
Development of a Biotechnological Strategy for CO2 Capture in Coke Plant Emissions Using Microalgae

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain
2.2. Initial Biomass Production
2.3. Effect of Photoperiod and Flue Gas Concentration on Biomass Production
2.4. Optimization of Photoperiod and Flue Gas Concentration
2.5. Biomass Quantification
- is the dry biomass per unit volume of the culture medium (g/L).
- is the final weight of the filter with the dry biomass (g).
- is the initial weight of the filter before filtration (g).
- is the volume of the aliquot filtered (L).
2.6. Biomass Analysis
2.6.1. Total Carbohydrates
2.6.2. Total Proteins
2.6.3. Total Lipids
2.6.4. Total Organic Carbon (TOC) Analysis
2.7. Field Verification
2.8. Biomass Production and Metabolite Concentration
- Mc represents the percentage of the metabolite (proteins, carbohydrates, or lipids) in the biomass (%);
- Ms is the mass of the specific metabolite in the sample (g);
- Bs is the total dry biomass in the sample (g).
3. Results
3.1. Effect of Photoperiod and CO2 Concentration on Biomass Production and CO2 Capture
3.1.1. Statistical Analysis
3.1.2. Visualization of Interactions Using Response Surfaces
3.1.3. Quadratic Model for Biomass Prediction
3.2. Process Optimization
Biomass Production and Biochemical Composition of Metabolites
4. Discussion
4.1. ANOVA Analysis
4.2. Response Surface Analysis
4.3. Quadratic Model
4.4. Optimal Conditions
4.5. Biomass Production and Biochemical Composition
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shindell, D.; Smith, C.J. Climate and Air-Quality Benefits of a Realistic Phase-out of Fossil Fuels. Nature 2019, 573, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Pendrill, F.; Persson, U.M.; Godar, J.; Kastner, T.; Moran, D.; Schmidt, S.; Wood, R. Agricultural and Forestry Trade Drives Large Share of Tropical Deforestation Emissions. Glob. Environ. Change 2019, 56, 1–10. [Google Scholar] [CrossRef]
- Verma, P.; Kumar Ghosh, P. REDD+ Strategy for Forest Carbon Sequestration in India. Holist. Approach Environ. 2022, 12, 117–130. [Google Scholar] [CrossRef]
- Gizer, S.G.; Polat, O.; Ram, M.K.; Sahiner, N. Recent Developments in CO2 Capture, Utilization, Related Materials, and Challenges. Int. J. Energy Res. 2022, 46, 16241–16263. [Google Scholar] [CrossRef]
- Regufe, M.J.; Pereira, A.; Ferreira, A.F.P.; Ribeiro, A.M.; Rodrigues, A.E. Current Developments of Carbon Capture Storage and/or Utilization–Looking for Net-Zero Emissions Defined in the Paris Agreement. Energies 2021, 14, 2406. [Google Scholar] [CrossRef]
- Seo, H.; Rahimi, M.; Hatton, T.A. Electrochemical Carbon Dioxide Capture and Release with a Redox-Active Amine. J. Am. Chem. Soc. 2022, 144, 2164–2170. [Google Scholar] [CrossRef]
- Sgouridis, S.; Carbajales-Dale, M.; Csala, D.; Chiesa, M.; Bardi, U. Comparative Net Energy Analysis of Renewable Electricity and Carbon Capture and Storage. Nat. Energy 2019, 4, 456–465. [Google Scholar] [CrossRef]
- Zhang, X.; Yuan, H.; Guan, L.; Wang, X.; Wang, Y.; Jiang, Z.; Cao, L.; Zhang, X. Influence of Photoperiods on Microalgae Biofilm: Photosynthetic Performance, Biomass Yield, and Cellular Composition. Energies 2019, 12, 3724. [Google Scholar] [CrossRef]
- Rangel-Basto, Y.A.; García-Ochoa, I.E.; Suarez-Gelvez, J.H.; Zuorro, A.; Barajas-Solano, A.F.; Urbina-Suarez, N.A. The Effect of Temperature and Enzyme Concentration in the Transesterification Process of Synthetic Microalgae Oil. Chem. Eng. Trans. 2018, 64, 331–336. [Google Scholar] [CrossRef]
- Ji, M.-K.; Abou-Shanab, R.A.I.; Kim, S.-H.; Salama, E.-S.; Lee, S.-H.; Kabra, A.N.; Lee, Y.-S.; Hong, S.; Jeon, B.-H. Cultivation of Microalgae Species in Tertiary Municipal Wastewater Supplemented with CO2 for Nutrient Removal and Biomass Production. Ecol. Eng. 2013, 58, 142–148. [Google Scholar] [CrossRef]
- Senatore, V.; Buonerba, A.; Zarra, T.; Oliva, G.; Belgiorno, V.; Boguniewicz-Zablocka, J.; Naddeo, V. Innovative Membrane Photobioreactor for Sustainable CO2 Capture and Utilization. Chemosphere 2021, 273, 129682. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Huang, Y.; Feng, J.; Sun, J.; Zhou, J.; Cen, K. Mutate Chlorella sp. by Nuclear Irradiation to Fix High Concentrations of CO2. Bioresour. Technol. 2013, 136, 496–501. [Google Scholar] [CrossRef]
- Schultze, L.K.P.; Simon, M.-V.; Li, T.; Langenbach, D.; Podola, B.; Melkonian, M. High Light and Carbon Dioxide Optimize Surface Productivity in a Twin-Layer Biofilm Photobioreactor. Algal Res. 2015, 8, 37–44. [Google Scholar] [CrossRef]
- Tan, K.M.; Kassim, M.A. Growth, Carbohydrate Productivity And Growth Kinetic Study of Halochlorella Rubescens Cultivated Under CO2-Rich Conditions. Malays. Appl. Biol. 2020, 49, 1–11. [Google Scholar] [CrossRef]
- Razzak, S.A.; Ali, S.A.M.; Hossain, M.M.; Mouanda, A.N. Biological CO2 Fixation Using Chlorella Vulgaris and Its Thermal Characteristics through Thermogravimetric Analysis. Bioprocess. Biosyst. Eng. 2016, 39, 1651–1658. [Google Scholar] [CrossRef]
- Kuo, C.-M.; Jian, J.-F.; Lin, T.-H.; Chang, Y.-B.; Wan, X.-H.; Lai, J.-T.; Chang, J.-S.; Lin, C.-S. Simultaneous Microalgal Biomass Production and CO2 Fixation by Cultivating Chlorella sp. GD with Aquaculture Wastewater and Boiler Flue Gas. Bioresour. Technol. 2016, 221, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Shakoor, A.; Ashraf, F.; Shakoor, S.; Mustafa, A.; Rehman, A.; Altaf, M.M. Biogeochemical Transformation of Greenhouse Gas Emissions from Terrestrial to Atmospheric Environment and Potential Feedback to Climate Forcing. Environ. Sci. Pollut. Res. 2020, 27, 38513–38536. [Google Scholar] [CrossRef]
- Borella, L.; Sforza, E.; Bertucco, A. Effect of Residence Time in Continuous Photobioreactor on Mass and Energy Balance of Microalgal Protein Production. New Biotechnol. 2021, 64, 46–53. [Google Scholar] [CrossRef]
- Sterman, J.; Moomaw, W.; Rooney-Varga, J.N.; Siegel, L. Does Wood Bioenergy Help or Harm the Climate? Bull. At. Sci. 2022, 78, 128–138. [Google Scholar] [CrossRef]
- García-Martínez, J.B.; Ayala-Torres, E.; Reyes-Gómez, O.; Zuorro, A.; Andrés, F.; Barajas-Solano, B.; Crisóstomo, C.; Barajas-Ferreira, B. Evaluation of a Two-Phase Extraction System of Carbohydrates and Proteins from Chlorella Vulgaris Utex 1803. Chem. Eng. Trans. 2016, 49, 355–360. [Google Scholar] [CrossRef]
- Choi, H.I.; Hwang, S.-W.; Sim, S.J. Comprehensive Approach to Improving Life-Cycle CO2 Reduction Efficiency of Microalgal Biorefineries: A Review. Bioresour. Technol. 2019, 291, 121879. [Google Scholar] [CrossRef]
- Barajas-Solano, A.F.; Gonzalez-Delgado, A.D.; Kafarov, V. Effect of Thermal Pre-Treatment on Fermentable Sugar Production of Chlorella Vulgaris. Chem. Eng. Trans. 2014, 37, 655–660. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The Promising Future of Microalgae: Current Status, Challenges, and Optimization of a Sustainable and Renewable Industry for Biofuels, Feed, and Other Products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef] [PubMed]
- Urbina-Suarez, N.A.; Ayala-González, D.D.; Rivera-Amaya, J.D.; Barajas-Solano, A.F.; Machuca-Martínez, F. Evaluation of the Light/Dark Cycle and Concentration of Tannery Wastewater in the Production of Biomass and Metabolites of Industrial Interest from Microalgae and Cyanobacteria. Water 2022, 14, 346. [Google Scholar] [CrossRef]
- Vuppaladadiyam, A.K.; Yao, J.G.; Florin, N.; George, A.; Wang, X.; Labeeuw, L.; Jiang, Y.; Davis, R.W.; Abbas, A.; Ralph, P.; et al. Impact of Flue Gas Compounds on Microalgae and Mechanisms for Carbon Assimilation and Utilization. ChemSusChem 2018, 11, 334–355. [Google Scholar] [CrossRef]
- Alami, A.H.; Alasad, S.; Ali, M.; Alshamsi, M. Investigating Algae for CO2 Capture and Accumulation and Simultaneous Production of Biomass for Biodiesel Production. Sci. Total Environ. 2021, 759, 143529. [Google Scholar] [CrossRef]
- Wang, H.; Nche-Fambo, F.A.; Yu, Z.; Chen, F. Using Microalgal Communities for High CO2-Tolerant Strain Selection. Algal Res. 2018, 35, 253–261. [Google Scholar] [CrossRef]
- Suresh, A.; Benor, S. Microalgae-Based Biomass Production for Control of Air Pollutants. In From Biofiltration to Promising Options in Gaseous Fluxes Biotreatment; Elsevier: Amsterdam, The Netherlands, 2020; pp. 345–372. [Google Scholar]
- Ma, S.; Li, D.; Yu, Y.; Li, D.; Yadav, R.S.; Feng, Y. Application of a Microalga, Scenedesmus obliquus PF3, for the Biological Removal of Nitric Oxide (NO) and Carbon Dioxide. Environ. Pollut. 2019, 252, 344–351. [Google Scholar] [CrossRef]
- Cheng, D.; Li, X.; Yuan, Y.; Yang, C.; Tang, T.; Zhao, Q.; Sun, Y. Adaptive Evolution and Carbon Dioxide Fixation of Chlorella sp. in Simulated Flue Gas. Sci. Total Environ. 2019, 650, 2931–2938. [Google Scholar] [CrossRef]
- Kassim, M.A.; Ramli, S.H.; Meng, T.K. Analysis of Microalgal Growth Kinetic Model and Carbohydrate Biosynthesis Cultivated Using Agro-Industrial Waste Residuals as Carbon Source. Prep. Biochem. Biotechnol. 2022, 52, 514–524. [Google Scholar] [CrossRef]
- Talib, R.M.; Dharma, A.; Syafrizayanti, K.F. The Impact of Gas from Coal Combustion toward the Degree of Acidity (pH), Biomass Productivity, and Chlorophyll A and B Content of Chlorella Emersonii. Int. J. Curr. Microbiol. Appl. Sci. 2022, 11, 120–127. [Google Scholar] [CrossRef]
- Kumar, P.K.; Krishna, S.V.; Verma, K.; Pooja, K.; Bhagawan, D.; Himabindu, V. Phycoremediation of Sewage Wastewater and Industrial Flue Gases for Biomass Generation from Microalgae. S. Afr. J. Chem. Eng. 2018, 25, 133–146. [Google Scholar] [CrossRef]
- Cheng, J.; Zhu, Y.; Zhang, Z.; Yang, W. Modification and Improvement of Microalgae Strains for Strengthening CO2 Fixation from Coal-Fired Flue Gas in Power Plants. Bioresour. Technol. 2019, 291, 121850. [Google Scholar] [CrossRef]
- Andersen, R.; Berges, J.; Harrison, P.; Watanabe, M. Recipes for Freshwater and Seawater Media. In Algal Culture Techniques; Academic Press: London, UK, 2005. [Google Scholar]
- Al-Mallahi, J.; Ishii, K.; Sato, M.; Ochiai, S. Static Supply of Different Simulated Flue Gases for Native Microalgae Cultivation in Diluted Cow Manure Digestate. J. Environ. Manag. 2023, 335, 117557. [Google Scholar] [CrossRef] [PubMed]
- Moheimani, N.R.; Borowitzka, M.A.; Isdepsky, A.; Sing, S.F. Standard Methods for Measuring Growth of Algae and Their Composition. In Algae for Biofuels and Energy; Springer: Dordrecht, The Netherlands, 2013; pp. 265–284. ISBN 978-94-007-5479-9. [Google Scholar]
- Slocombe, S.P.; Ross, M.; Thomas, N.; McNeill, S.; Stanley, M.S. A Rapid and General Method for Measurement of Protein in Micro-Algal Biomass. Bioresour. Technol. 2013, 129, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Suh, W.I.; Farooq, W.; Moon, M.; Shrivastav, A.; Park, M.S.; Yang, J.W. Rapid Quantification of Microalgal Lipids in Aqueous Medium by a Simple Colorimetric Method. Bioresour. Technol. 2014, 155, 330–333. [Google Scholar] [CrossRef]
- Liu, X.; Chen, G.; Tao, Y.; Wang, J. Application of Effluent from WWTP in Cultivation of Four Microalgae for Nutrients Removal and Lipid Production under the Supply of CO2. Renew. Energy 2020, 149, 708–715. [Google Scholar] [CrossRef]
- Aghaalipour, E.; Akbulut, A.; Güllü, G. Carbon Dioxide Capture with Microalgae Species in Continuous Gas-Supplied Closed Cultivation Systems. Biochem. Eng. J. 2020, 163, 107741. [Google Scholar] [CrossRef]
- Kong, W.; Kong, J.; Ma, J.; Lyu, H.; Feng, S.; Wang, Z.; Yuan, P.; Shen, B. Chlorella Vulgaris Cultivation in Simulated Wastewater for the Biomass Production, Nutrients Removal and CO2 Fixation Simultaneously. J. Environ. Manag. 2021, 284, 112070. [Google Scholar] [CrossRef]
- Hariz, H.B.; Takriff, M.S.; Mohd Yasin, N.H.; Ba-Abbad, M.M.; Mohd Hakimi, N.I.N. Potential of the Microalgae-Based Integrated Wastewater Treatment and CO2 Fixation System to Treat Palm Oil Mill Effluent (POME) by Indigenous Microalgae; Scenedesmus sp. and Chlorella sp. J. Water Process Eng. 2019, 32, 100907. [Google Scholar] [CrossRef]
- Rodas-Zuluaga, L.I.; Castañeda-Hernández, L.; Castillo-Vacas, E.I.; Gradiz-Menjivar, A.; López-Pacheco, I.Y.; Castillo-Zacarías, C.; Boully, L.; Iqbal, H.M.N.; Parra-Saldívar, R. Bio-Capture and Influence of CO2 on the Growth Rate and Biomass Composition of the Microalgae Botryococcus braunii and Scenedesmus sp. J. CO2 Util. 2021, 43, 101371. [Google Scholar] [CrossRef]
- Ramos-Ibarra, J.R.; Snell-Castro, R.; Neria-Casillas, J.A.; Choix, F.J. Biotechnological Potential of Chlorella sp. and Scenedesmus sp. Microalgae to Endure High CO2 and Methane Concentrations from Biogas. Bioprocess. Biosyst. Eng. 2019, 42, 1603–1610. [Google Scholar] [CrossRef]
- Bhola, V.; Swalaha, F.M.; Nasr, M.; Bux, F. Fuzzy Intelligence for Investigating the Correlation between Growth Performance and Metabolic Yields of a Chlorella sp. Exposed to Various Flue Gas Schemes. Bioresour. Technol. 2017, 243, 1078–1086. [Google Scholar] [CrossRef]
- Hussain, J.; Wang, X.; Sousa, L.; Ali, R.; Rittmann, B.E.; Liao, W. Using Non-Metric Multi-Dimensional Scaling Analysis and Multi-Objective Optimization to Evaluate Green Algae for Production of Proteins, Carbohydrates, Lipids, and Simultaneously Fix Carbon Dioxide. Biomass Bioenergy 2020, 141, 105711. [Google Scholar] [CrossRef]
- Quintero-Dallos, V.; García-Martínez, J.B.; Contreras-Ropero, J.E.; Barajas-Solano, A.F.; Barajas-Ferrerira, C.; Lavecchia, R.; Zuorro, A. Vinasse as a Sustainable Medium for the Production of Chlorella Vulgaris UTEX 1803. Water 2019, 11, 1526. [Google Scholar] [CrossRef]
- Ghosh, A.; Khanra, S.; Mondal, M.; Devi, T.I.; Halder, G.; Tiwari, O.N.; Bhowmick, T.K.; Gayen, K. Biochemical Characterization of Microalgae Collected from North East Region of India Advancing towards the Algae-based Commercial Production. Asia-Pac. J. Chem. Eng. 2017, 12, 745–754. [Google Scholar] [CrossRef]
- Vishwakarma, R.; Dhar, D.W.; Jena, M.; Shukla, M. Biochemical Parameters and 18S RRNA Gene Sequence Analysis amongst Green Microalgal Strains from Selected Aquatic Sites of Eastern India. Water Sci. Technol. 2020, 82, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Nordin, N.; Yusof, N.; Samsudin, S. Biomass Production of Chlorella sp., Scenedesmus sp., and Oscillatoria sp. in Nitrified Landfill Leachate. Waste Biomass Valorization 2017, 8, 2301–2311. [Google Scholar] [CrossRef]
- Gumbi, S.T.; Mutanda, T.; Olaniran, A.O. Nutrient Removal from Dairy and Poultry Wastewater with Simultaneous Biomass and Biodiesel Production by Chlorella sp. T4 Isolated from a Freshwater Stream in South Africa. Waste Biomass Valorization 2021, 12, 6931–6943. [Google Scholar] [CrossRef]
- Heffernan, J.K.; Valgepea, K.; de Souza Pinto Lemgruber, R.; Casini, I.; Plan, M.; Tappel, R.; Simpson, S.D.; Köpke, M.; Nielsen, L.K.; Marcellin, E. Enhancing CO2-Valorization Using Clostridium Autoethanogenum for Sustainable Fuel and Chemicals Production. Front. Bioeng. Biotechnol. 2020, 8, 204. [Google Scholar] [CrossRef]
- Deprá, M.C.; Dias, R.R.; Severo, I.A.; de Menezes, C.R.; Zepka, L.Q.; Jacob-Lopes, E. Carbon Dioxide Capture and Use in Photobioreactors: The Role of the Carbon Dioxide Loads in the Carbon Footprint. Bioresour. Technol. 2020, 314, 123745. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Samadhiya, K.; Kiran, B. Multi-Objective Tailored Optimization Deciphering Carbon Partitioning and Metabolomic Tuning in Response to Elevated CO2 Levels, Organic Carbon and Sparging Period. Environ. Res. 2022, 204, 112137. [Google Scholar] [CrossRef]
- Madhubalaji, C.K.; Chandra, T.S.; Chauhan, V.S.; Sarada, R.; Mudliar, S.N. Chlorella Vulgaris Cultivation in Airlift Photobioreactor with Transparent Draft Tube: Effect of Hydrodynamics, Light and Carbon Dioxide on Biochemical Profile Particularly ω-6/ω-3 Fatty Acid Ratio. J. Food Sci. Technol. 2020, 57, 866–876. [Google Scholar] [CrossRef] [PubMed]
- Ziganshina, E.E.; Bulynina, S.S.; Yureva, K.A.; Ziganshin, A.M. Optimization of Photoautotrophic Growth Regimens of Scenedesmaceae Alga: The Influence of Light Conditions and Carbon Dioxide Concentrations. Appl. Sci. 2023, 13, 12753. [Google Scholar] [CrossRef]
- Wang, B.; Xiong, W.; Yu, J.; Maness, P.-C.; Meldrum, D.R. Unlocking the Photobiological Conversion of CO2 to (R)-3-Hydroxybutyrate in Cyanobacteria. Green. Chem. 2018, 20, 3772–3782. [Google Scholar] [CrossRef]
- Gao, E.-B.; Wu, J.; Ye, P.; Qiu, H.; Chen, H.; Fang, Z. Rewiring Carbon Flow in Synechocystis PCC 6803 for a High Rate of CO2-to-Ethanol under an Atmospheric Environment. Front. Microbiol. 2023, 14, 1211004. [Google Scholar] [CrossRef]
- Choi, J.; Ko, J.; Ng, C.T.; Jeong, S.; Tenhunen, J.; Xue, W.; Cho, J. Quantification of CO2 Fluxes in Paddy Rice Based on the Characterization and Simulation of CO2 Assimilation Approaches. Agric. Meteorol. 2018, 249, 348–366. [Google Scholar] [CrossRef]
- Huang, Y.; Rorrer, G.L. Optimal Temperature and Photoperiod for the Cultivation of Agardhiella subulata Microplantlets in a Bubble-Column Photobioreactor. Biotechnol. Bioeng. 2002, 79, 135–144. [Google Scholar] [CrossRef]
- Mohammadi, A.; Abedini, H. Experimental Study and Numerical Modeling of CO2 Bio-Fixation in a Continues Photobioreactor. J. Chem. Pet. Eng. 2020, 54, 47–55. [Google Scholar] [CrossRef]
- Almomani, F. Kinetic Modeling of Microalgae Growth and CO2 Bio-Fixation Using Central Composite Design Statistical Approach. Sci. Total Environ. 2020, 720, 137594. [Google Scholar] [CrossRef]
- Shalaby, A.; Elkamel, A.; Douglas, P.L.; Zhu, Q.; Zheng, Q.P. A Machine Learning Approach for Modeling and Optimization of a CO2 Post-Combustion Capture Unit. Energy 2021, 215, 119113. [Google Scholar] [CrossRef]
- Mink, A.; Schediwy, K.; Posten, C.; Nirschl, H.; Simonis, S.; Krause, M.J. Comprehensive Computational Model for Coupled Fluid Flow, Mass Transfer, and Light Supply in Tubular Photobioreactors Equipped with Glass Sponges. Energies 2022, 15, 7671. [Google Scholar] [CrossRef]
- Molitor, H.R.; Schnoor, J.L. Using Simulated Flue Gas to Rapidly Grow Nutritious Microalgae with Enhanced Settleability. ACS Omega 2020, 5, 27269–27277. [Google Scholar] [CrossRef]
- Patrinou, V.; Daskalaki, A.; Kampantais, D.; Kanakis, D.C.; Economou, C.N.; Bokas, D.; Kotzamanis, Y.; Aggelis, G.; Vayenas, D.V.; Tekerlekopoulou, A.G. Optimization of Cultivation Conditions for Tetraselmis Striata and Biomass Quality Evaluation for Fish Feed Production. Water 2022, 14, 3162. [Google Scholar] [CrossRef]
- Gabrielyan, D.A.; Gabel, B.V.; Sinetova, M.A.; Gabrielian, A.K.; Markelova, A.G.; Shcherbakova, N.V.; Los, D.A. Optimization of CO2 Supply for the Intensive Cultivation of Chlorella sorokiniana IPPAS C-1 in the Laboratory and Pilot-Scale Flat-Panel Photobioreactors. Life 2022, 12, 1469. [Google Scholar] [CrossRef] [PubMed]
- Dey, S.; Bhattacharya, A.; Kumar, P.; Malik, A. High-Rate CO2 sequestration Using a Novel Venturi Integrated Photobioreactor and Subsequent Valorization to Microalgal Lipids. Green. Chem. 2020, 22, 7962–7973. [Google Scholar] [CrossRef]
- Metsoviti, M.N.; Papapolymerou, G.; Karapanagiotidis, I.T.; Katsoulas, N. Effect of Light Intensity and Quality on Growth Rate and Composition of Chlorella vulgaris. Plants 2019, 9, 31. [Google Scholar] [CrossRef]
- Yadav, G.; Dubey, B.K.; Sen, R. A Comparative Life Cycle Assessment of Microalgae Production by CO2 Sequestration from Flue Gas in Outdoor Raceway Ponds under Batch and Semi-Continuous Regime. J. Clean. Prod. 2020, 258, 120703. [Google Scholar] [CrossRef]
- Aslam, A.; Thomas-Hall, S.R.; Mughal, T.; Zaman, Q.; Ehsan, N.; Javied, S.; Schenk, P.M. Heavy Metal Bioremediation of Coal-Fired Flue Gas Using Microalgae under Different CO2 Concentrations. J. Environ. Manag. 2019, 241, 243–250. [Google Scholar] [CrossRef]
- Lage, S.; Toffolo, A.; Gentili, F.G. Microalgal Growth, Nitrogen Uptake and Storage, and Dissolved Oxygen Production in a Polyculture Based-Open Pond Fed with Municipal Wastewater in Northern Sweden. Chemosphere 2021, 276, 130122. [Google Scholar] [CrossRef]
- Almomani, F.; Ketife, A.A.; Judd, S.; Shurair, M.; Bhosale, R.R.; Znad, H.; Tawalbeh, M. Impact of CO2 Concentration and Ambient Conditions on Microalgal Growth and Nutrient Removal from Wastewater by a Photobioreactor. Sci. Total Environ. 2019, 662, 662–671. [Google Scholar] [CrossRef]
- Siddiki, S.Y.A.; Mofijur, M.; Kumar, P.S.; Ahmed, S.F.; Inayat, A.; Kusumo, F.; Badruddin, I.A.; Khan, T.M.Y.; Nghiem, L.D.; Ong, H.C.; et al. Microalgae Biomass as a Sustainable Source for Biofuel, Biochemical and Biobased Value-Added Products: An Integrated Biorefinery Concept. Fuel 2022, 307, 121782. [Google Scholar] [CrossRef]
- Li, G.; Xiao, W.; Yang, T.; Lyu, T. Optimization and Process Effect for Microalgae Carbon Dioxide Fixation Technology Applications Based on Carbon Capture: A Comprehensive Review. C 2023, 9, 35. [Google Scholar] [CrossRef]
- Rinanti, A.; Purwadi, R. Increasing Carbohydrate and Lipid Productivity in Tropical Microalgae Biomass as a Sustainable Biofuel Feed Stock. Energy Procedia 2019, 158, 1215–1222. [Google Scholar] [CrossRef]
- de Morais, M.G.; de Morais, E.G.; Duarte, J.H.; Deamici, K.M.; Mitchell, B.G.; Costa, J.A.V. Biological CO2 Mitigation by Microalgae: Technological Trends, Future Prospects and Challenges. World J. Microbiol. Biotechnol. 2019, 35, 78. [Google Scholar] [CrossRef]
- Klin, M.; Pniewski, F.; Latała, A. Growth Phase-Dependent Biochemical Composition of Green Microalgae: Theoretical Considerations for Biogas Production. Bioresour. Technol. 2020, 303, 122875. [Google Scholar] [CrossRef] [PubMed]
- Mandik, Y.I.; Cheirsilp, B.; Srinuanpan, S.; Maneechote, W.; Boonsawang, P.; Prasertsan, P.; Sirisansaneeyakul, S. Zero-Waste Biorefinery of Oleaginous Microalgae as Promising Sources of Biofuels and Biochemicals through Direct Transesterification and Acid Hydrolysis. Process Biochem. 2020, 95, 214–222. [Google Scholar] [CrossRef]
- Comley, J.G.; Scott, J.A.; Laamanen, C.A. Utilizing CO2 in Industrial Off-Gas for Microalgae Cultivation: Considerations and Solutions. Crit. Rev. Biotechnol. 2024, 44, 910–923. [Google Scholar] [CrossRef]
- Mehar, J.; Shekh, A.; Malchira, N.U.; Mudliar, S. Potential of Microalgae for Integrated Biomass Production Utilizing CO2 and Food Industry Wastewater. In Application of Microalgae in Wastewater Treatment; Springer International Publishing: Cham, Switzerland, 2019; pp. 41–67. [Google Scholar]
- Chen, Y.; Xu, C. How to Narrow the CO2 Gap from Growth-Optimal to Flue Gas Levels by Using Microalgae for Carbon Capture and Sustainable Biomass Production. J. Clean. Prod. 2021, 280, 124448. [Google Scholar] [CrossRef]


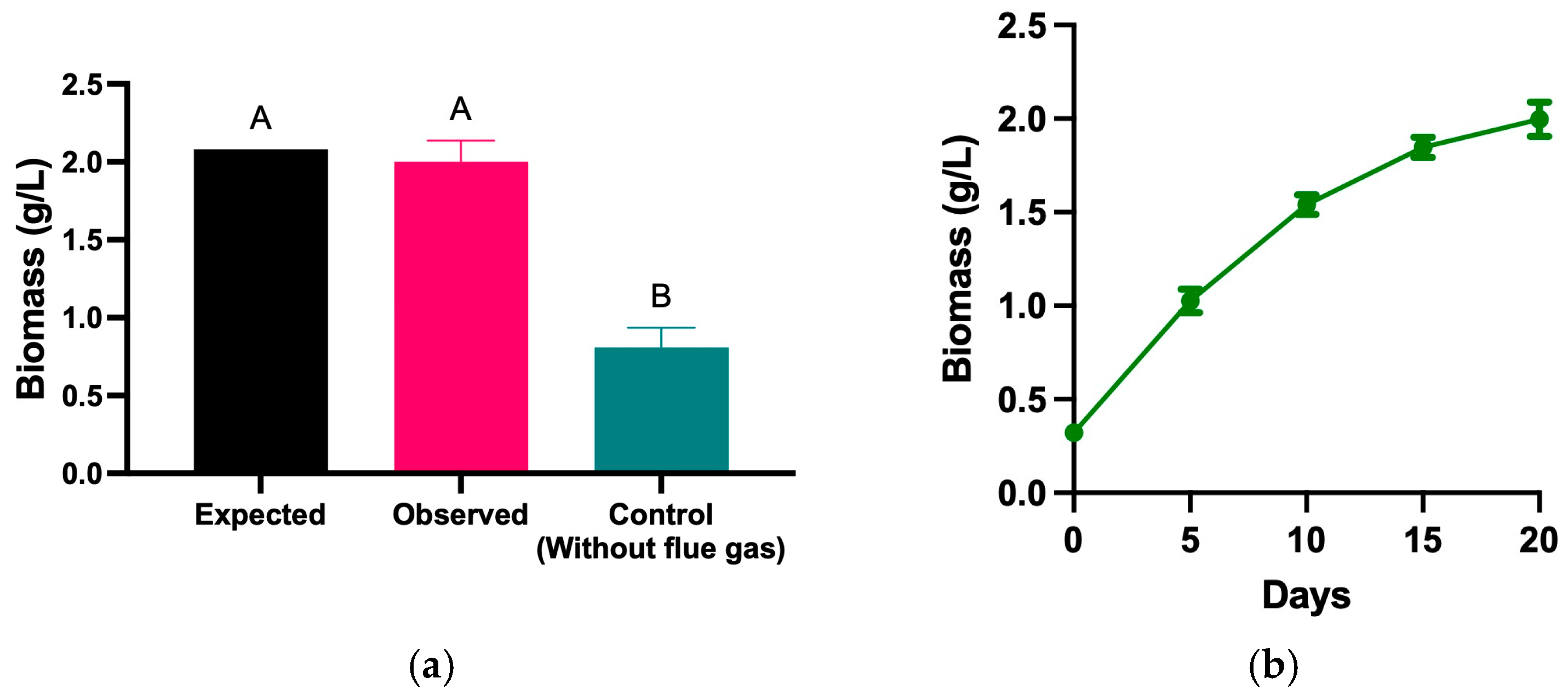


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Name | Units | Type | Levels | ||||
|---|---|---|---|---|---|---|---|---|
| −α | −1 | 0 | +1 | +α | ||||
| A | Photoperiod | h | Numeric (Continuous) | 2.1 | 7 | 12.5 | 18 | 22.9 |
| B | Flue gas | L/min | 0 | 0.1 | 0.55 | 1 | 1.4 | |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| Block | 0.0043 | 1 | 0.0043 | ||
| Model | 8.47 | 5 | 1.69 | 196.68 | <0.0001 * |
| A-Photoperiod | 0.1559 | 1 | 0.1559 | 18.10 | 0.0038 * |
| B-Flue gas | 0.1287 | 1 | 0.1287 | 14.95 | 0.0062 * |
| AB | 0.0036 | 1 | 0.0036 | 0.4181 | 0.5385 ** |
| A2 | 4.43 | 1 | 4.43 | 513.94 | <0.0001 * |
| B2 | 4.38 | 1 | 4.38 | 509.04 | <0.0001 * |
| Residual | 0.0603 | 7 | 0.0086 | ||
| Lack of Fit | 0.0106 | 3 | 0.0035 | 0.2838 | 0.8356 ** |
| Pure Error | 0.0497 | 4 | 0.0124 | ||
| Cor Total | 8.53 | 13 | |||
| R2 | 0.9929 | Adeq Precision | 27.1428 | C.V.% | 7.80 |
| Adjusted R2 | 0.9879 | Std. Dev. | 0.0928 | ||
| Predicted R2 | 0.9768 | Mean | 1.19 |
| Code | Variable | Unit | Value |
|---|---|---|---|
| A | Flue gas | L/min | 0.587 |
| B | Photoperiod | h | 12.6 |
| Z1 | Biomass | g/L | 2.08 |
| Algal Strain | CO2 (% v/v) | Temperature (°C) | Biomass (g/L) | Reference |
|---|---|---|---|---|
| Chlorella sp. | 5 | 27 | 0.36 | [40] |
| 25 | 40 | 1.2 | [30] | |
| 10 | 30 | 2.12 | [41] | |
| C. vulgaris | 10 | 27 | 0.43 | [42] |
| Dunaliella sp. | 3 | 27 | 0.17 | [43] |
| Scenedesmus sp. | 20 | 30 | 1.95 | [44] |
| S. obliquus | 15 | 30 | 2.25 | [45] |
| H. rubescens_UFPS013 | 4.2 | 27 ± 1.2 | 2.0 | This work |
| Strain | Biomass | Proteins | Carbohydrates | Lipids | Reference |
|---|---|---|---|---|---|
| (g/L) | (% w/w) | ||||
| Chlorella sp. | 2.6 | 48.5 | 23.8 | 17.7 | [46] |
| C. vulgaris | 6.6 | 53.0 | 22.0 | 21.9 | [47] |
| C. vulgaris UTEX 1803 | 5.6 | 49.1 | 23.0 | 15.3 | [48] |
| Chloroccocum sp. | 2.9 | 47.2 | 19.7 | 20.2 | [49] |
| Nostoc sp. PCC 9202 | 3.1 | 42.4 | 32.2 | 8.1 | [50] |
| N. punctiforme | 3 | 45.8 | 29.9 | 13.6 | [51] |
| S. vacuolatus | 8.3 | 44.6 | 20.3 | 28 | [52] |
| H. rubescens_UFPS013 | 2 | 48.5 | 29.6 | 9.6 | This work |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuorro, A.; Contreras-Ropero, J.E.; Parada-Solano, R.J.; García-Martínez, J.B.; López-Barrera, G.L.; Urbina-Suarez, N.A.; Barajas-Solano, A.F. Development of a Biotechnological Strategy for CO2 Capture in Coke Plant Emissions Using Microalgae. Processes 2025, 13, 2121. https://doi.org/10.3390/pr13072121
Zuorro A, Contreras-Ropero JE, Parada-Solano RJ, García-Martínez JB, López-Barrera GL, Urbina-Suarez NA, Barajas-Solano AF. Development of a Biotechnological Strategy for CO2 Capture in Coke Plant Emissions Using Microalgae. Processes. 2025; 13(7):2121. https://doi.org/10.3390/pr13072121
Chicago/Turabian StyleZuorro, Antonio, Jefferson E. Contreras-Ropero, Renso J. Parada-Solano, Janet B. García-Martínez, Germán L. López-Barrera, Néstor A. Urbina-Suarez, and Andrés F. Barajas-Solano. 2025. "Development of a Biotechnological Strategy for CO2 Capture in Coke Plant Emissions Using Microalgae" Processes 13, no. 7: 2121. https://doi.org/10.3390/pr13072121
APA StyleZuorro, A., Contreras-Ropero, J. E., Parada-Solano, R. J., García-Martínez, J. B., López-Barrera, G. L., Urbina-Suarez, N. A., & Barajas-Solano, A. F. (2025). Development of a Biotechnological Strategy for CO2 Capture in Coke Plant Emissions Using Microalgae. Processes, 13(7), 2121. https://doi.org/10.3390/pr13072121