

Clinical Utility of IFIT Proteins in Human Malignancies
 , and
, and {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
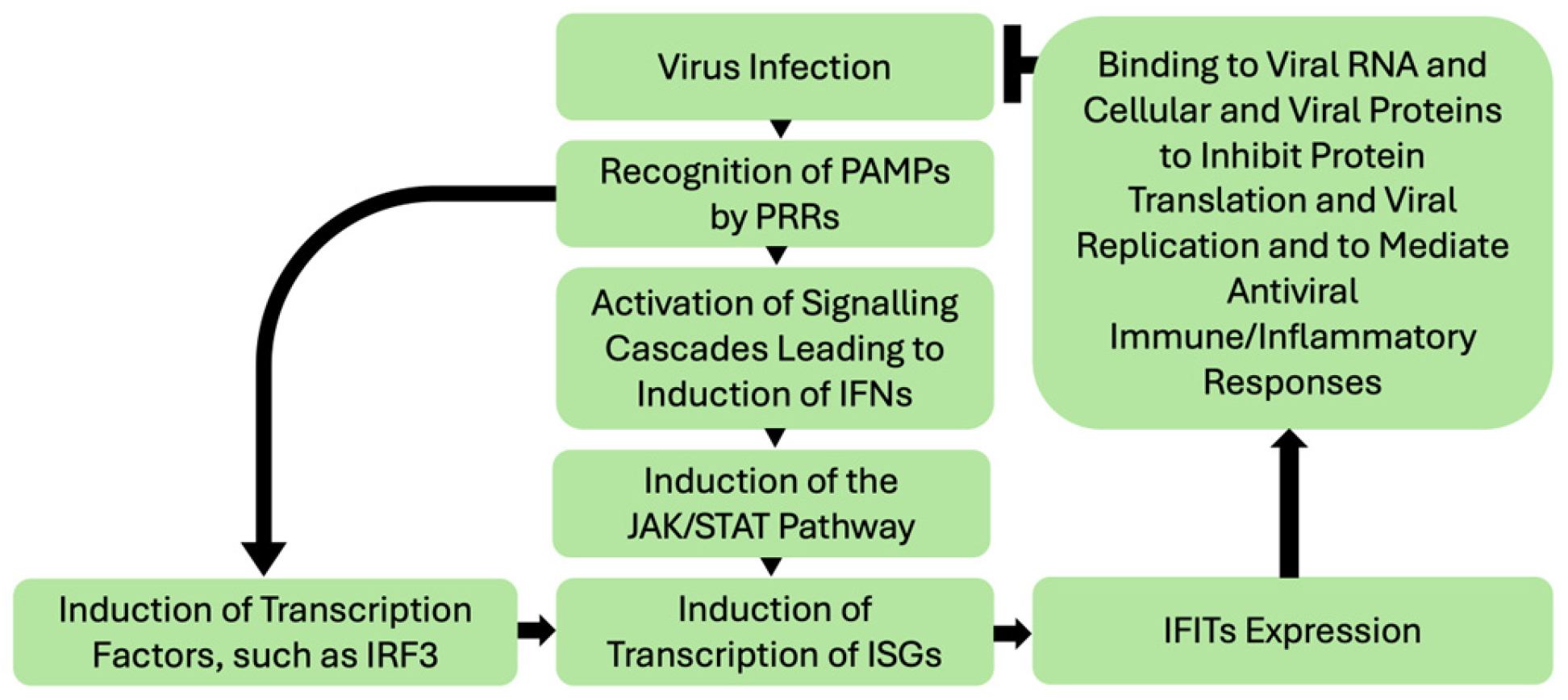


2. IFIT1
2.1. IFIT1 and Unfavorable Clinical Outcomes
2.2. IFIT1 and Favorable Clinical Outcomes
3. IFIT2
3.1. IFIT2 and Unfavorable Clinical Outcomes
3.2. IFIT2 and Favorable Clinical Outcomes
4. IFIT3
4.1. IFIT3 and Unfavorable Clinical Outcomes
4.2. IFIT3 and Favorable Clinical Outcomes
5. IFIT5
5.1. IFIT5 and Unfavorable Clinical Outcomes
5.2. IFIT5 and Favorable Clinical Outcomes
6. Discussion, Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, W.; Li, Y.; Xin, S.; Yang, L.; Jiang, M.; Xin, Y.; Wang, Y.; Cao, P.; Zhang, S.; Yang, Y.; et al. The emerging roles of IFIT3 in antiviral innate immunity and cellular biology. J. Med. Virol. 2023, 95, e28259. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S.; Farzan, M. The broad-spectrum antiviral functions of IFIT and IFITM proteins. Nat. Rev. Immunol. 2013, 13, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Fensterl, V.; Sen, G.C. Interferon-induced Ifit proteins: Their role in viral pathogenesis. J. Virol. 2015, 89, 2462–2468. [Google Scholar] [CrossRef]
- Vladimer, G.I.; Gorna, M.W.; Superti-Furga, G. IFITs: Emerging Roles as Key Anti-Viral Proteins. Front. Immunol. 2014, 5, 94. [Google Scholar] [CrossRef] [PubMed]
- Franco, J.H.; Chattopadhyay, S.; Pan, Z.K. How Different Pathologies Are Affected by IFIT Expression. Viruses 2023, 15, 342. [Google Scholar] [CrossRef]
- Johnson, B.; VanBlargan, L.A.; Xu, W.; White, J.P.; Shan, C.; Shi, P.Y.; Zhang, R.; Adhikari, J.; Gross, M.L.; Leung, D.W.; et al. Human IFIT3 Modulates IFIT1 RNA Binding Specificity and Protein Stability. Immunity 2018, 48, 487–499.e5. [Google Scholar] [CrossRef]
- Mears, H.V.; Sweeney, T.R. Better together: The role of IFIT protein-protein interactions in the antiviral response. J. Gen. Virol. 2018, 99, 1463–1477. [Google Scholar] [CrossRef]
- Pichlmair, A.; Lassnig, C.; Eberle, C.A.; Gorna, M.W.; Baumann, C.L.; Burkard, T.R.; Burckstummer, T.; Stefanovic, A.; Krieger, S.; Bennett, K.L.; et al. IFIT1 is an antiviral protein that recognizes 5′-triphosphate RNA. Nat. Immunol. 2011, 12, 624–630. [Google Scholar] [CrossRef]
- Wu, R.; Yang, H.; Liu, C. IFIT3: A crucial mediator in innate immunity and tumor progression with therapeutic implications. Front. Immunol. 2025, 16, 1515718. [Google Scholar] [CrossRef]
- Zhang, N.; Shi, H.; Yan, M.; Liu, G. IFIT5 Negatively Regulates the Type I IFN Pathway by Disrupting TBK1-IKKepsilon-IRF3 Signalosome and Degrading IRF3 and IKKepsilon. J. Immunol. 2021, 206, 2184–2197. [Google Scholar] [CrossRef]
- Zheng, C.; Zheng, Z.; Zhang, Z.; Meng, J.; Liu, Y.; Ke, X.; Hu, Q.; Wang, H. IFIT5 positively regulates NF-kappaB signaling through synergizing the recruitment of IkappaB kinase (IKK) to TGF-beta-activated kinase 1 (TAK1). Cell Signal. 2015, 27, 2343–2354. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, X.; Chen, W.; Chen, L. IFIT5 potentiates anti-viral response through enhancing innate immune signaling pathways. Acta Biochim. Biophys. Sin. 2013, 45, 867–874. [Google Scholar] [CrossRef]
- Choi, Y.J.; Bowman, J.W.; Jung, J.U. A Talented Duo: IFIT1 and IFIT3 Patrol Viral RNA Caps. Immunity 2018, 48, 474–476. [Google Scholar] [CrossRef] [PubMed]
- Pidugu, V.K.; Pidugu, H.B.; Wu, M.M.; Liu, C.J.; Lee, T.C. Emerging Functions of Human IFIT Proteins in Cancer. Front. Mol. Biosci. 2019, 6, 148. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.F.; Chen, Q.; Hua, S.H.; Yip, G.W. Roles of Interferon Induced Protein with Tetratricopeptide Repeats (IFIT) Family in Cancer. Curr. Med. Chem. 2021, 28, 5034–5047. [Google Scholar] [CrossRef]
- Pidugu, V.K.; Wu, M.M.; Yen, A.H.; Pidugu, H.B.; Chang, K.W.; Liu, C.J.; Lee, T.C. IFIT1 and IFIT3 promote oral squamous cell carcinoma metastasis and contribute to the anti-tumor effect of gefitinib via enhancing p-EGFR recycling. Oncogene 2019, 38, 3232–3247. [Google Scholar] [CrossRef]
- Li, T.H.; Zhao, B.B.; Qin, C.; Wang, Y.Y.; Li, Z.R.; Cao, H.T.; Yang, X.Y.; Zhou, X.T.; Wang, W.B. IFIT1 modulates the proliferation, migration and invasion of pancreatic cancer cells via Wnt/beta-catenin signaling. Cell. Oncol. 2024, 47, 1253–1265. [Google Scholar] [CrossRef]
- Liu, Y.J.; Li, J.P.; Han, M.; Li, J.X.; Ye, Q.W.; Lin, S.T.; Zhou, J.Y.; Liu, S.L.; Zou, X. IFIT1 + neutrophil is a causative factor of immunosuppressive features of poorly cohesive carcinoma (PCC). J. Transl. Med. 2024, 22, 580. [Google Scholar] [CrossRef]
- Zan, X.; Li, S.; Wei, S.; Gao, L.; Zhao, L.; Yan, X.; Zhao, Y.; Shi, J.; Wang, Y.; Liu, R.; et al. COL8A1 Promotes NSCLC Progression Through IFIT1/IFIT3-Mediated EGFR Activation. Front. Oncol. 2022, 12, 707525. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, C.; Zhang, J.; Han, D.; Shi, X. Comprehensive analysis of the prognosis and biological significance for IFIT family in skin cutaneous melanoma. Int. Immunopharmacol. 2021, 101, 108344. [Google Scholar] [CrossRef]
- Mbofung, R.M.; McKenzie, J.A.; Malu, S.; Zhang, M.; Peng, W.; Liu, C.; Kuiatse, I.; Tieu, T.; Williams, L.; Devi, S.; et al. HSP90 inhibition enhances cancer immunotherapy by upregulating interferon response genes. Nat. Commun. 2017, 8, 451. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Shi, L.Z.; Zhao, H.; Chen, J.; Xiong, L.; He, Q.; Chen, T.; Roszik, J.; Bernatchez, C.; Woodman, S.E.; et al. Loss of IFN-gamma Pathway Genes in Tumor Cells as a Mechanism of Resistance to Anti-CTLA-4 Therapy. Cell 2016, 167, 397–404.e9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.F.; Chen, Y.; Lin, G.S.; Zhang, J.D.; Tang, W.L.; Huang, J.H.; Chen, J.S.; Wang, X.F.; Lin, Z.X. High IFIT1 expression predicts improved clinical outcome, and IFIT1 along with MGMT more accurately predicts prognosis in newly diagnosed glioblastoma. Hum. Pathol. 2016, 52, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Chauvin, A.; Wang, C.S.; Geha, S.; Garde-Granger, P.; Mathieu, A.A.; Lacasse, V.; Boisvert, F.M. The response to neoadjuvant chemoradiotherapy with 5-fluorouracil in locally advanced rectal cancer patients: A predictive proteomic signature. Clin. Proteom. 2018, 15, 16. [Google Scholar] [CrossRef]
- He, Y.; Jiang, S.; Cui, Y.; Liang, J.; Zhong, Y.; Sun, Y.; Moran, M.F.; Huang, Z.; He, G.; Mao, X. Induction of IFIT1/IFIT3 and inhibition of Bcl-2 orchestrate the treatment of myeloma and leukemia via pyroptosis. Cancer Lett. 2024, 588, 216797. [Google Scholar] [CrossRef]
- Pidugu, V.K.; Yen, A.H.; Wu, M.M.; Liu, C.J.; Lee, T.C. Characterization of oncogenic activity of interferon-induced protein with tetratricopeptide repeats 1 and 3 in human oral squamous cell carcinoma progression. Abstract 3933 in Proceedings of the American Association for Cancer Research Annual Meeting 2017, Washington, DC, USA, 1–5 April 2017. Cancer Res. 2017, 77 (Suppl. S13), 3933. [Google Scholar] [CrossRef]
- Li, H.; Yang, L.L.; Wu, C.C.; Xiao, Y.; Mao, L.; Chen, L.; Zhang, W.F.; Sun, Z.J. Expression and Prognostic Value of IFIT1 and IFITM3 in Head and Neck Squamous Cell Carcinoma. Am. J. Clin. Pathol. 2020, 153, 618–629. [Google Scholar] [CrossRef]
- Pidugu, V.K.; Wu, M.M.; Pidugu, H.B.; Lee, T.C. IFIT1 and IFIT3 modulate the drug response in human oral squamous cell carcinoma through interaction and activation of Hsp90. Abstract 2098 in Proceedings of the American Association for Cancer Research Annual Meeting 2019, Atlanta, GA, USA, 29 March–3 April 2019. Cancer Res. 2019, 79 (Suppl. S13), 2098. [Google Scholar] [CrossRef]
- Pidugu, V.; Pidugu, H.; Teng, Y.-H.; Lee, T.C. IFIT1 and IFIT3 function as Hsp90 co-chaperones to modulate the drug response in human oral squamous cell carcinoma. Abstract B034 in Proceedings of the American Association for Cancer Research Annual Meeting, Boston, MA, USA, 26–30 October 2019. Mol. Cancer Ther. 2019, 18 (Suppl. S12), B034. [Google Scholar] [CrossRef]
- Pidugu, V.K.; Wu, M.M.; Pidugu, H.B.; Lee, T.C. IFIT1 and 3 modulate the drug sensitivity in human oral squamous cell carcinoma cells. Abstract A135 in Proceedings of the American Association for Cancer Research Annual Meeting 2017 Philadelphia, PA, USA 26-30 October 2017. Mol. Cancer Ther. 2018, 17 (Suppl. S1), A135. [Google Scholar] [CrossRef]
- Danish, H.H.; Goyal, S.; Taunk, N.K.; Wu, H.; Moran, M.S.; Haffty, B.G. Interferon-induced protein with tetratricopeptide repeats 1 (IFIT1) as a prognostic marker for local control in T1-2 N0 breast cancer treated with breast-conserving surgery and radiation therapy (BCS + RT). Breast J. 2013, 19, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Weichselbaum, R.R.; Ishwaran, H.; Yoon, T.; Nuyten, D.S.; Baker, S.W.; Khodarev, N.; Su, A.W.; Shaikh, A.Y.; Roach, P.; Kreike, B.; et al. An interferon-related gene signature for DNA damage resistance is a predictive marker for chemotherapy and radiation for breast cancer. Proc. Natl. Acad. Sci. USA 2008, 105, 18490–18495. [Google Scholar] [CrossRef]
- Ghufran, S.M.; Sharma, P.; Roy, B.; Jaiswal, S.; Aftab, M.; Sengupta, S.; Ghose, S.; Biswas, S. Transcriptome wide functional analysis of HBx expressing human hepatocytes stimulated with endothelial cell cross-talk. Genomics 2023, 115, 110642. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, Y.; Hou, J.; Bai, C.; Li, Z.; Fan, J.; Ng, I.O.L.; Zhou, W.; Sun, H.; Dong, Q.; et al. Hepatic IFIT3 predicts interferon-alpha therapeutic response in patients of hepatocellular carcinoma. Hepatology 2017, 66, 152–166. [Google Scholar] [CrossRef]
- Liu, G.; Sun, J.; Yang, Z.F.; Zhou, C.; Zhou, P.Y.; Guan, R.Y.; Sun, B.Y.; Wang, Z.T.; Zhou, J.; Fan, J.; et al. Cancer-associated fibroblast-derived CXCL11 modulates hepatocellular carcinoma cell migration and tumor metastasis through the circUBAP2/miR-4756/IFIT1/3 axis. Cell Death Dis. 2021, 12, 260. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Y.; Lu, W.; Sun, R.; Guo, R.; Cao, X.; Liu, X.; Lyu, C.; Zhao, M. The diagnostic/prognostic roles and biological function of the IFIT family members in acute myeloid leukemia. BMC Med. Genom. 2023, 16, 296. [Google Scholar] [CrossRef]
- Perez-Diez, I.; Andreu, Z.; Hidalgo, M.R.; Perpina-Clerigues, C.; Fantin, L.; Fernandez-Serra, A.; de la Iglesia-Vaya, M.; Lopez-Guerrero, J.A.; Garcia-Garcia, F. A Comprehensive Transcriptional Signature in Pancreatic Ductal Adenocarcinoma Reveals New Insights into the Immune and Desmoplastic Microenvironments. Cancers 2023, 15, 2887. [Google Scholar] [CrossRef]
- Nakamura, H.; Watanabe, M.; Takada, K.; Sato, T.; Hikage, F.; Umetsu, A.; Muramatsu, J.; Furuhashi, M.; Ohguro, H. Modulation of Epithelial-Mesenchymal Transition Is a Possible Underlying Mechanism for Inducing Chemoresistance in MIA PaCa-2 Cells against Gemcitabine and Paclitaxel. Biomedicines 2024, 12, 1011. [Google Scholar] [CrossRef]
- Su, W.; Xiao, W.; Chen, L.; Zhou, Q.; Zheng, X.; Ju, J.; Jiang, J.; Wang, Z. Decreased IFIT2 Expression in Human Non-Small-Cell Lung Cancer Tissues Is Associated with Cancer Progression and Poor Survival of the Patients. OncoTargets Ther. 2019, 12, 8139–8149. [Google Scholar] [CrossRef]
- Chen, L.; Zhai, W.; Zheng, X.; Xie, Q.; Zhou, Q.; Tao, M.; Zhu, Y.; Wu, C.; Jiang, J. Decreased IFIT2 Expression Promotes Gastric Cancer Progression and Predicts Poor Prognosis of the Patients. Cell. Physiol. Biochem. 2018, 45, 15–25. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Y.; Zhu, Y.; Chen, Y.; Feng, J.; Jiang, T.; Zheng, X.; Chen, L.; Jiang, J. STAT1/IFIT2 signaling pathway is involved in PD-L1-mediated epithelial-to-mesenchymal transition in human esophageal cancer. Clin. Transl. Oncol. 2022, 24, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Wang, Y.; Ma, Z.; Yang, R.; Liang, S.; Zhang, M.; Song, S.; Li, S.; Liu, G.; Fan, D.; et al. MicroRNA-645, up-regulated in human adencarcinoma of gastric esophageal junction, inhibits apoptosis by targeting tumor suppressor IFIT2. BMC Cancer 2014, 14, 633. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.G.; Hu, B.; Sun, H.X.; Sun, Q.M.; Sun, C.; Fu, P.Y.; Yang, Z.F.; Zhang, X.; Zhou, C.H.; Fan, J.; et al. Long non-coding RNA00364 represses hepatocellular carcinoma cell proliferation via modulating p-STAT3-IFIT2 signaling axis. Oncotarget 2017, 8, 102006–102019. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Zhan, M.; Zhang, Y.; Huang, S.; Xu, S.; Huang, X.; He, M.; Yao, Y.; Man, M.; Wang, J. PLZF inhibits proliferation and metastasis of gallbladder cancer by regulating IFIT2. Cell Death Dis. 2018, 9, 71. [Google Scholar] [CrossRef]
- Stawowczyk, M.; Van Scoy, S.; Kumar, K.P.; Reich, N.C. The interferon stimulated gene 54 promotes apoptosis. J. Biol. Chem. 2011, 286, 7257–7266. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, N.; Liu, S.; Jiang, M.; Wan, J.; Zhang, Y.; Wan, L.; Xie, C.; Le, A. Overexpression of IFIT2 inhibits the proliferation of chronic myeloid leukemia cells by regulating the BCR-ABL/AKT/mTOR pathway. Int. J. Mol. Med. 2020, 45, 1187–1194. [Google Scholar] [CrossRef]
- Zhang, Y.; Kong, Y.; Liu, S.; Zeng, L.; Wan, L.; Zhang, Z. Curcumin induces apoptosis in human leukemic cell lines through an IFIT2-dependent pathway. Cancer Biol. Ther. 2017, 18, 43–50. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.; Zheng, X.; Zhong, W.; Tian, X.; Yin, B.; Tian, K.; Zhang, W. Long non-coding RNA LINC00161 sensitises osteosarcoma cells to cisplatin-induced apoptosis by regulating the miR-645-IFIT2 axis. Cancer Lett. 2016, 382, 137–146. [Google Scholar] [CrossRef]
- Zhou, S.; Sun, Y.; Chen, T.; Wang, J.; He, J.; Lyu, J.; Shen, Y.; Chen, X.; Yang, R. The Landscape of the Tumor Microenvironment in Skin Cutaneous Melanoma Reveals a Prognostic and Immunotherapeutically Relevant Gene Signature. Front. Cell Dev. Biol. 2021, 9, 739594. [Google Scholar] [CrossRef]
- Gao, L.; Xiong, D.D.; He, R.Q.; Lai, Z.F.; Liu, L.M.; Huang, Z.G.; Yang, X.; Wu, H.Y.; Yang, L.H.; Ma, J.; et al. Identifying TF-miRNA-mRNA regulatory modules in nitidine chloride treated HCC xenograft of nude mice. Am. J. Transl. Res. 2019, 11, 7503–7522. [Google Scholar]
- Ohsugi, T.; Yamaguchi, K.; Zhu, C.; Ikenoue, T.; Furukawa, Y. Decreased expression of interferon-induced protein 2 (IFIT2) by Wnt/beta-catenin signaling confers anti-apoptotic properties to colorectal cancer cells. Oncotarget 2017, 8, 100176–100186. [Google Scholar] [CrossRef] [PubMed]
- Ohsugi, T.; Yamaguchi, K.; Zhu, C.; Ikenoue, T.; Takane, K.; Shinozaki, M.; Tsurita, G.; Yano, H.; Furukawa, Y. Anti-apoptotic effect by the suppression of IRF1 as a downstream of Wnt/beta-catenin signaling in colorectal cancer cells. Oncogene 2019, 38, 6051–6064. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, S.; Xu, F.; Kong, Y.; Wan, L.; Zhang, Y.; Zhang, Z. Inhibition of Proteasome Activity Induces Aggregation of IFIT2 in the Centrosome and Enhances IFIT2-Induced Cell Apoptosis. Int. J. Biol. Sci. 2017, 13, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Song, L.; Cong, Q.; Wang, J.; Xu, H.; Chu, Y.; Li, Q.; Zhang, Y.; Zou, X.; Zhang, C.; et al. The LIM protein AJUBA promotes colorectal cancer cell survival through suppression of JAK1/STAT1/IFIT2 network. Oncogene 2017, 36, 2655–2666. [Google Scholar] [CrossRef]
- Xu, Q.C.; Tien, Y.C.; Shi, Y.H.; Chen, S.; Zhu, Y.Q.; Huang, X.T.; Huang, C.S.; Zhao, W.; Yin, X.Y. METTL3 promotes intrahepatic cholangiocarcinoma progression by regulating IFIT2 expression in an m(6)A-YTHDF2-dependent manner. Oncogene 2022, 41, 1622–1633. [Google Scholar] [CrossRef]
- Xu, B.; Zhu, Y.L.; Fan, J.L.; Chen, L.J.; Jiang, J.T. Clinical significance of IFIT2 expression in human renal cancer tissues. Transl. Cancer Res. 2020, 9, 3214–3221. [Google Scholar] [CrossRef]
- Ognibene, M.; De Marco, P.; Amoroso, L.; Cangelosi, D.; Zara, F.; Parodi, S.; Pezzolo, A. Multiple Genes with Potential Tumor Suppressive Activity Are Present on Chromosome 10q Loss in Neuroblastoma and Are Associated with Poor Prognosis. Cancers 2023, 15, 2035. [Google Scholar] [CrossRef]
- Lai, K.C.; Chang, K.W.; Liu, C.J.; Kao, S.Y.; Lee, T.C. IFN-induced protein with tetratricopeptide repeats 2 inhibits migration activity and increases survival of oral squamous cell carcinoma. Mol. Cancer Res. 2008, 6, 1431–1439. [Google Scholar] [CrossRef]
- Lai, K.C.; Liu, C.J.; Chang, K.W.; Lee, T.C. Depleting IFIT2 mediates atypical PKC signaling to enhance the migration and metastatic activity of oral squamous cell carcinoma cells. Oncogene 2013, 32, 3686–3697. [Google Scholar] [CrossRef]
- Lai, K.C.; Liu, C.J.; Lin, T.J.; Mar, A.C.; Wang, H.H.; Chen, C.W.; Hong, Z.X.; Lee, T.C. Blocking TNF-alpha inhibits angiogenesis and growth of IFIT2-depleted metastatic oral squamous cell carcinoma cells. Cancer Lett. 2016, 370, 207–215. [Google Scholar] [CrossRef]
- Lai, K.C.; Regmi, P.; Liu, C.J.; Lo, J.F.; Lee, T.C. IFIT2 Depletion Promotes Cancer Stem Cell-like Phenotypes in Oral Cancer. Biomedicines 2023, 11, 896. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.C.; Hong, Z.X.; Hsieh, J.G.; Lee, H.J.; Yang, M.H.; Hsieh, C.H.; Yang, C.H.; Chen, Y.R. IFIT2-depleted metastatic oral squamous cell carcinoma cells induce muscle atrophy and cancer cachexia in mice. J. Cachexia Sarcopenia Muscle 2022, 13, 1314–1328. [Google Scholar] [CrossRef] [PubMed]
- Regmi, P.; Lai, K.C.; Liu, C.J.; Lee, T.C. SAHA Overcomes 5-FU Resistance in IFIT2-Depleted Oral Squamous Cell Carcinoma Cells. Cancers 2020, 12, 3527. [Google Scholar] [CrossRef] [PubMed]
- Jing, D.; Zhou, W.; Shen, L.; Zhang, Q.; Xie, W.T.; Shen, E.; Li, Z.; Shen, L.F.; Sun, L.Q. RIG-I promotes IFN/JAK2 expression and the endoplasmic reticulum stress response to inhibit chemoradiation resistance in nasopharyngeal carcinoma. Cancer Med. 2019, 8, 6344–6357. [Google Scholar] [CrossRef]
- Zenga, J.; Awan, M.J.; Frei, A.; Massey, B.; Bruening, J.; Shukla, M.; Sharma, G.P.; Shreenivas, A.; Wong, S.J.; Zimmermann, M.T.; et al. Type I interferon signaling promotes radioresistance in head and neck cancer. Transl. Cancer Res. 2024, 13, 2535–2543. [Google Scholar] [CrossRef]
- Koh, S.Y.; Moon, J.Y.; Unno, T.; Cho, S.K. Baicalein Suppresses Stem Cell-Like Characteristics in Radio- and Chemoresistant MDA-MB-231 Human Breast Cancer Cells through Up-Regulation of IFIT2. Nutrients 2019, 11, 624. [Google Scholar] [CrossRef]
- Gao, C.; Li, L.; Jin, X.; Song, X.; Li, H.; Xu, X.; Dong, C.; Ma, B. The Involvement of Insulin-Like Growth Factor 2 Messenger Ribonucleic Acid-Binding Protein 2 in the Regulation of the Expression of Breast Cancer-Related Genes. Breast Cancer 2022, 14, 311–322. [Google Scholar] [CrossRef]
- Liu, P.; Kong, X.; Yi, S.; Chen, Y.; Luo, W. IFIT3 accelerates the progression of head and neck squamous cell carcinoma by targeting PD-L1 to activate PI3K/AKT signaling pathway. World J. Surg. Oncol. 2024, 22, 34. [Google Scholar] [CrossRef]
- Niess, H.; Camaj, P.; Mair, R.; Renner, A.; Zhao, Y.; Jackel, C.; Nelson, P.J.; Jauch, K.W.; Bruns, C.J. Overexpression of IFN-induced protein with tetratricopeptide repeats 3 (IFIT3) in pancreatic cancer: Cellular “pseudoinflammation” contributing to an aggressive phenotype. Oncotarget 2015, 6, 3306–3318. [Google Scholar] [CrossRef]
- Chai, B.; Li, Y.; Guo, Y.; Zhang, Z.; Jia, K.; Chai, X.; Suo, Y. ETV7 promotes colorectal cancer progression through upregulation of IFIT3. Funct. Integr. Genom. 2024, 24, 8. [Google Scholar] [CrossRef]
- Naranjo, N.M.; Salem, I.; Harris, M.A.; Languino, L.R. IFIT3 (interferon induced protein with tetratricopeptide repeats 3) modulates STAT1 expression in small extracellular vesicles. Biochem. J. 2021, 478, 3905–3921. [Google Scholar] [CrossRef] [PubMed]
- Thakore, V.P.; Patel, K.D.; Vora, H.H.; Patel, P.S.; Jain, N.K. Up-regulation of extracellular-matrix and inflammation related genes in oral squamous cell carcinoma. Arch. Oral Biol. 2024, 161, 105925. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Wang, X.; Liu, W.; Ji, P.; Shang, A.; Wu, J.; Zhou, H.; Quan, W.; Yao, Y.; Yang, Y.; et al. Novel evidence for retinoic acid-induced G (Rig-G) as a tumor suppressor by activating p53 signaling pathway in lung cancer. FASEB J. 2020, 34, 11900–11912. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Zhang, Q.; Xuan, Y.; Ou, Z.; Yu, Q.; Zhu, D.; Xu, E. The expression and prognostic value of IFIT3 in esophageal squamous cell carcinoma. Transl. Cancer Res. 2024, 13, 6219–6234. [Google Scholar] [CrossRef]
- Zhao, Y.; Altendorf-Hofmann, A.; Pozios, I.; Camaj, P.; Daberitz, T.; Wang, X.; Niess, H.; Seeliger, H.; Popp, F.; Betzler, C.; et al. Elevated interferon-induced protein with tetratricopeptide repeats 3 (IFIT3) is a poor prognostic marker in pancreatic ductal adenocarcinoma. J. Cancer Res. Clin. Oncol. 2017, 143, 1061–1068. [Google Scholar] [CrossRef]
- Li, H.; Zhou, J.; Zhou, L.; Zhang, X.; Shang, J.; Feng, X.; Yu, L.; Fan, J.; Ren, J.; Zhang, R.; et al. Identification of the shared gene signatures and molecular pathways in systemic lupus erythematosus and diffuse large B-cell lymphoma. J. Gene Med. 2023, 25, e3558. [Google Scholar] [CrossRef]
- Zhu, H.R.; Wei, Y.B.; Guo, J.Q.; Liu, X.F. Double-negative T cells with a distinct transcriptomic profile are abundant in the peripheral blood of patients with breast cancer. Breast Cancer Res. Treat. 2025, 209, 103–115. [Google Scholar] [CrossRef]
- Wang, Z.; Tu, L.; Chen, M.; Tong, S. Identification of a tumor microenvironment-related seven-gene signature for predicting prognosis in bladder cancer. BMC Cancer 2021, 21, 692. [Google Scholar] [CrossRef]
- Zhen, J.; Song, Z.; Su, W.; Zeng, Q.C.; Li, J.; Sun, Q. Integrated analysis of RNA-binding proteins in thyroid cancer. PLoS ONE 2021, 16, e0247836. [Google Scholar] [CrossRef]
- Thakore, V.P.; Patel, K.D.; Bhadresha, K.P.; Patel, P.S.; Jain, N.K. An integrative analysis to enumerate candidate genes for clinical use in oral cancer. J. Cancer Res. Ther. 2022, 18, S182–S190. [Google Scholar] [CrossRef]
- Korpela, S.P.; Hinz, T.K.; Oweida, A.; Kim, J.; Calhoun, J.; Ferris, R.; Nemenoff, R.A.; Karam, S.D.; Clambey, E.T.; Heasley, L.E. Role of epidermal growth factor receptor inhibitor-induced interferon pathway signaling in the head and neck squamous cell carcinoma therapeutic response. J. Transl. Med. 2021, 19, 43. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Zhang, Q.; Su, Y.; Pan, L. Network-Based Differential Analysis to Identify Molecular Features of Tumorigenesis for Esophageal Squamous Carcinoma. Molecules 2018, 23, 88. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Zhang, B.; Zhang, C.; Kwong, D.L.; Chang, Z.; Li, S.; Wang, Z.; Han, H.; Li, J.; Zhong, Y.; et al. Single-Cell Transcriptomic Analysis of Primary and Metastatic Tumor Ecosystems in Esophageal Squamous Cell Carcinoma. Adv. Sci. 2023, 10, e2204565. [Google Scholar] [CrossRef]
- Popp, M.C.; Klippstein, M.; Lohneis, P.; Kalinski, T.; Li, J.; Quaas, A.; Bludau, M.; Wang, Z.; Waldschmidt, D.; Kunzmann, V.; et al. Interferon-Induced Protein With Multiple Tetratricopeptide Repeats 3 Is Associated With Response to Chemotherapy and Recurrence but Not With Survival. Pancreas 2020, 49, 1307–1314. [Google Scholar] [CrossRef]
- Van den Broeck, A.; Vankelecom, H.; Van Eijsden, R.; Govaere, O.; Topal, B. Molecular markers associated with outcome and metastasis in human pancreatic cancer. J. Exp. Clin. Cancer Res. 2012, 31, 68. [Google Scholar] [CrossRef]
- Hu, H.; Li, B.; Chen, H.; Fan, G.; Ye, Z.; Ji, S.; Yu, X.; Xu, X.; Qin, Y. NMI promotes tumor progression and gemcitabine resistance in pancreatic cancer via STAT3-IFIT3 axis. Mol. Carcinog. 2024, 63, 195–208. [Google Scholar] [CrossRef]
- Wang, Z.; Qin, J.; Zhao, J.; Li, J.; Li, D.; Popp, M.; Popp, F.; Alakus, H.; Kong, B.; Dong, Q.; et al. Inflammatory IFIT3 renders chemotherapy resistance by regulating post-translational modification of VDAC2 in pancreatic cancer. Theranostics 2020, 10, 7178–7192. [Google Scholar] [CrossRef]
- Wang, F.; Tian, J.; Pang, L.; Wu, J.; Shang, A.; Sun, Z.; Li, D.; Yan, J.; Quan, W. Retinoic Acid-Induced Gene G(RIG-G) as a Novel Monitoring Biomarker in Leukemia and Its Clinical Applications. Genes 2021, 12, 1035. [Google Scholar] [CrossRef]
- Takashima, Y.; Hamano, M.; Yoshii, K.; Hayano, A.; Fukai, J.; Iwadate, Y.; Kajiwara, K.; Hondoh, H.; Yamanaka, R. Reciprocal expression of the immune response genes CXCR3 and IFI44L as module hubs are associated with patient survivals in primary central nervous system lymphoma. Int. J. Clin. Oncol. 2023, 28, 468–481. [Google Scholar] [CrossRef]
- Cheung, C.H.Y.; Cheng, C.K.; Leung, K.T.; Zhang, C.; Ho, C.Y.; Luo, X.; Kam, A.Y.F.; Xia, T.; Wan, T.S.K.; Pitts, H.A.; et al. C-terminal binding protein 2 is a novel tumor suppressor targeting the MYC-IRF4 axis in multiple myeloma. Blood Adv. 2024, 8, 2217–2234. [Google Scholar] [CrossRef]
- Wu, H.; Qin, J.; Zhao, Q.; Lu, L.; Li, C. Microdissection of the Bulk Transcriptome at Single-Cell Resolution Reveals Clinical Significance and Myeloid Cells Heterogeneity in Lung Adenocarcinoma. Front. Immunol. 2021, 12, 723908. [Google Scholar] [CrossRef]
- Li, D.; Sun, J.; Liu, W.; Wang, X.; Bals, R.; Wu, J.; Quan, W.; Yao, Y.; Zhang, Y.; Zhou, H.; et al. Rig-G is a growth inhibitory factor of lung cancer cells that suppresses STAT3 and NF-kappaB. Oncotarget 2016, 7, 66032–66050. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Huang, D.; Liu, Y.; Zhang, G.; Zheng, X.; Guan, B.; Chen, A.; Wu, J.; Luo, S.M.; Liu, Z.; et al. FAM210B activates STAT1/IRF9/IFIT3 axis by upregulating IFN-alpha/beta expression to impede the progression of lung adenocarcinoma. Cell Death Dis. 2025, 16, 63. [Google Scholar] [CrossRef]
- Peng, D.; Liang, M.; Li, L.; Yang, H.; Fang, D.; Chen, L.; Guan, B. Circ_BBS9 as an early diagnostic biomarker for lung adenocarcinoma: Direct interaction with IFIT3 in the modulation of tumor immune microenvironment. Front. Immunol. 2024, 15, 1344954. [Google Scholar] [CrossRef] [PubMed]
- Nushtaeva, A.A.; Stepanov, G.A.; Semenov, D.V.; Juravlev, E.S.; Balahonova, E.A.; Gerasimov, A.V.; Sidorov, S.V.; Savelyev, E.I.; Kuligina, E.V.; Richter, V.A.; et al. Characterization of primary normal and malignant breast cancer cell and their response to chemotherapy and immunostimulatory agents. BMC Cancer 2018, 18, 728. [Google Scholar] [CrossRef]
- Mares-Quinones, M.D.; Galan-Vasquez, E.; Perez-Rueda, E.; Perez-Ishiwara, D.G.; Medel-Flores, M.O.; Gomez-Garcia, M.D.C. Identification of modules and key genes associated with breast cancer subtypes through network analysis. Sci. Rep. 2024, 14, 12350. [Google Scholar] [CrossRef]
- Huang, Z.; Bu, D.; Yang, N.; Huang, W.; Zhang, L.; Li, X.; Ding, B.S. Integrated analyses of single-cell transcriptomics identify metastasis-associated myeloid subpopulations in breast cancer lung metastasis. Front. Immunol. 2023, 14, 1180402. [Google Scholar] [CrossRef]
- Huang, J.; Lo, U.G.; Wu, S.; Wang, B.; Pong, R.C.; Lai, C.H.; Lin, H.; He, D.; Hsieh, J.T.; Wu, K. The roles and mechanism of IFIT5 in bladder cancer epithelial-mesenchymal transition and progression. Cell Death Dis. 2019, 10, 437. [Google Scholar] [CrossRef]
- Lo, U.G.; Bao, J.; Cen, J.; Yeh, H.C.; Luo, J.; Tan, W.; Hsieh, J.T. Interferon-induced IFIT5 promotes epithelial-to-mesenchymal transition leading to renal cancer invasion. Am. J. Clin. Exp. Urol. 2019, 7, 31–45. [Google Scholar]
- Lo, U.G.; Pong, R.C.; Yang, D.; Gandee, L.; Hernandez, E.; Dang, A.; Lin, C.J.; Santoyo, J.; Ma, S.; Sonavane, R.; et al. IFNgamma-Induced IFIT5 Promotes Epithelial-to-Mesenchymal Transition in Prostate Cancer via miRNA Processing. Cancer Res. 2019, 79, 1098–1112. [Google Scholar] [CrossRef]
- Lou, N.; Zheng, C.; Wang, Y.; Liang, C.; Tan, Q.; Luo, R.; Zhang, L.; Xie, T.; Shi, Y.; Han, X. Identification of novel serological autoantibodies in Chinese prostate cancer patients using high-throughput protein arrays. Cancer Immunol. Immunother. 2023, 72, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Yang, L.; Liu, C.; Jiang, R.; Huang, Q.; Wang, Q.; Wu, X. A novel N7-Methylguanine-related gene signature for predicting prognosis in acute myeloid leukemia: Bioinformatic analysis and experimental verification. Hematology 2024, 29, 2433905. [Google Scholar] [CrossRef] [PubMed]
- Felipe Fumero, E.; Walter, C.; Frenz, J.M.; Seifert, F.; Alla, V.; Hennig, T.; Angenendt, L.; Hartmann, W.; Wolf, S.; Serve, H.; et al. Epigenetic control over the cell-intrinsic immune response antagonizes self-renewal in acute myeloid leukemia. Blood 2024, 143, 2284–2299. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, S.; Qin, W.; Wang, Y.; Li, L.; Li, Q.; Yuan, X. A Novel RNA Binding Protein-Related Prognostic Signature for Hepatocellular Carcinoma. Front. Oncol. 2020, 10, 580513. [Google Scholar] [CrossRef]
- Xiong, D.; Yang, J.; Li, D.; Wang, J. Exploration of Key Immune-Related Transcriptomes Associated with Doxorubicin-Induced Cardiotoxicity in Patients with Breast Cancer. Cardiovasc. Toxicol. 2023, 23, 329–348. [Google Scholar] [CrossRef]
- Zhang, X.; Miao, Y.; Sun, H.W.; Wang, Y.X.; Zhao, W.M.; Pang, A.Y.; Wu, X.Y.; Shen, C.C.; Chen, X.D. Integrated analysis from multi-center studies identities m7G-derived modification pattern and risk stratification system in skin cutaneous melanoma. Front. Immunol. 2022, 13, 1034516. [Google Scholar] [CrossRef]
- Deng, J.; Lin, J.; Liu, C.; Li, J.; Cai, J.; Zhou, X.; Li, X. N7-methylguanosine methylation-related regulator genes as biological markers in predicting prognosis for melanoma. Sci. Rep. 2022, 12, 21082. [Google Scholar] [CrossRef]
- Xiao, S.; Li, D.; Zhu, H.Q.; Song, M.G.; Pan, X.R.; Jia, P.M.; Peng, L.L.; Dou, A.X.; Chen, G.Q.; Chen, S.J.; et al. RIG-G as a key mediator of the antiproliferative activity of interferon-related pathways through enhancing p21 and p27 proteins. Proc. Natl. Acad. Sci. USA 2006, 103, 16448–16453. [Google Scholar] [CrossRef]
- von Locquenghien, M.; Rozalen, C.; Celia-Terrassa, T. Interferons in cancer immunoediting: Sculpting metastasis and immunotherapy response. J. Clin. Investig. 2021, 131, e143296. [Google Scholar] [CrossRef]
- Abdolvahab, M.H.; Darvishi, B.; Zarei, M.; Majidzadeh, A.K.; Farahmand, L. Interferons: Role in cancer therapy. Immunotherapy 2020, 12, 833–855. [Google Scholar] [CrossRef]
- Jorgovanovic, D.; Song, M.; Wang, L.; Zhang, Y. Roles of IFN-gamma in tumor progression and regression: A review. Biomark. Res. 2020, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Benci, J.L.; Xu, B.; Qiu, Y.; Wu, T.J.; Dada, H.; Twyman-Saint Victor, C.; Cucolo, L.; Lee, D.S.M.; Pauken, K.E.; Huang, A.C.; et al. Tumor Interferon Signaling Regulates a Multigenic Resistance Program to Immune Checkpoint Blockade. Cell 2016, 167, 1540–1554.e12. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Michal, J.J.; Zhang, L.; Ding, B.; Lunney, J.K.; Liu, B.; Jiang, Z. Interferon induced IFIT family genes in host antiviral defense. Int. J. Biol. Sci. 2013, 9, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Peng, C.; Jin, C.; Wang, Y.; Wang, T.; Yang, P.; Peng, W.; Sun, Q.; Xu, H.; Nie, H.; et al. PJA2 Suppresses Colorectal Cancer Progression by Controlling HDAC2 Degradation and Stability. Adv. Sci. 2025, 12, e2401964. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parsyan, A.; Kochiyanil, A.; Bonvissuto, A.C.; Bhat, V.; Allan, A.L. Clinical Utility of IFIT Proteins in Human Malignancies. Biomedicines 2025, 13, 1435. https://doi.org/10.3390/biomedicines13061435
Parsyan A, Kochiyanil A, Bonvissuto AC, Bhat V, Allan AL. Clinical Utility of IFIT Proteins in Human Malignancies. Biomedicines. 2025; 13(6):1435. https://doi.org/10.3390/biomedicines13061435
Chicago/Turabian StyleParsyan, Armen, Arpitha Kochiyanil, Anne C. Bonvissuto, Vasudeva Bhat, and Alison L. Allan. 2025. "Clinical Utility of IFIT Proteins in Human Malignancies" Biomedicines 13, no. 6: 1435. https://doi.org/10.3390/biomedicines13061435
APA StyleParsyan, A., Kochiyanil, A., Bonvissuto, A. C., Bhat, V., & Allan, A. L. (2025). Clinical Utility of IFIT Proteins in Human Malignancies. Biomedicines, 13(6), 1435. https://doi.org/10.3390/biomedicines13061435