
Storage Stability and Lipidomic Analysis Reveal the Effect of Frozen Storage Temperature on Pacific Saury (Cololabis saira)


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Reagents
2.3. Lipid Composition and Lipid Oxidation Assay
2.4. Determination of Lipid Profile
2.4.1. Lipid Extraction
2.4.2. LC-MS/MS Method for Lipid Analysis
2.4.3. Identification by Lipid Search
2.5. Statistical Analysis
3. Results
3.1. Lipid Composition Change and Lipid Oxidation Analysis of C. saira at Different Storage Temperatures
3.2. Lipidomics Analysis of C. saira at Different Storage Temperatures
3.3. Multivariate Statistical Analysis of C. saira at Different Storage Temperatures
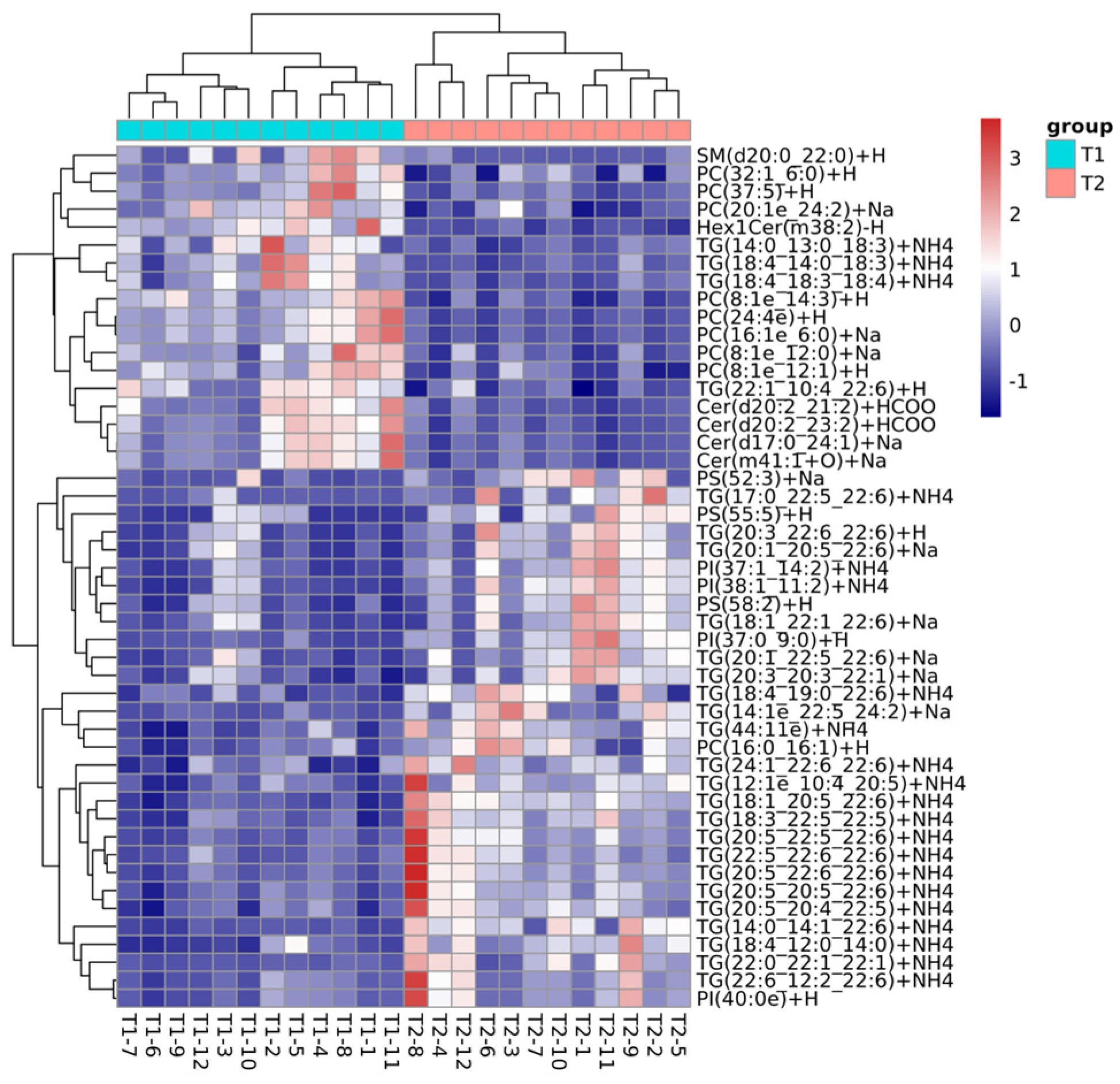
3.4. Differentially Abundant Lipid Analysis
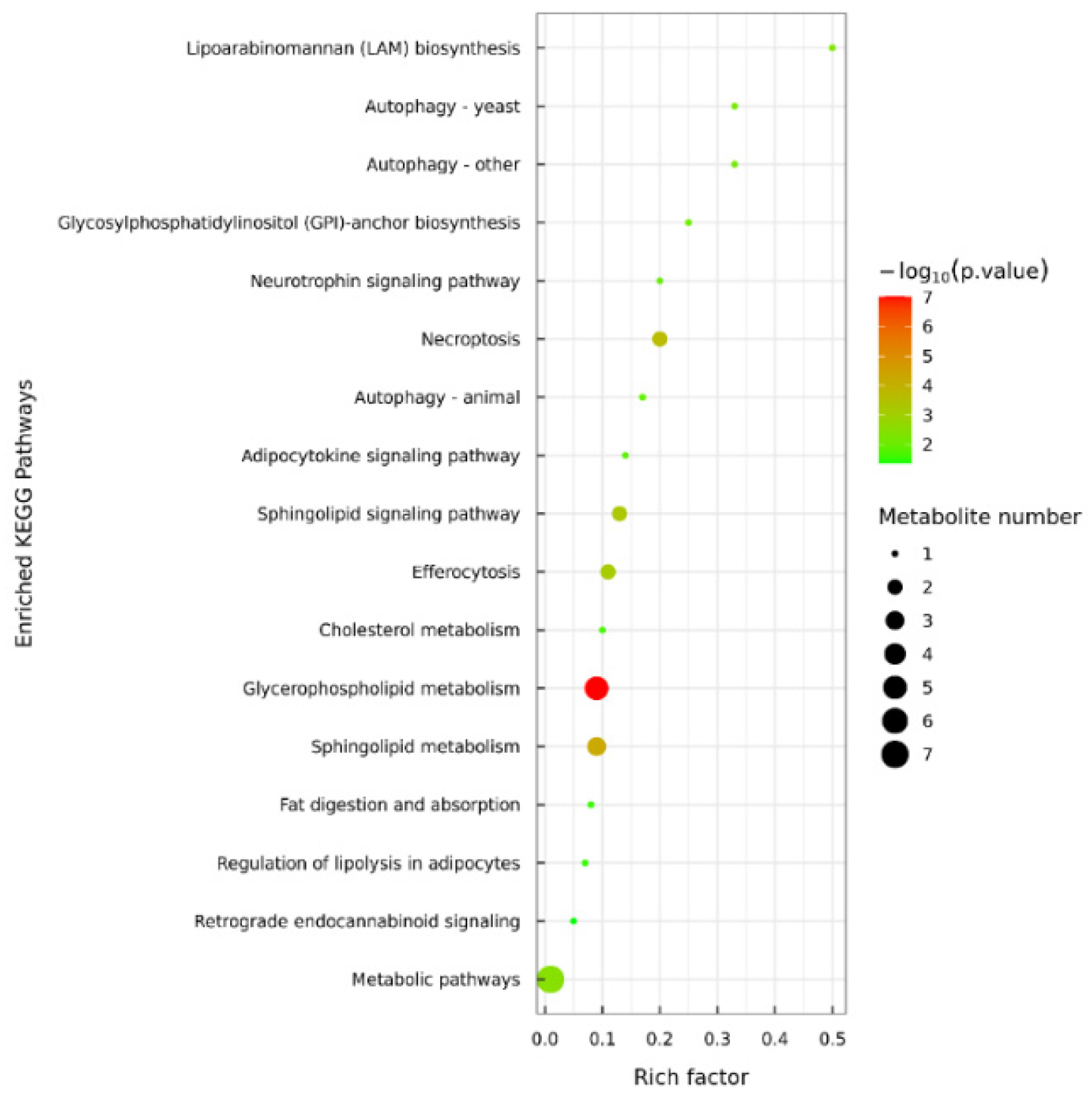
3.5. Lipid Metabolic Pathway Analysis of C. saira During Storage Process
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, X. International fisheries dynamics. Fish. Inf. Strategy 2023, 38, 317–324. [Google Scholar]
- NPFC. NPFC-2023-AR-Annual Summary Footprint-Pacific Saury. 2023. Available online: https://www.npfc.int/summary-footprint-pacific-saury-fisheries (accessed on 11 July 2024).
- Zhang, J.; Tao, N.; Wang, M.; Shi, W.; Ye, B.; Wang, X.; Zhu, Q.; Hua, C. Characterization of phospholipids from Pacific saury (Cololabis saira) viscera and their neuroprotective activity. Food Biosci. 2018, 24, 120–126. [Google Scholar] [CrossRef]
- Liu, H.M.; Yang, G.Q.; Zhao, Q.; Li, H.Y.; Niu, L.H.; Wu, H.Y.; Yu, H. Antioxidant effects of Stevia rebaudiana leaf and stem extracts on lipid oxidation in salted Pacific Saury (Cololabis saira) during processing. Eur. J. Lipid Sci. Technol. 2022, 124, 210022. [Google Scholar] [CrossRef]
- Tao, X.Y.; Yin, M.Y.; Liu, L.; Song, R.Z.; Wang, X.D.; Tao, N.P.; Wang, X.C. UPLC-ESI-MS/MS strategy to analyze fatty acids composition and lipid profiles of Pacific saury (Cololabis saira). Food Chem. X 2024, 23, 101682. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Medina, M.D.; Sáez-Casado, M.; Martínez-Moya, T.; Rincón-Cervera, M.Á. The Effect of Low Temperature Storage on the Lipid Quality of Fish, Either Alone or Combined with Alternative Preservation Technologies. Foods 2024, 13, 1097. [Google Scholar] [CrossRef] [PubMed]
- Indergård, E.; Tolstorebrov, I.; Larsen, H.; Eikevik, T.M. The influence of long-term storage, temperature and type of packaging materials on the quality characteristics of frozen farmed Atlantic Salmon (Salmo salar). Int. J. Refrig. 2014, 41, 27–36. [Google Scholar] [CrossRef]
- Lan, W.Q.; Liu, L.; Xiao, L.; Mei, J.; Xie, J. Effect of Temperature Fluctuation on the Quality of Big-Eye Tuna (Thunnus obesus) during Low Temperature Circulation. Food Sci. 2021, 42, 205–212. [Google Scholar]
- Zhao, L.; Wang, L.; Cao, R.; Liu, Q.; Su, D.; Zhang, Y.T.; Yu, Y.Q. The role of ultraviolet radiation in the flavor formation during drying processing of Pacific saury (Cololabis saira). J. Sci. Food Agric. 2024, 104, 8099–8108. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.Y.; Chen, Y.; Gong, Y.S. Improvement of Theaflavins on Glucose and Lipid Metabolism in Diabetes Mellitus. Foods 2024, 13, 1763. [Google Scholar] [CrossRef]
- Luo, X.; Sun, W.C.; Luo, Y.H. Research progress in detection and function of sphingomyelin in food. Food Res. Dev. 2020, 41, 211–218. [Google Scholar]
- Luo, H.B.; Wang, W.H.; Chen, W.; Tang, H.Q.; Jiang, L.; Yu, Z.F. Effect of incorporation of natural chemicals in water ice-glazing on freshness and shelf-life of Pacific saury (Cololabis saira) during −18 °C frozen storage. J. Sci. Food Agric. 2018, 9, 3309–3314. [Google Scholar] [CrossRef] [PubMed]
- Ding, K.H.; Wang, Y.F.; Luan, D.L. Effects of high-temperature short-time processing on nutrition quality of Pacific saury (Cololabis saira) using extracted fatty acids as the indicator. Food Sci. Nutr. 2022, 1, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, R.; Naiki, K.; Tsuji, K.; Nomata, H.; Sugiura, Y.; Matsushita, T.; Kimura, I. Effect of antioxidative treatment on lipid oxidation in skinless fillet of Pacific Saury (Cololabis saira) in frozen storage. J. Food Process. Preserv. 2013, 37, 325–334. [Google Scholar] [CrossRef]
- Wang, S.Y.; Jian, C.; Hu, M.Y.; Zhao, L.; Sun, H.H.; Liu, Q.; Cao, R.; Xue, Y. Lipid changes and volatile compounds formation in different processing stages of dry-cured Spanish mackerel. Food Qual. Saf. 2024, 8, fyae026. [Google Scholar] [CrossRef]
- GB 5009.181-2016; National Standards for Food Safety Determination of Malondialdehyde in Food. The National Health and Family Planning Commission of the People’s Republic of China: Beijing, China, 2016.
- GB 5009.227-2016; National Standards for Food Safety Determination of Peroxide Value in Food. The National Health and Family Planning Commission of the People’s Republic of China: Beijing, China, 2016.
- GB 5009.229-2016; National Standards for Food Safety Determination of Acid Value in Food. The National Health and Family Planning Commission of the People’s Republic of China: Beijing, China, 2016.
- Shi, C.P.; Guo, H.; Wu, T.T.; Tao, N.P.; Wang, X.C.; Zhong, J. Effect of three types of thermal processing methods on the lipidomics profile of tilapia fillets by UPLC-Q-Extractive Orbitrap mass spectrometry. Food Chem. 2019, 298, 125029. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Q.; Zhao, M.T.; Wang, X.W.; Li, C.; Liu, Z.Y.; Shen, X.R.; Zhou, D.Y. Investigation of oyster Crassostrea gigas lipid profile from three sea areas of China based on non-targeted lipidomics for their geographic region traceability. Food Chem. 2022, 386, 132748. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, X.C.; Ding, Y.T.; Ke, Z.G.; Zhou, X.X.; Zhang, J.Y. Diversity and succession of the microbial community and its correlation with lipid oxidation in dry-cured black carp (Mylopharyngodon piceus) during storage. Food Microbiol. 2021, 98, 103686. [Google Scholar] [CrossRef]
- Fang, C.D.; Chen, H.S.; Yan, H.B.; Shui, S.S.; Benjakul, S.; Zhang, B. Investigation of the changes in the lipid profiles in hairtail (Trichiurus haumela) muscle during frozen storage using chemical and LC/MS-based lipidomics analysis. Food Chem. 2022, 390, 133140. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Caputo, S.; Pantusa, M.; Perri, E.; Sindona, G.; Sportelli, L. Amino acids as modulators of lipoxygenase oxidation mechanism. The identification and structural characterization of spin adducts intermediates by electron spin resonance and tandem mass spectrometry. Food Chem. 2010, 119, 533–538. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Cai, Z.C.; Sang, X.H.; Deng, W.T.; Zeng, L.X.; Wang, J.M.; Zhang, J.H. LC-MS-based lipidomics analyses of alterations in lipid profiles of Asian sea bass (Lates calcarifer) induced by plasma-activated water treatment. Food Res. Int. 2024, 177, 113866. [Google Scholar] [CrossRef] [PubMed]
- Soyer, A.; Özalp, B.; Dalmıs, Ü.; Bilgin, V. Effects of freezing temperature and duration of frozen storage on lipid and protein oxidation in chicken meat. Food Chem. 2010, 120, 1025–1103. [Google Scholar] [CrossRef]
- Hu, L.L.; Ren, S.J.; Shen, Q.; Ye, X.Q.; Chen, J.C.; Ling, J.G. Protein oxidation and proteolysis during roasting and in vitro digestion of fish (Acipenser gueldenstaedtii). J. Sci. Food Agric. 2018, 98, 5344–5351. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Kong, Q.; Sun, Z.T.; Liu, J.Y. Freshness analysis based on lipidomics for farmed Atlantic salmon (Salmo salar L.) stored at different times. Food Chem. 2022, 373, 131564. [Google Scholar] [CrossRef]
- Wang, L.; Zang, M.W.; Cheng, X.Y.; Wang, S.W.; Zhao, X.; Zhao, B.; Li, D. Evaluation of changes in the lipid profiles of dried shrimps (Penaeus vannamei) during accelerated storage based on chemical and lipidomics analysis. LWT—Food Sci. Technol. 2024, 191, 115564. [Google Scholar] [CrossRef]
- Guo, X.; Shi, D.; Liu, C.J.; Huang, Y.L.; Wang, Q.L.; Wang, J.Y.; Pei, L.Y.; Lu, S.L. UPLC-MS-MS-based lipidomics for the evaluation of changes in lipids during dry-cured mutton ham processing. Food Chem. 2022, 377, 131977. [Google Scholar] [CrossRef]
- Fu, Y.H.; Cao, S.Y.; Yang, L.; Li, Z.L. Flavor formation based on lipid in meat and meat products: A review. J. Food Biochem 2022, 46, e14439. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Sun, Z.X.; Qu, X.C.; Cao, J.; Shen, X.R.; Li, C. A comprehensive study of lipid profiles of round scad (Decapterus maruadsi) based on lipidomic with UPLCQ-Exactive Orbitrap-MS. Food Res. Int. 2020, 133, 109138. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.M.; Gang, K.Q.; Li, C.; Wang, J.H.; Liu, Y.X.; Zhou, D.Y.; Zhu, B.W. Change of lipids in whelks (Neptunea arthritica cumingi Crosse and Neverita didyma) during cold storage. Food Res. Int. 2020, 136, 109330. [Google Scholar] [CrossRef]
- Wang, X.S.; Zhang, H.W.; Song, Y.; Cong, P.X.; Li, Z.J.; Xu, J.; Xue, C.H. Comparative Lipid Profile Analysis of Four Fish Species by Ultraperformance Liquid Chromatography Coupled with Quadrupole Time-of-Flight Mass Spectrometry. J. Agric. Food Chem. 2019, 67, 9423–9431. [Google Scholar] [CrossRef]
- Cui, Z.; Liu, C.; RAO, W.X.; Chen, P.; Lei, K.K.; Mai, K.S.; Zhang, W.B. Dietary phospholipids improve growth performance and change the lipid composition and volatile flavor compound profiles in the muscle of abalone Haliotis discus hannai by affecting the glycerophospholipid metabolism. Aquac. Rep. 2023, 30, 101567. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lipid Subclass | Number of Compounds |
|---|---|
| Triglycerides, TG | 1182 |
| Phosphatidylcholine, PC | 430 |
| Phosphatidylethanolamine, PE | 409 |
| Phosphatidylinositol, PI | 330 |
| Phosphatidylglycerol, PG | 327 |
| Diglyceride, DG | 316 |
| Phosphatidylserine, PS | 224 |
| Cardiolipin, CL | 205 |
| Ceramide, Cer | 127 |
| Sulfatide, ST | 108 |
| Hexosylceramide, Hex1Cer | 97 |
| Sphingomyelin, SM | 75 |
| Dihexosylceramide, Hex2Cer | 74 |
| Lysophosphatidylcholine, LPC | 58 |
| Simple Glc series, Hex3Cer | 51 |
| N-acetylhexosyl ceramide, CerG2GNAc1 | 45 |
| Phytosphingosine, phSM | 44 |
| Lysophosphatidylethanolamine, LPE | 39 |
| Phosphatidylinositol phosphate, PIP | 29 |
| Phosphatidylinositol bisphosphate, PIP2 | 29 |
| Wax exters, WE | 26 |
| Aacyl carnitine, AcCa | 22 |
| Sphingosine bases, SPH | 22 |
| Monoglyceride, MG | 21 |
| Phosphatidic acid, PA | 20 |
| Ganglioside, monosialo trihexosyl ceramide, GM3 | 16 |
| Lysophosphatidylglycerol, LPG | 14 |
| Ceramides phosphate, CerP | 13 |
| Fatty acid, FA | 13 |
| Lysophosphatidylinositol, LPI | 10 |
| Lysophosphatidylserine, LPS | 9 |
| Zymosterol ester, ZyE | 8 |
| (O-acyl)-1-hydroxy fatty acid, OAHFA | 4 |
| Cholesteryl esters, ChE | 3 |
| Coenzyme, Co | 3 |
| Phosphatidylinositol triphosphate, PIP3 | 3 |
| Ganglioside, disialo tetrahexosyl ceramide, GD1a | 2 |
| Ganglioside, disialo dihexosyl ceramide, GD2 | 2 |
| monosialotetrahexosyl ganglioside, GM1 | 2 |
| Ganglioside, trisialo trihexosyl, GT3 | 2 |
| Sphin-gosine phosphate, SPHP | 2 |
| Dihexosyl N-acetylhexosyl ceramide, CerG3GNAc1 | 1 |
| Disialoganglioside, GD3 | 1 |
| Lysophosphatidic acid, LPA | 1 |
| Sitosteryl ester, SiE | 1 |
| Stigmasterol ester, StE | 1 |
| Number | Lipid lon | Class | Fold Change | p-Value | VIP |
|---|---|---|---|---|---|
| 1 | PE(18:1p_22:6)+H | PE | 0.499098 | 6.44 × 10−5 | 1.042086 |
| 2 | PC(39:6)+H | PC | 0.496688 | 8.43 × 10−5 | 1.38837 |
| 3 | TG(16:0_14:4_22:6)+NH4 | TG | 0.492149 | 2.84 × 10−6 | 1.881346 |
| 4 | TG(20:5_13:0_22:6)+NH4 | TG | 0.488128 | 3.24 × 10−6 | 3.105524 |
| 5 | TG(22:6_14:1_22:6)+NH4 | TG | 0.487257 | 0.000175 | 3.042199 |
| 6 | PE(16:0_22:6)−H | PE | 0.486444 | 1.94 × 10−6 | 2.852529 |
| 7 | PE(35:2e)+Na | PE | 0.48616 | 0.000594 | 1.108355 |
| 8 | SM(d20:0_24:5)+H | SM | 0.480833 | 0.00303 | 1.631861 |
| 9 | CL(78:11)−2H | CL | 0.480744 | 1.49 × 10−6 | 2.871019 |
| 10 | TG(22:6_14:3_22:6)+NH4 | TG | 0.47991 | 0.002727 | 1.572998 |
| 11 | PS(43:1e)+H | PS | 0.478902 | 3.56 × 10−5 | 2.906576 |
| 12 | PC(42:9)+H | PC | 0.47835 | 0.001059 | 1.089234 |
| 13 | TG(18:4_14:0_20:5)+NH4 | TG | 0.477786 | 0.001276 | 9.197348 |
| 14 | PC(22:3_14:1)+H | PC | 0.47614 | 1.19 × 10−5 | 1.162553 |
| 15 | TG(18:4_14:0_18:4)+NH4 | TG | 0.475079 | 4.68 × 10−5 | 6.977423 |
| 16 | PI(16:0_22:6)−H | PI | 0.469722 | 0.000323 | 1.639228 |
| 17 | PC(22:1_11:2)+Na | PC | 0.467923 | 1.05 × 10−5 | 4.199848 |
| 18 | PG(43:3e)+NH4 | PG | 0.466328 | 3.02 × 10−6 | 3.178235 |
| 19 | PE(10:0e_6:0)+H | PE | 0.463883 | 0.002008 | 1.500992 |
| 20 | ST(d17:1_25:0)+NH4 | ST | 0.457943 | 0.000698 | 1.70078 |
| 21 | PI(40:0e)+H | PI | 0.457439 | 0.000708 | 1.704323 |
| 22 | PC(17:1_22:6)+HCOO | PC | 0.455686 | 1.43 × 10−6 | 1.153681 |
| 23 | PE(16:0_18:2)+Na | PE | 0.454466 | 0.003023 | 1.162741 |
| 24 | TG(18:4_12:0_18:4)+NH4 | TG | 0.444249 | 0.001779 | 2.19355 |
| 25 | TG(18:4_18:0_22:6)+H | TG | 0.441096 | 4.30 × 10−6 | 1.808455 |
| 26 | PE(18:1p_20:5)+H | PE | 0.440797 | 2.07 × 10−5 | 1.528535 |
| 27 | TG(18:4_18:4_22:6)+H | TG | 0.439262 | 1.60 × 10−6 | 1.637022 |
| 28 | PG(32:2e)+NH4 | PG | 0.436992 | 1.91 × 10−5 | 1.147128 |
| 29 | TG(18:4_18:3_20:5)+NH4 | TG | 0.435734 | 0.000153 | 4.625362 |
| 30 | LPE(16:0)−H | LPE | 0.435246 | 0.002236 | 2.086989 |
| 31 | TG(22:6_12:2_22:6)+NH4 | TG | 0.431817 | 0.000265 | 2.296603 |
| 32 | PG(18:1e_23:1)+NH4 | PG | 0.427813 | 4.44 × 10−5 | 1.735106 |
| 33 | TG(18:4_14:1_18:4)+NH4 | TG | 0.425433 | 0.000918 | 1.645381 |
| 34 | PC(20:5_22:6)+HCOO | PC | 0.424829 | 0.000338 | 1.648933 |
| 35 | PC(44:11)+H | PC | 0.417744 | 0.000639 | 1.998421 |
| 36 | PC(22:6_22:6)+HCOO | PC | 0.414333 | 1.13 × 10−5 | 2.111974 |
| 37 | TG(18:4_13:0_22:6)+NH4 | TG | 0.411841 | 2.24 × 10−6 | 3.32173 |
| 38 | TG(18:4_18:4_20:5)+NH4 | TG | 0.411485 | 6.69 × 10−5 | 5.206043 |
| 39 | TG(18:4_18:3_18:4)+NH4 | TG | 0.411481 | 9.19 × 10−5 | 2.389019 |
| 40 | TG(18:4_13:0_20:5)+NH4 | TG | 0.408598 | 1.28 × 10−5 | 1.884269 |
| 41 | PE(18:2e_22:6)−H | PE | 0.4084 | 2.98 × 10−5 | 1.0287 |
| 42 | PE(18:1_22:6)−H | PE | 0.404044 | 0.005235 | 1.597052 |
| 43 | CL(82:13)−2H | CL | 0.403207 | 0.005028 | 1.601206 |
| 44 | TG(20:4_22:6_23:1)+NH4 | TG | 0.402977 | 0.001303 | 1.084609 |
| 45 | ST(m42:0 + O)+NH4 | ST | 0.401593 | 7.04 × 10−5 | 4.169806 |
| 46 | ST(d15:0_25:0)+NH4 | ST | 0.398087 | 0.001072 | 1.264458 |
| 47 | PI(33:1)+NH4 | PI | 0.397697 | 1.66 × 10−6 | 1.082869 |
| 48 | DG(21:5e)+NH4 | DG | 0.39523 | 0.000806 | 1.177416 |
| 49 | PG(31:2e)+NH4 | PG | 0.390305 | 3.11 × 10−6 | 1.162939 |
| 50 | TG(16:0_14:1_22:6)+H | TG | 0.389657 | 4.88 × 10−5 | 1.350519 |
| 51 | TG(18:4_12:0_20:5)+NH4 | TG | 0.387985 | 2.06 × 10−6 | 2.156707 |
| 52 | TG(22:4_12:4_22:4)+NH4 | TG | 0.3853 | 7.13 × 10−5 | 2.344739 |
| 53 | PC(20:5_22:6)+H | PC | 0.383516 | 0.00035 | 1.10243 |
| 54 | PC(17:1_22:6)+H | PC | 0.375831 | 0.002524 | 2.368831 |
| 55 | PC(37:5)+H | PC | 0.372954 | 3.74 × 10−5 | 2.477881 |
| 56 | TG(18:4_14:3_22:6)+NH4 | TG | 0.372685 | 0.000631 | 1.676399 |
| 57 | PI(42:1e)+H | PI | 0.364115 | 1.67 × 10−5 | 1.105667 |
| 58 | SPH(m18:1)+H | SPH | 0.363043 | 4.33 × 10−5 | 1.437811 |
| 59 | PE(16:0_20:5)−H | PE | 0.362863 | 0.001141 | 1.06783 |
| 60 | PE(39:2)+Na | PE | 0.359881 | 0.003976 | 1.106558 |
| 61 | PE(29:1_11:3)+Na | PE | 0.356409 | 0.00195 | 2.475733 |
| 62 | TG(18:4_18:4_18:4)+NH4 | TG | 0.350723 | 4.43 × 10−5 | 3.849431 |
| 63 | PE(20:1_22:6)−H | PE | 0.348642 | 0.000763 | 1.674806 |
| 64 | TG(18:4_14:2_20:5)+NH4 | TG | 0.348242 | 0.000135 | 1.611142 |
| 65 | TG(11:0_18:4_22:6)+NH4 | TG | 0.344356 | 2.29 × 10−5 | 1.024028 |
| 66 | PE(16:1e_22:6)−H | PE | 0.343037 | 1.16 × 10−5 | 1.600446 |
| 67 | TG(18:4_14:3_20:5)+NH4 | TG | 0.342678 | 7.13 × 10−5 | 1.382108 |
| 68 | TG(22:1_10:4_22:6)+H | TG | 0.339259 | 2.49 × 10−8 | 1.694093 |
| 69 | ST(d17:1_24:0)+NH4 | ST | 0.324952 | 5.26 × 10−6 | 1.074646 |
| 70 | TG(20:5_12:1_22:6)+H | TG | 0.324425 | 0.001746 | 1.569025 |
| 71 | SPH(d20:0)+H-H2O | SPH | 0.314818 | 1.68 × 10−5 | 1.334997 |
| 72 | ST(d48:3)+Na | ST | 0.306939 | 7.41 × 10−6 | 1.166141 |
| 73 | PE(18:2_22:6)+Na | PE | 0.305299 | 0.000763 | 1.272167 |
| 74 | TG(20:2_14:4_22:5)+NH4 | TG | 0.285599 | 0.003817 | 1.646126 |
| 75 | PE(22:6_22:6)−H | PE | 0.280681 | 6.49 × 10−6 | 2.91534 |
| 76 | PE(20:3_22:6)+Na | PE | 0.27609 | 3.58 × 10−5 | 3.615534 |
| 77 | PE(20:5_22:6)−H | PE | 0.271036 | 0.000203 | 1.163258 |
| 78 | TG(18:4_20:5_22:6)+H | TG | 0.255424 | 3.27 × 10−8 | 1.147957 |
| 79 | TG(20:5_14:2_22:6)+H | TG | 0.236534 | 6.83 × 10−7 | 1.309602 |
| 80 | TG(18:1_11:3_21:0)+Na | TG | 0.224134 | 5.49 × 10−7 | 1.208512 |
| 81 | TG(18:4_18:4_20:5)+H | TG | 0.202142 | 0.000627 | 2.141881 |
| 82 | TG(22:6_11:2_21:0)+H | TG | 0.201635 | 3.72 × 10−6 | 2.215741 |
| 83 | Hex2Cer(m39:3)+H-H2O | Hex2Cer | 0.199907 | 0.001191 | 1.061588 |
| 84 | SM(d20:0_22:0)+H | SM | 0.166036 | 0.002705 | 8.710905 |
| 85 | PI(25:1_11:2)+NH4 | PI | 0.071681 | 0.005298 | 7.23351 |
| Number | Lipidlon | Class | Fold Change | p-Value | VIP |
|---|---|---|---|---|---|
| 1 | TG(20:5_22:1_22:6)+NH4 | TG | 0.49851 | 5.68 × 10−7 | 5.215054 |
| 2 | TG(20:5_22:6_22:6)+NH4 | TG | 0.491599 | 0.000221 | 2.567459 |
| 3 | TG(18:1_20:5_22:6)+NH4 | TG | 0.490677 | 6.05 × 10−8 | 4.438805 |
| 4 | TG(20:0_22:3_23:0)+Na | TG | 0.489055 | 7.87 × 10−6 | 4.994488 |
| 5 | TG(24:1_22:1_24:1)+NH4 | TG | 0.486977 | 0.006783 | 1.564398 |
| 6 | TG(22:6_14:1_22:6)+NH4 | TG | 0.486156 | 0.000355 | 2.537809 |
| 7 | TG(18:4_16:0_20:5)+NH4 | TG | 0.482545 | 4.57 × 10−6 | 9.422562 |
| 8 | TG(18:4_18:4_20:5)+NH4 | TG | 0.475521 | 0.000612 | 3.973146 |
| 9 | TG(20:5_14:1_22:6)+NH4 | TG | 0.471436 | 8.49 × 10−6 | 1.658027 |
| 10 | TG(22:4_12:4_22:4)+NH4 | TG | 0.470116 | 0.000904 | 1.769338 |
| 11 | TG(26:1_22:1_22:6)+NH4 | TG | 0.467412 | 0.001272 | 1.034124 |
| 12 | TG(20:5_18:2_22:6)+NH4 | TG | 0.460452 | 1.21 × 10−5 | 3.218636 |
| 13 | TG(24:1_22:1_22:1)+NH4 | TG | 0.456162 | 0.000451 | 4.562797 |
| 14 | ST(m42:0 + O)+NH4 | ST | 0.451775 | 0.000622 | 3.284288 |
| 15 | TG(18:4_16:1_20:5)+NH4 | TG | 0.449565 | 3.97 × 10−5 | 4.416179 |
| 16 | TG(20:5_20:5_22:6)+NH4 | TG | 0.440813 | 2.87 × 10−6 | 5.177796 |
| 17 | TG(22:5_20:4_22:6)+NH4 | TG | 0.433843 | 0.000243 | 1.074286 |
| 18 | TG(22:1_22:1_22:1)+NH4 | TG | 0.432794 | 8.21 × 10−6 | 9.731679 |
| 19 | ST(d46:2)+NH4 | ST | 0.432073 | 8.33 × 10−6 | 5.279914 |
| 20 | SM(d18:2_24:1)+H | SM | 0.430173 | 4.26 × 10−5 | 1.308388 |
| 21 | TG(18:4_20:5_22:6)+NH4 | TG | 0.429418 | 1.56 × 10−5 | 6.177515 |
| 22 | TG(14:0_20:5_22:6)+NH4 | TG | 0.429232 | 1.12 × 10−5 | 2.991447 |
| 23 | TG(20:1_11:3_22:6)+NH4 | TG | 0.426299 | 3.61 × 10−6 | 1.616654 |
| 24 | TG(18:4_14:1_18:4)+NH4 | TG | 0.424032 | 0.000601 | 1.434173 |
| 25 | TG(18:4_18:3_22:6)+NH4 | TG | 0.41912 | 7.34 × 10−6 | 1.093412 |
| 26 | TG(16:0_14:1_22:6)+H | TG | 0.406657 | 9.14 × 10−5 | 1.130789 |
| 27 | TG(20:5_12:1_22:6)+H | TG | 0.395436 | 0.008554 | 1.019339 |
| 28 | TG(16:0_14:4_22:6)+NH4 | TG | 0.395024 | 4.52 × 10−7 | 1.802223 |
| 29 | TG(18:4_14:3_16:0)+NH4 | TG | 0.389088 | 4.19 × 10−5 | 1.988512 |
| 30 | PG(43:3e)+NH4 | PG | 0.380387 | 2.56 × 10−5 | 2.920494 |
| 31 | PI(37:0_9:0)+H | PI | 0.371176 | 0.008509 | 1.172954 |
| 32 | SM(d20:0_24:3)+H | SM | 0.36418 | 5.99 × 10−6 | 2.831523 |
| 33 | SM(d20:0_22:2)+H | SM | 0.35894 | 0.000375 | 2.3627 |
| 34 | TG(22:6_11:2_21:0)+H | TG | 0.355089 | 0.001193 | 1.531593 |
| 35 | TG(18:4_12:0_18:4)+NH4 | TG | 0.338135 | 0.000114 | 2.230751 |
| 36 | TG(16:1_14:0_20:5)+NH4 | TG | 0.331458 | 8.32 × 10−7 | 7.620491 |
| 37 | TG(12:0_14:0_22:6)+NH4 | TG | 0.328791 | 0.000737 | 3.230222 |
| 38 | TG(18:4_12:0_20:5)+NH4 | TG | 0.327852 | 7.66 × 10−7 | 1.988383 |
| 39 | TG(18:4_12:0_14:0)+NH4 | TG | 0.31954 | 0.000463 | 1.849902 |
| 40 | TG(18:4_14:0_20:5)+NH4 | TG | 0.318295 | 7.37 × 10−5 | 10.2416 |
| 41 | TG(18:4_14:3_20:5)+NH4 | TG | 0.311052 | 2.61 × 10−5 | 1.263194 |
| 42 | TG(18:4_14:2_20:5)+NH4 | TG | 0.270653 | 1.31 × 10−5 | 1.556192 |
| 43 | TG(28:1_22:3_22:6)+NH4 | TG | 0.269109 | 1.76 × 10−7 | 1.040046 |
| 44 | SM(d20:0_24:5)+H | SM | 0.259478 | 2.83 × 10−5 | 1.932346 |
| 45 | TG(18:4_13:0_22:6)+NH4 | TG | 0.237456 | 3.20 × 10−7 | 3.431831 |
| 46 | PI(42:1e)+H | PI | 0.233915 | 8.54 × 10−7 | 1.113729 |
| 47 | PS(43:1e)+H | PS | 0.232173 | 8.28 × 10−7 | 3.321953 |
| 48 | SPH(m18:1)+H | SPH | 0.195451 | 9.37 × 10−8 | 1.459295 |
| 49 | ST(d17:1_25:0)+NH4 | ST | 0.19223 | 5.03 × 10−7 | 2.081955 |
| 50 | PI(40:0e)+H | PI | 0.191481 | 5.31 × 10−7 | 2.086004 |
| 51 | ST(d15:0_25:0)+NH4 | ST | 0.18241 | 8.61 × 10−6 | 1.444491 |
| 52 | SPH(d20:0)+H-H2O | SPH | 0.181566 | 3.37 × 10−7 | 1.301676 |
| 53 | TG(22:6_12:2_22:6)+NH4 | TG | 0.180922 | 2.34 × 10−7 | 2.711327 |
| 54 | TG(18:4_14:3_22:6)+NH4 | TG | 0.176824 | 8.09 × 10−6 | 1.854093 |
| 55 | TG(22:6_14:3_22:6)+NH4 | TG | 0.134366 | 6.24 × 10−7 | 2.144745 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Wang, S.; Liu, Q.; Cao, R.; Zhang, Y.; Su, D.; Yu, Y. Storage Stability and Lipidomic Analysis Reveal the Effect of Frozen Storage Temperature on Pacific Saury (Cololabis saira). Foods 2025, 14, 756. https://doi.org/10.3390/foods14050756
Zhao L, Wang S, Liu Q, Cao R, Zhang Y, Su D, Yu Y. Storage Stability and Lipidomic Analysis Reveal the Effect of Frozen Storage Temperature on Pacific Saury (Cololabis saira). Foods. 2025; 14(5):756. https://doi.org/10.3390/foods14050756
Chicago/Turabian StyleZhao, Ling, Shanyu Wang, Qi Liu, Rong Cao, Yating Zhang, Dong Su, and Yueqin Yu. 2025. "Storage Stability and Lipidomic Analysis Reveal the Effect of Frozen Storage Temperature on Pacific Saury (Cololabis saira)" Foods 14, no. 5: 756. https://doi.org/10.3390/foods14050756
APA StyleZhao, L., Wang, S., Liu, Q., Cao, R., Zhang, Y., Su, D., & Yu, Y. (2025). Storage Stability and Lipidomic Analysis Reveal the Effect of Frozen Storage Temperature on Pacific Saury (Cololabis saira). Foods, 14(5), 756. https://doi.org/10.3390/foods14050756