

Aqueous Extract from Cuminum cyminum L. Seed Alleviates Ovalbumin-Induced Allergic Rhinitis in Mouse via Balancing of Helper T Cells


Abstract
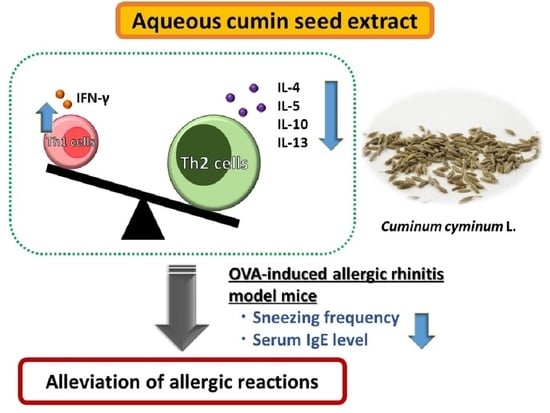
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Sample Preparation
2.3. Animals
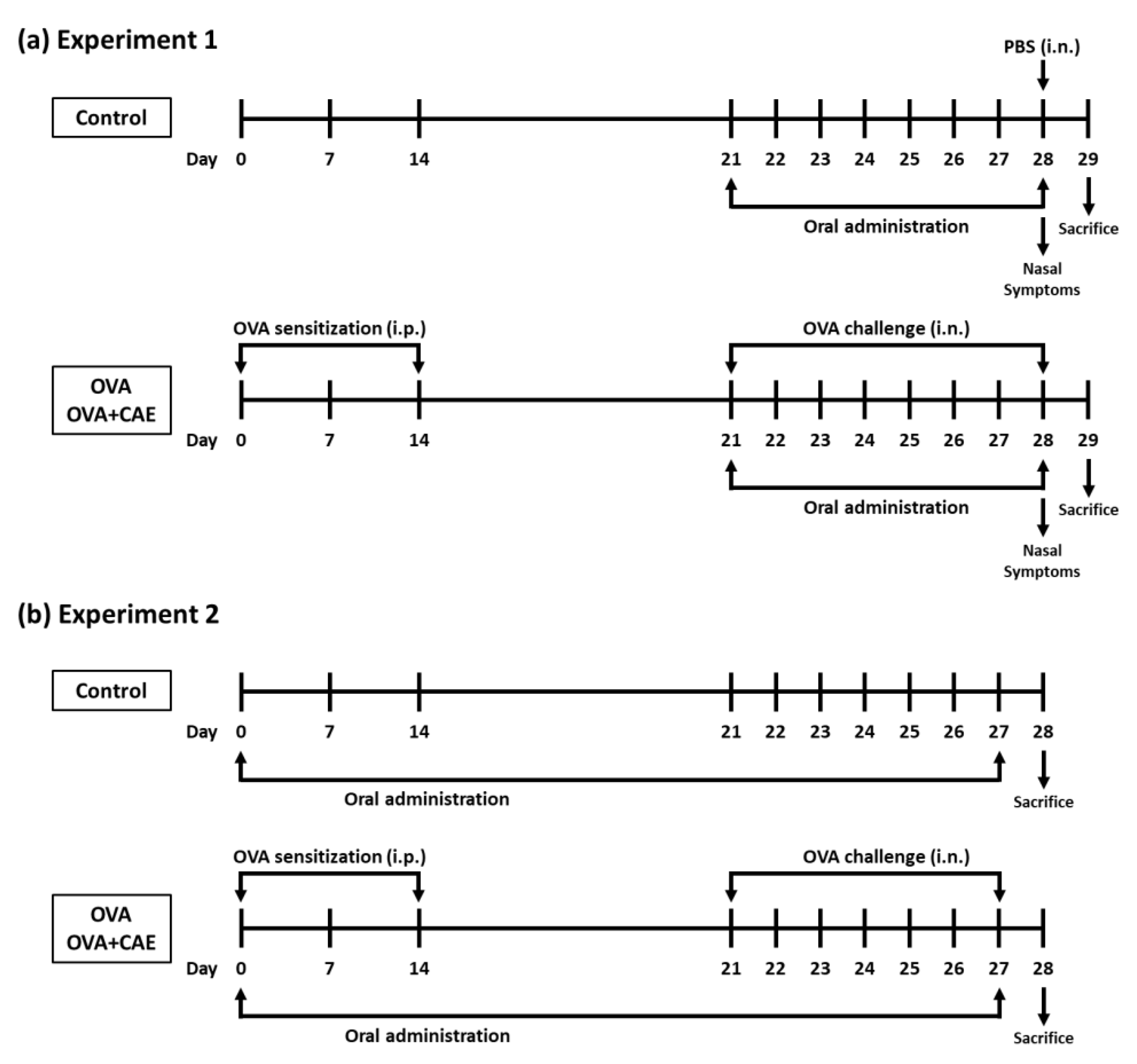
2.4. Mouse Model of OVA-Induced Allergic Rhinitis
2.5. Biochemical Measurement in the Serum of OVA-Induced Allergic Rhinitis Mice
2.6. Preparation of Splenocytes
2.7. Flow Cytometric Analysis
2.8. Statistical Analysis
3. Results
3.1. Effect of Oral Administration of CAE on Allergy Symptoms in OVA-Induced Allergic Rhinitis Mice
3.2. Effect of CAE on Serum Levels of Immunoglobulins and Cytokines of OVA-Induced Allergic Rhinitis Mice
3.3. Effect of CAE on Production of Immunoglobulins and Cytokines in Splenocytes of OVA-Induced Allergic Rhinitis Mice
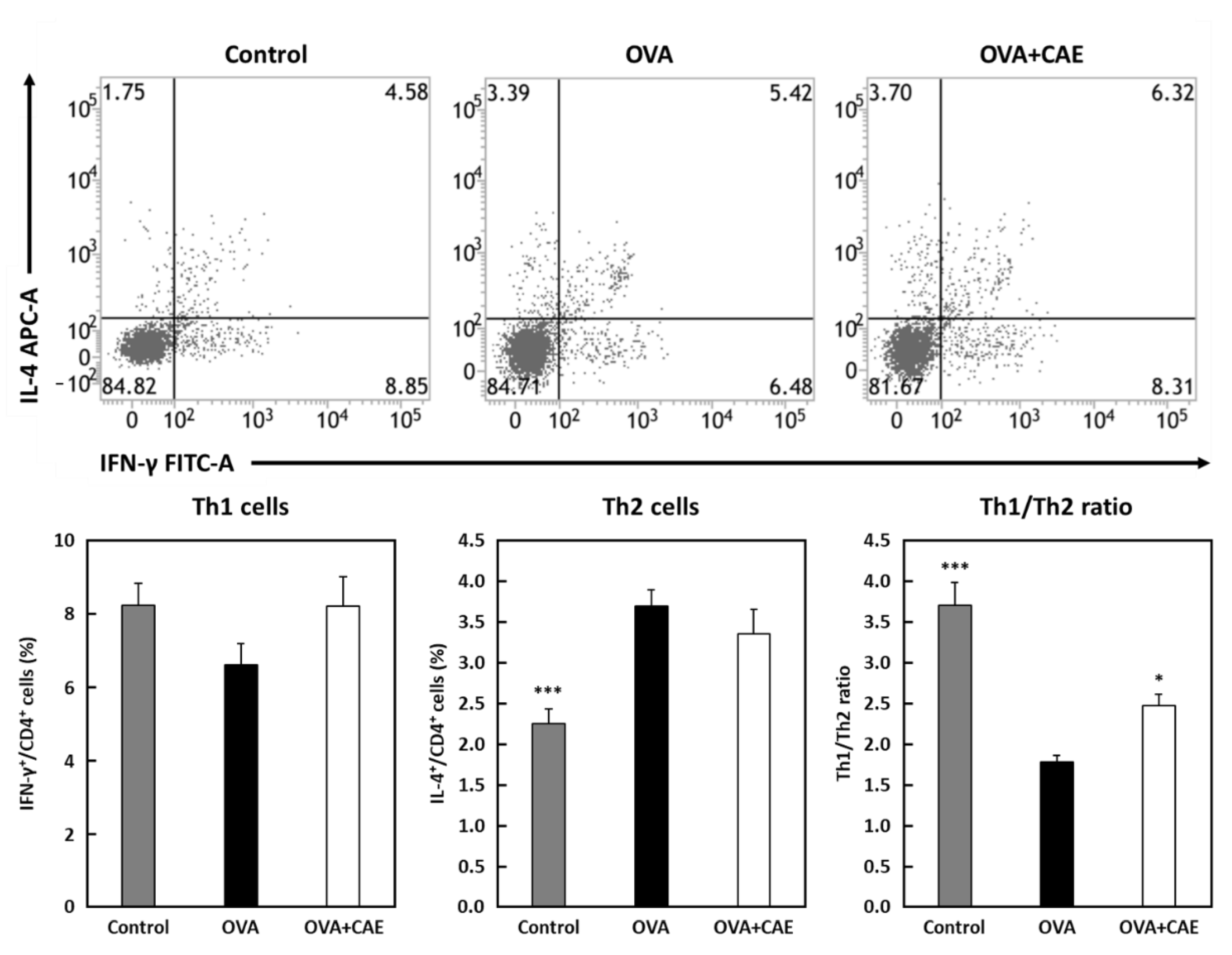
3.4. Effect of CAE on Th1/Th2 Balance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turner, H.; Kinet, J.-P. Signalling through the high-affinity IgE receptor FcεRI. Nature 1999, 402, 24–30. [Google Scholar] [CrossRef]
- Metcalfe, D.D.; Peavy, R.D.; Gilfillan, A.M. Mechanisms of mast cell signaling in anaphylaxis. J. Allergy Clin. Immunol. 2009, 124, 639–646. [Google Scholar] [CrossRef] [PubMed]
- White, M. Mediators of inflammation and the inflammatory process. J. Allergy Clin. Immunol. 1999, 103, S378–S381. [Google Scholar] [CrossRef]
- Ohno, T.; Morita, H.; Arae, K.; Matsumoto, K.; Nakae, S. Interleukin-33 in allergy. Allergy 2012, 67, 1203–1214. [Google Scholar] [CrossRef]
- Saluja, R.; Khan, M.; Church, M.K.; Maurer, M. The role of IL-33 and mast cells in allergy and inflammation. Clin. Transl. Allergy 2015, 5, 33. [Google Scholar] [CrossRef]
- Kubo, M. Innate and adaptive type 2 immunity in lung allergic inflammation. Immunol. Rev. 2017, 278, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Cayrol, C.; Girard, J.-P. Interleukin-33 (IL-33): A nuclear cytokine from the IL-1 family. Immunol. Rev. 2017, 281, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Murdaca, G.; Greco, M.; Tonacci, A.; Negrini, S.; Borro, M.; Puppo, F.; Gangemi, S. IL-33/IL-31 Axis in Immune-Mediated and Allergic Diseases. Int. J. Mol. Sci. 2019, 20, 5856. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Nie, Y.; Chong, L.; Cai, X.; Zhang, H.; Lin, B.; Liang, Y.; Li, C. PI3K and Notch signal pathways coordinately regulate the activation and proliferation of T lymphocytes in asthma. Life Sci. 2013, 92, 890–895. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.-L.; Li, F.-H.; Zhang, Y.; Zhang, X.-F.; Yang, J. Tangeretin has anti-asthmatic effects via regulating PI3K and Notch signaling and modulating Th1/Th2/Th17 cytokine balance in neonatal asthmatic mice. Braz. J. Med. Biol. Res. 2017, 50, e5991. [Google Scholar] [CrossRef]
- Huang, L.; Wang, M.; Yan, Y.; Gu, W.; Zhang, X.; Tan, J.; Sun, H.; Ji, W.; Chen, Z. OX40L induces helper T cell differentiation during cell immunity of asthma through PI3K/AKT and P38 MAPK signaling pathway. J. Transl. Med. 2018, 16, 74. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Athari, S.S.; Nasab, E.M.; Zhao, L. PI3K/AKT/mTOR and TLR4/MyD88/NF-κB signaling inhibitors attenuate pathological mechanisms of allergic asthma. Inflammation 2021, 44, 1895–1907. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Liu, Y.; Yu, J.; Dai, M.; Zhu, Y.; Bao, Y.; Peng, H.; Liu, K.; Zhu, X. Gene knockdown of CCR3 reduces eosinophilic inflammation and the Th2 immune response by inhibiting the PI3K/AKT pathway in allergic rhinitis mice. Sci. Rep. 2022, 12, 5411. [Google Scholar] [CrossRef] [PubMed]
- Korinek, M.; Handoussa, H.; Tsai, Y.-H.; Chen, Y.-Y.; Chen, M.-H.; Chiou, Z.-W.; Fang, Y.; Chang, F.-R.; Yen, C.-H.; Hsieh, C.-F.; et al. Anti-Inflammatory and Antimicrobial Volatile Oils: Fennel and Cumin Inhibit Neutrophilic Inflammation via Regulating Calcium and MAPKs. Front. Pharmacol. 2021, 12, 674095. [Google Scholar] [CrossRef]
- Allahghadri, T.; Rasooli, I.; Owlia, P.; Nadooshan, M.J.; Ghazanfari, T.; Taghizadeh, M.; Astaneh, S.D.A. Antimicrobial Property, Antioxidant Capacity, and Cytotoxicity of Essential Oil from Cumin Produced in Iran. J. Food Sci. 2010, 75, H54–H61. [Google Scholar] [CrossRef]
- Patil, S.B.; Takalikar, S.S.; Joglekar, M.M.; Haldavnekar, V.S.; Arvindekar, A.U. Insulinotropic and β-cell protective action of cuminaldehyde, cuminol and an inhibitor isolated from Cuminum cyminum in streptozotocin-induced diabetic rats. Br. J. Nutr. 2013, 110, 1434–1443. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, X.; Bi, Y.; Miao, R.; Zhang, Z.; Su, H. Anti-Inflammatory Effects of Cumin Essential Oil by Blocking JNK, ERK, and NF-κB Signaling Pathways in LPS-Stimulated RAW 264.7 Cells. Evid. Based Complement. Alternat. Med. 2015, 2015, 474509. [Google Scholar] [CrossRef]
- Haque, M.R.; Ansari, S.H. Aromatic aldehyde compound cuminaldehyde protects nonalcoholic fatty liver disease in rats feeding high fat diet. Hum. Exp. Toxicol. 2019, 38, 823–832. [Google Scholar] [CrossRef]
- Monteiro-Neto, V.; De Souza, C.D.; Gonzaga, L.F.; Da Silveira, B.C.; Sousa, N.C.F.; Pontes, J.P.; Santos, D.M.; Martins, W.C.; Pessoa, J.F.V.; Junior, A.R.C.; et al. Cuminaldehyde potentiates the antimicrobial actions of ciprofloxacin against Staphylococcus aureus and Escherichia coli. PLoS ONE 2020, 15, e0232987. [Google Scholar] [CrossRef]
- Hada, M.; Nishi, K.; Ishida, M.; Onda, H.; Nishimoto, S.; Sugahara, T. Inhibitory effect of aqueous extract of Cuminum cyminum L. seed on degranulation of RBL-2H3 cells and passive cutaneous anaphylaxis reaction in mice. Cytotechnology 2019, 71, 599–609. [Google Scholar] [CrossRef]
- Nakae, S.; Morita, H.; Ohno, T.; Arae, K.; Matsumoto, K.; Saito, H. Role of Interleukin-33 in Innate-Type Immune Cells in Allergy. Allergol. Int. 2013, 62, 13–20. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Matsushita, K. Innate-Type and Acquired-Type Allergy Regulated by IL-33. Allergol. Int. 2014, 63, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Mizusaki, A.; Nishi, K.; Nishiwaki, H.; Ishida, M.; Tamamoto, T.; Sugahara, T. Suppressive effect of ethanol extract from passion fruit seeds on IgE production. J. Funct. Foods 2017, 32, 176–184. [Google Scholar] [CrossRef]
- Romagnani, S. The role of lymphocytes in allergic disease. J. Allergy Clin. Immunol. 2000, 105, 399–408. [Google Scholar] [CrossRef]
- Nagase, H.; Ueki, S.; Fujieda, S. The roles of IL-5 and anti-IL-5 treatment in eosinophilic diseases: Asthma, eosinophilic granulomatosis with polyangiitis, and eosinophilic chronic rhinosinusitis. Allergol. Int. 2020, 69, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Mäkelä, M.J.; Kanehiro, A.; Borish, L.; Dakhama, A.; Loader, J.; Joetham, A.; Xing, Z.; Jordana, M.; Larsen, G.L.; Gelfand, E.W. IL-10 is necessary for the expression of airway hyperresponsiveness but not pulmonary inflammation after allergic sensitization. Proc. Natl. Acad. Sci. USA 2000, 97, 6007–6012. [Google Scholar] [CrossRef]
- Polukort, S.H.; Rovatti, J.; Carlson, L.; Thompson, C.; Ser-Dolansky, J.; Kinney, S.R.M.; Schneider, S.S.; Mathias, C.B. IL-10 enhances IgE-mediated mast cell responses and is essential for the development of experimental food allergy in IL-10-deficient mice. J. Immunol. 2016, 196, 4865–4876. [Google Scholar] [CrossRef] [PubMed]
- Trinchieri, G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat. Rev. Immunol. 2003, 3, 133–146. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Gu, W.; Sun, B. TH1/TH2 cell differentiation and molecular signals. Adv. Exp. Med. Biol. 2014, 841, 15–44. [Google Scholar] [PubMed]
- Ishida, M.; Ohara, R.; Miyagawa, F.; Kikuzaki, H.; Nishi, K.; Onda, H.; Yoshino, N.; Sugahara, T. Umbelliferose Isolated from Cuminum cyminum L. Seeds Inhibits Antigen-Induced Degranulation in Rat Basophilic Leukemia RBL-2H3 Cells. Molecules 2022, 27, 4101. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, B. Apigenin protects ovalbumin-induced asthma through the regulation of Th17 cells. Fitoterapia 2013, 91, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; He, D.; Yan, B. Apigenin attenuates allergic responses of ovalbumin-induced allergic rhinitis through modulation of Th1/Th2 responses in experimental mice. Dose Response 2020, 18, 1559325820904799. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wuniqiemu, T.; Tang, W.; Teng, F.; Bian, Q.; Yi, L.; Qin, J.; Zhu, X.; Wei, Y.; Dong, J. Luteolin inhibits autophagy in allergic asthma by activating PI3K/Akt/mTOR signaling and inhibiting Beclin-1-PI3KC3 complex. Int. Immunopharmacol. 2021, 94, 107460. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Xu, O.; Wang, J.; Shan, C.; Ren, X. Luteolin ameliorates inflammation and Th1/Th2 imbalance via regulating the TLR4/NF-κB pathway in allergic rhinitis rats. Immunopharmacol. Immunotoxicol. 2021, 43, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Sonoyama, K.; Watanabe, J.; Yamaguchi, N.; Kikuchi, H.; Nagura, T.; Aritsuka, T.; Fukumoto, K.; Kasai, T. Reduction of allergic airway eosinophilia by dietary raffinose in Brown Norway rats. Br. J. Nutr. 2004, 92, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Sonoyama, K.; Watanabe, H.; Watanabe, J.; Yamaguchi, N.; Yamashita, A.; Hashimoto, H.; Kishino, E.; Fujita, K.; Okada, M.; Mori, S.; et al. Allergic Airway Eosinophilia Is Suppressed in Ovalbumin-Sensitized Brown Norway Rats Fed Raffinose and α-Linked Galactooligosaccharide. J. Nutr. 2005, 135, 538–543. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Parameters | Control | OVA | OVA + CAE | OVA vs. Control p-Value | OVA vs. OVA + CAE p-Value |
|---|---|---|---|---|---|
| Mean ± SEM (n = 5) | |||||
| Body weight (g) on day 29 | 23.6 ± 0.3 | 21.1 ± 0.7 | 21.0 ± 0.5 | 0.010 | 0.995 |
| OVA challenge on day 28 | |||||
| Sneezing (number) | 6.8 ± 1.6 | 50.0 ± 10.8 | 27.0 ± 6.0 | 0.002 | 0.076 |
| Rubbing (number) | 21.6 ± 7.9 | 44.2 ± 5.2 | 38.8 ± 9.0 | 0.098 | 0.834 |
| Serum Levels | Control | OVA | OVA + CAE | OVA vs. Control p-Value | OVA vs. OVA + CAE p-Value |
|---|---|---|---|---|---|
| Mean ± SEM (n = 5) | |||||
| IgE (μg/mL) | 0.9 ± 0.3 | 41.9 ± 3.7 | 29.3 ± 1.4 | <0.001 | 0.004 |
| IgG1 (mg/mL) | 0.8 ± 0.2 | 5.9 ± 0.2 | 5.6 ± 0.3 | <0.001 | 0.438 |
| IL-4 (pg/mL) | 1.3 ± 0.2 | 9.5 ± 2.4 | 3.7 ± 0.6 | 0.003 | 0.023 |
| IFN-γ (pg/mL) | 769.5 ± 344.9 | 62.0 ± 37.3 | 716.1 ± 474.2 | 0.276 | 0.325 |
| Serum Levels | Control | OVA | OVA + CAE | OVA vs. Control p-Value | OVA vs. OVA + CAE p-Value |
|---|---|---|---|---|---|
| Mean ± SEM (n = 5) | |||||
| IgE (ng/mL) | 6.5 ± 2.1 | 37.7 ± 6.7 | 24.4 ± 6.9 | 0.004 | 0.207 |
| IgG1 (ng/mL) | 7.9 ± 4.5 | 131.1 ± 5.0 | 124.9 ± 10.3 | <0.001 | 0.767 |
| IL-4 (pg/mL) | 7.6 ± 0.6 | 875.6 ± 178.3 | 381.2 ± 121.1 | <0.001 | 0.029 |
| IL-5 (pg/mL) | 8.7 ± 0.3 | 744.9 ± 73.7 | 331.0 ± 96.0 | <0.001 | 0.002 |
| IL-10 (pg/mL) | 31.7 ± 4.0 | 829.9 ± 47.8 | 556.3 ± 118.9 | <0.001 | 0.041 |
| IL-13 (ng/mL) | 0.1 ± 0.0 | 3.3 ± 0.4 | 2.0 ± 0.6 | <0.001 | 0.066 |
| IFN-γ (ng/mL) | 1.7 ± 0.4 | 1.0 ± 0.2 | 1.5 ± 0.6 | 0.429 | 0.650 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishida, M.; Miyagawa, F.; Nishi, K.; Sugahara, T. Aqueous Extract from Cuminum cyminum L. Seed Alleviates Ovalbumin-Induced Allergic Rhinitis in Mouse via Balancing of Helper T Cells. Foods 2022, 11, 3224. https://doi.org/10.3390/foods11203224
Ishida M, Miyagawa F, Nishi K, Sugahara T. Aqueous Extract from Cuminum cyminum L. Seed Alleviates Ovalbumin-Induced Allergic Rhinitis in Mouse via Balancing of Helper T Cells. Foods. 2022; 11(20):3224. https://doi.org/10.3390/foods11203224
Chicago/Turabian StyleIshida, Momoko, Fuka Miyagawa, Kosuke Nishi, and Takuya Sugahara. 2022. "Aqueous Extract from Cuminum cyminum L. Seed Alleviates Ovalbumin-Induced Allergic Rhinitis in Mouse via Balancing of Helper T Cells" Foods 11, no. 20: 3224. https://doi.org/10.3390/foods11203224
APA StyleIshida, M., Miyagawa, F., Nishi, K., & Sugahara, T. (2022). Aqueous Extract from Cuminum cyminum L. Seed Alleviates Ovalbumin-Induced Allergic Rhinitis in Mouse via Balancing of Helper T Cells. Foods, 11(20), 3224. https://doi.org/10.3390/foods11203224