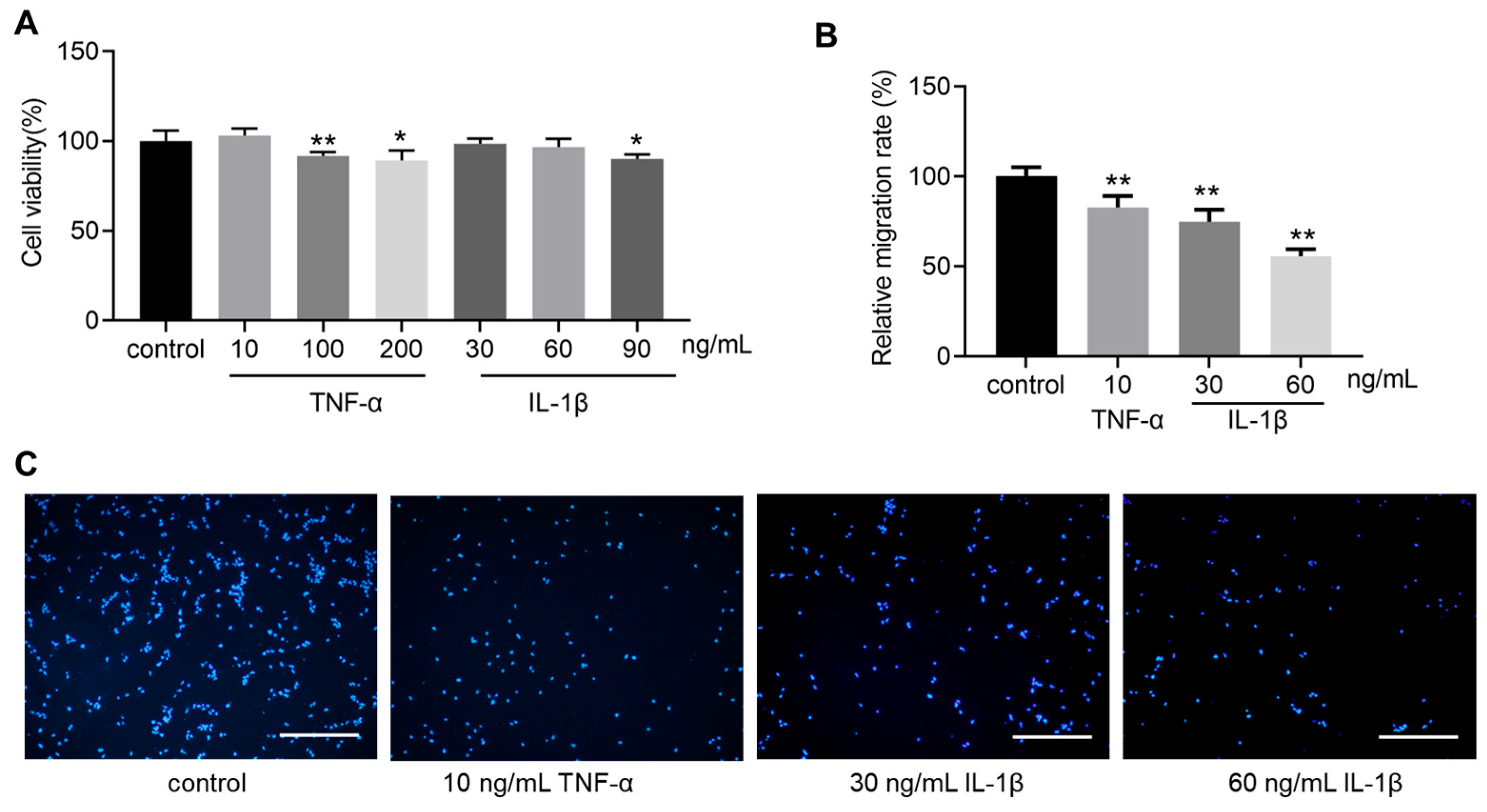
3.2. TNF-α and IL-1β Inhibit hOPC Migration
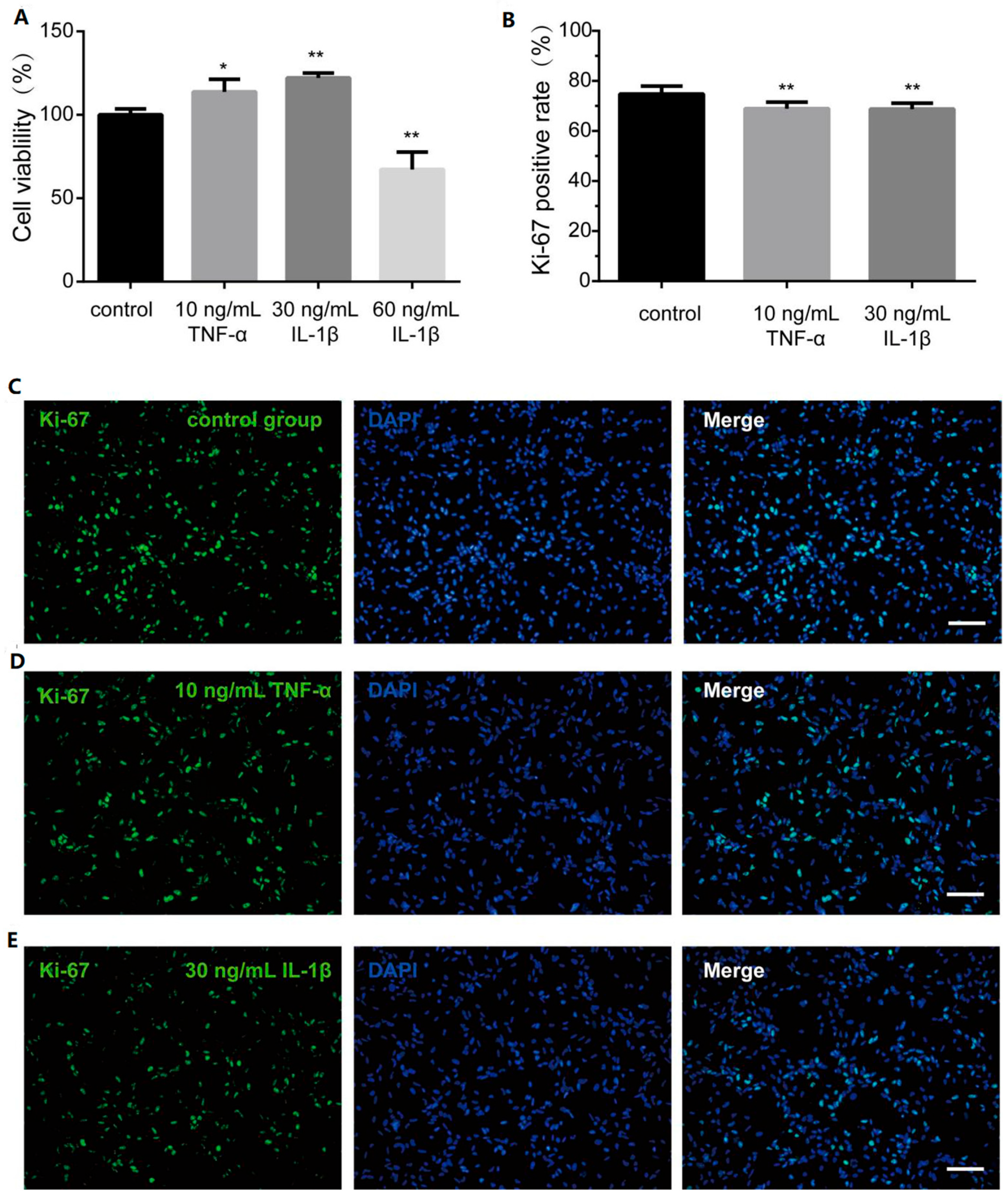
3.3. TNF-α and IL-1β Inhibit hOPC Proliferation
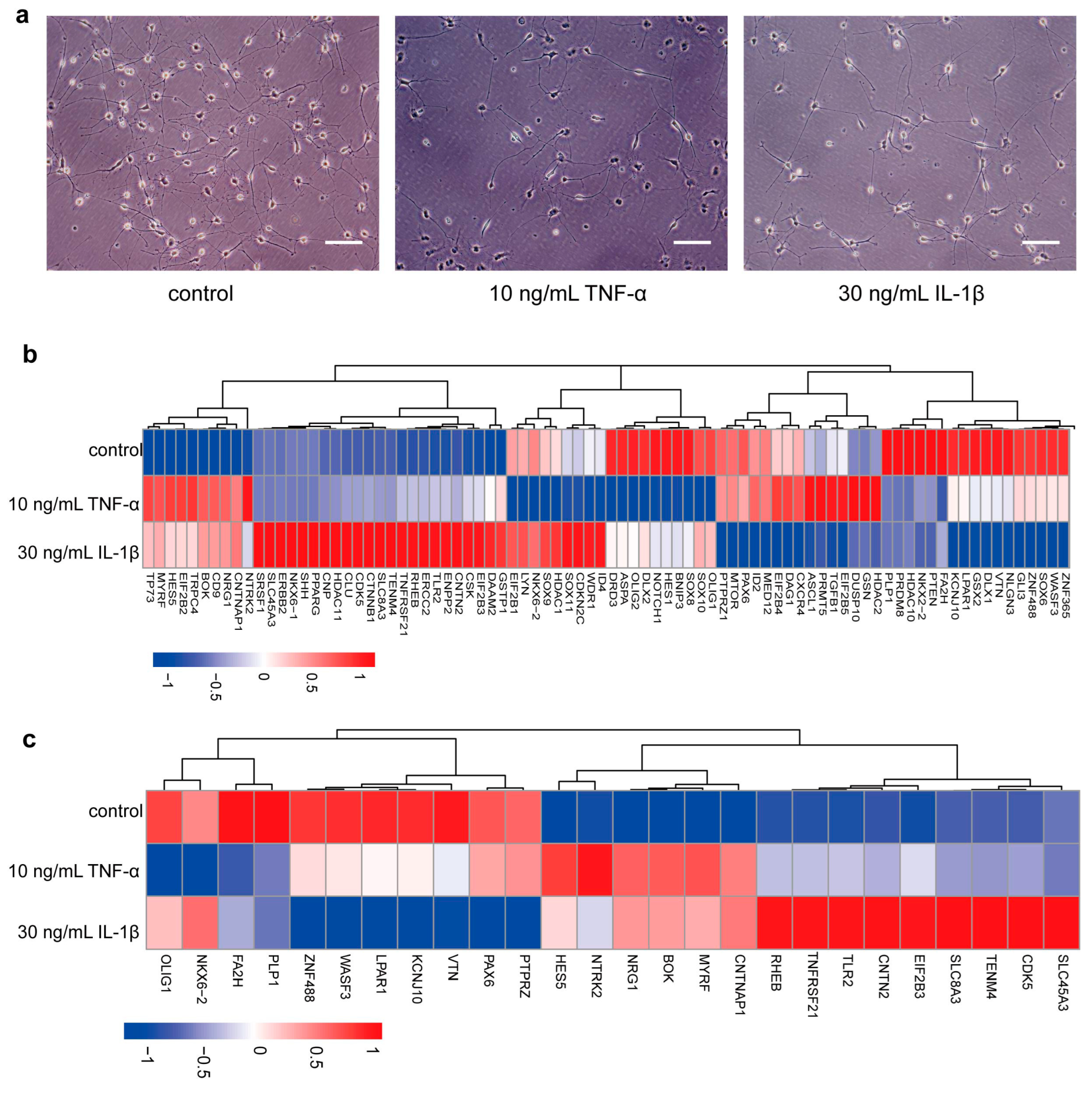
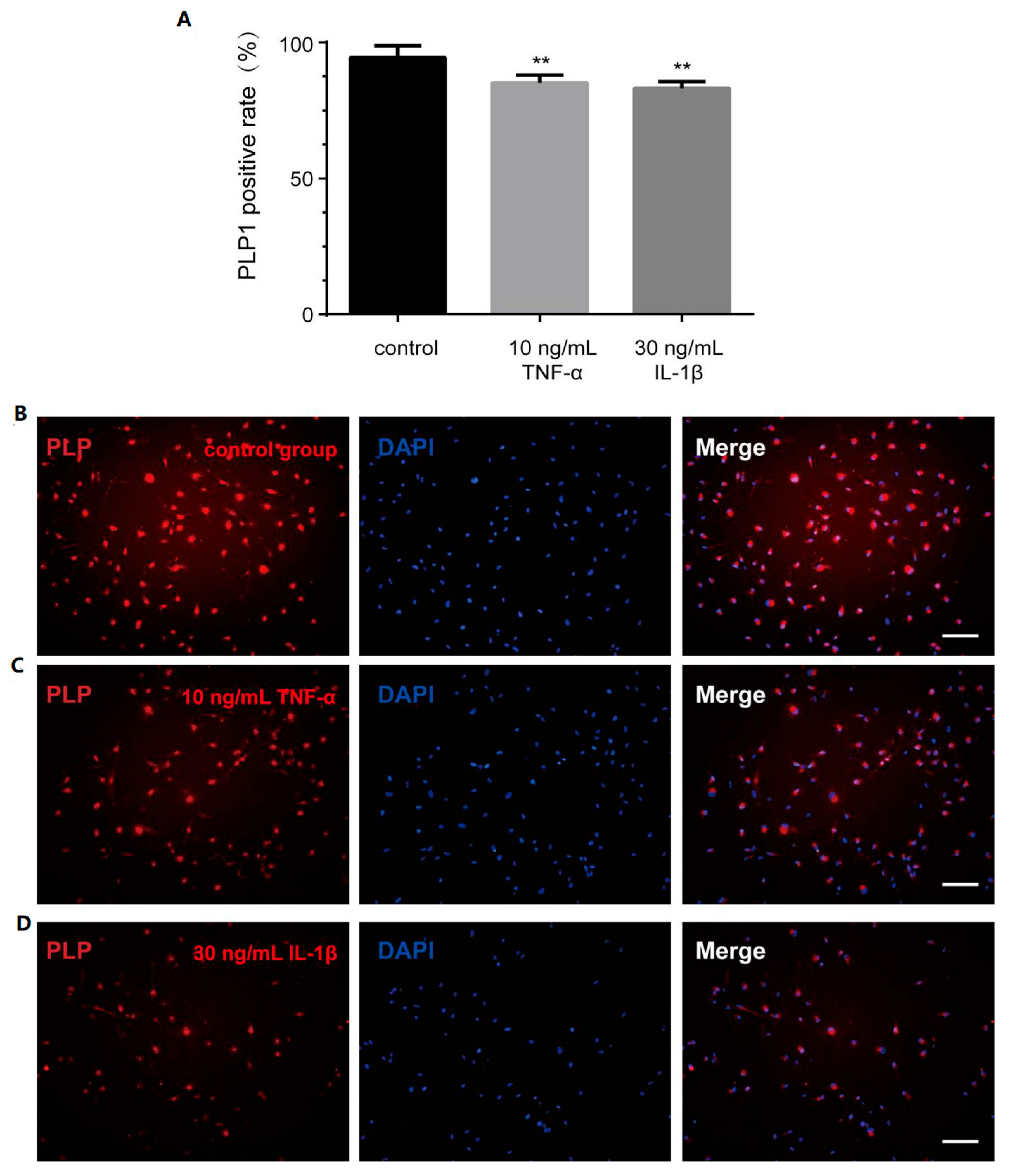
3.4. TNF-α and IL-1β Inhibit PLP1 Expression in Differentiated Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- van Tilborg, E.; Heijnen, C.J.; Benders, M.J.; van Bel, F.; Fleiss, B.; Gressens, P.; Nijboer, C.H. Impaired oligodendrocyte maturation in preterm infants: Potential therapeutic targets. Prog. Neurobiol. 2016, 136, 28–49. [Google Scholar] [CrossRef] [PubMed]
- UNICEF. Levels & Trends in Child Mortality: Report 2019. In Estimates Developed by the United Nations Inter-agency Group for Child Mortality Estimation; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Back, S.A. White matter injury in the preterm infant: Pathology and mechanisms. Acta Neuropathol. 2017, 134, 331–349. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Concepcion, K.; Meng, X.; Zhang, L. Brain-immune interactions in perinatal hypoxic-ischemic brain injury. Prog. Neurobiol. 2017, 159, 50–68. [Google Scholar] [CrossRef] [PubMed]
- Banisadr, G.; Frederick, T.J.; Freitag, C.; Ren, D.; Jung, H.; Miller, S.D.; Miller, R.J. The role of CXCR4 signaling in the migration of transplanted oligodendrocyte progenitors into the cerebral white matter. Neurobiol. Dis. 2011, 44, 19–27. [Google Scholar] [CrossRef]
- Kuhn, S.; Gritti, L.; Crooks, D.; Dombrowski, Y. Oligodendrocytes in Development, Myelin Generation and Beyond. Cells 2019, 8, 1424. [Google Scholar] [CrossRef]
- Nevin, Z.S.; Factor, D.C.; Karl, R.T.; Douvaras, P.; Laukka, J.; Windrem, M.S.; Goldman, S.A.; Fossati, V.; Hobson, G.M.; Tesar, P.J. Modeling the Mutational and Phenotypic Landscapes of Pelizae-us-Merzbacher Disease with Human iPSC-Derived Oligodendrocytes. Am. J. Hum. Genet. 2017, 100, 617–634. [Google Scholar] [CrossRef]
- Ogawa, S.; Hagiwara, M.; Misumi, S.; Tajiri, N.; Shimizu, T.; Ishida, A.; Suzumori, N.; Sugiura-Ogasawara, M.; Hida, H. Transplanted Oligodendrocyte Progenitor Cells Survive in the Brain of a Rat Neonatal White Matter Injury Model but Less Mature in Comparison with the Normal Brain. Cell Transplant. 2020, 29, 963689720946092. [Google Scholar] [CrossRef]
- Piao, J.; Major, T.; Auyeung, G.; Policarpio, E.; Menon, J.; Droms, L.; Gutin, P.; Uryu, K.; Tchieu, J.; Soulet, D.; et al. Human embryonic stem cell-derived oligodendrocyte progenitors re-myelinate the brain and rescue behavioral deficits following radiation. Cell Stem Cell 2015, 16, 198–210. [Google Scholar] [CrossRef]
- Wang, S.; Bates, J.; Li, X.; Schanz, S.; Chandler-Militello, D.; Levine, C.; Maherali, N.; Studer, L.; Hochedlinger, K.; Windrem, M.; et al. Human iPSC-derived oligodendrocyte progenitor cells can myelinate and rescue a mouse model of congenital hypomyelination. Cell Stem Cell 2013, 12, 252–264. [Google Scholar] [CrossRef]
- Kaur, C.; Ling, E.A. Periventricular white matter damage in the hypoxic neonatal brain: Role of microglial cells. Prog. Neurobiol. 2009, 87, 264–280. [Google Scholar] [CrossRef]
- Pellegatta, S.; Tunici, P.; Poliani, P.L.; Dolcetta, D.; Cajola, L.; Colombelli, C.; Ciusani, E.; Di Donato, S.; Finocchiaro, G. The therapeutic potential of neural stem/progenitor cells in murine globoid cell leukodystrophy is conditioned by macrophage/microglia activation. Neurobiol. Dis. 2006, 21, 314–323. [Google Scholar] [CrossRef]
- Kuai, X.L.; Ni, R.Z.; Zhou, G.X.; Mao, Z.B.; Zhang, J.F.; Yi, N.; Liu, Z.X.; Shao, N.; Ni, W.K.; Wang, Z.W. Transplantation of mouse embryonic stem cell-derived oligoden-drocytes in the murine model of globoid cell leukodystrophy. Stem Cell Res. Ther. 2015, 6, 30. [Google Scholar] [CrossRef] [PubMed]
- Giannakopoulou, A.; Grigoriadis, N.; Polyzoidou, E.; Lourbopoulos, A.; Michaloudi, E.; Papadopoulos, G.C. Time-dependent fate of transplanted neural precursor cells in experimental autoimmune encephalomyelitis mice. Exp. Neurol. 2011, 230, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.J.; Tatara, A.; McCreedy, D.A.; Shiu, A.; Sakiyama-Elbert, S.E. Tissue-engineered fibrin scaffolds containing neural progenitors enhance functional recovery in a subacute model of SCI. Soft Matter 2010, 6, 5127–5137. [Google Scholar] [CrossRef] [PubMed]
- Karimi-Abdolrezaee, S.; Eftekharpour, E.; Wang, J.; Morshead, C.M.; Fehlings, M.G. Delayed transplantation of adult neural precursor cells promotes remyelination and functional neurological recovery after spinal cord injury. J. Neurosci. 2006, 26, 3377–3389. [Google Scholar] [CrossRef]
- Lu, Y.; Yang, Y.; Wang, Z.; Wang, C.; Du, Q.; Wang, Q.; Luan, Z. Isolation and culture of human oligodendrocyte precursor cells from neurospheres. Brain Res. Bull. 2015, 118, 17–24. [Google Scholar] [CrossRef]
- Sandrock, R.W.; Wheatley, W.; Levinthal, C.; Lawson, J.; Hashimoto, B.; Rao, M.; Campanelli, J.T. Isolation, characterization and preclinical development of human glial-restricted progenitor cells for treatment of neurological disorders. Regen. Med. 2010, 5, 381–394. [Google Scholar] [CrossRef]
- Ninos, J.M.; Jefferies, L.C.; Cogle, C.R.; Kerr, W.G. The thrombopoietin receptor, c-Mpl, is a selective surface marker for human hematopoietic stem cells. J. Transl. Med. 2006, 4, 9. [Google Scholar] [CrossRef]
- Quah, B.J.; Barlow, V.P.; McPhun, V.; Matthaei, K.I.; Hulett, M.D.; Parish, C.R. Bystander B cells rapidly acquire antigen receptors from activated B cells by membrane transfer. Proc. Natl. Acad. Sci. USA 2008, 105, 4259–4264. [Google Scholar] [CrossRef]
- Lüders, K.A.; Nessler, S.; Kusch, K.; Patzig, J.; Jung, R.B.; Möbius, W.; Nave, K.A.; Werner, H.B. Maintenance of high proteolipid protein level in adult central nervous system myelin is required to preserve the integrity of myelin and axons. Glia 2019, 67, 634–649. [Google Scholar] [CrossRef]
- Natalucci, G.; Latal, B.; Koller, B.; Rüegger, C.; Sick, B.; Held, L.; Bucher, H.U.; Fauchère, J.C. Swiss EPO Neuroprotection Trial Group. Effect of Early Prophylactic High-Dose Recombinant Human Erythropoietin in Very Preterm Infants on Neurodevelopmental Outcome at 2 Years: A Randomized Clinical Trial. JAMA 2016, 315, 2079–2085. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Park, D.; Ban, Y.H.; Cha, Y.; An, E.S.; Choi, J.; Choi, E.K.; Kim, Y.B. Improvement by Human Oligodendrocyte Progenitor Cells of Neurobehavioral Disorders in an Experimental Model of Neonatal Periventricular Leu-komalacia. Cell Transpl. 2018, 27, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.X.; Ma, S.M.; Zhang, P.; Fan, Z.C.; Xiong, M.; Cheng, G.Q.; Yang, Y.; Qiu, Z.L.; Zhou, W.H.; Li, J. Neuroprotective effects of oligodendrocyte progenitor cell transplantation in premature rat brain following hypoxic-ischemic injury. PLoS ONE 2015, 10, e0115997. [Google Scholar]
- Webber, D.J.; van Blitterswijk, M.; Chandran, S. Neuroprotective effect of oligodendrocyte precursor cell transplantation in a long-term model of periventricular leukomalacia. Am. J. Pathol. 2009, 175, 2332–2342. [Google Scholar] [CrossRef]
- Windrem, M.S.; Nunes, M.C.; Rashbaum, W.K.; Schwartz, T.H.; Goodman, R.A.; McKhann, G., 2nd; Roy, N.S.; Goldman, S.A. Fetal and adult human oligodendrocyte progenitor cell isolates myelinate the congenitally dysmyelinated brain. Nat. Med. 2004, 10, 93–97. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, X.Y.; Casella, G.; Tian, J.; Ye, Z.Q.; Yang, T.; Han, J.J.; Jia, L.Y.; Rostami, A.; Li, X. Generation of Oligodendrocyte Progenitor Cells from Mouse Bone Marrow Cells. Front. Cell. Neurosci. 2019, 13, 247. [Google Scholar] [CrossRef]
- Roy, N.S.; Cleren, C.; Singh, S.K.; Yang, L.; Beal, M.F.; Goldman, S.A. Functional engraftment of human ES cell-derived dopaminergic neurons enriched by coculture with telomerase-immortalized midbrain astrocytes. Nat. Med. 2006, 12, 1259–1268. [Google Scholar] [CrossRef]
- Capetian, P.; Azmitia, L.; Pauly, M.G.; Krajka, V.; Stengel, F.; Bernhardi, E.M.; Klett, M.; Meier, B.; Seibler, P.; Stanslowsky, N.; et al. Plasmid-Based Generation of Induced Neural Stem Cells from Adult Human Fibroblasts. Front. Cell. Neurosci. 2016, 10, 245. [Google Scholar] [CrossRef]
- van Tilborg, E.; de Theije, C.G.M.; van Hal, M.; Wagenaar, N.; de Vries, L.S.; Benders, M.J.; Rowitch, D.H.; Nijboer, C.H. Origin and dynamics of oligodendrocytes in the developing brain: Implications for perinatal white matter injury. Glia 2018, 66, 221–238. [Google Scholar] [CrossRef]
- Vela, J.M.; Molina-Holgado, E.; Arévalo-Martín, A.; Almazán, G.; Guaza, C. Interleukin-1 regulates proliferation and differentiation of oligodendrocyte progenitor cells. Mol. Cell Neurosci. 2002, 20, 489–502. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, J.; Wang, L.; Chen, Y.; Wan, Y.; He, Y.; Jiang, L.; Ma, J.; Liao, R.; Zhang, X.; et al. Interleukin-1β impedes oligodendrocyte progenitor cell recruitment and white matter repair following chronic cerebral hypoperfusion. Brain Behav. Immun. 2017, 60, 93–105. [Google Scholar] [CrossRef]
- Patel, J.R.; Williams, J.L.; Muccigrosso, M.M.; Liu, L.; Sun, T.; Rubin, J.B.; Klein, R.S. Astrocyte TNFR2 is required for CXCL12-mediated regulation of oligodendrocyte progenitor proliferation and differentiation within the adult CNS. Acta Neuropathol. 2012, 124, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Dziembowska, M.; Tham, T.N.; Lau, P.; Vitry, S.; Lazarini, F.; Dubois-Dalcq, M. A role for CXCR4 signaling in survival and migration of neural and oligodendrocyte precursors. Glia 2005, 50, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.; Shen, F.; He, S.; Chen, M.; Han, Q.; Fang, M.; Zeng, H.; Chen, C.; Deng, Y. IL-1β induces hypomyelination in the periventricular white matter through inhibition of oligodendrocyte progenitor cell maturation via FYN/MEK/ERK signaling pathway in septic neonatal rats. Glia 2016, 64, 583–602. [Google Scholar] [CrossRef]
- Lentferink, D.H.; Jongsma, J.M.; Werkman, I.; Baron, W. Grey matter OPCs are less mature and less sensitive to IFNγ than white matter OPCs: Consequences for remyelination. Sci. Rep. 2018, 8, 2113. [Google Scholar] [CrossRef] [PubMed]
- Favrais, G.; van de Looij, Y.; Fleiss, B.; Ramanantsoa, N.; Bonnin, P.; Stoltenburg-Didinger, G.; Lacaud, A.; Saliba, E.; Dammann, O.; Gallego, J.; et al. Systemic inflammation disrupts the developmental program of white matter. Ann. Neurol. 2011, 70, 550–565. [Google Scholar] [CrossRef]
- Flygt, J.; Ruscher, K.; Norberg, A.; Mir, A.; Gram, H.; Clausen, F.; Marklund, N. Neutralization of Interleukin-1β following Diffuse Traumatic Brain Injury in the Mouse Attenuates the Loss of Mature Oligodendrocytes. J. Neurotrauma 2018, 35, 2837–2849. [Google Scholar] [CrossRef]
- Duncan, I.D.; Kondo, Y.; Zhang, S.C. The myelin mutants as models to study myelin repair in the leukodystrophies. Neurotherapeutics 2011, 8, 607–624. [Google Scholar] [CrossRef]
- Bonora, M.; De Marchi, E.; Patergnani, S.; Suski, J.M.; Celsi, F.; Bononi, A.; Giorgi, C.; Marchi, S.; Rimessi, A.; Duszyński, J.; et al. Tumor necrosis factor-α impairs oligodendroglial differentiation through a mitochondria-dependent process. Cell Death Differ. 2014, 21, 1198–1208. [Google Scholar] [CrossRef]
- De Nuccio, C.; Bernardo, A.; Cruciani, C.; De Simone, R.; Visentin, S.; Minghetti, L. Peroxisome proliferator activated receptor-γ agonists protect oligodendrocyte progenitors against tumor necrosis factor-alpha-induced damage: Effects on mitochondrial functions and differentiation. Exp. Neurol. 2015, 271, 506–514. [Google Scholar] [CrossRef]
- Xin, M.; Yue, T.; Ma, Z.; Wu, F.F.; Gow, A.; Lu, Q.R. Myelinogenesis and axonal recognition by oligodendrocytes in brain are uncoupled in Olig1-null mice. J. Neurosci. 2005, 25, 1354–1365. [Google Scholar] [CrossRef] [PubMed]
- Katanov, C.; Novak, N.; Vainshtein, A.; Golani, O.; Dupree, J.L.; Peles, E. N-Wasp Regulates Oligodendrocyte Myelination. J. Neurosci. 2020, 40, 6103–6111. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Lee, X.; Hu, Y.; Ji, B.; Shao, Z.; Yang, W.; Huang, G.; Walus, L.; Rhodes, K.; Gong, B.J.; et al. Death receptor 6 negatively regulates oligodendrocyte survival, maturation and myelination. Nat. Med. 2011, 17, 816–821. [Google Scholar] [CrossRef]
- Xu, X.; Yu, Q.; Fang, M.; Yi, M.; Yang, A.; Xie, B.; Yang, J.; Zhang, Z.; Dai, Z.; Qiu, M. Stage-specific regulation of oligodendrocyte development by Hedgehog signaling in the spinal cord. Glia 2020, 68, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Greer, J.M.; Lees, M.B. Myelin proteolipid protein—The first 50 years. Int. J. Biochem. Cell Biol. 2002, 34, 211–215. [Google Scholar] [CrossRef]
- Ozgen, H.; Baron, W.; Hoekstra, D.; Kahya, N. Oligodendroglial membrane dynamics in relation to myelin biogenesis. Cell Mol. Life Sci. 2016, 73, 3291–3310. [Google Scholar] [CrossRef]
- Jana, M.; Pahan, K. Redox regulation of cytokine-mediated inhibition of myelin gene expression in human primary oligodendrocytes. Free Radic. Biol. Med. 2005, 39, 823–831. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; He, Y.; Wang, Q.; Liu, W.; Yang, Y.; Zhou, H.; Ma, X.; Hu, C.; Luan, Z.; Qu, S. The Effect of Tumor Necrosis Factor-α and Interleu-Kin-1β on the Restorative Properties of Human Oligodendrocyte Precursor Cells In Vitro. Bioengineering 2025, 12, 457. https://doi.org/10.3390/bioengineering12050457
Wang Z, He Y, Wang Q, Liu W, Yang Y, Zhou H, Ma X, Hu C, Luan Z, Qu S. The Effect of Tumor Necrosis Factor-α and Interleu-Kin-1β on the Restorative Properties of Human Oligodendrocyte Precursor Cells In Vitro. Bioengineering. 2025; 12(5):457. https://doi.org/10.3390/bioengineering12050457
Chicago/Turabian StyleWang, Zhaoyan, Ying He, Qian Wang, Weipeng Liu, Yinxiang Yang, Haipeng Zhou, Xuexia Ma, Caiyan Hu, Zuo Luan, and Suqing Qu. 2025. "The Effect of Tumor Necrosis Factor-α and Interleu-Kin-1β on the Restorative Properties of Human Oligodendrocyte Precursor Cells In Vitro" Bioengineering 12, no. 5: 457. https://doi.org/10.3390/bioengineering12050457
APA StyleWang, Z., He, Y., Wang, Q., Liu, W., Yang, Y., Zhou, H., Ma, X., Hu, C., Luan, Z., & Qu, S. (2025). The Effect of Tumor Necrosis Factor-α and Interleu-Kin-1β on the Restorative Properties of Human Oligodendrocyte Precursor Cells In Vitro. Bioengineering, 12(5), 457. https://doi.org/10.3390/bioengineering12050457