

Essential Oil of Greek Citrus sinensis cv New Hall - Citrus aurantium Pericarp: Effect upon Cellular Lipid Composition and Growth of Saccharomyces cerevisiae and Antimicrobial Activity against Bacteria, Fungi, and Human Pathogenic Microorganisms

 ,
,  ,
,  ,
, 

 ,
, 


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction of Essential Oil
2.3. Gas Chromatography–Mass Spectrometry (GC-MS) Analysis
2.4. Antimicrobial Activity of the EO
2.5. Saccharomyces cerevisiae Culture Conditions
2.6. Determinations and Analyses
2.7. Statistical Analysis
3. Results and Discussion
3.1. Composition of the EO
3.2. Antibacterial Activity of the EO
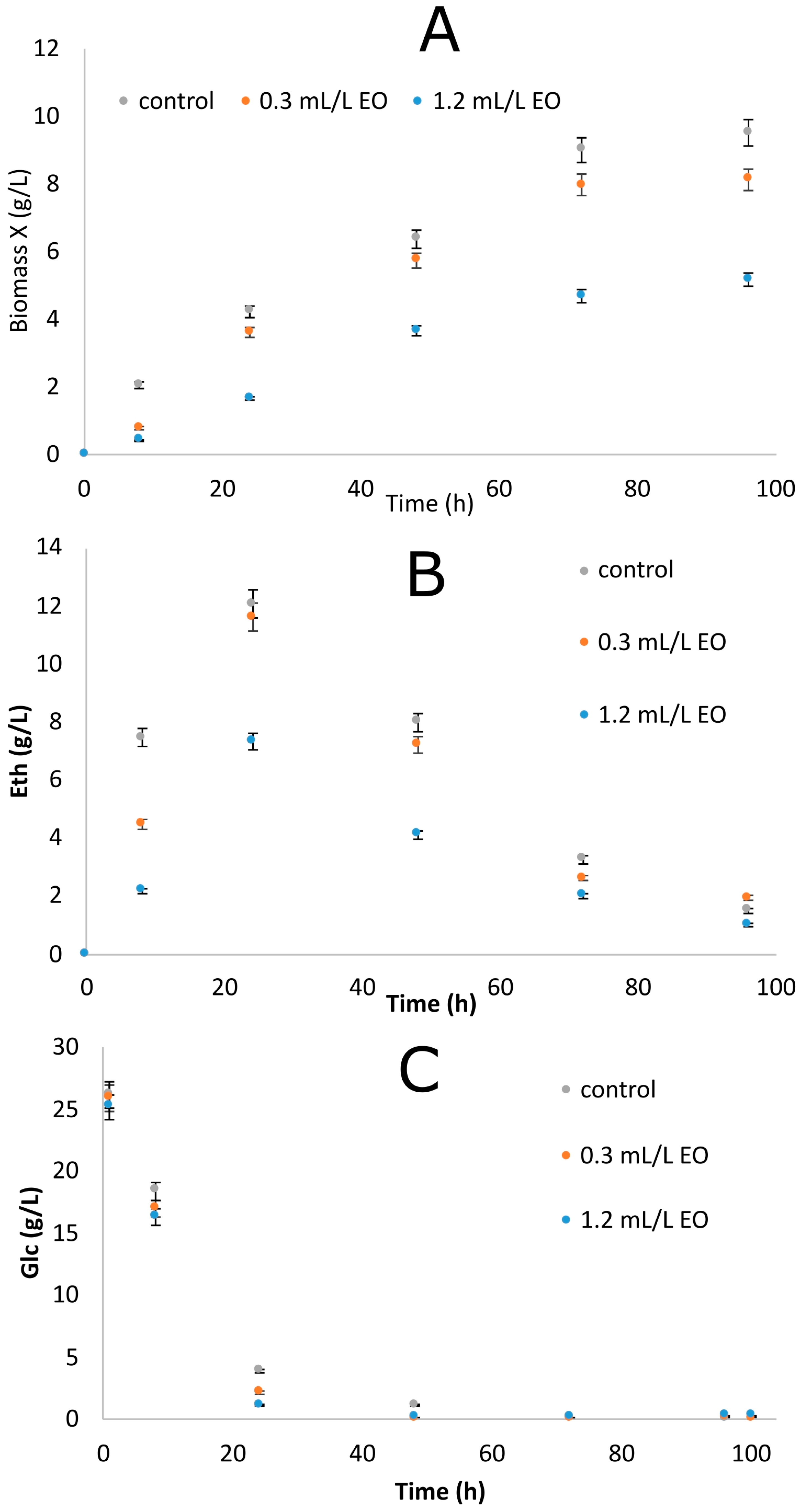
3.3. Effect of the EO on Saccharomyces cerevisiae Growth
3.4. Effect of the EO on the Lipids of Saccharomyces cerevisiae
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Appelhans, M.S.; Bayly, M.J.; Heslewood, M.M.; Groppo, M.; Verboom, G.A.; Forster, P.I.; Kallunki, J.A.; Duretto, M.F. A new subfamily classification of the Citrus family (Rutaceae) based on six nuclear and plastid markers. Taxon 2021, 70, 1035–1061. [Google Scholar] [CrossRef]
- Nicolosi, E.; Deng, Z.N.; Gentile, A.; La Malfa, S.; Continella, G.; Tribulato, E. Citrus phylogeny and genetic origin of important species as investigated by molecular markers. Theor. Appl. Genet. 2000, 100, 1155–1166. [Google Scholar] [CrossRef]
- Schwartz, T.; Nylinder, S.; Ramadugu, C.; Antonelli, A.; Pfeil, B.E. The Origin of Oranges: A Multi-locus Phylogeny of Rutaceae Subfamily Aurantioideae. Syst. Bot. 2016, 40, 1053–1062. [Google Scholar] [CrossRef]
- Guo, Q.; Liu, K.; Deng, W.; Zhong, B.; Yang, W.; Chun, J. Chemical composition and antimicrobial activity of Gannan navel orange (Citrus sinensis Osbeck cv. Newhall) peel essential oils. Food Sci. Nutr. 2018, 6, 1431–1437. [Google Scholar] [CrossRef]
- Sharmeen, J.B.; Mahomoodally, F.M.; Zengin, G.; Maggi, F. Essential oils as natural sources of fragrance compounds for cosmetics and cosmeceuticals. Molecules 2021, 26, 666. [Google Scholar] [CrossRef]
- Razola-Díaz, M.d.C.; Guerra-Hernández, E.J.; García-Villanova, B.; Verardo, V. Recent developments in extraction and encapsulation techniques of orange essential oil. Food Chem. 2021, 354, 129575. [Google Scholar] [CrossRef] [PubMed]
- Verzera, A.; Trozzi, A.; Gazea, F.; Cicciarello, G.; Cotroneo, A. Effects of rootstock on the composition of bergamot (Citrus bergamia Risso et Poiteau) essential oil. J. Agric. Food Chem. 2003, 51, 206–210. [Google Scholar] [CrossRef]
- Bora, H.; Kamle, M.; Mahato, D.K.; Tiwari, P.; Kumar, P. Citrus essential oils (CEOs) and their applications in food: An overview. Plants 2020, 9, 357. [Google Scholar] [CrossRef]
- González-Mas, M.C.; Rambla, J.L.; López-Gresa, M.P.; Blázquez, M.A.; Granell, A. Volatile Compounds in Citrus Essential Oils: A Comprehensive Review. Front. Plant Sci. 2019, 10, 12. [Google Scholar] [CrossRef]
- Gioffrè, G.; Ursino, D.; Labate, M.L.C.; Giuffrè, A.M. The peel essential oil composition of bergamot fruit (Citrus bergamia, Risso) of Reggio Calabria (Italy): A review. Emir. J. Food Agric. 2020, 32, 835. [Google Scholar] [CrossRef]
- Rowshan, V.; Najafian, S. Changes of Peel Essential Oil Composition of Citrus aurantium L. During Fruit Maturation in Iran. J. Essent. Oil-Bearing Plants 2015, 18, 1006–1012. [Google Scholar] [CrossRef]
- Maria, G.A.; Riccardo, N. Citrus bergamia, Risso: The peel, the juice and the seed oil of the bergamot fruit of Reggio Calabria (South Italy). Emir. J. Food Agric. 2020, 32, 522–532. [Google Scholar] [CrossRef]
- Ferhat, M.A.; Boukhatem, M.N.; Hazzit, M.; Meklati, B.Y.; Chemat, F. Cold Pressing, Hydrodistillation and Microwave Dry Distillation of Citrus Essential Oil from Algeria: A Comparative Study. Electron. J. Biol. 2016, 1, 30–41. [Google Scholar]
- Moufida, S.; Marzouk, B. Biochemical characterization of blood orange, sweet orange, lemon, bergamot and bitter orange. Phytochemistry 2003, 62, 1283–1289. [Google Scholar] [CrossRef]
- Tranchida, P.Q.; Bonaccorsi, I.; Dugo, P.; Mondello, L.; Dugo, G. Analysis of Citrus essential oils: State of the art and future perspectives. A review. Flavour Fragr. J. 2012, 27, 98–123. [Google Scholar] [CrossRef]
- Kademi, H.I.; Garba, U. Citrus peel essential oils: A review on composition and antimicrobial activities. Int. J. Food Saf. Nutr. Public Health Technol. 2017, 9, 38–44. [Google Scholar]
- Tamang, J.P.; Shin, D.-H.; Jung, S.-J.; Chae, S.-W. Functional Properties of Microorganisms in Fermented Foods. Front. Microbiol. 2016, 7, 578. [Google Scholar] [CrossRef]
- Benjamin, B.; Bakare, D.V.; Effiong, T.E. Saccharomyces cerevisiae Bio-Ethanol Production as an Alternative Source of Sustainable Energy Ethanol Production using Saccharomyces cerevisiae. Int. J. Res. Appl. Sci. Biotechnol. 2020, 7, 190–194. [Google Scholar] [CrossRef]
- Kasavi, C.; Finore, I.; Lama, L.; Nicolaus, B.; Oliver, S.G.; Toksoy Oner, E.; Kirdar, B. Evaluation of industrial Saccharomyces cerevisiae strains for ethanol production from biomass. Biomass Bioenergy 2012, 45, 230–238. [Google Scholar] [CrossRef]
- Akhtar, N.; Karnwal, A.; Upadhyay, A.K.; Paul, S.; Mannan, M.A.U. Saccharomyces cerevisiae bio-ethanol production, a sustainable energy alternative. Asian J. Microbiol. Biotechnol. Environ. Sci. 2018, 20, S200–S204. [Google Scholar]
- Ceccato-Antonini, S.R.; Shirahigue, L.D.; Varano, A.; da Silva, B.N.; Brianti, C.S.; de Azevedo, F.A. Citrus essential oil: Would it be feasible as antimicrobial in the bioethanol industry? Biotechnol. Lett. 2023, 45, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yi, F.; Jin, R.; Sun, J.; Ma, B.; Bao, X. Evaluation of mechanical-pressed essential oil from Nanfeng mandarin (Citrus reticulata Blanco cv. Kinokuni) as a food preservative based on antimicrobial and antioxidant activities. LWT 2018, 95, 346–353. [Google Scholar] [CrossRef]
- Drive, G.; Suite, A.; Stream, C.; Spectrometry, C.M.; Adams, R.P. Identification of Essential Oil Components by Gas Review: Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2005; Volume 8, ISBN 9781932633214. [Google Scholar]
- Pyrgioti, E.; Graikou, K.; Cheilari, A.; Chinou, I. Assessment of Antioxidant and Antimicrobial Properties of Selected Greek Propolis Samples (North East Aegean Region Islands). Molecules 2022, 27, 8198. [Google Scholar] [CrossRef] [PubMed]
- Widelski, J.; Graikou, K.; Ganos, C.; Skalicka-Wozniak, K.; Chinou, I. Volatiles from Selected Apiaceae Species Cultivated in Poland—Antimicrobial Activities. Processes 2021, 9, 695. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Gortzi, O.; Margeli, E.; Chinou, I.; Galiotou-Panayotou, M.; Lalas, S. Effect of Citrus essential oil addition upon growth and cellular lipids of Yarrowia lipolytica yeast. Eur. J. Lipid Sci. Technol. 2008, 110, 997–1006. [Google Scholar] [CrossRef]
- Wood, I.P.; Elliston, A.; Ryden, P.; Bancroft, I.; Roberts, I.N.; Waldron, K.W. Rapid quantification of reducing sugars in biomass hydrolysates: Improving the speed and precision of the dinitrosalicylic acid assay. Biomass Bioenergy 2012, 44, 117–121. [Google Scholar] [CrossRef]
- Gaff, M.; Esteban-Decloux, M.; Giampaoli, P. Bitter orange peel essential oil: A review of the different factors and chemical reactions influencing its composition. Flavour Fragr. J. 2020, 35, 247–269. [Google Scholar] [CrossRef]
- Farahmandfar, R.; Tirgarian, B.; Dehghan, B.; Nemati, A. Comparison of different drying methods on bitter orange (Citrus aurantium L.) peel waste: Changes in physical (density and color) and essential oil (yield, composition, antioxidant and antibacterial) properties of powders. J. Food Meas. Charact. 2020, 14, 862–875. [Google Scholar] [CrossRef]
- Ngan, T.T.K.; Nguyen, O.B.; Muoi, N.V.; Truc, T.T.; My, V.T.N. Chemical composition and antibacterial activity of orange (citrus sinensis) essential oils obtained by hydrodistillation and solvent free microwave extraction. In Proceedings of the IOP Conference Series: Materials Science and Engineering; IOP Publishing: Bristol, UK, 2020; Volume 991. [Google Scholar]
- Lin, X.; Cao, S.; Sun, J.; Lu, D.; Zhong, B.; Chun, J. The chemical compositions, and antibacterial and antioxidant activities of four types of citrus essential oils. Molecules 2021, 26, 3412. [Google Scholar] [CrossRef] [PubMed]
- Ben Hsouna, A.; Ben Halima, N.; Smaoui, S.; Hamdi, N. Citrus lemon essential oil: Chemical composition, antioxidant and antimicrobial activities with its preservative effect against Listeria monocytogenes inoculated in minced beef meat. Lipids Health Dis. 2017, 16, 146. [Google Scholar] [CrossRef]
- de Souza Pedrosa, G.T.; de Carvalho, R.J.; Berdejo, D.; de Souza, E.L.; Pagán, R.; Magnani, M. Control of Autochthonous Spoilage Lactic Acid Bacteria in Apple and Orange Juices by Sensorially Accepted Doses of Citrus Spp. Essential Oils Combined with Mild Heat Treatments. J. Food Sci. 2019, 84, 848–858. [Google Scholar] [CrossRef] [PubMed]
- Prabuseenivasan, S.; Jayakumar, M.; Ignacimuthu, S. In vitro antibacterial activity of some plant essential oils. BMC Complement. Altern. Med. 2006, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Sarris, D.; Papanikolaou, S. Biotechnological production of ethanol: Biochemistry, processes and technologies. Eng. Life Sci. 2016, 16, 307–329. [Google Scholar] [CrossRef]
- Terpou, A.; Dimopoulou, M.; Belka, A.; Kallithraka, S.; Nychas, G.-J.E.; Papanikolaou, S. Effect of Myclobutanil Pesticide on the Physiological Behavior of Two Newly Isolated Saccharomyces cerevisiae Strains during Very-High-Gravity Alcoholic Fermentation. Microorganisms 2019, 7, 666. [Google Scholar] [CrossRef]
- Aggelis, G.; Athanassopoulos, N.; Paliogianni, A.; Komaitis, M. Effect of a Teucrium polium L. extract on the growth and fatty acid composition of Saccharomyces cerevisiae and Yarrowia lipolytica. Int. J. Gen. Mol. Microbiol. 1998, 73, 195–198. [Google Scholar] [CrossRef]
- Parapouli, M.; Vasileiadis, A.; Afendra, A.S.; Hatziloukas, E. Saccharomyces cerevisiae and its industrial applications. AIMS Microbiol. 2020, 6, 1–31. [Google Scholar] [CrossRef]
- Roukas, T. Ethanol production from non-sterilized beet molasses by free and immobilized Saccharomyces cerevisiae cells using fed-batch culture. J. Food Eng. 1996, 27, 87–96. [Google Scholar] [CrossRef]
- Roukas, T. Kinetics of ethanol production from carob pods extract by immobilized Saccharomyces cerevisiae cells. Appl. Biochem. Biotechnol. 1994, 44, 49–64. [Google Scholar] [CrossRef]
- Çaylak, B.; Vardar Sukan, F. Comparison of different production processes for bioethanol. Turk. J. Chem. 1998, 22, 351–359. [Google Scholar]
- Wang, R.; Ji, Y.; Melikoglu, M.; Koutinas, A.; Webb, C. Optimization of innovative ethanol production from wheat by response surface methodology. Process Saf. Environ. Prot. 2007, 85, 404–412. [Google Scholar] [CrossRef]
- Lin, Y.; Tanaka, S. Ethanol fermentation from biomass resources: Current state and prospects. Appl. Microbiol. Biotechnol. 2006, 69, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Sarris, D.; Matsakas, L.; Aggelis, G.; Koutinas, A.A.; Papanikolaou, S. Aerated vs non-aerated conversions of molasses and olive mill wastewaters blends into bioethanol by Saccharomyces cerevisiae under non-aseptic conditions. Ind. Crops Prod. 2014, 56, 83–93. [Google Scholar] [CrossRef]
- Roukas, T.; Kotzekidou, P. From food industry wastes to second generation bioethanol: A review. Rev. Environ. Sci. Biotechnol. 2022, 21, 299–329. [Google Scholar] [CrossRef]
- Basa, K.; Papanikolaou, S.; Dimopoulou, M.; Terpou, A.; Kallithraka, S.; Nychas, G.J.E. Trials of Commercial-and Wild-Type Saccharomyces cerevisiae Strains under Aerobic and Microaerophilic/Anaerobic Conditions: Ethanol Production and Must Fermentation from Grapes of Santorini (Greece) Native Varieties. Fermentation 2022, 8, 249. [Google Scholar] [CrossRef]
- del Castillo Agudo, L. Lipid content of Saccharomyces cerevisiae strains with different degrees of ethanol tolerance. Appl. Microbiol. Biotechnol. 1992, 37, 647–651. [Google Scholar] [CrossRef]
- You, K.M.; Rosenfield, C.L.; Knipple, D.C. Ethanol tolerance in the yeast Saccharomyces cerevisiae is dependent on cellular oleic acid content. Appl. Environ. Microbiol. 2003, 69, 1499–1503. [Google Scholar] [CrossRef]
- Moreton, R.S. Modification of fatty acid composition of lipid accumulating yeasts with cyclopropene fatty acid desaturase inhibitors. Appl. Microbiol. Biotechnol. 1985, 22, 42–45. [Google Scholar] [CrossRef]
- Chatzifragkou, A.; Petrou, I.; Gardeli, C.; Komaitis, M.; Papanikolaou, S. Effect of Origanum vulgare L. essential oil on growth and lipid profile of Yarrowia lipolytica cultivated on glycerol-based media. J. Am. Oil Chem. Soc. 2011, 88, 1955–1964. [Google Scholar] [CrossRef]
- Xenopoulos, E.; Giannikakis, I.; Chatzifragkou, A.; Koutinas, A.; Papanikolaou, S. Lipid Production by Yeasts Growing on Commercial Xylose in Submerged Cultures with Process Water Being Partially Replaced by Olive Mill Wastewaters. Processes 2020, 8, 819. [Google Scholar] [CrossRef]
- Filippousi, R.; Diamantopoulou, P.; Stavropoulou, M.; Makris, D.P.; Papanikolaou, S. Lipid production by Rhodosporidium toruloides from biodiesel-derived glycerol in shake flasks and bioreactor: Impact of initial C/N molar ratio and added onion-peel extract. Process Biochem. 2022, 123, 52–62. [Google Scholar] [CrossRef]


{kind=link}
| No | Compound | % | No | Compound | % |
|---|---|---|---|---|---|
| 1 | α-Pinene | 1.07 ± 0.02 * | 25 | Geraniol | 0.14 ± 0.01 |
| 2 | Sabinene | 0.49 ± 0.01 | 26 | Geranial | 0.97 ± 0.03 |
| 3 | Myrcene | 1.12 ± 0.03 | 27 | Limonen-10-ol | 0.13 ± 0.01 |
| 4 | β-Pinene | 4.14 ± 0.18 | 28 | Carvacrol | 0.16 ± 0.01 |
| 5 | Limonene | 76.25 ± 2.06 | 29 | Decadienal <trans-2,4-> | 0.12 ± 0.01 |
| 6 | γ-Terpinene | 0.19 ± 0.01 | 30 | Citronellyl acetate | 0.19 ± 0.01 |
| 7 | Octanol <n-> | 0.55 ± 0.02 | 31 | Eugenol | 0.15 ± 0.01 |
| 8 | Terpinolene | 0.18 ± 0.01 | 32 | α-Copaene | 0.23 ± 0.01 |
| 9 | Linaool | 1.77 ± 0.04 | 33 | β-Cubebene | 0.45 ± 0.02 |
| 10 | Menthatriene <1,3,8-para-> | 0.16 ± 0.01 | 34 | Dodecanal | 0.22 ± 0.01 |
| 11 | Mentha-2,8-dien-1-ol <trans-para-> | 0.17 ± 0.01 | 35 | Caryophyllene <trans-> | 0.16 ± 0.01 |
| 12 | Limonene oxide <cis-> | 0.12 ± 0.01 | 36 | α-Bergamotene | 0.13 ± 0.01 |
| 13 | Limonene oxide <trans-> | 0.23 ± 0.01 | 37 | Aromadendrene | 0.10 ± 0.01 |
| 14 | Camphor | 0.11 ± 0.01 | 38 | α-Humulene | 0.18 ± 0.01 |
| 15 | β-Terpineol | 0.15 ± 0.01 | 39 | Germacrene-d | 0.30 ± 0.01 |
| 16 | Citronellal | 0.10 ± 0.01 | 40 | Valencene | 1.15 ± 0.03 |
| 17 | Terpinene-4-ol | 0.44 ± 0.02 | 41 | α-Farnesene | 0.17 ± 0.01 |
| 18 | α-Terpineol | 1.29 ± 0.03 | 42 | δ-Cadinene | 0.40 ± 0.01 |
| 19 | Decanal | 0.60 ± 0.01 | 43 | Elemol | 0.11 ± 0.01 |
| 20 | Octanol acetate | 0.11 ± 0.01 | 44 | Nerolidol | 0.14 ± 0.01 |
| 21 | Carveol <trans-> | 0.26 ± 0.01 | 45 | γ-Eudesmol | 0.18 ± 0.01 |
| 22 | Citronellol | 0.27 ± 0.01 | 46 | Muurolol <T-> | 0.10 ± 0.01 |
| 23 | Neral | 0.35 ± 0.02 | 47 | Farnesol | 0.67 ± 0.03 |
| 24 | Carvone | 0.13 ± 0.01 | 48 | α-Sinensal | 0.10 ± 0.01 |
| Total | 96.68 | ||||
| S. aureus | S. epidermidis | P. aeruginosa | K. pneumoniae | E. cloacae | E. coli | C. albicans | C. tropicalis | C. glabrata | |
|---|---|---|---|---|---|---|---|---|---|
| Citrus studied EO | 520 ± 16 | 460 ± 18 | 880 ± 21 | 860 ± 23 | 900 ± 28 | 740 ± 23 | 1100 ± 24 | 1050 ± 29 | 940 ± 21 |
| Netilmicin | 3.5 ± 0.1 | 3.4 ± 0.1 | 7.2 ± 0.2 | 7.7 ± 0.2 | 6.8 ± 0.2 | 3.6 ± 0.1 | - | - | - |
| Amoxicillin | 2 ± 0.1 | 1.8 ± 0.1 | 2 ± 0.1 | 2 ± 0.1 | 2.5 ± 0.1 | 2 ± 0.1 | - | - | - |
| 5-Flucytosine | - | - | - | - | - | - | 1.2 ± 0.1 | 9.4 ± 0.3 | 9.5 ± 0.2 |
| Amphotericin B | - | - | - | - | - | - | 1 ± 0.1 | 4.2 ± 0.2 | 3.8 ± 0.1 |
| Essential Oil (mL/L) | Fermentation Time (h) | Glcr (g/L) | X (g/L) | Eth (g/L) | YEth/Glc (g/g) |
|---|---|---|---|---|---|
| 0 | 24 | 0.9 ± 0.02 a,* | 4.2 ± 0.08 a | 11.9 ± 0.35 a | 0.47 |
| 72 | 0.3 ± 0.01 A | 9.5 ± 0.30 A | 3.1 ± 0.06 A | ||
| 0.3 | 24 | 0.2 ± 0.01 b | 3.6 ± 0.12 b | 11.8 ± 0.33 a | 0.46 |
| 72 | 0.0 ± 0.00 B | 8.0 ± 0.27 B | 2.6 ± 0.05 B | ||
| 1.2 | 24 | 3.9 ± 0.15 c | 1.7 ± 0.05 c | 7.3 ± 0.19 b | 0.33 |
| 72 | 0.1 ± 0.01 C | 5.1 ± 0.13 C | 1.0 ± 0.02 C |
| C16:0 | Δ9C16:1 | C18:0 | Δ9C18:1 | Δ9,Δ12C18:2 | |
|---|---|---|---|---|---|
| Control experiment (no EO added) | |||||
| Exponential phase (8 h) | 10.4 a,* | 33.0 a | 3.5 a | 47.0 a | 6.0 a |
| Mid-stationary (24 h) | 13.2 b | 30.5 b | 4.4 b | 45.0 b | 6.4 a |
| Late stationary (72 h) | 14.5 c | 29.0 c | 5.6 c | 42.0 c | 8.5 b |
| EO added (0.3 mL/L) | |||||
| Exponential phase (8 h) | 26.7 d | 15.6 d | 7.2 d | 42.0 c | 8.5 b |
| Mid-stationary (24 h) | 25.4 e | 18.0 e | 7.0 d | 40.7 d | 8.0 b |
| Late stationary (72 h) | 24.2 f | 19.4 f | 6.0 c | 42.8 c | 7.2 c |
| EO added (1.2 mL/L) | |||||
| Exponential phase (8 h) | 25.0 e | 6.0 g | 7.7 d | 42.0 c | 11.8 d |
| Mid-stationary (24 h) | 25.5 e | 6.5 g | 7.5 d | 41.5 c | 12.4 d |
| Late stationary (72 h) | 27.2 g | 8.3 h | 6.0 c | 39.4 e | 13.2 e |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bozinou, E.; Athanasiadis, V.; Chatzimitakos, T.; Ganos, C.; Gortzi, O.; Diamantopoulou, P.; Papanikolaou, S.; Chinou, I.; Lalas, S.I. Essential Oil of Greek Citrus sinensis cv New Hall - Citrus aurantium Pericarp: Effect upon Cellular Lipid Composition and Growth of Saccharomyces cerevisiae and Antimicrobial Activity against Bacteria, Fungi, and Human Pathogenic Microorganisms. Processes 2023, 11, 394. https://doi.org/10.3390/pr11020394
Bozinou E, Athanasiadis V, Chatzimitakos T, Ganos C, Gortzi O, Diamantopoulou P, Papanikolaou S, Chinou I, Lalas SI. Essential Oil of Greek Citrus sinensis cv New Hall - Citrus aurantium Pericarp: Effect upon Cellular Lipid Composition and Growth of Saccharomyces cerevisiae and Antimicrobial Activity against Bacteria, Fungi, and Human Pathogenic Microorganisms. Processes. 2023; 11(2):394. https://doi.org/10.3390/pr11020394
Chicago/Turabian StyleBozinou, Eleni, Vassilis Athanasiadis, Theodoros Chatzimitakos, Christos Ganos, Olga Gortzi, Panagiota Diamantopoulou, Seraphim Papanikolaou, Ioanna Chinou, and Stavros I. Lalas. 2023. "Essential Oil of Greek Citrus sinensis cv New Hall - Citrus aurantium Pericarp: Effect upon Cellular Lipid Composition and Growth of Saccharomyces cerevisiae and Antimicrobial Activity against Bacteria, Fungi, and Human Pathogenic Microorganisms" Processes 11, no. 2: 394. https://doi.org/10.3390/pr11020394
APA StyleBozinou, E., Athanasiadis, V., Chatzimitakos, T., Ganos, C., Gortzi, O., Diamantopoulou, P., Papanikolaou, S., Chinou, I., & Lalas, S. I. (2023). Essential Oil of Greek Citrus sinensis cv New Hall - Citrus aurantium Pericarp: Effect upon Cellular Lipid Composition and Growth of Saccharomyces cerevisiae and Antimicrobial Activity against Bacteria, Fungi, and Human Pathogenic Microorganisms. Processes, 11(2), 394. https://doi.org/10.3390/pr11020394