Physalis angulata Calyces Modulate Macrophage Polarization and Alleviate Chemically Induced Intestinal Inflammation in Mice


Abstract
1. Introduction
2. Experimental Section
2.1. Reagents and Chemicals
2.2. Plant Collection and Preparation of the Dichloromethane Fraction from P. angulata
2.3. Cell Culture
2.4. Macrophage Activation and Gene Expression
2.5. Animals
2.6. DSS-Induced Colitis Model
2.7. Histologic Analysis
2.8. Myeloperoxidase (MPO) Activity Assessment
2.9. Determination of Pro-Inflammatory Cytokines in Colon and Mesenteric Lymph Nodes
2.10. Statistical Analysis
3. Results
3.1. Extraction of P. angulata and Preliminary Phytochemical Study of PADF
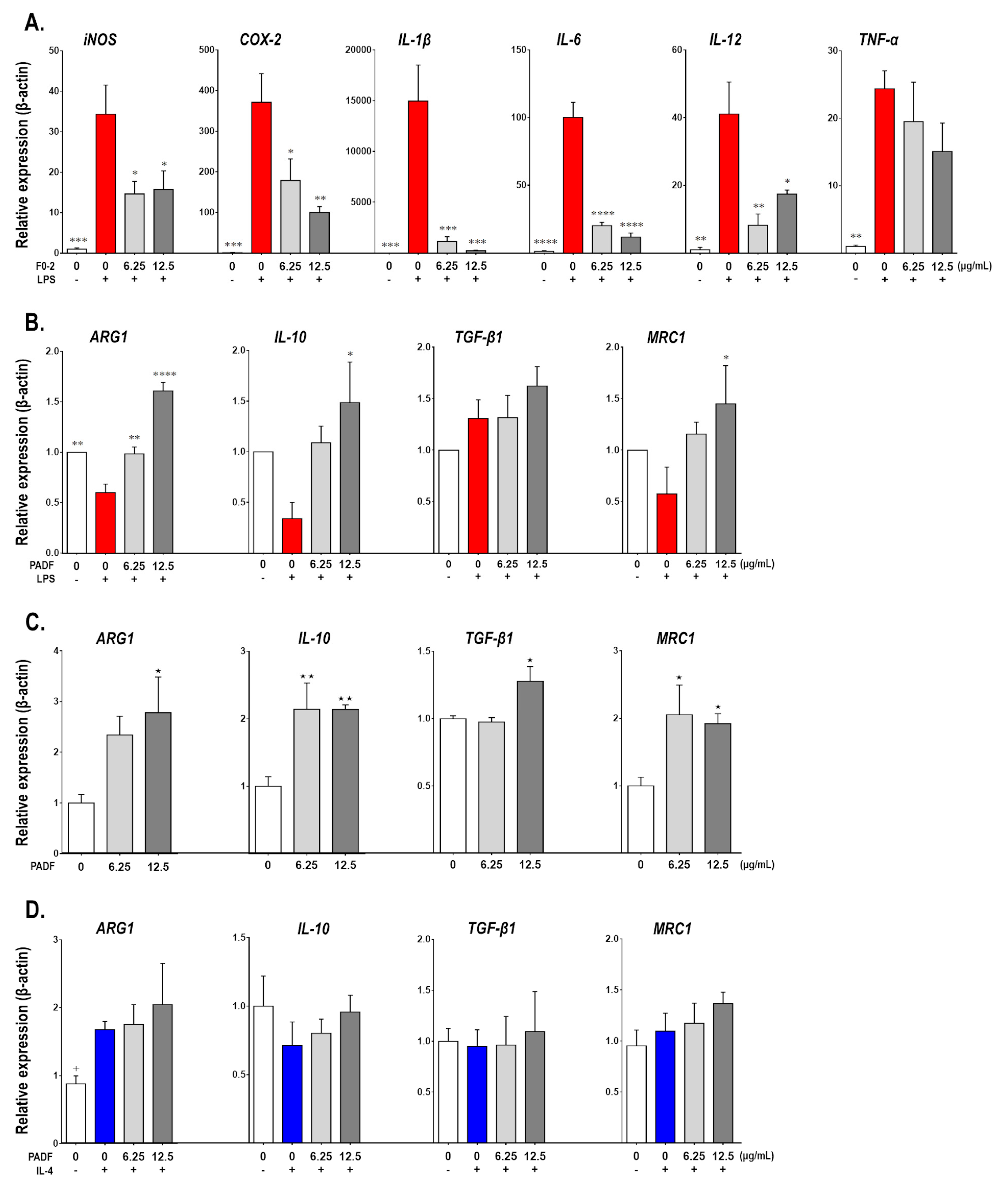
3.2. PADF Modifies the Gene Expression Profile of Resting and LPS-Stimulated RAW 264.7 Macrophages Promoting an Anti-Inflammatory Polarization
3.3. P. angulata Fraction (PADF) Ameliorates DSS-Induced Colitis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PADF | Physalis angulata dichloromethane fraction |
| LPS | Lipopolysaccharide |
| IL | Interleukin |
| TNF- α | Tumor Necrosis Factor α |
| COX-2 | Cyclooxygenase-2 |
| iNOS | Inducible nitric oxide synthase |
| ARG1 | Arginase 1 |
| MRC1 | Mannose receptor C type 1 |
| DSS | Dextran Sodium Sulfate |
| DAI | Disease Activity Index |
| MPO | Myeloperoxidase |
| MLN | Mesenteric Lymph Nodes |
References
- Tambo, E.; Khater, E.I.M.; Chen, J.-H.; Bergquist, R.; Zhou, X.-N. Nobel prize for the artemisinin and ivermectin discoveries: A great boost towards elimination of the global infectious diseases of poverty. Infect. Dis. Poverty 2015, 4, 58. [Google Scholar] [CrossRef]
- WHO. WHO Traditional Medicine Strategy: 2014–2023; WHO Press: Hong Kong, China, 2013; pp. 1–76. [Google Scholar]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef]
- Zhang, W.-N.; Tong, W.-Y. Chemical Constituents and Biological Activities of Plants from the Genus Physalis. Chem. Biodivers. 2016, 13, 48–65. [Google Scholar] [CrossRef]
- Cicchetti, E.; Duroure, L.; Le Borgne, E.; Laville, R. Upregulation of Skin-Aging Biomarkers in Aged NHDF Cells by a Sucrose Ester Extract from the Agroindustrial Waste of Physalis peruviana Calyces. J. Nat. Prod. 2018, 81, 1946–1955. [Google Scholar] [CrossRef]
- Li, A.-L.; Chen, B.-J.; Li, G.-H.; Zhou, M.-X.; Li, Y.-R.; Ren, D.-M.; Lou, H.-X.; Wang, X.-N.; Shen, T. Physalis alkekengi L. var. franchetii (Mast.) Makino: An ethnomedical, phytochemical and pharmacological review. J. Ethnopharmacol. 2018, 210, 260–274. [Google Scholar] [CrossRef]
- Rivera, D.E.; Ocampo, Y.C.; Castro, J.P.; Barrios, L.; Diaz, F.; Franco, L.A. A screening of plants used in Colombian traditional medicine revealed the anti-inflammatory potential of Physalis angulata calyces. Saudi J. Biol. Sci. 2018. [Google Scholar] [CrossRef]
- Morales Saavedra, J.d.C.; Rodríguez Zaragoza, F.A.; Cabrera Toledo, D.; Sánchez Hernández, C.V.; Vargas-Ponce, O. Agromorphological characterization of wild and weedy populations of Physalis angulata in Mexico. Sci. Hortic. 2019, 246, 86–94. [Google Scholar] [CrossRef]
- Rengifo Salgado, E.; Vargas Arana, G. Physalis angulata L. (Bolsa Mullaca): Revisión de Usos Tradicionales, Química y Farmacología. Boletín Latinoamericano y del Caribe de Plantas Medicinales y Aromáticas 2013, 12, 5. [Google Scholar]
- Kindscher, K.; Long, Q.; Corbett, S.; Bosnak, K.; Loring, H.; Cohen, M.; Timmermann, B.N. The Ethnobotany and Ethnopharmacology of Wild Tomatillos, Physalis longifolia Nutt., and Related Physalis Species: A Review. Econ. Bot. 2012, 66, 298–310. [Google Scholar] [CrossRef]
- Ding, N.; Wang, Y.; Dou, C.; Liu, F.; Guan, G.; Wei, K.; Yang, J.; Yang, M.; Tan, J.; Zeng, W.; et al. Physalin D regulates macrophage M1/M2 polarization via the STAT1/6 pathway. J. Cell. Physiol. 2019, 234, 8788–8796. [Google Scholar] [CrossRef]
- Wirtz, S.; Neufert, C.; Weigmann, B.; Neurath, M.F. Chemically induced mouse models of intestinal inflammation. Nat. Protoc. 2007, 2, 541. [Google Scholar] [CrossRef]
- Bradley, P.P.; Priebat, D.A.; Christensen, R.D.; Rothstein, G. Measurement of Cutaneous Inflammation: Estimation of Neutrophil Content with an Enzyme Marker. J. Investig. Dermatol. 1982, 78, 206–209. [Google Scholar] [CrossRef]
- Zheng, Y.; Luan, L.; Chen, Y.; Ren, Y.; Wu, Y. Characterization of physalins and fingerprint analysis for the quality evaluation of Physalis alkekengi L. var. franchetii by ultra-performance liquid chromatography combined with diode array detection and electrospray ionization tandem mass spectrometry. J. Pharm. Biomed. Anal. 2012, 71, 54–62. [Google Scholar] [CrossRef]
- Franco, L.A.; Ocampo, Y.C.; Gómez, H.A.; De la Puerta, R.; Espartero, J.L.; Ospina, L.F. Sucrose Esters from Physalis peruviana Calyces with Anti-Inflammatory Activity. Planta Med. 2014, 80, 1605–1614. [Google Scholar] [CrossRef]
- Mima, H.; Kitamori, N. Chromatographic analysis of sucrose esters of long chain fatty acids. J. Am. Oil Chem. Soc. 1964, 41, 198–200. [Google Scholar] [CrossRef]
- Devkar, S.T.; Suryapujary, S.M.; Jagtap, S.D.; Katyare, S.S.; Hegde, M.V. Effect of macronutrient deficiency on withanolides content in the roots of Withania somnifera and its correlationship with molybdenum content. Pharm. Biol. 2015, 53, 518–523. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Vargas-Ponce, O.; Sánchez Martínez, J.; Zamora Tavares, M.d.P.; Valdivia Mares, L.E. Traditional management of a small-scale crop of Physalis angulata in Western Mexico. Genet. Resour. Crop. Evol. 2016, 63, 1383–1395. [Google Scholar] [CrossRef]
- Ligarreto, G.A.; Lobo, M.; Correa, A. Recursos geneticos del genero Physalis en Colombia. In Avances en Cultivo, Poscosecha y Exportación de la Uchuva (Physalis peruviana L.) en Colombia; Fischer, G., Miranda, D., Piedrahita, W., Romero, J., Eds.; Universidad Nacional de Colombia: Bogota, Colombia, 2005; Volume 1, pp. 14–15. [Google Scholar]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage Activation and Polarization: Nomenclature and Experimental Guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Lin, Y.-S.; Chiang, H.-C.; Kan, W.-S.; Hone, E.; Shih, S.-J.; Won, M.-H. Immunomodulatory Activity of Various Fractions Derived from Physalis angulata L Extract. Am. J. Chin. Med. 1992, 20, 233–243. [Google Scholar] [CrossRef]
- Krause, P.; Morris, V.; Greenbaum, J.A.; Park, Y.; Bjoerheden, U.; Mikulski, Z.; Muffley, T.; Shui, J.-W.; Kim, G.; Cheroutre, H.; et al. IL-10-producing intestinal macrophages prevent excessive antibacterial innate immunity by limiting IL-23 synthesis. Nat. Commun. 2015, 6, 7055. [Google Scholar] [CrossRef]
- Perše, M.; Cerar, A. Dextran Sodium Sulphate Colitis Mouse Model: Traps and Tricks. J. Biomed. Biotechnol. 2012, 2012, 718617. [Google Scholar] [CrossRef]
- Almeida Junior, L.D.; Quaglio, A.E.V.; de Almeida Costa, C.A.R.; Di Stasi, L.C. Intestinal anti-inflammatory activity of Ground Cherry (Physalis angulata L.) standardized CO2 phytopharmaceutical preparation. World J. Gastroenterol. 2017, 23, 4369–4380. [Google Scholar] [CrossRef]
- Zhang, C.-R.; Khan, W.; Bakht, J.; Nair, M.G. New antiinflammatory sucrose esters in the natural sticky coating of tomatillo (Physalis philadelphica), an important culinary fruit. Food Chem. 2016, 196, 726–732. [Google Scholar] [CrossRef]
- Pérez-Castorena, A.-L.; Martínez, M.; Maldonado, E. Labdanes and Sucrose Esters from Physalis sordida. J. Nat. Prod. 2010, 73, 1271–1276. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Official Name | Primer Sequence 1 |
|---|---|---|
| ACTB (β-actin) | Actin, beta | F: TAGGCGGACTGTTACTGAGC R: TGCTCCAACCAACTGCTGTC |
| ARG1 | Arginase | F: TCGTGTACATTGGCTTGCGA R: GCCAATCCCCAGCTTGTCTA |
| PTGS2 (COX-2) | Prostaglandin endoperoxide synthase 2 | F: TTCAACACACTCTATCACTGGC R: AGAAGCGTTTGCGGTACTCAT |
| IL-1β | Interleukin 1 beta | F: CTTCCTTGTGCAAGTGTCTG R: GCCTGAAGCTCTTGTTGATG |
| IL-6 | Interleukin 6 | F: CTGCAAGAGACTTCCATCCAG R: AGTGGTATAGACAGGTCTGTTGG |
| IL-10 | Interleukin 10 | F: TGCCTGCTCTTACTGACTGG R: CTGGGAAGTGGGTGCAGTTA |
| IL-12 | Interleukin 12 | F: TGGTTTGCCATCGTTTTGCTG R: ACAGGTGAGGTTCACTGTTTCT |
| iNOS | Nitric oxide synthase 2, inducible | F: ACATCGACCCGTCCACAGTAT R: CAGAGGGGTAGGCTTGTCTC |
| MRC1 | Mannose receptor, C type 1 | F: GCTTCCGTCACCCTGTATGC R: TCATCCGTGGTTCCATAGACC |
| TGF-β1 | Transforming growth factor, beta 1 | F: ACTGGAGTTGTACGGCAGTG R: TCATGTCATGGATGGTGCCC |
| TNF (TNF-α) | Tumor necrosis factor | F: ACCCTCACACTCAGATCATC R: GAGTAGACAAGGTACAACCC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera, D.; Ocampo, Y.; Franco, L.A. Physalis angulata Calyces Modulate Macrophage Polarization and Alleviate Chemically Induced Intestinal Inflammation in Mice. Biomedicines 2020, 8, 24. https://doi.org/10.3390/biomedicines8020024
Rivera D, Ocampo Y, Franco LA. Physalis angulata Calyces Modulate Macrophage Polarization and Alleviate Chemically Induced Intestinal Inflammation in Mice. Biomedicines. 2020; 8(2):24. https://doi.org/10.3390/biomedicines8020024
Chicago/Turabian StyleRivera, David, Yanet Ocampo, and Luis A. Franco. 2020. "Physalis angulata Calyces Modulate Macrophage Polarization and Alleviate Chemically Induced Intestinal Inflammation in Mice" Biomedicines 8, no. 2: 24. https://doi.org/10.3390/biomedicines8020024
APA StyleRivera, D., Ocampo, Y., & Franco, L. A. (2020). Physalis angulata Calyces Modulate Macrophage Polarization and Alleviate Chemically Induced Intestinal Inflammation in Mice. Biomedicines, 8(2), 24. https://doi.org/10.3390/biomedicines8020024