
Authenticity of Hay Milk vs. Milk from Maize or Grass Silage by Lipid Analysis

 ,
,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Milk Samples
2.2. Chemicals and Reagents
2.3. Sample Preparation and Analysis via GC-MS
2.3.1. Milk Fat Extraction
2.3.2. Transesterification
2.3.3. Analysis of the Cyclopropane Fatty Acid Dihydrosterculic Acid (DHSA)
2.4. Sample Preparation and Analysis via HPLC-HRMS
2.4.1. Milk Fat Extraction
2.4.2. High-Performance Liquid Chromatography Coupled to High Resolution Mass Spectrometry (HPLC-HRMS)
2.5. Data Processing and Statistical Analysis
3. Results and Discussion
3.1. GC-MS Analysis of DHSA
3.2. Milk Fat Analysis by HPLC-HRMS
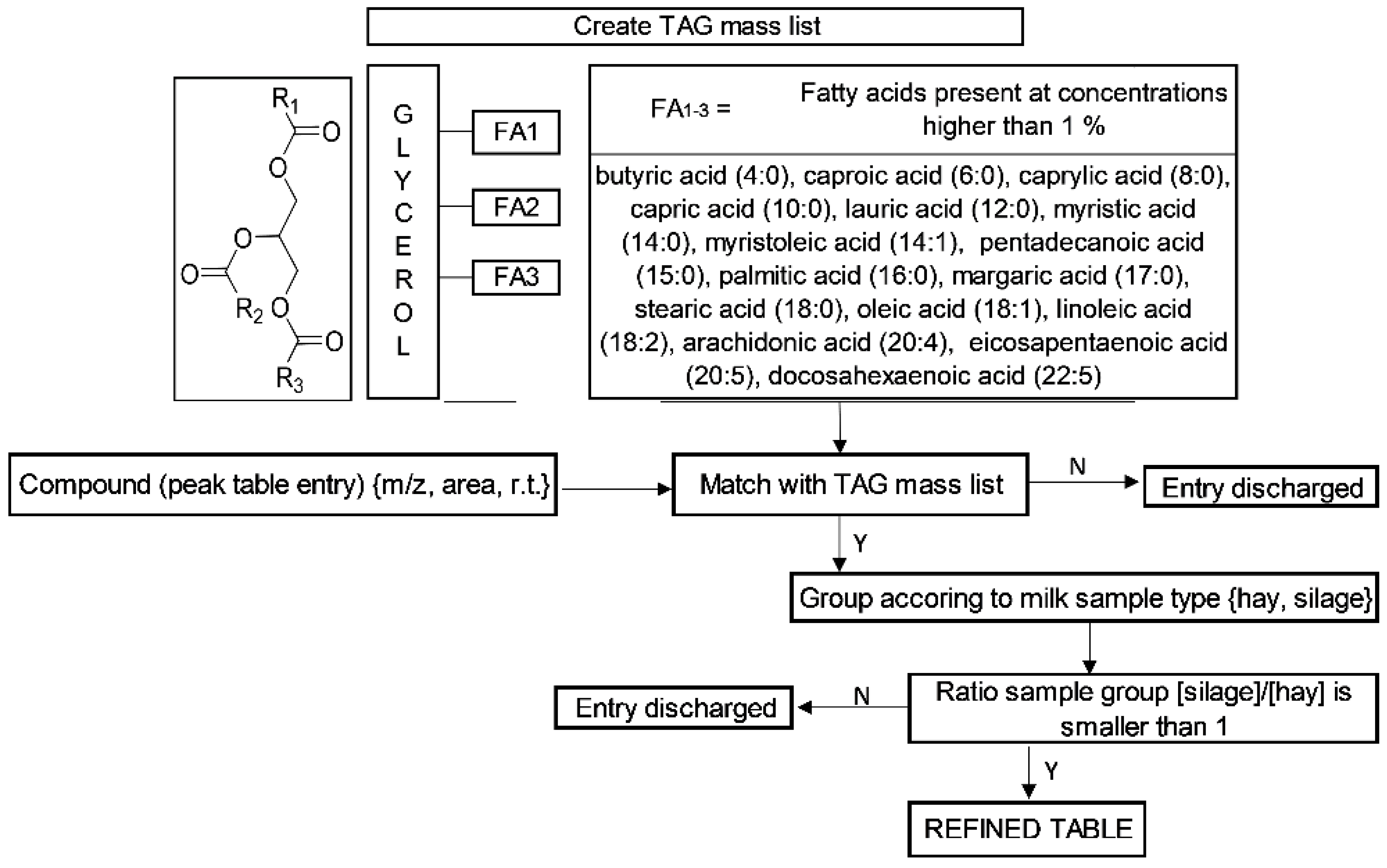
3.3. Tentative Identification of TAGs Marker
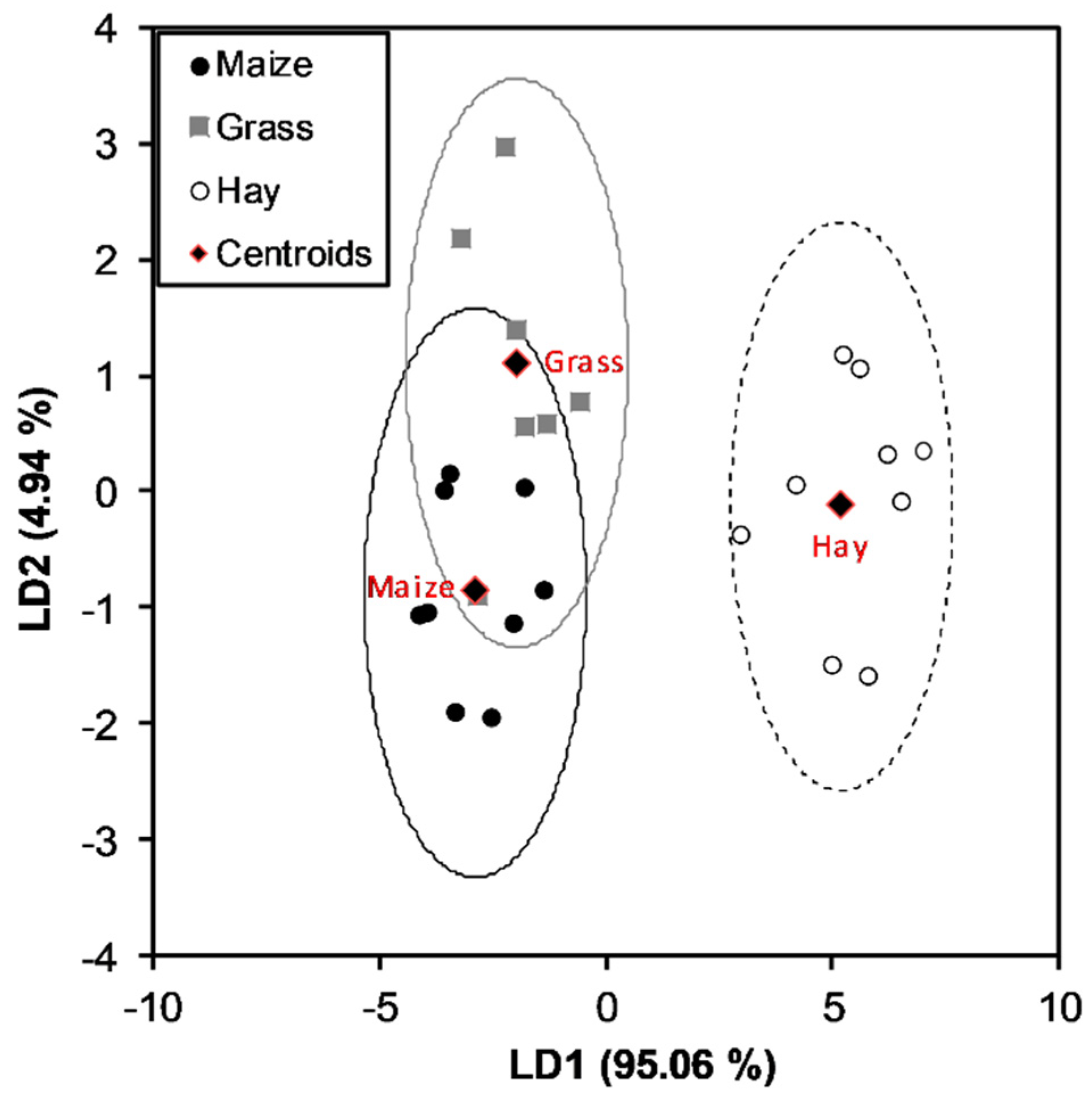
3.4. Discrimination of Milk Samples Using Multivariate Statistical Analysis
3.5. Comparison between DHSA and TAGs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacDonald, D.; Crabtree, J.; Wiesinger, G.; Dax, T.; Stamou, N.; Fleury, P.; Gutierrez Lazpita, J.; Gibon, A. Agricultural abandonment in mountain areas of Europe: Environmental consequences and policy response. J. Environ. Manag. 2000, 59, 47–69. [Google Scholar] [CrossRef]
- Busch, G.; Kühl, S.; Gauly, M. Consumer expectations regarding hay and pasture-raised milk in South Tyrol. Austrian J. Agric. Econ. Rural. Stud. 2018, 27, 79–86. [Google Scholar]
- Tosato, A. The Protection of Traditional Foods in the EU: Traditional Specialities Guaranteed. Eur. Law J. 2013, 19, 545–576. [Google Scholar] [CrossRef]
- Wölk, M.; Milkovska-Stamenova, S.; Schröter, T.; Hoffmann, R. Influence of seasonal variation and processing on protein glycation and oxidation in regular and hay milk. Food Chem. 2021, 337, 127690. [Google Scholar] [CrossRef]
- Palmieri, N.; Pesce, A.; Verrascina, M.; Perito, M.A. Market Opportunities for Hay Milk: Factors Influencing Perceptions among Italian Consumers. Animals 2021, 11, 431. [Google Scholar] [CrossRef] [PubMed]
- Commission Implementing Regulation (EU) 2016/304 of 2 March 2016. Available online: https://eur-lex.europa.eu/eli/reg_impl/2016/304/oj (accessed on 29 September 2021).
- Chilliard, Y.; Ferlay, A.; Doreau, M. Effect of different types of forages, animal fat or marine oils in cow’s diet on milk fat secretion and composition, especially conjugated linoleic acid (CLA) and polyunsaturated fatty acids. Livest. Prod. Sci. 2001, 70, 31–48. [Google Scholar] [CrossRef]
- Cabiddu, A.; Peratoner, G.; Valenti, B.; Monteils, V.; Martin, B.; Coppa, M. A quantitative review of on-farm feeding practices to enhance the quality of grassland-based ruminant dairy and meat products. Animal 2021, 100375. [Google Scholar] [CrossRef]
- Marseglia, A.; Caligiani, A.; Comino, L.; Righi, F.; Quarantelli, A.; Palla, G. Cyclopropyl and ω-cyclohexyl fatty acids as quality markers of cow milk and cheese. Food Chem. 2013, 140, 711–716. [Google Scholar] [CrossRef]
- Bergamaschi, M.; Cipolat-Gotet, C.; Cecchinato, A.; Schiavon, S.; Bittante, G. Chemometric authentication of farming systems of origin of food (milk and ripened cheese) using infrared spectra, fatty acid profiles, flavor fingerprints, and sensory descriptions. Food Chem. 2020, 305, 125480. [Google Scholar] [CrossRef] [PubMed]
- Caligiani, A.; Nocetti, M.; Lolli, V.; Marseglia, A.; Palla, G. Development of a Quantitative GC-MS Method for the Detection of Cyclopropane Fatty Acids in Cheese as New Molecular Markers for Parmigiano Reggiano Authentication. J. Agric. Food Chem. 2016, 64, 4158–4164. [Google Scholar] [CrossRef]
- Borreani, G.; Coppa, M.; Revello-Chion, A.; Comino, L.; Giaccone, D.; Ferlay, A.; Tabacco, E. Effect of different feeding strategies in intensive dairy farming systems on milk fatty acid profiles, and implications on feeding costs in Italy. J. Dairy Sci. 2013, 96, 6840–6855. [Google Scholar] [CrossRef] [Green Version]
- Butler, G.; Collomb, M.; Rehberger, B.; Sanderson, R.; Eyre, M.; Leifert, C. Conjugated linoleic acid isomer concentrations in milk from high- and low-input management dairy systems. J. Sci. Food Agric. 2009, 89, 697–705. [Google Scholar] [CrossRef]
- Collomb, M.; Bütikofer, U.; Sieber, R.; Jeangros, B.; Bosset, J.-O. Correlation between fatty acids in cows’ milk fat produced in the Lowlands, Mountains and Highlands of Switzerland and botanical composition of the fodder. Int. Dairy J. 2002, 12, 661–666. [Google Scholar] [CrossRef]
- Disciplinare e Normative del Consorzio-Parmigiano Reggiano. Available online: https://www.parmigianoreggiano.com/it/consorzio-disciplinare-normative/ (accessed on 10 June 2021).
- Chen, Y.Y.; Gänzle, M.G. Influence of cyclopropane fatty acids on heat, high pressure, acid and oxidative resistance in Escherichia coli. Int. J. Food Microbiol. 2016, 222, 16–22. [Google Scholar] [CrossRef]
- Russell, N.J.; Evans, R.I.; Steeg, P.F.; Hellemons, J.; Verheul, A.; Abee, T. Membranes as a target for stress adaptation. Int. J. Food Microbiol. 1995, 28, 255–261. [Google Scholar] [CrossRef]
- Montanari, C.; Sado Kamdem, S.L.; Serrazanetti, D.I.; Etoa, F.-X.; Guerzoni, M.E. Synthesis of cyclopropane fatty acids in Lactobacillus helveticus and Lactobacillus sanfranciscensis and their cellular fatty acids changes following short term acid and cold stresses. Food Microbiol. 2010, 27, 493–502. [Google Scholar] [CrossRef]
- Brown, J.L.; Ross, T.; McMeekin, T.A.; Nichols, P.D. Acid habituation of Escherichia coli and the potential role of cyclopropane fatty acids in low pH tolerance. Int. J. Food Microbiol. 1997, 37, 163–173. [Google Scholar] [CrossRef]
- Chang, Y.Y.; Cronan, J.E. Membrane cyclopropane fatty acid content is a major factor in acid resistance of Escherichia coli. Mol. Microbiol. 1999, 33, 249–259. [Google Scholar] [CrossRef]
- Woolford, M.K. The Silage Fermentation; Microbiological Series, No.14; Marcel Dekker: New York, NY, USA; Basle, Switzerland, 1984. [Google Scholar]
- Lima, E.; Abdalla, D. High-performance liquid chromatography of fatty acids in biological samples. Anal. Chim. Acta 2002, 465, 81–91. [Google Scholar] [CrossRef]
- Tzompa-Sosa, D.A.; Meurs, P.P.; van Valenberg, H.J.F. Triacylglycerol Profile of Summer and Winter Bovine Milk Fat and the Feasibility of Triacylglycerol Fragmentation. Eur. J. Lipid Sci. Technol. 2018, 120, 1700291. [Google Scholar] [CrossRef]
- Lopez, C.; Briard-Bion, V.; Menard, O.; Rousseau, F.; Pradel, P.; Besle, J.-M. Phospholipid, sphingolipid, and fatty acid compositions of the milk fat globule membrane are modified by diet. J. Agric. Food Chem. 2008, 56, 5226–5236. [Google Scholar] [CrossRef]
- Craige Trenerry, V.; Akbaridoust, G.; Plozza, T.; Rochfort, S.; Wales, W.J.; Auldist, M.; Ajlouni, S. Ultra-high-performance liquid chromatography–ion trap mass spectrometry characterisation of milk polar lipids from dairy cows fed different diets. Food Chem. 2013, 141, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Lock, A.L.; Garnsworthy, P.C. Technical note: A rapid lipid separation method for determining fatty acid composition of milk. J. Dairy Sci. 2004, 87, 3785–3788. [Google Scholar] [CrossRef] [Green Version]
- Christie, W.W. A simple procedure for rapid transmethylation of glycerolipids and cholesteryl esters. J. Lipid Res. 1982, 23, 1072–1075. [Google Scholar] [CrossRef]
- Breitkopf, S.B.; Ricoult, S.J.H.; Yuan, M.; Xu, Y.; Peake, D.A.; Manning, B.D.; Asara, J.M. A relative quantitative positive/negative ion switching method for untargeted lipidomics via high resolution LC-MS/MS from any biological source. Metabolomics 2017, 13, 30. [Google Scholar] [CrossRef] [Green Version]
- Matyash, V.; Liebisch, G.; Kurzchalia, T.V.; Shevchenko, A.; Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. J. Lipid Res. 2008, 49, 1137–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, R.G. Handbook of Milk Composition; Academic Press: San Diego, CA, USA, 1995; ISBN 0123844304. [Google Scholar]
- Jensen, R.G. The Composition of Bovine Milk Lipids: January 1995 to December 2000. J. Dairy Sci. 2002, 85, 295–350. [Google Scholar] [CrossRef]
- Bugaud, C.; Buchin, S.; Coulon, J.-B.; Hauwuy, A.; Dupont, D. Influence of the nature of alpine pastures on plasmin activity, fatty acid and volatile compound composition of milk. Lait 2001, 81, 401–414. [Google Scholar] [CrossRef]
- Decaen, C.; Adda, J. Changes in secretion of fatty acids in milk fat of cows during lactation. Ann. Biol. Anim. Biochim. Biophys. 1970, 10, 659–677. [Google Scholar] [CrossRef]
- Bartsch, B.D.; Graham, E.R.; McLean, D.M. Protein and fat composition and some manufacturing properties of milk from dairy cows fed on hay and concentrate in various ratios. Aust. J. Agric. Res. 1979, 30, 191. [Google Scholar] [CrossRef]
- Doreau, M.; Chilliard, Y. Effects of ruminal or postruminal fish oil supplementation on intake and digestion in dairy cows. Reprod. Nutr. Dev. 1997, 37, 113–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shabala, L.; Ross, T. Cyclopropane fatty acids improve Escherichia coli survival in acidified minimal media by reducing membrane permeability to H+ and enhanced ability to extrude H+. Res. Microbiol. 2008, 159, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Cossignani, L.; Pollini, L.; Blasi, F. Invited review: Authentication of milk by direct and indirect analysis of triacylglycerol molecular species. J. Dairy Sci. 2019, 102, 5871–5882. [Google Scholar] [CrossRef] [PubMed]
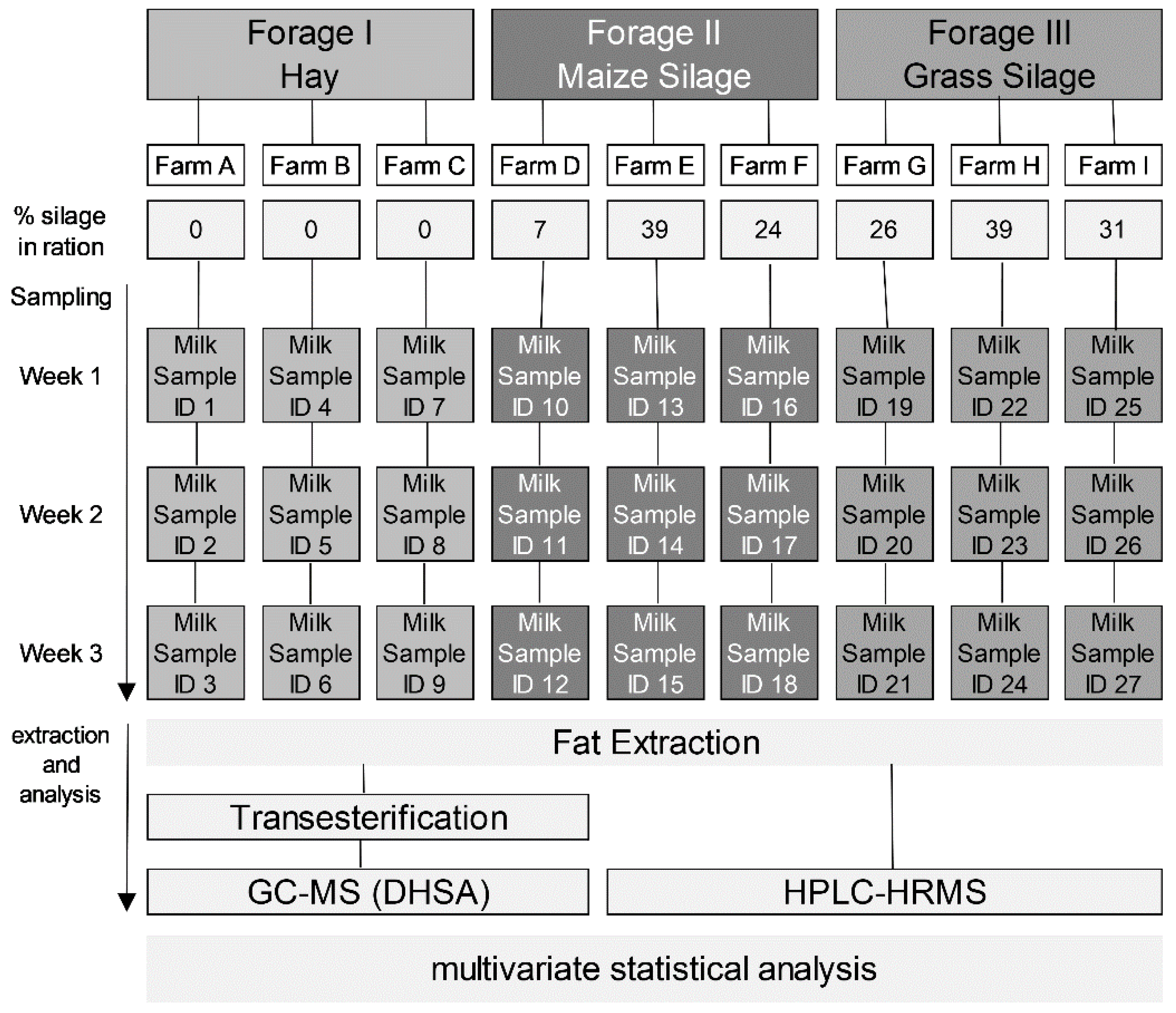
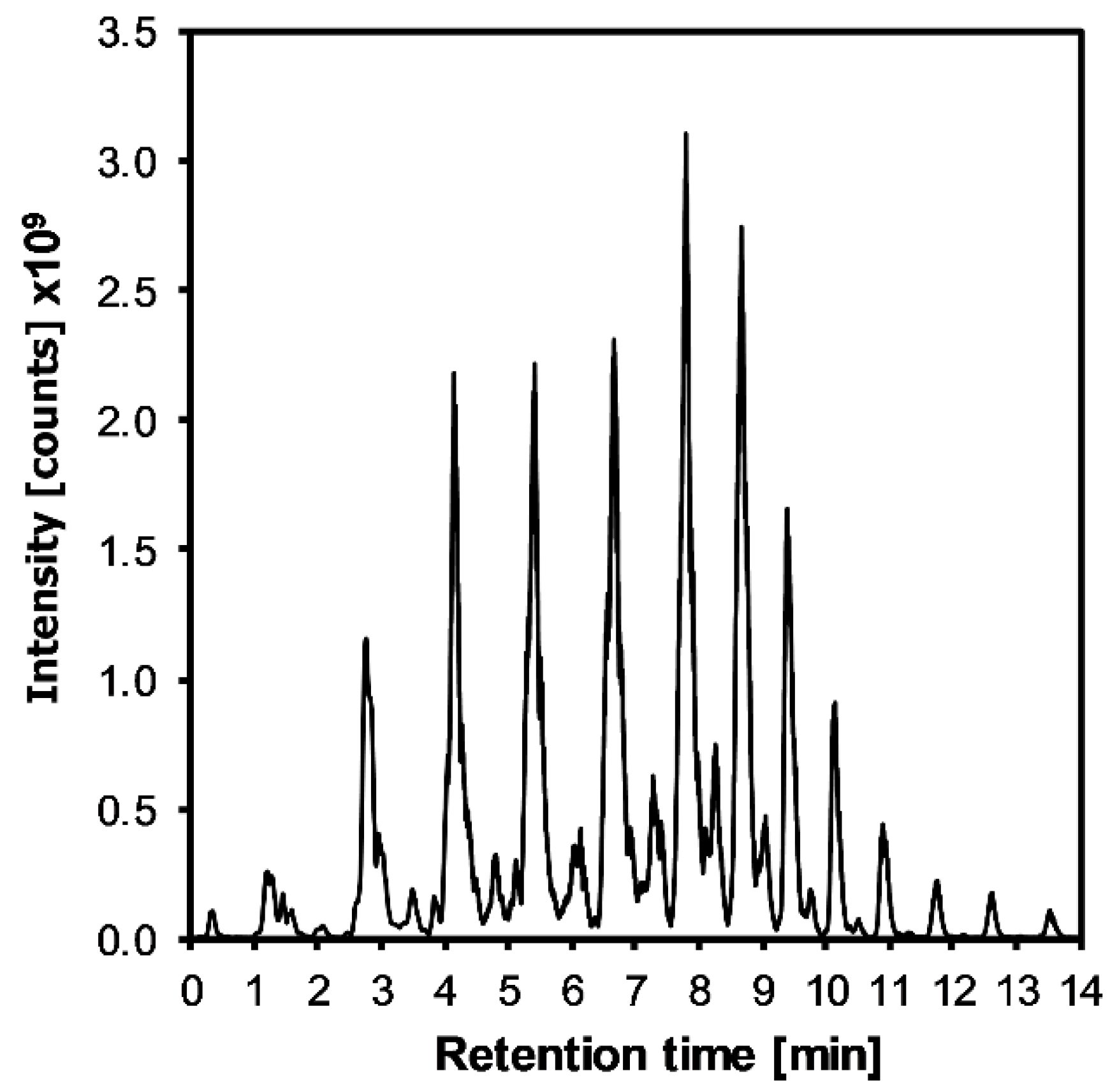
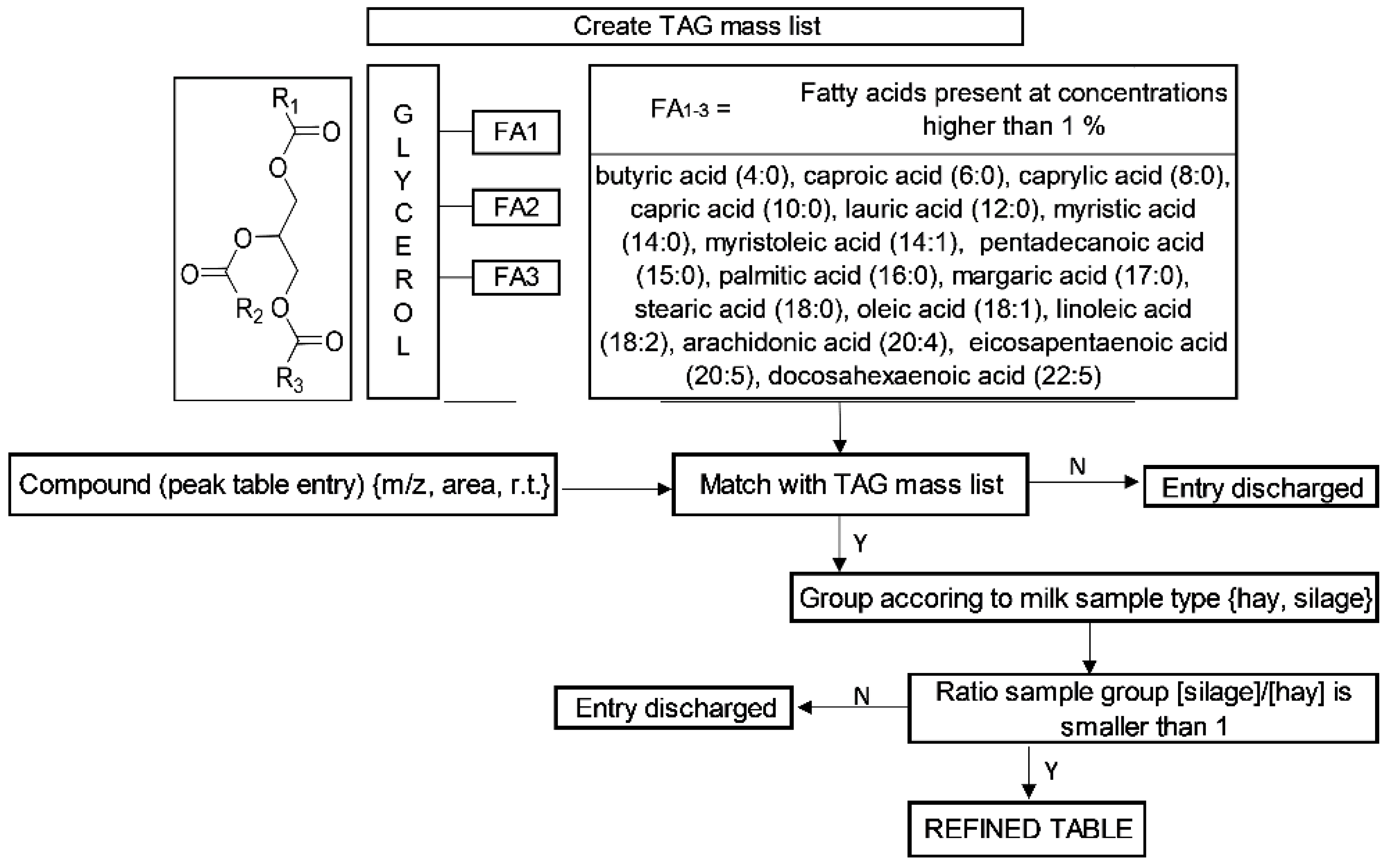
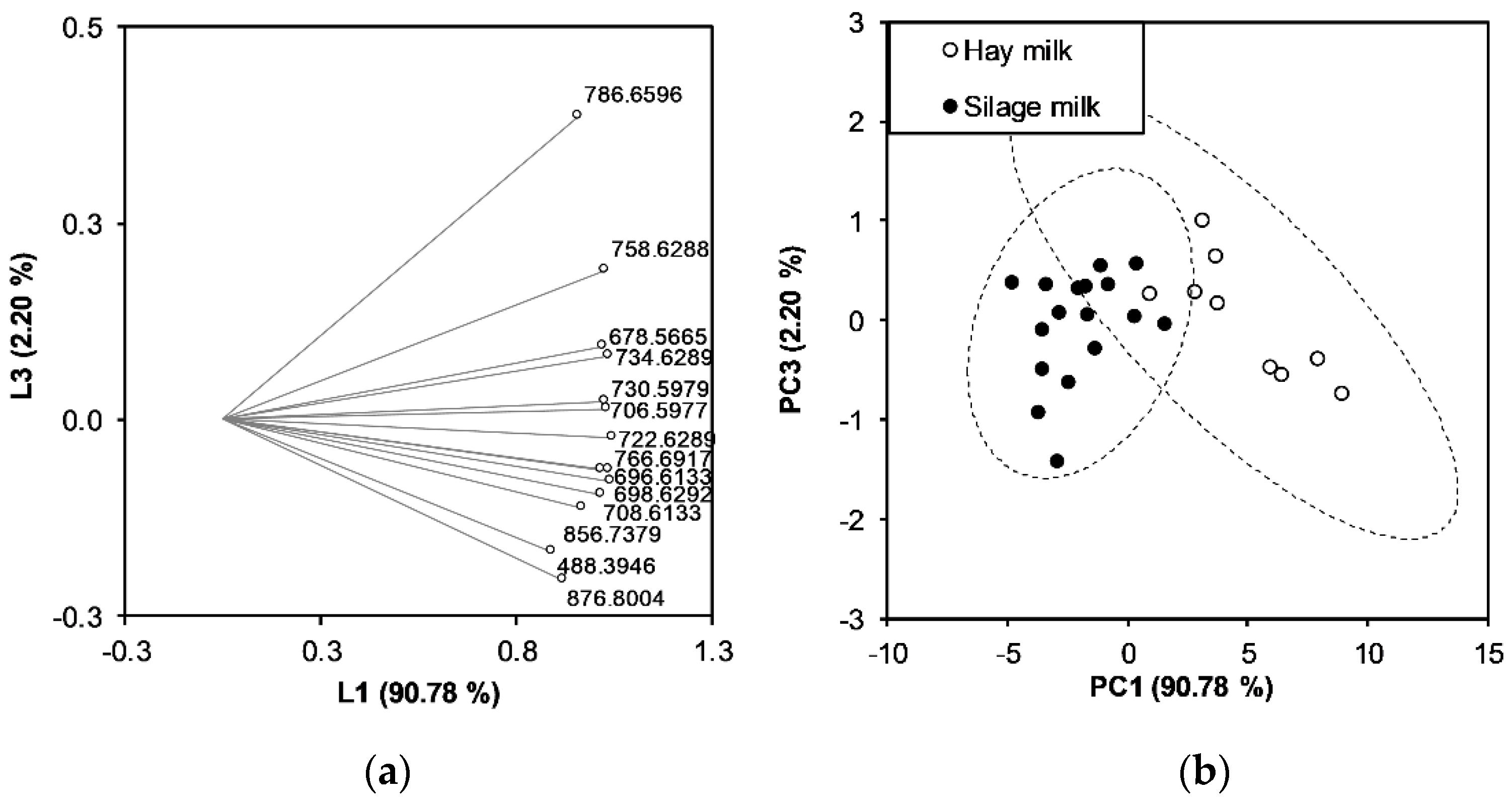

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Milk Sample | Farm | DHSA (mg/kg Fat) | RSD (%) |
|---|---|---|---|
| HM | A | <LOD | |
| B | <LOD | ||
| C | <LOD | ||
| SM-M | D | 94 ± 13 | 13 |
| E | 94 ± 68 | 72 | |
| F | 52 ± 10 | 19 | |
| SM-G | G | <LOQ | |
| H | <LOD | ||
| I | 30 ± 11 | 28 |
| m/z [M+NH4]+ | Predicted Formula | Classif. (CN:DB *) | m/z Fragments | Tentatively Identified Fatty Acid Moieties |
|---|---|---|---|---|
| 488.3946 | C27H50O6 | TG 24:0 | 355.2843, 327.253, 299.2217, 155.143, 127.1117, 109.1012, 99.0804, 81.0699 | butyric (4:0); caproic (6:0); caprylic (8:0) |
| 678.5665 | C41H73O6 | TG 38:3 | 573.4869, 405.3011, 383.3157, 261.2213, 239.2355, 71.04924 | butyric (4:0); palmitic (16:0); linolenic (18:3) |
| 696.6133 | C42H78O6 | TG 39:1 | 591.5349, 437.3627, 409.3312, 409.3258, 397.3312, 397.3220, 265.2524, 99.0804, 71.0492 | butyric (4:0); caproic (6:0); pentadecanoic (15:0); margaric (17:0); oleic (18:1) |
| 698.6292 | C42H80O6 | TG 39:0 | 593.5503, 565.5195, 509.4567, 453.3939, 439.3783, 425.3625, 411.3469, 397.3313, 313.2728, 267.2680, 253.2525, 239.2368, 235.2419, 225.2211, 221.2262, 211.2056, 207.2107, 193.1951, 173.1171, 155.1431, 137.1325, 127.1117, 99.0804, 81.0699, 71.0855, 53.0025 | butyric (4:0); caproic (6:0); caprylic (8:0), capric (10:0); pentadecanoic (15:0); margaric (17:0); stearic (18:0) |
| 706.5977 | C43H76O6 | TG 40:3 | 409.3316,407.3159, 99.0805, 411.3470, 601.5199, 265.2528, 145.0860, 263.2371, 261.2213, 405.3002, 433.3314, 119.0857, 127.1118, 573.4892, 247.2422, 245.2266, 243.2115, 239.2368, 173.1324, 313.2731, 339.2894, 155.1433, 53.0026, 99.1169, 71.0492 | butyric (4:0); caproic (6:0); palmitic (16:0); stearic (18:0); oleic (18:1); linoleic (18:2); linolenic (18:3) |
| 708.6133 | C43H78O6 | TG 40:2 | 603.5757, 409.3601, 339.2887, 265.2522, 247.2463, 145.1018, 71.04978 | butyric (4:0); oleic (18:1) |
| 722.6289 | C44H80O6 | TG 41:2 | 589.5209, 435.3455, 425.3624 | caproic (6:0); margaric (17:0); linoleic (18:2); |
| 730.5979 | C45H76O6 | TG42:5 | 625.5178, 457.3311, 383.3153, 313.2517, 239.2368, 145.1012, 71.0492 | docosapentaenoic (22:5); palmitic (16:0); butyric (4:0) |
| 734.6289 | C45H80O6 | TG 42:3 | 717.6027, 601.5206, 435.3476, 437.3621, 265.2532, 263.2367, 99.0805, 81.0699 | caproic (6:0); oleic (18:1); linoleic (18:2) |
| 758.6288 | C47H80O6 | TG 44:5 | 411.3466, 285.0094, 239.0951, 201.1638, 109.1014 | caprylic (8:0); capric (10:0); lauric (12:0); myristic (14:0); docosapentaenoic (22:5); |
| 766.6917 | C47H88O6 | TG 44:1 | 605.5507, 577.5189, 549.4877, 523.4722, 521.4564, 493.4251, 467.4093, 465.3938, 109.1011, 95.0854, 85.1011 81.0698, 71.0855, 57.0701 | caprylic (8:0); capric (10:0); lauric (12:0); myristoleic (14:1); myristic (14:0); palmitic (16:0); oleic (18:1); stearic (18:0) |
| 786.6596 | C49H84O6 | TG 46:5 | 569.456, 313.278256, 467.4245, 239.2007, 211.2057 | linoleic (12:0); myristic (14:0); docosapentaenoic (22:5); eicosapentaenoic (20:5) |
| 856.7379 | C54H94O6 | TG 51:5 | 639.5344, 571.4151, 537.4889, 507.1079, 373,7151 239.2007, 299.2578, 191.1788 | lauric (12:0); myristoleic (14:1); pentadecenoic (15:1); myristic (17:0); myristoleic (17:1); docosapentaenoic (22:5); docosatetraenoic (22:4); docosatrienoic (22:3) |
| 876.8004 | C55H102O6 | TG 52:2 | 221.2262, 239.2367, 245.2261, 263.2366, 267.2684, 313.2734, 575.5031, 579.5311, 603.5344 | linoleic (18:2); stearic (18:0); palmitic (16:0) |
| Class | Hay | Silage | Sub-Class | Grass | Maize | Total | |
|---|---|---|---|---|---|---|---|
| Fitting | Hay | 9 (100%) | 0 (%) | 0 (%) | 0 (%) | 9 | |
| Silage | 0 (%) | 18 (100%) | Grass | 8 (89%) | 1 (11%) | 18 | |
| Maize | 1 (11%) | 8 (89%) | |||||
| Cross validation, leave one out | Hay | 9 (100%) | 0 (%) | 0 (%) | 0 (%) | 9 | |
| Silage | 0 (%) | 18 (100%) | Grass | 8 (89%) | 1 (11%) | 18 | |
| Maize | 1 (11%) | 8 (89%) | |||||
| DHSA present | Hay | 9 (100%) | 0 (%) | 0 (%) | 0 (%) | 9 | |
| Silage | 4 (22%) | 14 (78%) | Grass | 5 (56%) | - | 18 | |
| Maize | - | 9 (100%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imperiale, S.; Kaneppele, E.; Morozova, K.; Fava, F.; Martini-Lösch, D.; Robatscher, P.; Peratoner, G.; Venir, E.; Eisenstecken, D.; Scampicchio, M. Authenticity of Hay Milk vs. Milk from Maize or Grass Silage by Lipid Analysis. Foods 2021, 10, 2926. https://doi.org/10.3390/foods10122926
Imperiale S, Kaneppele E, Morozova K, Fava F, Martini-Lösch D, Robatscher P, Peratoner G, Venir E, Eisenstecken D, Scampicchio M. Authenticity of Hay Milk vs. Milk from Maize or Grass Silage by Lipid Analysis. Foods. 2021; 10(12):2926. https://doi.org/10.3390/foods10122926
Chicago/Turabian StyleImperiale, Sebastian, Elke Kaneppele, Ksenia Morozova, Federico Fava, Demian Martini-Lösch, Peter Robatscher, Giovanni Peratoner, Elena Venir, Daniela Eisenstecken, and Matteo Scampicchio. 2021. "Authenticity of Hay Milk vs. Milk from Maize or Grass Silage by Lipid Analysis" Foods 10, no. 12: 2926. https://doi.org/10.3390/foods10122926
APA StyleImperiale, S., Kaneppele, E., Morozova, K., Fava, F., Martini-Lösch, D., Robatscher, P., Peratoner, G., Venir, E., Eisenstecken, D., & Scampicchio, M. (2021). Authenticity of Hay Milk vs. Milk from Maize or Grass Silage by Lipid Analysis. Foods, 10(12), 2926. https://doi.org/10.3390/foods10122926