Cytotoxicity of Extracts from New Zealand Surf Clams Against Organ Cancer Cell Lines




Abstract
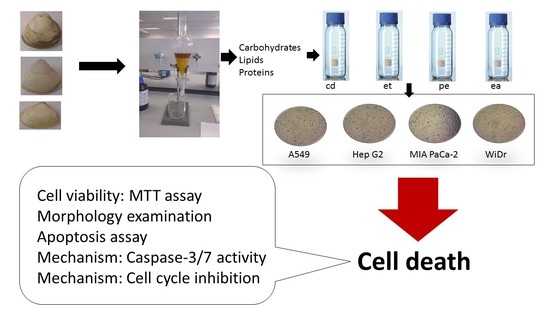
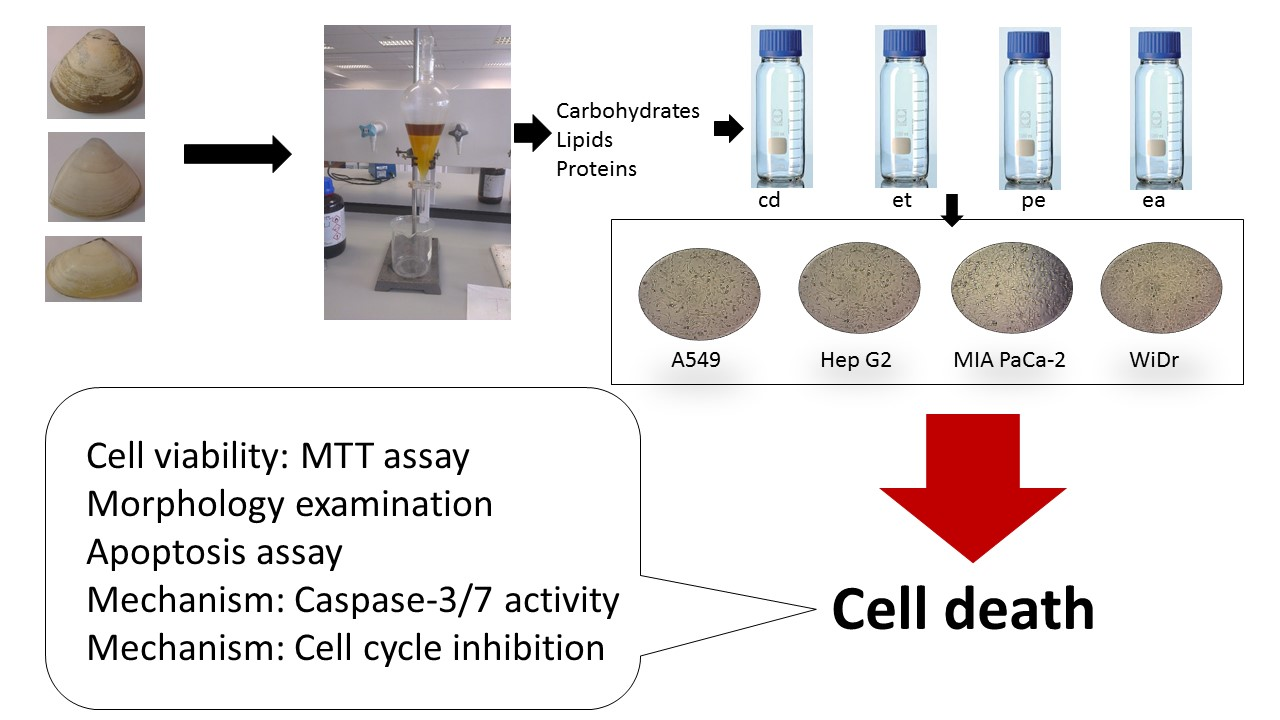{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
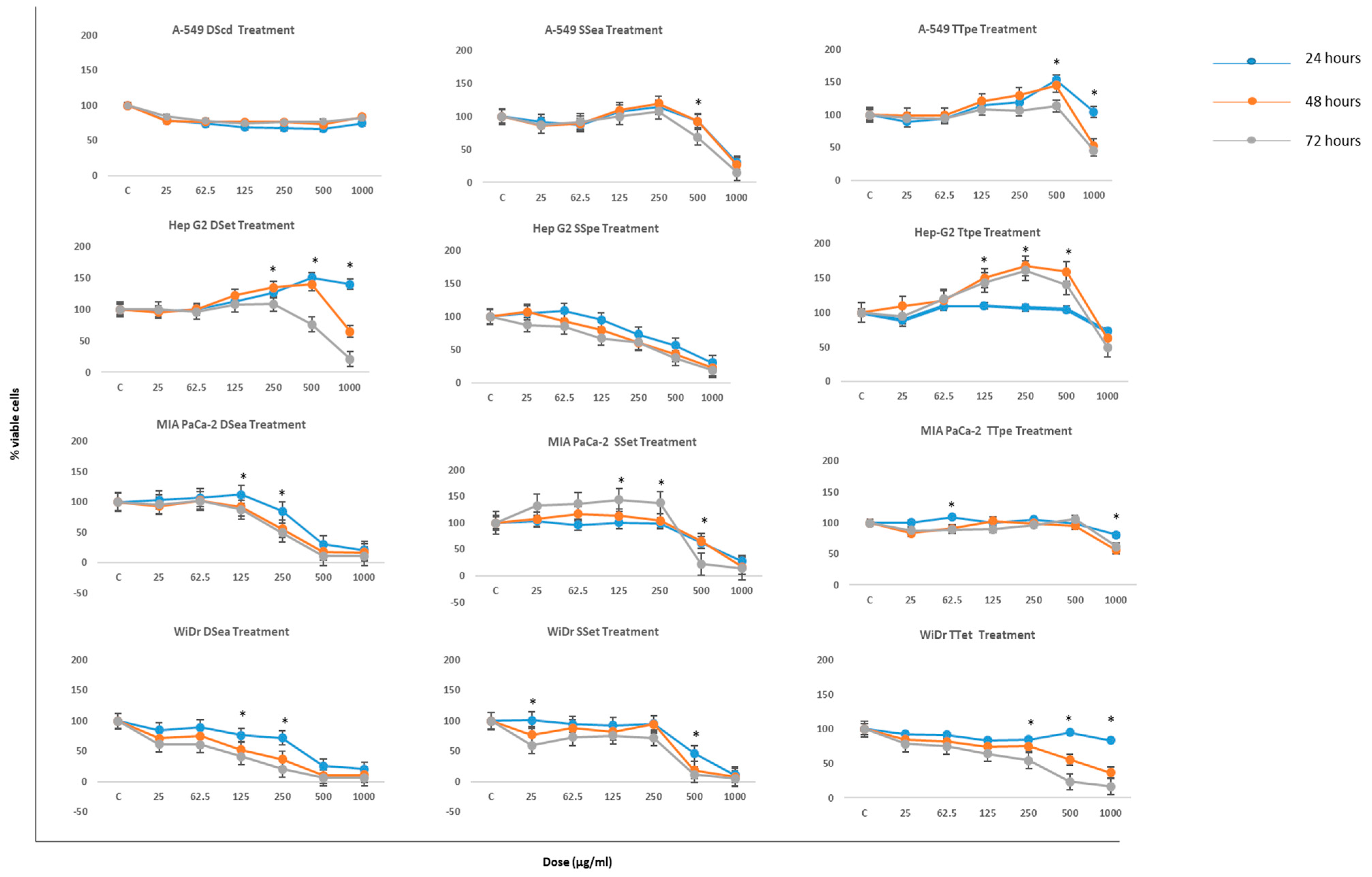
2.1. Antiproliferative Activities of NZ Surf Clam Extracts
2.2. Morphological Analysis
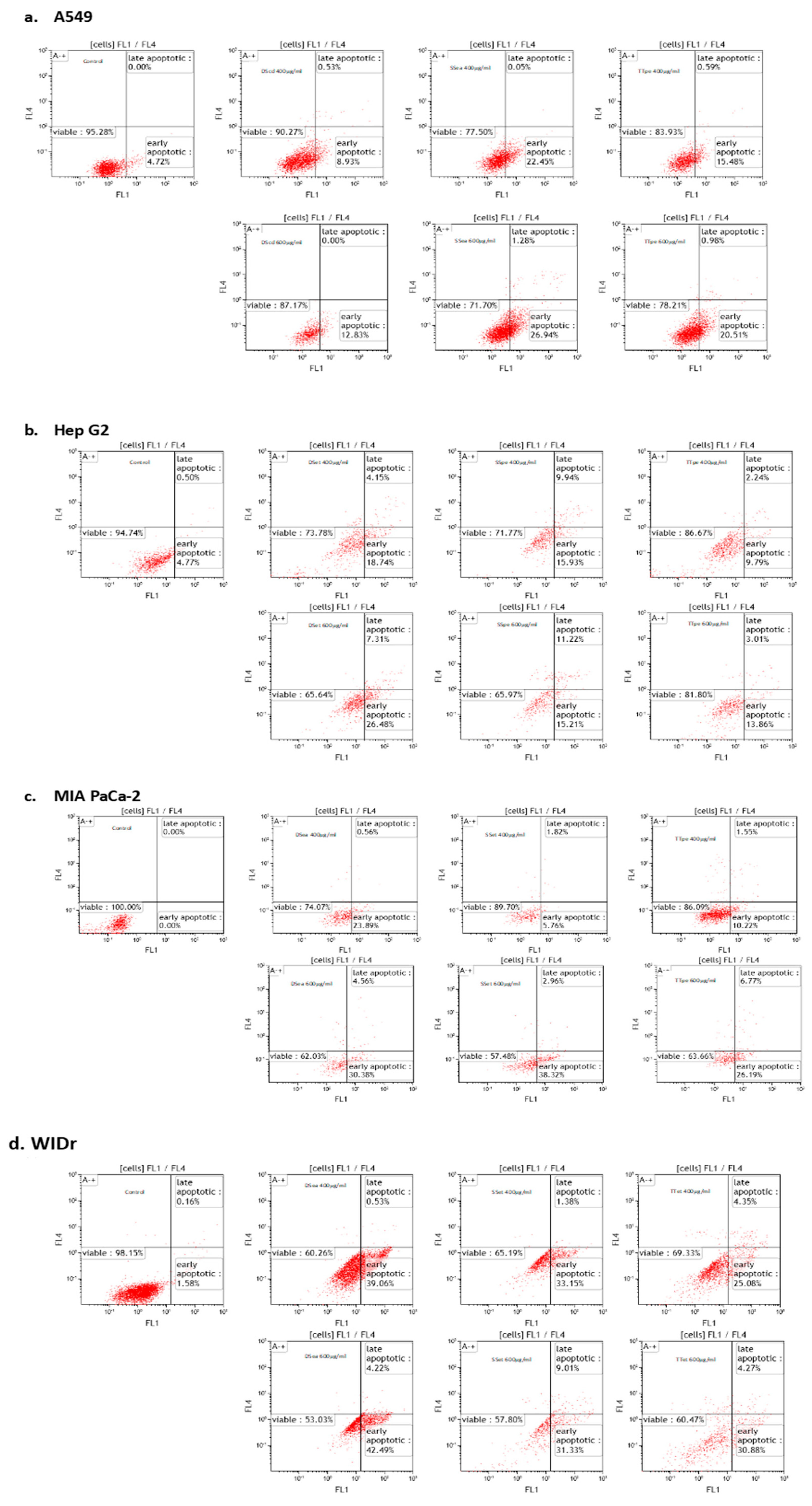
2.3. Apoptosis Inducing Activity of NZ Surf Clam Extracts
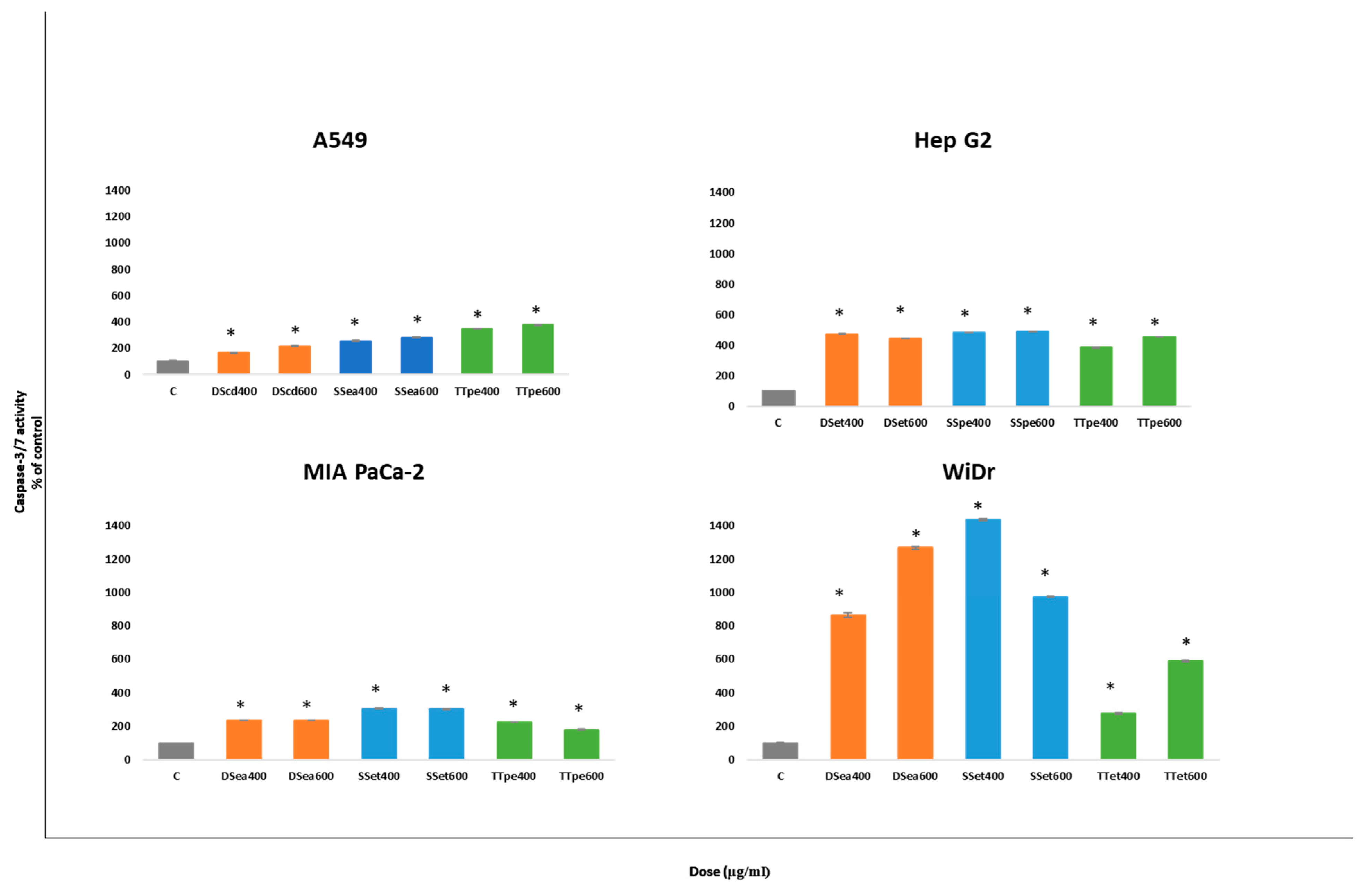
2.4. Caspase-3/7 Activity of NZ Surf Clam Extracts
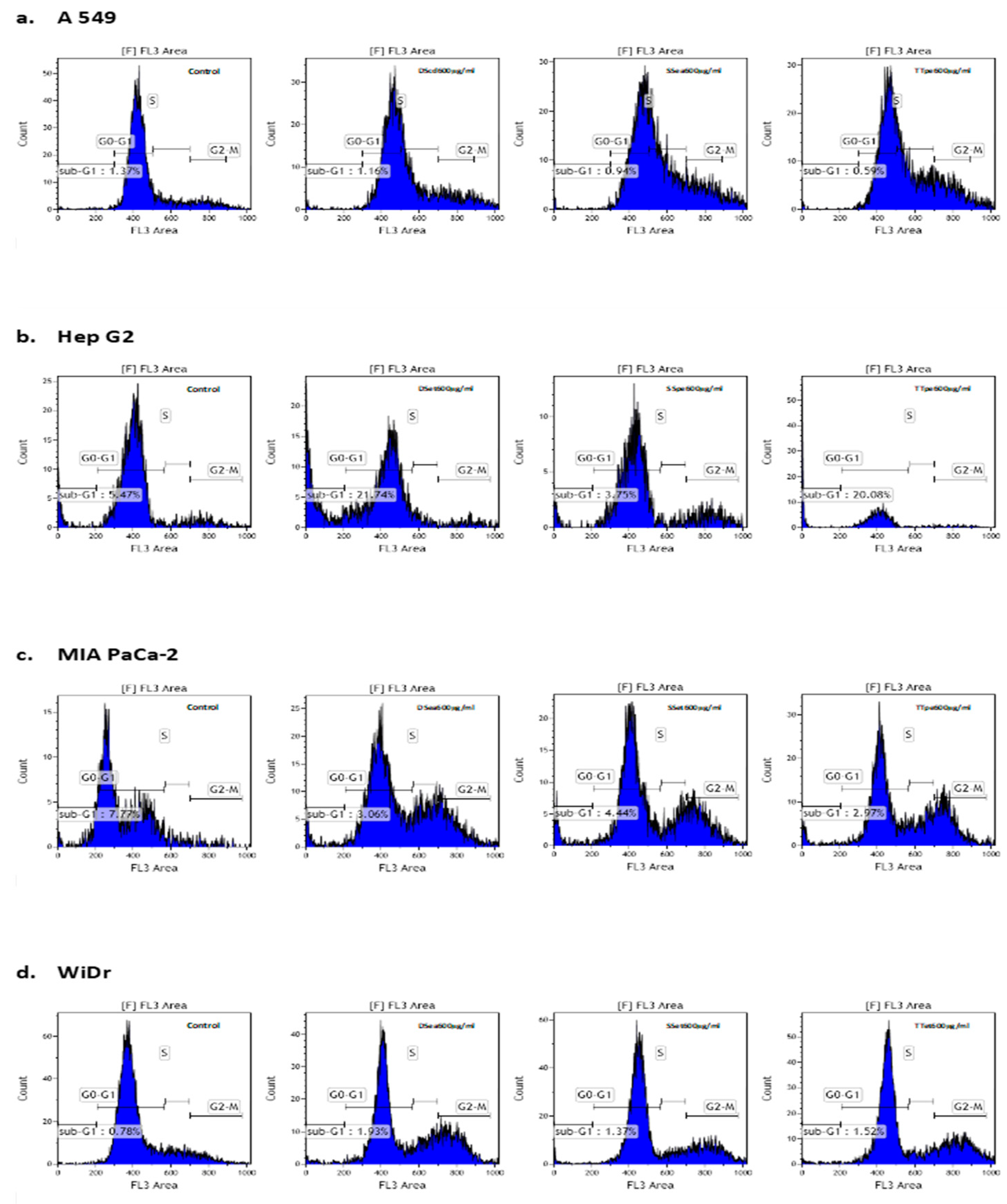
2.5. Cell Cycle Alterations by NZ Surf Clam Extracts
2.6. Chemical Composition of Extracts
3. Materials and Methods
3.1. Materials
3.2. Extraction and Fractionation of Clam Samples
3.3. Cell Proliferation Assay
Data Interpretation
3.4. Cell Apoptosis Assay
3.5. Cell Cycle Assay
3.6. The Apo-ONE® Homogeneous Caspase-3/7 Assay
3.7. Statistical Analysis
4. Conclusions
5. Future Research
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Singh, R.; Sharma, M.; Joshi, P.; Rawat, D.S. Clinical Status of Anti-Cancer Agents Derived from Marine Sources. Anti-Cancer Agents Med. Chem. (Former. Curr. Med. Chem. Anti-Cancer Agents) 2008, 8, 603–617. [Google Scholar] [CrossRef]
- Jung, W.-K.; Kim, S.K. Isolation and characterisation of an anticoagulant oligopeptide from blue mussel, Mytilus edulis. Food Chem. 2009, 117, 687–692. [Google Scholar] [CrossRef]
- Mohan, K.; Abirami, P.; Kanchana, S.; Arumugam, M. Isolation and characterization of Molluscan glycosaminoglycans from Pazhayar, South-East coast of India. Int. J. Fish. Aquat. Stud. 2016, 4, 100–105. [Google Scholar]
- Gomes, A.M.; Kozlowski, E.O.; Pomin, V.H.; de Barros, C.M.; Zaganeli, J.L.; Pavão, M.S. Unique extracellular matrix heparan sulfate from the bivalve Nodipecten nodosus (Linnaeus, 1758) safely inhibits arterial thrombosis after photochemically induced endothelial lesion. J. Biol. Chem. 2010, 285, 7312–7323. [Google Scholar] [CrossRef]
- Lee, H.-J.; Saravana, P.S.; Cho, Y.-N.; Haq, M.; Chun, B.-S. Extraction of bioactive compounds from oyster (Crassostrea gigas) by pressurized hot water extraction. J. Supercrit. Fluids 2018. [Google Scholar] [CrossRef]
- Kumar, D.S.; Janakiram, P.; Kumar, M.M.K.; Geetha, G.K. Inhibitory activity of bioactive compounds isolated from Anadara granosa in shrimp health management. World J. Microbiol. Biotechnol. 2017, 33, 207. [Google Scholar] [CrossRef]
- Zhang, Y.; Dai, B.; Deng, Y.; Zhao, Y. In vitro anti-inflammatory and antioxidant activities and protein quality of high hydrostatic pressure treated squids (Todarodes pacificus). Food Chem. 2016, 203, 258–266. [Google Scholar] [CrossRef]
- Salem, R.B.S.-B.; Ktari, N.; Bkhairia, I.; Nasri, R.; Mora, L.; Kallel, R.; Hamdi, S.; Jamoussi, K.; Boudaouara, T.; El-Feki, A.; et al. In vitro and in vivo anti-diabetic and anti-hyperlipidemic effects of protein hydrolysates from Octopus vulgaris in alloxanic rats. Food Res. Int. 2018, 106, 952–963. [Google Scholar] [CrossRef]
- Odeleye, T.; Li, Y.; White, W.L.; Nie, S.; Chen, S.; Wang, J.; Lu, J. The antioxidant potential of the New Zealand surf clams. Food Chem. 2016, 204, 141–149. [Google Scholar] [CrossRef]
- Pettit, G.R.; Day, J.F.; Hartwell, J.L.; Wood, H.B. Antineoplastic components of marine animals. Nature 1970, 227, 962. [Google Scholar] [CrossRef]
- Benkendorff, K. Molluscan biological and chemical diversity: Secondary metabolites and medicinal resources produced by marine molluscs. Biol. Rev. 2010, 85, 757–775. [Google Scholar] [CrossRef]
- Hua, K.F.; Chen, K.F.; Ho, C.L.; Chen, M.C.; Chen, Y.L.; Chen, W.J.; Huang, J.F.; Perng, Y.S.; Lin, C.C. Freshwater clam extract inhibits inflammatory responses in LPS-activated macrophages by reducing the activation of mitogen-activated protein kinases and NF-kappaB. Nat. Prod. Commun. 2012, 7, 1435–1440. [Google Scholar] [CrossRef]
- Joy, M.; Chakraborty, K. First report of two new antioxidative meroterpeno 2H-pyranoids from short-necked yellow-foot clam Paphia malabarica (family: Veneridae) with bioactivity against pro-inflammatory cyclooxygenases and lipoxygenase. Nat. Prod. Res. 2017, 31, 615–625. [Google Scholar] [CrossRef]
- Joy, M.; Chakraborty, K. Previously undisclosed bioactive sterols from Corbicula bivalve clam Villorita cyprinoides with anti-inflammatory and antioxidant potentials. Steroids 2018, 135, 1–8. [Google Scholar] [CrossRef]
- Ramasamy, M.; Balasubramanian, U. Identification of bioactive compounds and antimicrobial activity of marine clam Anadara granosa (linn.). Int. J. Sci. Nat. 2012, 3, 263–266. [Google Scholar]
- Tsai, J.S.; Lin, T.C.; Chen, J.L.; Pan, B.S. The inhibitory effects of freshwater clam (Corbicula fluminea, Muller) muscle protein hydrolysates on angiotensin I converting enzyme. Process Biochem. 2006, 41, 2276–2281. [Google Scholar] [CrossRef]
- Ajithkumar, P.; Jeganathan, N.S.; Balamurugan, K.; Manvalan, R.; Radha, K. Evaluation of anti-ulcer activity of Villorita cyprinoides extract (black water clams) against immobilization stress induced ulcer in albino rats. J. Pharm. Res. Opin. 2012, 2, 55–57. [Google Scholar]
- Lin, Y.-H.; Tsai, J.-S.; Hung, L.-B.; Pan, B.-S. Hypocholesterolemic effect of compounded freshwater clam protein hydrolysate and Gracilaria. Food Chem. 2010, 123, 395–399. [Google Scholar] [CrossRef]
- GI Cancer Institute. GI Cancer Explained. 2017. Available online: https://gicancer.org.au/gi-cancer-explained/ (accessed on 27 December 2018).
- Song, L.-M.K.; Wilson, B.C. Endoscopic detection of early upper GI cancers. Best Pract. Res. Clin. Gastroenterol. 2005, 19, 833–856. [Google Scholar] [CrossRef]
- British Columbia Cancer. Digestive System. 2018. Available online: http://www.bccancer.bc.ca/health-info/types-of-cancer/digestive-system (accessed on 27 December 2018).
- NIH. Cancer Stat Facts: Lung and Bronchus Cancer. 2018. Available online: https://seer.cancer.gov/statfacts/html/lungb.html (accessed on 27 December 2018).
- Liao, N.; Chen, S.; Ye, X.; Zhong, J.; Wu, N.; Dong, S.; Yang, B.; Liu, D. Antioxidant and anti-tumor activity of a polysaccharide from freshwater clam, Corbicula fluminea. Food Funct. 2013, 4, 539–548. [Google Scholar] [CrossRef]
- Liao, N.; Zhong, J.; Zhang, R.; Ye, X.; Zhang, Y.; Wang, W.; Wang, Y.; Chen, S.; Liu, D.; Liu, R. Protein-Bound Polysaccharide from Corbicula fluminea Inhibits Cell Growth in MCF-7 and MDA-MB-231 Human Breast Cancer Cells. PLoS ONE 2016, 11, e0167889. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Huang, Y.T.; Chang, C.I.; Ho, C.T.; Pan, B.S. Apoptotic-inducing epidioxysterols identified in hard clam (Meretrix lusoria). Food Chem. 2007, 102, 788–795. [Google Scholar] [CrossRef]
- Chi, C.-F.; Hu, F.-Y.; Wang, B.; Li, T.; Ding, G.-F. Antioxidant and anticancer peptides from the protein hydrolysate of blood clam (Tegillarca granosa) muscle. J. Funct. Foods 2015, 15, 301–313. [Google Scholar] [CrossRef]
- Luan, H.-M.; Wang, L.-C.; Wu, H.; Jin, Y.; Ji, J. Antioxidant activities and antioxidative components in the surf clam, Mactra veneriformis. Nat. Prod. Res. 2011, 25, 1838–1848. [Google Scholar] [CrossRef]
- Buranrat, B.; Mairuae, N.; Kanchanarach, W. Cytotoxic and antimigratory effects of Cratoxy formosum extract against Hep G2 liver cancer cells. Biomed. Rep. 2017, 6, 441–448. [Google Scholar] [CrossRef]
- Huang, F.; Zhao, S.; Yu, F.; Yang, Z.; Ding, G. Protective Effects and Mechanism of Meretrix meretrix Oligopeptides against Nonalcoholic Fatty Liver Disease. Mar. Drugs 2017, 15, 31. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, K.C.; Bonzon, C.; Green, D.R. The machinery of programmed cell death. Pharmacol. Ther. 2001, 92, 57–70. [Google Scholar] [CrossRef]
- Huang, S.-T.; Yang, R.-C.; Yang, L.-J.; Pang, J.-H.S. Phyllanthus urinaria triggers the apoptosis and Bcl-2 down-regulation in Lewis lung carcinoma cells. Life Sci. 2003, 72, 1705–1716. [Google Scholar] [CrossRef]
- Invitrogen. Alexa Fluor® 488 annexin V/Dead Cell Apoptosis Kit with Alexa® Fluor 488 annexin V and PI for Flow Cytometry. 2010. Available online: https://assets.thermofisher.com/TFS-Assets/LSG/manuals/mp13241.pdf (accessed on 24 December 2018).
- Huang, F.; Yang, Z.; Yu, D.; Wang, J.; Li, R.; Ding, G. Sepia Ink Oligopeptide Induces Apoptosis in Prostate Cancer Cell Lines via Caspase-3 Activation and Elevation of Bax/Bcl-2 Ratio. Mar. Drugs 2012, 10, 2153–2165. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.M.; Shen, J.Z.; Wang, Y.; Lu, A.X.; Ho, W.S. Anti-oxidant and anti-cancer activities of Angelica dahurica extract via induction of apoptosis in colon cancer cells. Phytomedicine 2016, 23, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-T.; Huang, Y.-H.; Hour, T.-C.; Pan, B.-S.; Liu, Y.-C.; Pan, M.-H. Apoptosis-inducing active components from Corbicula fluminea through activation of caspase-2 and production of reactive oxygen species in human leukemia HL-60 cells. Food Chem. Toxicol. 2006, 44, 1261–1272. [Google Scholar] [CrossRef]
- Song, E.-J.; Chan, M.; Shin, J.W.; Chen, C.-C. Hard clam extracts induce atypical apoptosis in human gastric cancer cells. Exp. Ther. Med. 2017, 14, 1409–1418. [Google Scholar] [CrossRef] [PubMed]
- Chijimatsu, T.; Umeki, M.; Okuda, Y.; Yamada, K.; Oda, H.; Mochizuki, S. The fat and protein fractions of freshwater clam (Corbicula fluminea) extract reduce serum cholesterol and enhance bile acid biosynthesis and sterol excretion in hypercholesterolaemic rats fed a high-cholesterol diet. Br. J. Nutr. 2011, 105, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Chijimatsu, T.; Umeki, M.; Kataoka, Y.; Kobayashi, S.; Yamada, K.; Oda, H.; Mochizuki, S. Lipid components prepared from a freshwater clam (Corbicula fluminea) extract ameliorate hypercholesterolaemia in rats fed high-cholesterol diet. Food Chem. 2013, 136, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, M.W.; Macrides, T.A.; Kalafatis, N.; Betts, W.H.; Haynes, D.R.; Broadbent, J. Anti-inflammatory activity of a lipid fraction (lyprinol) from the NZ green-lipped mussel. Inflammol. Pharmacol. 1997, 5, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Gauvin, A.; Smadja, J.; Aknin, M.; Faure, R.; Gaydou, E.-M. Isolation of bioactive 5α,8α-epidioxysterols from the mare sponge Luffariella cf. variabilis. Can. J. Chem. 2000, 78, 986–992. [Google Scholar]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Odeleye, T.; White, W.L.; Lu, J. Cytotoxicity of Extracts from New Zealand Surf Clams Against Organ Cancer Cell Lines. Biomedicines 2019, 7, 25. https://doi.org/10.3390/biomedicines7020025
Odeleye T, White WL, Lu J. Cytotoxicity of Extracts from New Zealand Surf Clams Against Organ Cancer Cell Lines. Biomedicines. 2019; 7(2):25. https://doi.org/10.3390/biomedicines7020025
Chicago/Turabian StyleOdeleye, Tinu, William Lindsey White, and Jun Lu. 2019. "Cytotoxicity of Extracts from New Zealand Surf Clams Against Organ Cancer Cell Lines" Biomedicines 7, no. 2: 25. https://doi.org/10.3390/biomedicines7020025
APA StyleOdeleye, T., White, W. L., & Lu, J. (2019). Cytotoxicity of Extracts from New Zealand Surf Clams Against Organ Cancer Cell Lines. Biomedicines, 7(2), 25. https://doi.org/10.3390/biomedicines7020025