Abstract
Background: Liver diseases are a global health issue with an annual mortality of 80,000 patients, mainly due to complications that arise during disease progression, as effective treatments are lacking. Objectives: This study evaluated the hepatoprotective effects of two derivatives of cinnamic acid, LQM717 and LQM755, in a murine model of acute liver damage induced by carbon tetrachloride (CCl4, 4 g/kg, single dose p.o.). Methods: Male Wistar rats were pretreated with five doses of LQM717 (20 mg/kg i.p.) or LQM755 (equimolar dose), starting 2 days before inducing hepatotoxic damage with CCl4. Results: The key parameters of hepatocellular function and damage showed significant increases in ALT, ALP, GGT, and total and direct bilirubin in rats intoxicated with CCl4, with decreased liver glycogen and serum albumin. Macroscopic and microscopic liver examinations revealed reduced inflammation, necrosis, and steatosis in animals pretreated with LQM717 or LQM755. Hepatomegaly was observed only in the LQM717 + CCl4 group. LQM755 statistically provided partial protection against increases in ALT and ALP and completely prevented elevations in GGT and total and direct bilirubin. LQM755 completely prevented albumin reduction, while LQM717 only partially prevented it. Both compounds partially prevented glycogen depletion. Bioinformatic analysis identified 32 potential liver protein targets for LQM717 and 36 for LQM755. Conclusions: These findings suggest that LQM717 and LQM755 have significant hepatoprotective effects against CCl4-induced acute liver injury, providing information for future studies in other acute and chronic models, as well as to elucidate their mechanisms of action.
1. Introduction
Liver diseases are responsible for more than two million deaths annually, accounting for 4% of all deaths worldwide [1,2]. Approximately 80 million individuals worldwide are estimated to be afflicted with various forms of liver disease. The economic burden of this condition is substantial, with costs amounting to EUR 27 billion in the 12 member states of the European Union in 2008, USD 19.2 billion in the United States in 2004, AUD 2.5 billion in Australia in 2010, and CNY 35 billion in China in 2006. The elevated risk of liver failure, which can be fatal, further exacerbates this situation. Furthermore, in recent years, the incidence of liver disease has increased both in the Western world and in several developing areas [3,4,5,6].
The most common liver diseases include viral hepatitis, cirrhosis, hepatocellular carcinoma (HCC), nonalcoholic fatty liver disease (NAFLD), fatty liver, hepatitis, hepatocirrhosis, hepatoma, biliary cysts, cholecystitis, and biliary cholangitis. The increasing incidence of these diseases is partly because, despite advances in the search for new treatments, effective drugs have not yet been developed. There are some treatments of HCC, such as Sorafenib, Lenvatinib, Regorafenib, Cabozantinib, and Ramucirumab, which have been shown to have antineoplastic activities. Nevertheless, liver transplantation (LT) remains the treatment of choice for patients with advanced liver damage, although it is associated with posttransplant complications and long waiting times [7,8,9,10,11,12,13,14].
Over the years, an expanding body of research has delved into the therapeutic properties of cinnamic acid derivatives (CADs). These compounds have demonstrated a broad spectrum of activities, including anti-inflammatory, antioxidant, antiviral, antiplatelet, antifungal, antibacterial, and anticancer properties. Cinnamic acid and its hydroxylated and methoxylated analogs have been found in numerous natural environments, either in their free or conjugated state, through ester links, the C1 ether of the aromatic ring, or O-glycoside links [15,16,17].
Cinnamic acid and the phenethyl ester of caffeic acid (CAPE) are intermediates and end products in many secondary metabolic pathways in plants. Many of the hydroxystyrene derivatives present in plants are generated by the hydroxylation of cinnamic acid, formic acid, and alanine. In combination with other constituent units, such as glucose, CADs are found in plants and form basic lignocellulosic structures. The p-coumarate units in CADs are used specifically in plants for the construction of lignin polymers, which are present in hardwoods, grasses, and other plants, such as mosses, in various chemical forms. Numerous CADs have been shown to have chemotherapeutic and chemoprophylactic value, such as LQM755, which has a cytotoxic effect on a human gastric adenocarcinoma cell-line [18], and LQM717, which reduces the formation of liver cancer in the resistant hepatocyte modified model in rats [19]. These products, which are consumed through plant-based diets, play important physiological roles in the human body. Furthermore, phenolic compounds derived from hydroxycinnamic acid, which are present in high concentrations in tea, play a key role in the potential health benefits of these compounds. Some specialty teas have been shown to improve liver function. Similarly, components such as cinnamic acids and hydroxystyrenes have been identified in products derived from bees and vegetable oils [20,21,22].
Several in vitro investigations have demonstrated that CADs possess the capacity to mitigate the impacts of various hepatotoxic agents [23] in experimental models, predominantly concentrated on hepatocellular carcinomas, or liver damage induced by substances such as paracetamol [24] or carbon tetrachloride (CCl4) [25]. However, studies have focused on other virus-driven cell signaling mechanisms that contribute to liver disease. For example, Li and colleagues reported that ursodeoxycholic acid–cinnamic acid hybrids inhibit inflammation Akt/NF-κB and MAPK signaling pathways [26].
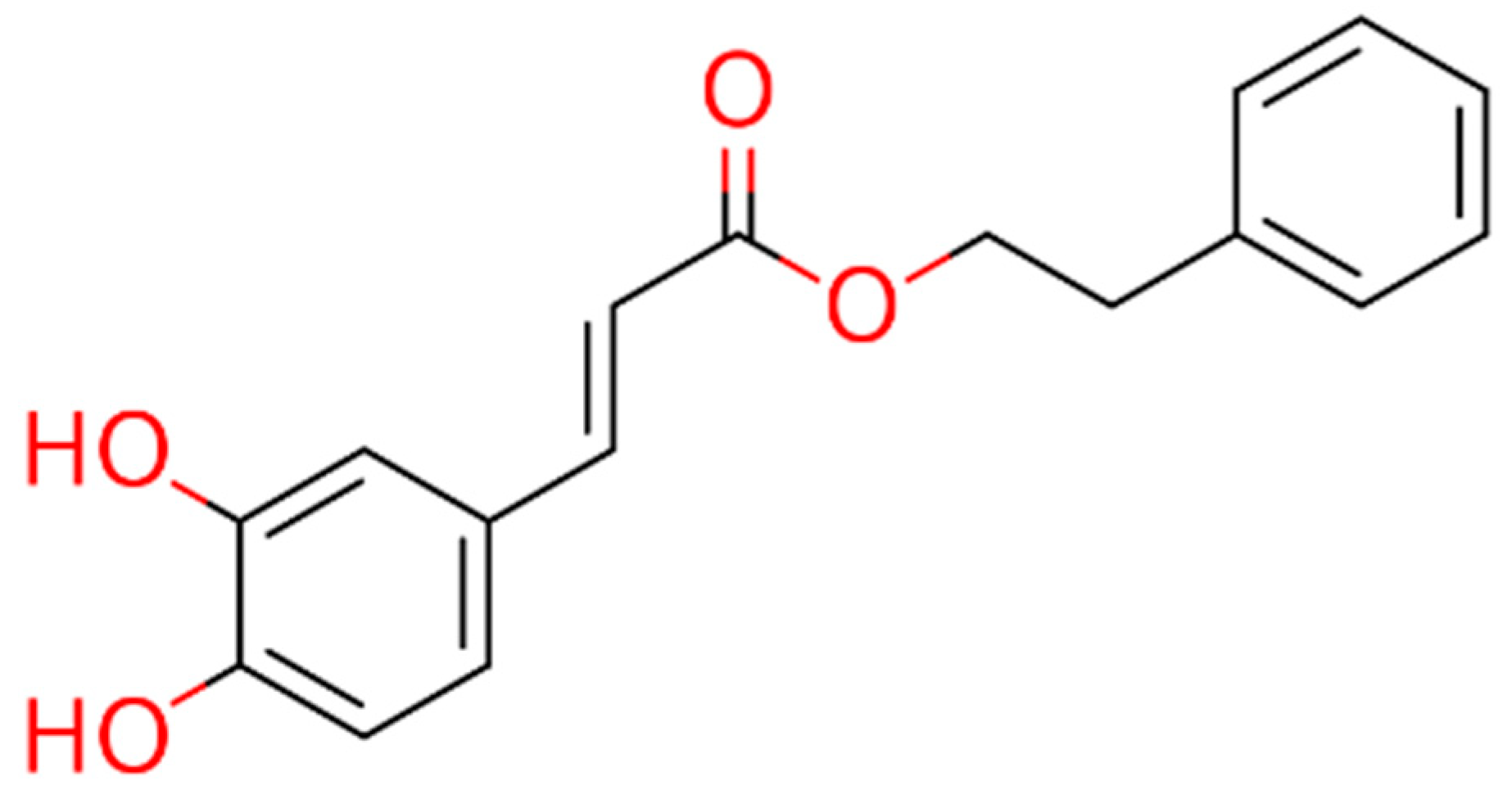
These observations prompted the idea of using cinnamic acid to generate derivatives, specifically amides, that could offer metabolic advantages over esters, such as CAPE (Figure 1), as cinnamic acid is not an esterase substrate and therefore has greater stability in biological systems.
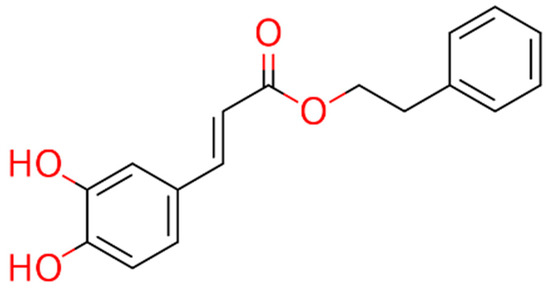
Figure 1.
Chemical structure of CAPE.
Our research group has synthesized several derivatives of cinnamic acid, reporting significant effects on gastric cancer [18,19,27] and viral infections [unpublished results]. In this work, we present the hepatoprotective activity of two derivatives, LQM717 and LQM755, which have shown promising results (Table 1).

Table 1.
Physicochemical characteristics of LQM717 and LQM755.
2. Materials and Methods
2.1. Animal Testing
Male Wistar rats (Rattus norvegicus, n = 31) weighing between 280 and 300 g were used. All experimental procedures were approved by the Research Ethics Committee of Facultad de Estudios Profesionales de Zona Huasteca, Ciudad Valles, México, and were carried out in accordance with international guidelines and technical specifications for the production, care, and use of animals established in the official Mexican standard NOM-062-ZOO-1999 [28]. The rats were maintained under controlled conditions, with a standard diet (LabDiet 5008®, PMI, St. Louis, MO, USA), free access to drinking water, a temperature of 22 ± 2 °C, a relative humidity of 50–60%, and a 12 h light–dark cycle.
2.2. Chemical Products
Chemicals
The reagents used in this investigation included carbon tetrachloride (CCl4, Malincrodt, St. Louis, MO, USA), mineral oil (Azumex, Puebla, Mexico), acetone and formaldehyde (Hycel, Jalisco, Mexico), xylene and sodium hydroxide (JT Baker, Madrid, Spain), ammonia water (CF-Industries, Deerfield, IL, USA), glycine (Santa Cruz Biotechnology, Dallas, TX, USA), potassium dichromate (Karal, Guanajuato, Mexico), basic fuchsin (Golden Bell, Jalisco, Mexico), and sodium metabisulfite (Karal, Guanajuato, Mexico). From Jalmek (NL, Mexico), hydrochloric acid, potassium hydroxide, sulfuric acid, magnesium chloride, Harris hematoxylin, yellowish eosin, and picric acid were used. Merck chemicals (Darmstadt, Germany) include p-nitrophenylphosphate, 2,4-dinitrophenylhydrazine, DL-alanine, p-nitrophenol, α-ketoglutaric acid, sodium pyruvate, p-nitroaniline, gamma-glutamyl-p-nitroanilide, dextrose, anthrone, phosphate disodium, monosodium phosphate, ketamine, xylazine, silane (silicon hydride), dimethylsulfoxide (DMSO, ACS reagent ≥ 99.9%), and Entellan.
2.3. Preparation of LQM717 and LQM755
2.3.1. Experimental Section
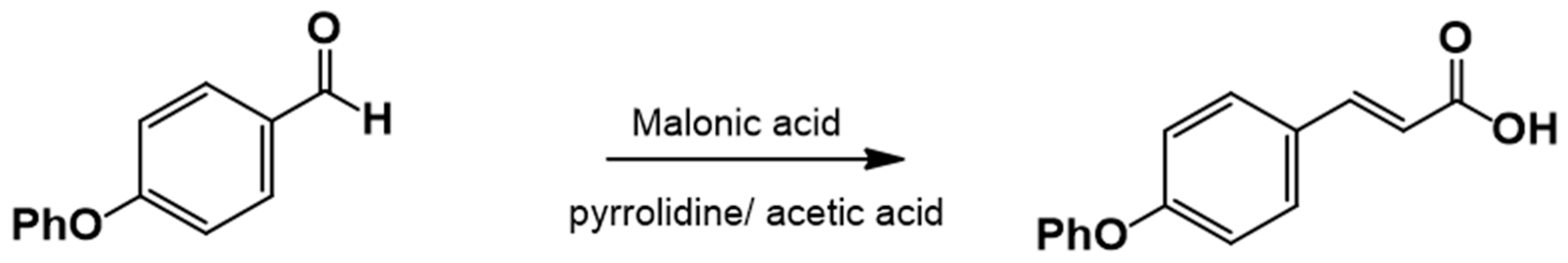
The cinnamic acid analog 4-phenoxycinnamic acid, which is not commercially available, was synthesized via Knövenagel–Döbner condensation using piperidine as a catalyst and glacial acetic acid as a solvent. Phenylacetic acid was the only reagent purchased from Sigma-Aldrich Co, SLM, St. Louis, MO, USA.
2.3.2. Procedure for the Synthesis of 3-(4-Phenoxyphenyl)-2-Propenoic Acid
Equimolar amounts of 4-Phenoxybenzaldehyde and malonic acid were placed in a round-bottom flask along with piperidine (1.25 equiv.) and glacial acetic acid (1.77 equiv.) (Figure 2). The reaction mixture was heated under reflux at 140–160 °C and monitored by TLC (hexane:EtOAc, 80:20). Upon completion, the reaction mixture was poured into ice-cold water to precipitate the product, which was then filtered and washed with water (3 × 100 mL). The crude product was recrystallized from EtOAc. If precipitation did not occur, the reaction mixture was extracted with EtOAc (5 × 50 mL), washed with brine (3 × 30 mL), dried over anhydrous sodium sulfate, and recrystallized.
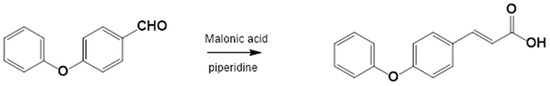
Figure 2.
3-(4-Phenoxyphenyl)-2-propenoic acid. Yield: 85%; mp 156–157 °C. IR (diamond, cm−1): νmax 1588, 3031, 3200–2200. 1H NMR (300 MHz, CDCl3): δ 6.30 (1H, d), 6.90 (5H, m), 7.24 (4H, m), 7.64 (1H, d, J = 15.6 Hz), 8.95 (1H, s). 13C NMR (75 MHz, CDCl3): δ 116.02, 117.02, 117.32, 121.61, 125.69, 128.21, 128.91, 144.64, 155.32, 156.13, 169.64.
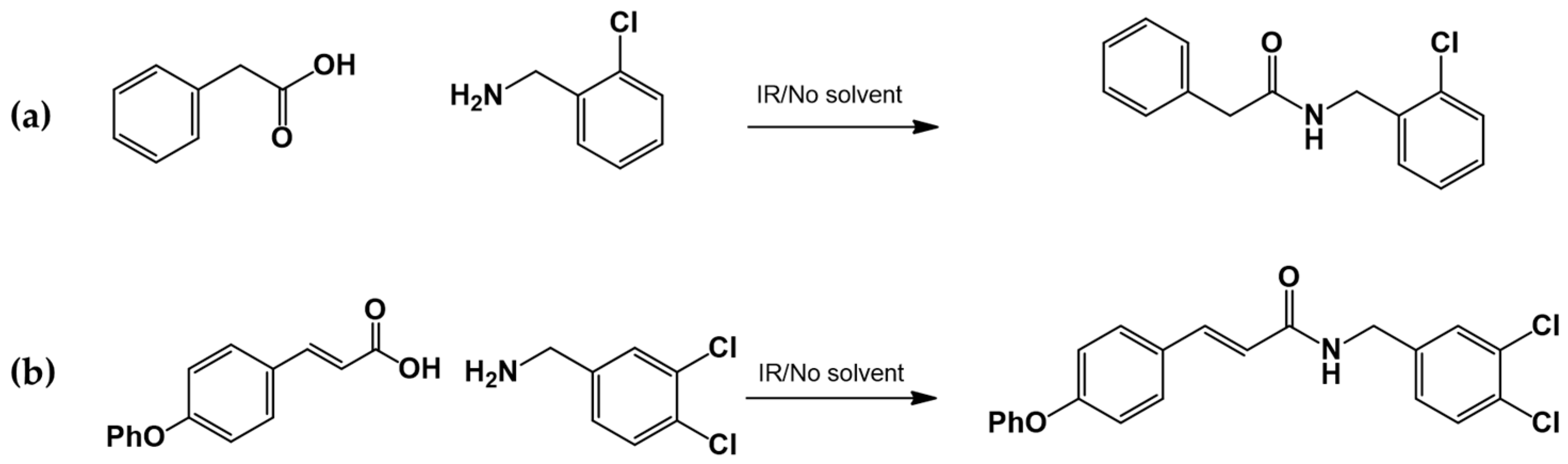
2.3.3. Amide Synthesis
The corresponding cinnamic acid and amine (1:1 molar ratio) were combined in a round-bottom flask and heated at 140–160 °C using a 300 W infrared lamp (Figure 3 and Figure 4). Reaction progress was monitored by TLC (hexane:EtOAc, 50:50). Upon completion, EtOAc was added to dissolve the product. Activated charcoal was added, and the mixture was filtered through diatomaceous earth (Hyflo Super Cel®, Merck KGaA, Darmstadt, Germany). The solvent was removed under reduced pressure, and the residue was recrystallized from EtOAc. Further purification was performed using EtOAc:hexane (90:10).
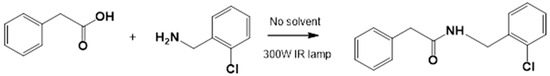
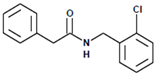
Figure 3.
LQM717—N-(2-Chlorobenzyl)-2-phenylacetamide (11). Yield: 3.9 g (79.83%); mp 118–120 °C. IR (diamond, cm−1): νmax 1546, 2923, 3274. 1H NMR (300 MHz, CDCl3, Me4Si): δ 3.63 (2H, s), 4.46 (2H, d, J = 4.0 Hz), 5.90 (1H, br s, NH), 7.29 (9H, m). 13C NMR (75 MHz, CDCl3, Me4Si): δ 42.05, 44.18, 127.44, 127.85, 129.28, 129.48, 129.88, 129.89, 130.32, 133.91, 135.05, 135.86, 171.29. Elemental analysis calcd. for C15H14ClNO: C, 69.33; H, 5.43; Cl, 13.64; N, 5.39; O, 6.16. Found: C, 69.44; H, 5.05; Cl, 13.50; N, 5.71; O, 6.22.
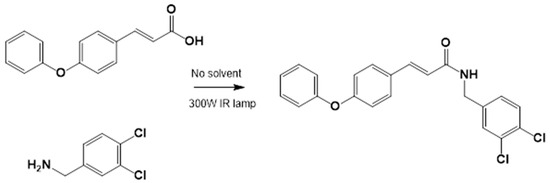
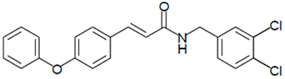
Figure 4.
LQM755.—3-(4-Phenoxy)phenyl-N-[(3,4-dichlorophenyl)methyl]prop-2-enamide. Yield: 41.98%; mp 136 °C; white solid. IR (diamond, cm−1): νmax 3400–3100 (N–H), 3043.48 (C–H sp2), 2921.22 (C–H sp3), 2478.05, 2401.64, 2328.29, 1606.98 (C=O), 1495 (C=C). 1H NMR (300 MHz, CDCl3, Me4Si): δ 8.67 (1H, t, NH), 7.57–7.56 (1H, m), 7.55–7.54 (2H, m), 7.50 (2H, d), 7.42 (1H, d, =CH), 7.40–7.37 (1H, m), 7.26–7.24 (1H, m), 7.16–7.13 (1H, m), 7.04–7.02 (2H, m), 6.96 (2H, d), 6.55 (1H, d, =CH), 4.36 (2H, d, CH2). 13C NMR (75 MHz, CDCl3, Me4Si): δ 165.79, 158.60, 156.38, 141.37, 139.13, 131.42, 131.04, 130.71, 129.79, 128.22, 130.36, 130.03, 124.60, 129.86, 121.10, 118.92, 119.82, 41.79. Elemental analysis calcd. for C22H17Cl2NO2: C, 66.30; H, 4.30; Cl, 17.80; N, 3.52; O, 8.04. Found: C, 69.35; H, 5.05; Cl, 17.60; N, 3.49; O, 8.01.
2.4. Preparation of LQM717 and LQM755 for Intraperitoneal Administration
The dose of LQM717 was 20 mg/kg, i.p., as in previous works [19], while the dose of LQM755 was equimolar to that of LQM717. In both cases, DMSO was used as a vehicle (Table 2). After their preparation, the compounds were immediately administered to each rat.
Table 2.
Intraperitoneal dosing of LQM717 and LQM755.
2.5. In Vivo Experimental Protocol
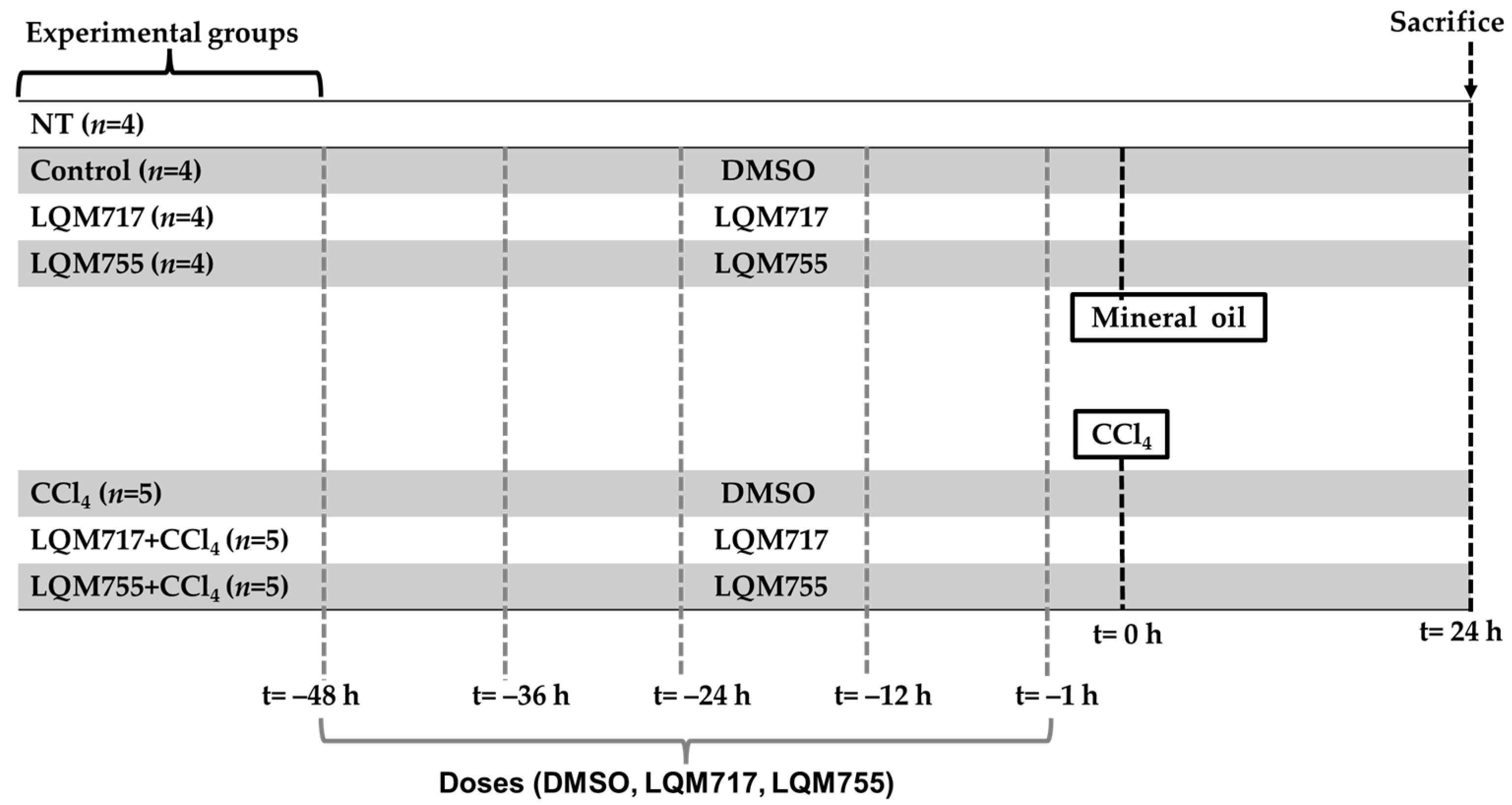
The experimental rats were randomly divided into seven groups (Figure 5):
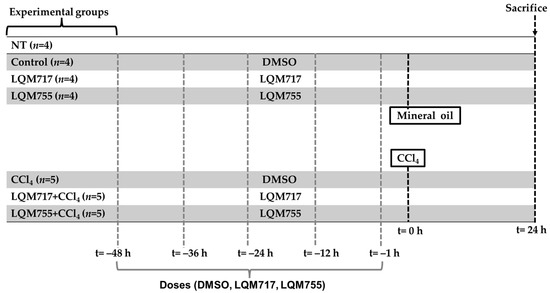
Figure 5.
Experimental groups and dosing schedule.
Healthy control groups:
- i.
- NT (untreated): Without any administration or treatment.
- ii.
- Control: Five administrations of DMSO (100 µL, i.p.) and one administration of mineral oil (250 µL/100 g, p.o.), which are the vehicles for the compounds and CCl4, respectively.
- iii.
- LQM717: Rats pretreated with five doses of LQM717 and one administration of mineral oil.
- iv.
- LQM755: Rats pretreated with five doses of LQM755 and one administration of mineral oil.
Groups with liver damage:
- v.
- CCl4: Rats were treated five times with DMSO and were intoxicated with a sublethal dose of CCl4 (4 g/kg, p.o.).
- vi.
- LQM717 + CCl4: Rats pretreated with five doses of LQM717 and intoxicated with a sublethal dose of CCl4.
- vii.
- LQM755 + CCl4: Rats pretreated with five doses of LQM755 and intoxicated with a sublethal dose of CCl4.
The animals were sacrificed 24 h after CCl4 or mineral oil administration. All the animals were weighed at the beginning of the treatments and on the day of sacrifice.
The NT (i) group was not subjected to any experimental manipulation or intervention. The Control (ii) and CCl4 (v) groups received DMSO every 12 h for 2 days (5 times) prior to the administration of mineral oil or CCl4, respectively. The LQM717 (iii) and LQM717 + CCl4 (vi) groups were pretreated with LQM717 (20 mg/kg, i.p., every 12 h for 2 days, for a total of 5 doses) before the administration of mineral oil or CCl4, respectively. The LQM755 (iv) and LQM755 + CCl4 (vii) groups were pretreated with LQM755 (equimolar dose to LQM717, i.p., every 12 h for 2 days, for a total of 5 doses) before the administration of mineral oil or CCl4, respectively.
2.6. Sacrifice of the Animals
Experimental animals were sedated with an intraperitoneal mixture of ketamine (75 mg/kg) and xylazine (10 mg/kg) and subsequently sacrificed by exsanguination via cardiac puncture. Animal death was confirmed by the absence of a heartbeat and the cessation of thermoregulation [29]. Blood samples were centrifuged at 1200× g for 10 min in a Sorvell® centrifuge (Waltham, MA, USA) to perform liver function tests on the serum.
2.7. In Situ Liver Evaluation
Macroscopic evaluation of the livers was carried out by taking photographs in situ before being extracted from the abdominal cavity. The color and texture of the livers from each experimental group were subsequently recorded.
2.8. Histochemical Procedures
2.8.1. Paraffin Embedding Tissue Samples and Sectioning
The liver was carefully removed, immediately washed with cold sterile physiological saline, weighed, and sectioned into small pieces with a scalpel. A portion of these fragments was fixed in glass bottles with a 4% solution of p-formaldehyde in phosphate buffer at pH = 7.
Liver samples fixed with p-formaldehyde were subjected to dehydration by immersion in ethanol (70%, 85%, 96%, and ≥99%, 1 h each at 60 °C) and clearance by successive immersions in absolute ethanol:xylene (1:1) and xylene for 1 h each at 60 °C. The tissues were then impregnated with liquid paraffin at 60 °C, forming a paraffin block with the tissue inside. Subsequently, 4 µm thick sections were obtained from each tissue via a microtome (ECOSHEL 202A, Pharr, TX, USA); several sections were fixed on slides previously treated with silane.
2.8.2. H&E Staining
Liver sections (4 µm) were deparaffinized at 60 °C overnight and then successively dipped in xylene, absolute ethanol:xylene (1:1), ethanol (≥99%, 96%, 80%), distilled water, hematoxylin, distilled water, acid alcohol, distilled water, ammonia water, eosin, ethanol 80%, 96%, ≥99%, absolute ethanol:xylene (1:1), and xylene. Finally, they were mounted with Entellan [30].
2.8.3. PAS Staining
Liver sections (4 µm) were fixed on slides previously treated with silane, deparaffinized at 60 °C overnight, and then dipped in xylene, absolute ethanol:xylene (1:1), ethanol (96%, 80%), distilled water, periodic acid, distilled water, Schiff’s reagent (37 °C), distilled water, hematoxylin, distilled water, ethanol (80%, 96%, and ≥99%), absolute ethanol:xylene (1:1), or xylene [30,31].
2.8.4. Biochemical Marker of Liver Necrosis
Liver damage was assessed by quantifying serum alanine aminotransferase (ALT) activity according to the specifications described by Reitman [32].
2.8.5. Biochemical Markers of Cholestasis
Biochemical markers of cholestasis were evaluated by measuring the serum activity of alkaline phosphatase (ALP) following the protocol described by Berger and Rudolph [33]. Serum gamma-glutamyl transpeptidase (GGT) activity was determined according to the instructions of the manufacturer of the commercial SPINREACT kit (γ-GT; 1001185). Additionally, direct and total bilirubin serum concentrations were measured via the SPINREACT protocol (Bilirubin T & D-SPINREACT-1001044).
2.8.6. Biochemical Markers of Liver Function
Liver function was assessed by measuring the serum albumin concentration according to the procedure indicated by the kit supplier (Albumin-SPINREACT-1001020). Furthermore, liver glycogen was assessed via the anthrone method described by Seifter and Dayton [34].
2.8.7. Statistical Analysis
The results of body/liver weights and biochemical tests were subjected to statistical analysis to assess their level of significance, which was expressed as the means ± SD, through a one-way analysis of variance (ANOVA), followed by a Tukey–Kramer test with a significance of * p < 0.05 via GraphPad Prism 8.00 software.
2.8.8. Bioinformatic Analysis
In this study, open-access bioinformatic platforms were used to predict possible protein targets in the liver that could interact with compounds LQM717 and LQM755. To achieve this, the ZINC20 page https://zinc.docking.org/ (accessed on 11 January 2024) was consulted to obtain simplified molecular input line entry system (SMILES) codes, which describe the connectivity of atoms in a molecular structure and are frequently used in bioinformatic studies [35]. With the SMILES codes of the compounds LQM717 and LQM755, possible protein targets were predicted via eight specialized online open-access platforms: SwissTarget Prediction [36] http://www.swisstargetprediction.ch (accessed on 11 January 2024), Super-PRED https://prediction.charite.de/ (accessed on 11 January 2024) [37], ChEMBL [38] https://www.ebi.ac.uk/chembl (accessed on 11 January 2024), PharmMapper [39] http://www.lilab-ecust.cn/pharmmapper/ (accessed on 11 January 2024), Pharos [40] https://pharos.nih.gov/ (accessed on 16 January 2024) the similarity ensemble approach [41] https://sea.bkslab.org/ (accessed on 16 January 2024), TargetNet [42] https://targetnet.scbdd.com (accessed on 15 January 2024), and BindingDB [43] https://www.bindingdb.org/rwd/bind/index.jsp (accessed on 16 January 2024). Possible targets whose probability value was equal to or greater than 50% (equivalent to a declared value of 0.5 on some platforms) were selected. Only those that were expressed in liver cells were subsequently selected via the Human Protein Atlas https://www.proteinatlas.org/ (accessed on 17 January 2024).
3. Results
3.1. Chemistry
3.1.1. Synthesis of 4-Phenoxy Cinnamic Acid
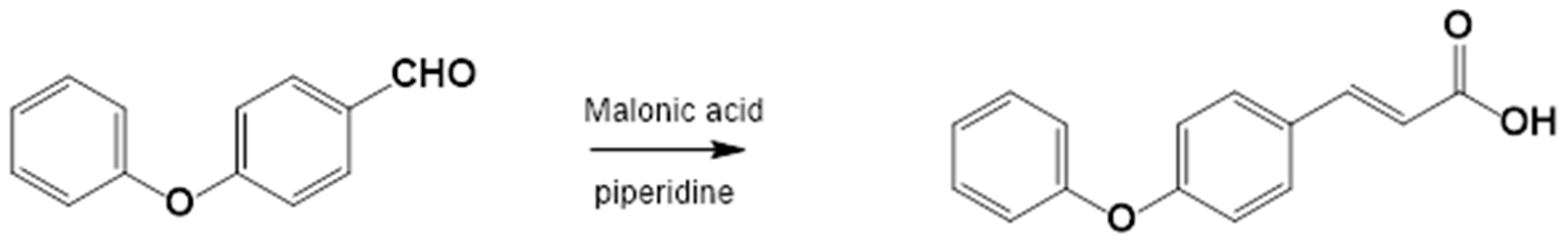
The corresponding substituted 4-phenoxybenzaldehyde and malonic acid, weighed in a 1:1 ratio, were placed in a flask with piperidine (1 mL for each part of the substituted benzaldehyde, ratio of 1.25:1) and glacial acetic acid (2.5 mL for each part of the substituted benzaldehyde, ratio of 1.77:1). This flask was connected to the reflux condenser at 140–160 °C, which is the temperature necessary to perform the reaction, followed by TLC (hexane:EtOAc, 80:20). Upon completion of the reaction, ice or cold water was added to the flask until the acid precipitated; the mixture was then filtered and washed repeatedly with water (3 × 100 mL). Prior to recrystallization, the acids were dissolved in AcOEt, and anhydrous sodium sulfate was added to eliminate residual water. Recrystallization from EtOAc produces the corresponding 4-phenoxycinnamic acid (Figure 6).
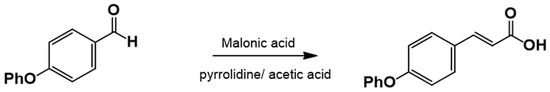
Figure 6.
Synthesis of 4-phenoxy cinnamic acid.
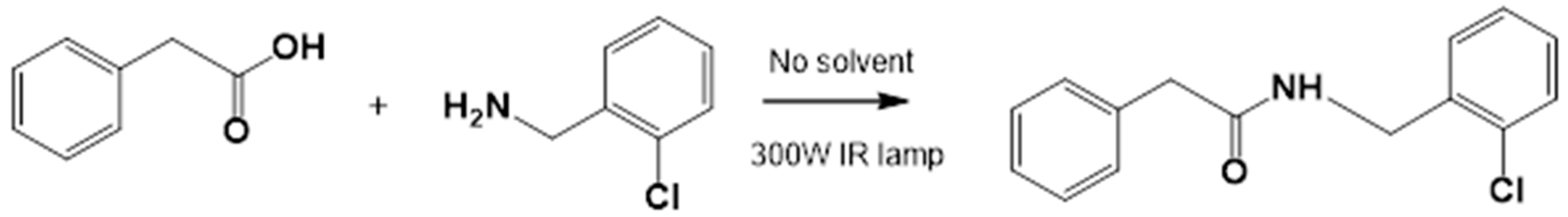
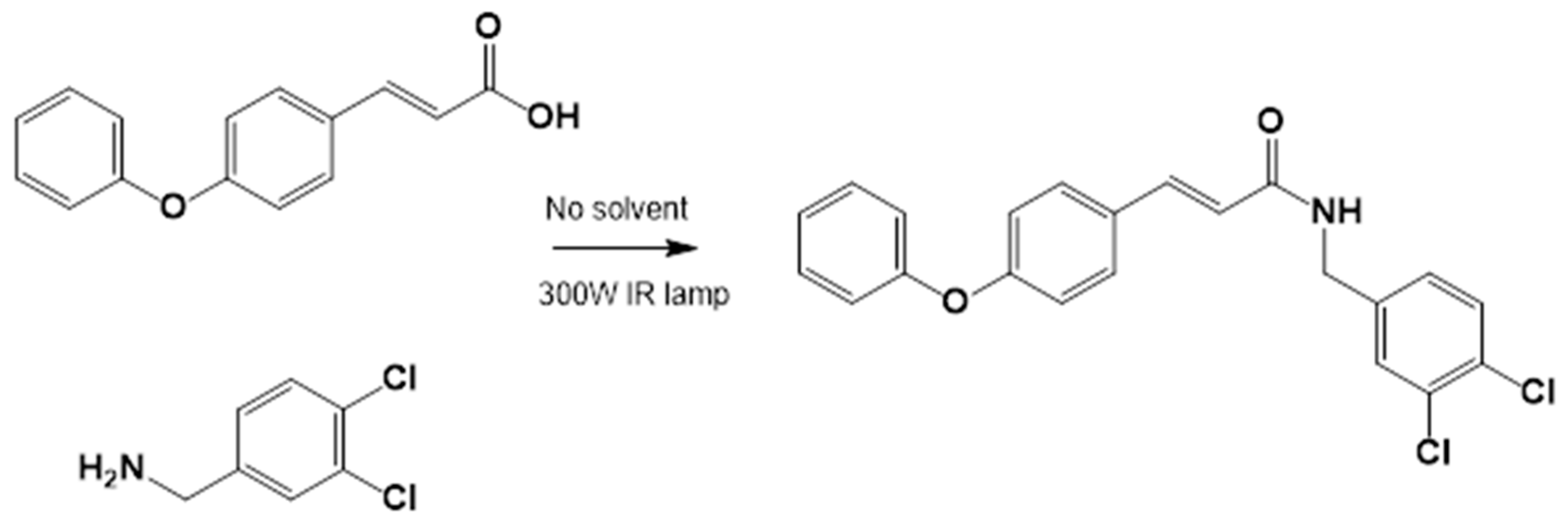
3.1.2. General Procedure for the Preparation of Amides
Acids and amines, which were weighed in a 1:1.2 ratio, were placed in a flask and connected to the reflux apparatus at 140–160 °C with a 300 W IR light bulb, followed by TLC (hexane: EtOAc, 50:50). Upon completion of the reaction, sufficient AcOEt was added to dissolve the reaction product. A small amount of activated charcoal was added to the reaction mixture and filtered over diatomaceous earth (Hyflo Super Cel® Diatomaceous Earth from Merck KGaA, Darmstadt, Germany) to remove the activated charcoal. Recrystallization from EtOAc produced the corresponding amide (Figure 7). Further recrystallization was performed with an AcOEt or an AcOEt:hexane 90:10 mixture only when necessary.
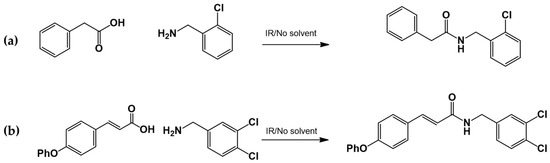
Figure 7.
Preparation of LQM717 and LQM755. (a) LQM717: N-(2-Chlorobenzyl)-2-phenylacetamide, yield (3.9 g, 79.83%); mp 118 –120 °C. IR (Diamond, cm−1): ν max 1546, 2923, 3274. 1H NMR (300 MHz; CDCl3; Me4Si) δ (ppm)3.63 (2H, s), 4.46 (2H, d, J = 4.0), 5.90 (1H, br, s, NH), 7.29 (9H, m); 13C NMR (75 MHz; CDCl3; Me4Si) δ (ppm) 42.05 (CH2), 44.18 (CH2), 127.44, 127.85, 129.28, 129.48, 129.88, 129.89, 130.32, 133.91, 135.05, 135.86 (Ph), 171.29 (-CO-). Analysis Calc. For C15H14ClNO: C, 69.3; H, 5.4; N, 5.3. Found C, 69.35; H, 5.05; N, 5.71. (b) LQM755: Yield (42%), mp 135–136 °C. IR (Diamond, cm−1): ν max 1495, 1606, 2478, 2401, 2328, 2921, 3043, 3100–3400. 1H NMR (500 MHz; DMSO-D6; Me4Si) δ (ppm) 4.35 (2H, d), 6.55 (1H, d), 6.96 (2H, d), 7.03 (2H, m), 7.14 (1H, m), 7.24 (1H, m), 7.38 (1H, m), 7.42 (1H, d), 7.50 (2H, d), 7.54 (2H, m), 7.56 (1H, m), 8.66 (1H, t). 13C NMR (125 MHz DMSO-D6; Me4Si) δ (ppm) 41.78 (-CH2-), 119.82 (-CH=), 118.92, 121.09, 124.60, 128.21, 129.85, 129.79, 130.36, 130.71, 131.04, 131.42, 139.13 (Ph), 141.36 (-CH=), 15637, 158.59 (Ph), 165.79 (C=O).
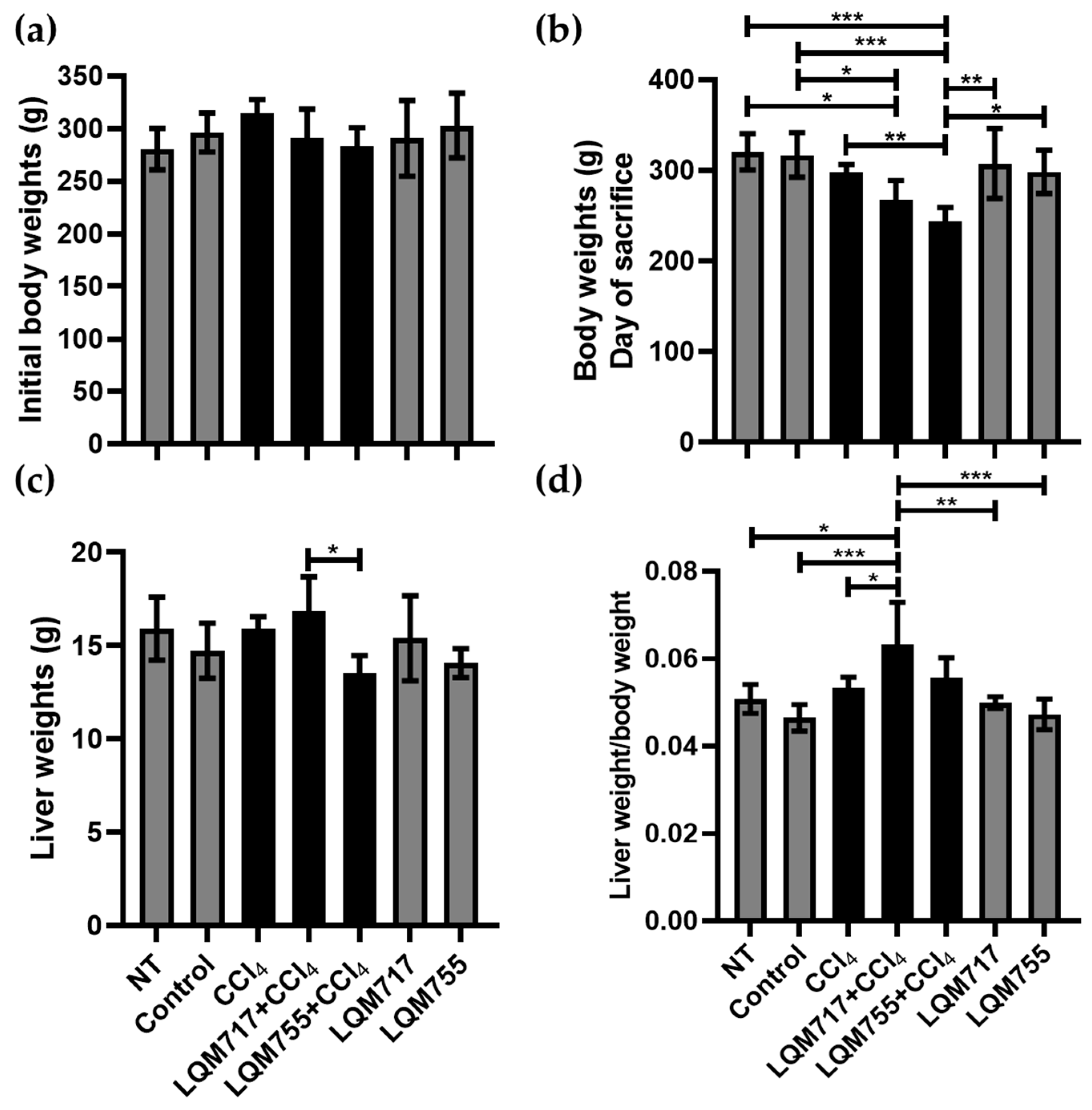
3.2. Body and Liver Weights
In the analysis of body weights at the beginning of the treatments, the averages of each experimental group did not differ significantly (Figure 8a). However, on the day of sacrifice (Figure 8b), the healthy groups (NT, Control, LQM717, and LQM755) had similar average weights without significant differences. In contrast, the average weight of the LQM717 + CCl4 group was significantly lower than that of the NT (* p < 0.05) and Control (* p < 0.05) groups. The LQM755 + CCl4 group presented a significant reduction in weight compared with the CCl4 (** p < 0.01), NT (*** p < 0.001), Control (*** p < 0.001), LQM717 (** p < 0.01), and LQM755 (* p < 0.05) groups.
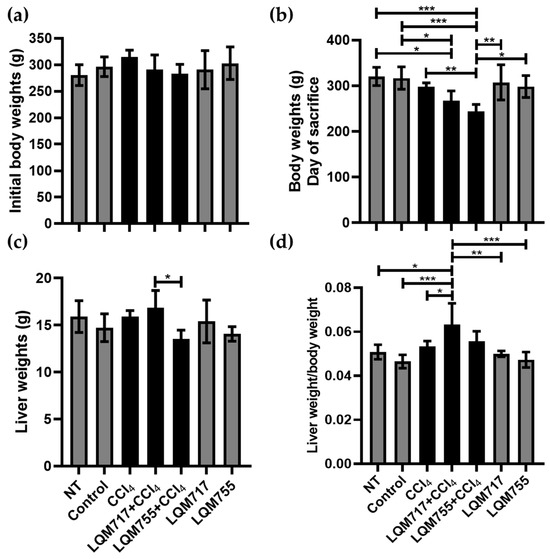
Figure 8.
Analysis of body and liver weights. (a) Body weights at the beginning of the experiment. (b) Body weights on the day of sacrifice. (c) Liver Weights after sacrifice. (d) Ratio between liver weight and body weight. Results show the average of each group ± SD. Statistically significant differences using a one-way ANOVA with a Tukey–Kramer post hoc test (* p < 0.05, ** p < 0.01, *** p < 0.001).
However, the average liver weights obtained at sacrifice were significantly lower in the LQM755 + CCl4 group in relation to the LQM717 + CCl4 group (* p < 0.05) (Figure 8c). Finally, the liver/body weight index was greater only in the LQM717 + CCl4 group than in the NT (* p < 0.05), Control (*** p < 0.001), CCl4 (* p < 0.05), and LQM717 (** p < 0.01) and LQM755 (*** p < 0.001) groups (Figure 8d).
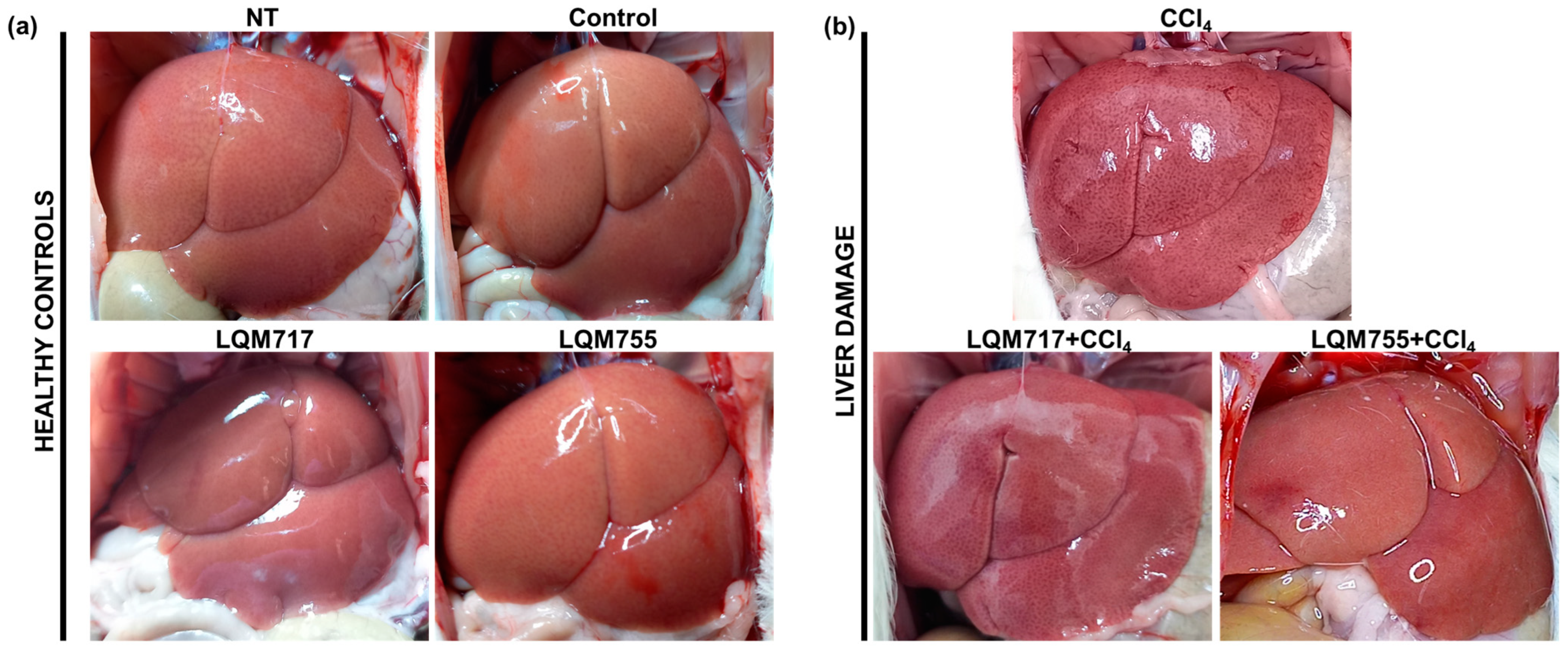
3.3. In Situ Liver Observations
Macroscopic analysis of the livers of the healthy groups (NT, Control, LQM717, and LQM755) revealed a normal architecture with a uniform red-brown color and a smooth and shiny surface (Figure 9a). In contrast, the CCl4 group presented changes in macroscopic morphology, with an opaque brown coloration and numerous necrotic spots on the tissue surface. However, in the groups pretreated with LQM717 or LQM755, the morphological alterations were less pronounced than those in the CCl4 group (Figure 9b).
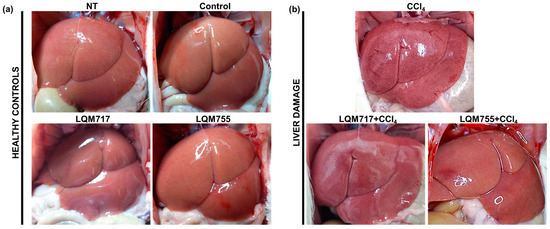
Figure 9.
Representative images of livers in situ. (a) Healthy controls. (b) Group intoxicated with only hepatotoxic CCl4 and groups pretreated with LQM717 or LQM755 and intoxicated with CCl4.
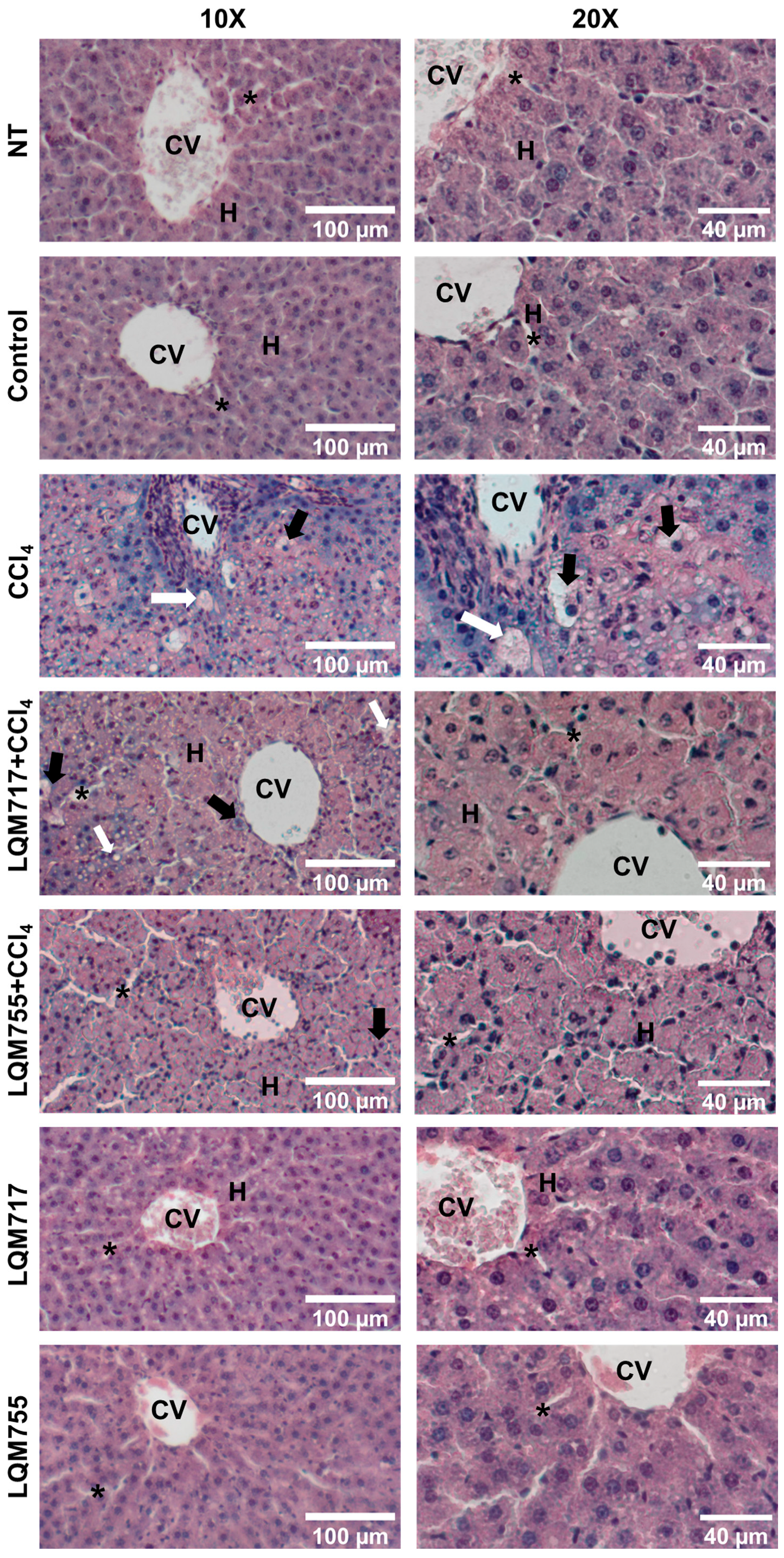
3.4. Evaluation of Microscopic Liver Damage via Hematoxylin and Eosin (H&E) Staining
Histological liver sections stained with H&E (Figure 10) revealed that healthy animals (NT, Control, LQM717, and LQM755 groups) had a normal liver parenchyma without morphological alterations, with sinusoids and cords of hepatocytes radiating from centrilobular veins to the periphery of the lobule. In contrast, the CCl4 group presented evident alterations in liver architecture, with a liver parenchyma that presented abundant necrotic lesions and balloon degeneration, characterized by enlarged hepatocytes with granular content due to cytoskeleton collapse, in addition to the presence of steatosis and inflammatory infiltration. However, the LQM717 + CCl4 and LQM755 + CCl4 groups showed significant prevention, with a notable decrease in necrotic lesions, inflammatory infiltration, and steatosis compared with those of the CCl4 group.
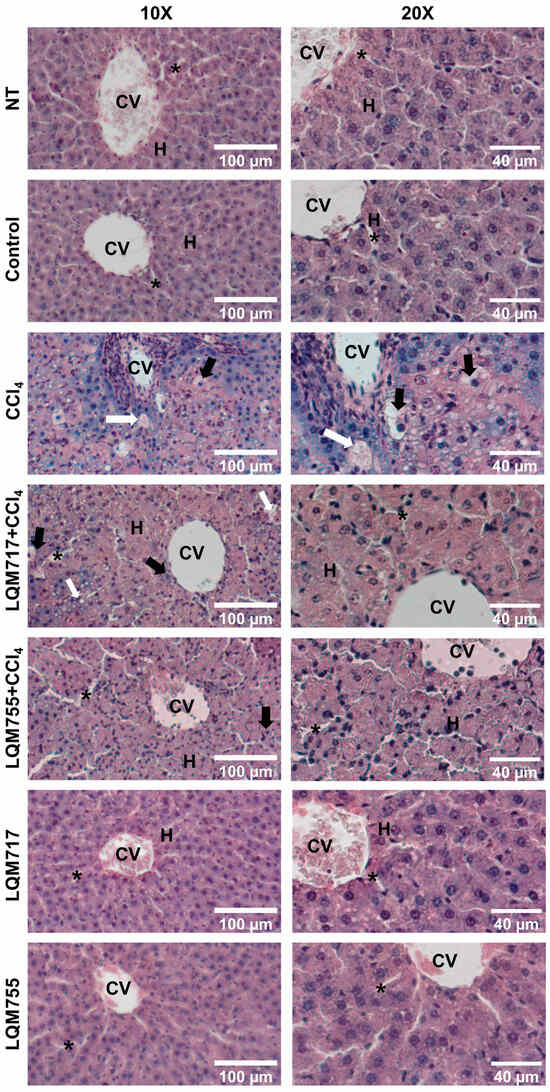
Figure 10.
Representative images of histological liver sections stained with H&E. CV = central vein; H = hepatocytes; white arrow = steatosis; black arrow = ballooning necrosis; asterisk = sinusoid.
3.5. Liver Function Tests
Table 3 presents the results of the liver function tests. The serum enzymatic activity of ALT in healthy groups (NT, Control, LQM717, and LQM755) was significantly different (p < 0.0001) in relation to the CCl4, LQM717 + CCl4, and LQM755 + CCl4 groups. Furthermore, the LQM717 + CCl4 and LQM755 + CCl4 groups showed a partial prevention to increased ALT activity, with significant differences compared to the CCl4 group (p = 0.0045, p = 0.0401, respectively).
Table 3.
Biochemical tests on serum and liver samples.
The enzymatic activity of ALP was partially and significantly lower only in the LQM755 + CCl4 group with a statistical significance (p < 0.0001) in relation to CCl4. On the other hand, all healthy controls showed significant differences (**** p < 0.0001) with the CCl4 and LQM717 + CCl4 groups, while the NT (p = 0.0012), LQM717 (p = 0.0070), LQM755 (p = 0.0030), and Control (p = 0.0009) groups were statistically significant with respect to the LQM755 + CCl4 group.
GGT serum activity and total bilirubin did not increase in the LQM755 + CCl4 group, without significant differences from those of the healthy groups and with significant differences from those of the CCl4 group (p = 0.0006, p = 0.0349, respectively). The LQM717 + CCl4 group only showed partial prevention to an increase in GGT activity (p = 0.0473) compared to CCl4. Direct bilirubin was significantly greater in the CCl4 group than in the NT (p = 0.0093), Control (p = 0.0098), LQM717, and LQM755 (p = 0.0085) groups, whereas the LQM717 + CCl4 and LQM755 + CCl4 groups did not differ significantly from healthy animals and tended to reduce this parameter in relation to the CCl4 group.
The serum albumin and liver glycogen levels were significantly lower (p < 0.0001) in the CCl4 group than in the healthy control groups (NT, Control, LQM717, and LQM755). In the LQM717 + CCl4 group, the reduction in serum albumin and liver glycogen contents was partially prevented (p = 0.0281, p = 0.035, respectively, with respect to CCl4). In the LQM755 + CCl4 group, the glycogen content reduction was partially prevented compared to the CCl4 group (p = 0.0400) and healthy controls (p < 0.0001), whereas serum albumin reduction was completely prevented. In all tests, healthy animals (NT, Control, LQM717, and LQM755) did not differ significantly.
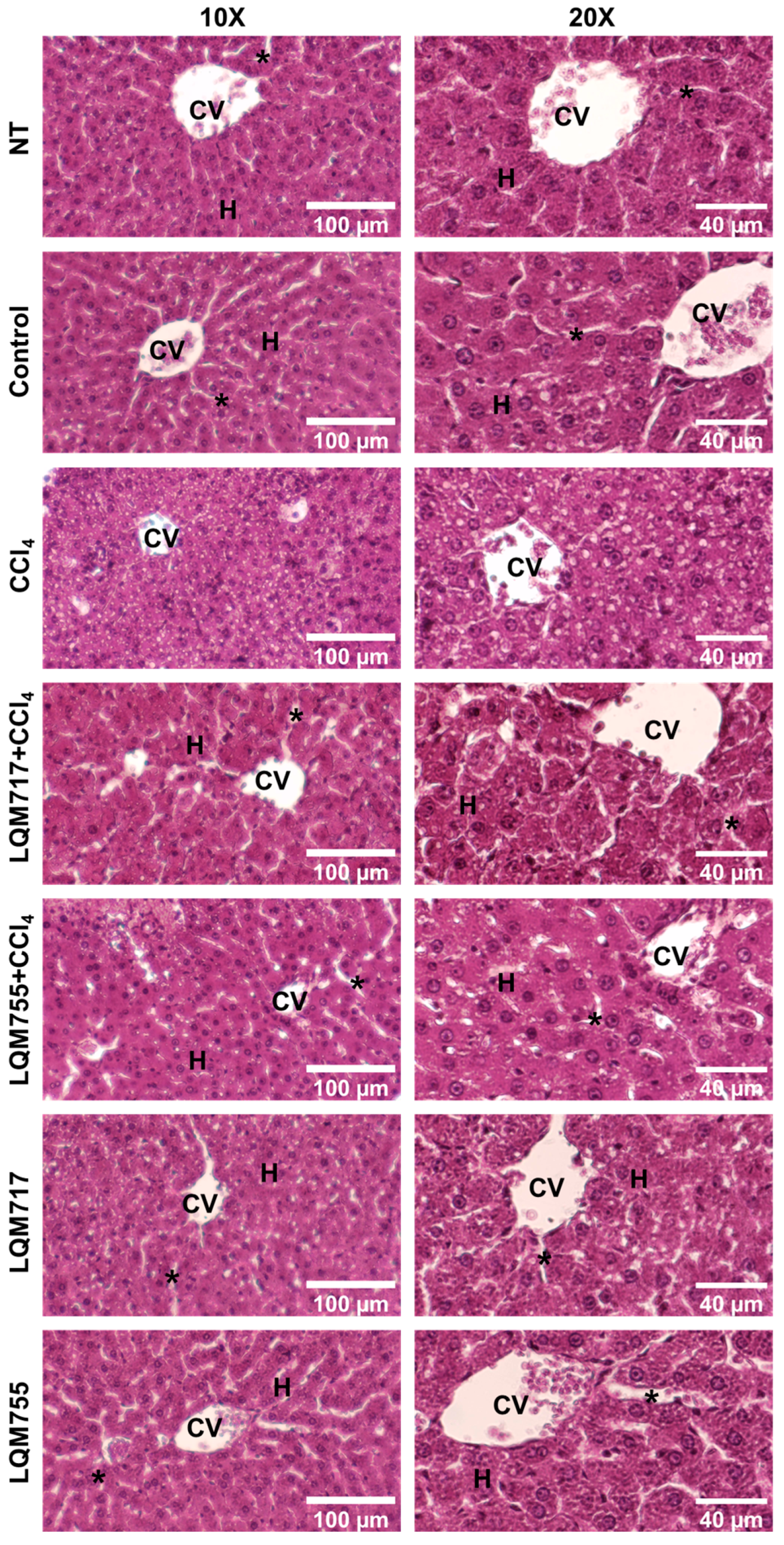
3.6. Detection of Polysaccharides with Periodic Acid-Schiff (PAS) Staining
PAS staining allows the detection of polysaccharides, especially glycogen deposits in liver tissue, which is reflected in a red-purple hue in the tissue [31]. This result is consistent with the biochemical quantification of liver glycogen (Table 3). In the healthy groups (NT, Control, LQM717, and LQM755), an intense and uniform red-purple coloration was observed in the cytoplasm of hepatocytes (Figure 11), suggesting the presence of abundant carbohydrates. In contrast, the tissues of the CCl4 group presented a significant decrease in the intensity of the red-purple color, indicating a decrease in carbohydrates in the liver parenchyma compared with those of the healthy groups. Finally, the LQM717 + CCl4 and LQM755 + CCl4 groups presented preservation of the red-purple hue, suggesting the presence of carbohydrates in the tissue, in contrast to the CCl4 group.
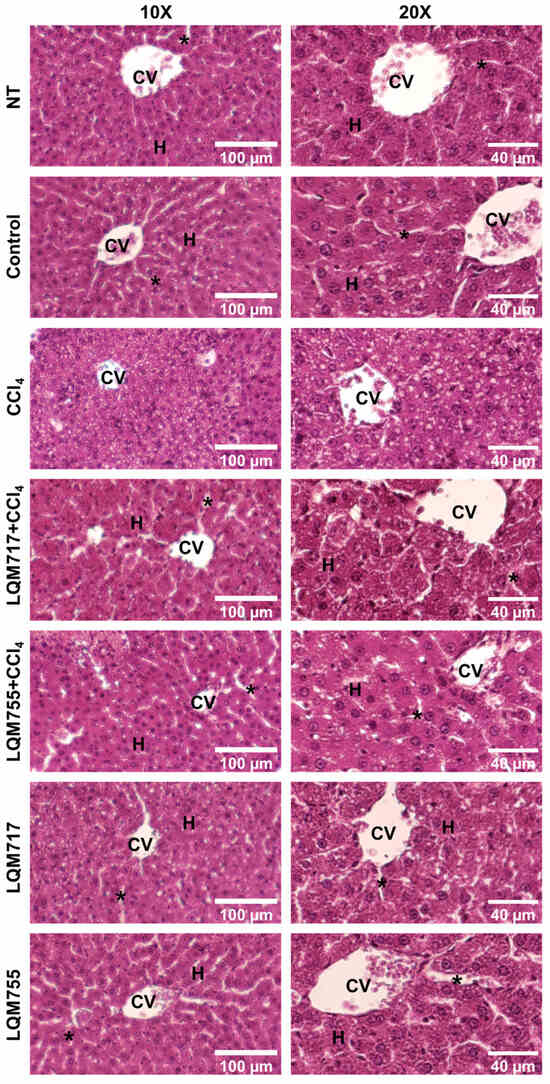
Figure 11.
Representative images of histological liver sections stained with PAS. CV = central vein; H = hepatocytes; asterisk = sinusoid.
3.7. Identification of Possible Protein Targets in the Liver
The identification of the possible mechanisms of action of the compounds LQM717 and LQM755 was carried out via five bioinformatic platforms specializing in the prediction of protein targets: Super-PRED (SP), Pharos (P), SEA (S), TargetNet (TN), and Binding DB (BDB). Protein targets expressed in parenchymal and nonparenchymal cells of the liver that perform liver functions or that are prognostic markers in liver cancer were selected. The results allowed the identification of 55 potential protein targets that could interact with the compounds LQM717 and LQM755 in the liver (Table 4, Table 5 and Table 6). Among these genes, 21 were identified as prognostic markers in liver cancer. In total, 32 possible protein targets were identified for compound LQM717 (Table 4 and Table 6), and 36 were identified for compound LQM755 (Table 5 and Table 6), sharing 13 potential protein targets between the two compounds (Table 6).
Table 4.
Bioinformatic analysis of putative protein targets to LQM717 in liver.
Table 5.
Bioinformatic analysis of putative protein targets to LQM755 in liver.
Table 6.
Bioinformatic analysis of putative protein targets that could interact with both LQM717 and LQM755 in liver.
4. Discussion
In the present work, an experimental model of acute liver damage induced by CCl4 was established. This model is capable of generating acute liver damage and centrilobular necrosis by administering a single sublethal dose of 4 g/kg orally (p.o.) [44,45]. The results revealed that the ratio between liver weight and body weight in the LQM717 + CCl4 group increased significantly, suggesting the presence of hepatomegaly. This condition could be associated with inflammation caused by the hepatotoxic effect of CCl4 [46]. In addition, the observed hepatomegaly could also be related to the defense mechanisms used by the body to maintain homeostasis. This process involves an increase in metabolic activity, which is necessary for the repair of damaged tissue [47]. Furthermore, inflammation could be an adaptive response to limit damage and promote liver regeneration.
Importantly, hepatomegaly and inflammation are key indicators of liver response to CCl4-induced injury. These findings underscore the relevance of compounds LQM717 and LQM755 in modulating the liver response to damage, suggesting their therapeutic potential in the prevention and treatment of toxin-induced liver diseases.
Macroscopic (Figure 9) and microscopic (Figure 10 and Figure 11) observations of the livers revealed that the compounds LQM717 and LQM755 reduce acute liver damage induced by a sublethal dose of CCl4. These observations are consistent with the results of the liver function tests (Table 3). The NT and Control groups, as well as the groups treated with LQM717 and LQM755, did not differ significantly, suggesting that at the doses used in this study, these compounds do not have hepatotoxic effects.
In contrast, the CCl4-treated group presented significant increases in ALT, ALP, and GGT enzymatic activity, as well as total and direct bilirubin levels, and a decrease in serum albumin and liver glycogen content. This is because CCl4 generates acute toxicity through its metabolism mediated by CYP450, producing free radicals such as the trichloromethyl radical (CCl3˙) and the trichloromethylperoxy radical (CCl3OO ˙), which react with proteins, nucleic acids, and lipids [46].
Pretreatment with LQM717 or LQM755 significantly prevented CCl4-induced liver damage. However, differences were observed between the two compounds derived from cinnamic acid (Table 3). LQM755 showed greater hepatoprotective activity than LQM717 since it partially but significantly prevented increases in ALT and ALP, whereas LQM717 only showed partial ALT activity. Further, LQM755 demonstrated a higher decrease in cholestatic biomarkers than LQM717, completely preventing an increase in GGT, whereas LQM717 only partially did so. Total bilirubin increase was also prevented with LQM755 pretreatment, but not with LQM717.
Cinnamic acid (20 mg/kg) has been shown to attenuate cisplatin-induced hepatotoxicity, resulting in partial prevention of increased total serum bilirubin [48]. Furthermore, at a dose of 50 mg/kg, the serum enzymatic activity of ALT, but not that of ALP, decreased in rats treated with gentamicin [23].
Considering that serum albumin is synthesized exclusively by hepatocytes and that its levels tend to decrease in chronic liver diseases and in cases of severe acute liver damage [49], the results obtained indicate that LQM755 could be a promising drug for the treatment of hepatic cholestasis; therefore, further studies will be required in cholestasis models, such as the bile duct ligation model in rats. This compound completely prevented the reduction in the serum albumin concentration, while LQM717 only partially did so. These findings suggest that LQM755 helps maintain liver function despite CCl4-induced damage.
Evaluation of liver biosynthetic capacity includes the quantification of liver glycogen and histological evaluation via PAS staining. Our results indicate that both LQM717 and LQM755 significantly prevent CCl4-induced carbohydrate depletion. These findings are related to those of a previous study with cinnamic acid (50 mg/kg), which only modestly prevented liver glycogen depletion in experimental animals challenged with CCl4 [25]. In the present work, the results for serum albumin and liver glycogen suggest that LQM717 and LQM755 help to preserve the biosynthetic capacity of the liver on acute liver damage, a condition not previously reported. The hepatoprotective effects described above may be related to previous studies evaluating the biological activity of these compounds. LQM717 prevented diethylnitrosamine-induced necrosis in a resistant hepatocyte modified model in rats [19]. LQM755 has shown antineoplastic activity by inhibiting breast cancer growth in the MCF-7 cell line and lung cancer [50].
Since the mechanism of action of LQM717 and LQM755 is still unknown, bioinformatic platforms were used to predict possible target proteins in the liver (Table 4, Table 5 and Table 6). Among the results obtained for LQM717 (Table 4), the following stand out:
- (a)
- Carboxylesterase 1 (CES1, P23141) is involved in the detoxification of xenobiotics, and its inhibition is beneficial for the treatment of metabolic disorders, such as obesity and fatty liver disease [51];
- (b)
- CYP450 3A4 (P08684) is considered an important CYP450 whose function is to detoxify bile acids, and the stimulation of its activity could be useful for the treatment of cholestatis [52];
- (c)
- The DNA excision repair protein ERCC-1 (P07992) may influence survival time after chemotherapy [53].
In the case of LQM755 (Table 5), the following targets stand out:
- (a)
- MMP-1 (P03956);
- (b)
- MMP-9 (P14780);
- (c)
- MMP-12 (P39900).
Chronic liver damage promotes fibrosis progression, a pathological condition that involves mainly the activation of liver stellate cells, which leads to excessive accumulation of the extracellular matrix (ECM) [54]. Matrix metalloproteinases (MMPs) play complex roles in modulating ECM degradation. Increasing MMP-1 reduces fibrosis progression by degrading the fibrillar ECM; MMP-9 is a matrix-degrading enzyme expressed in the early stages of liver damage through nuclear factor kappa-B (NF-κB) and mitogen-activated protein kinase, and MMP-12 facilitates the degradation of elastin [55].
In particular, NF-κB (Q04206) is another potential target protein of LQM755 (Table 5). This nuclear transcription factor is closely related to liver damage, and its inhibition delays the inflammatory response and oxidative stress and ultimately reduces hepatocellular death [56]. In addition, 13 possible target proteins were found that coincided with both compounds of interest (Table 6), including the Nuclear factor-erythroid 2 related factor 2 (Nrf2, Q16236). Dysregulation of Nrf2 signaling is related to oxidative stress and inflammation in diseases such as neurological diseases, diabetes mellitus, liver diseases, and cancer [57,58,59,60]. Therefore, there is a complex and bidirectional relationship between NF-κB and Nrf2, because these factors regulate opposite cellular pathways. On the one hand, NF-κB is a master regulator of inflammation, immune responses, and cell survival [61]. On the other hand, Nrf2 is a master transcriptional regulator of antioxidant and detoxification responses [62]. This could partly explain the superior effect of LQM755 compared to LQM717, as shown in Table 5 and Table 6; NF-κB and Nrf2 are potential protein targets for LQM755, perhaps modulating the activation of the Nrf2 antioxidant pathway and, in turn, inhibiting the NF-κB pro-inflammatory pathway. It is important to highlight that the potential therapeutic targets were identified through bioinformatic analysis and remain to be experimentally validated in future research.
Notably, the present work showed similarities between the hepatoprotective action of both compounds; LQM717 and LQM755 partially prevent increases in ALT enzymatic activity. However, LQM755 stands out by completely preventing an increase in GGT and total bilirubin, while LQM717 only partially prevents an increase in GGT. Likewise, it is highlighted that LQM755 maintains serum albumin production and significantly reduces glycogen depletion, while LQM717 only partially maintains both parameters.
5. Conclusions
The results of our study indicate that synthetic cinnamic acid analogs, specifically LQM717 and LQM755, have the ability to significantly prevent CCl4-induced acute liver damage. These compounds demonstrated remarkable hepatoprotection, in addition to maintaining the biosynthetic capacity of the liver. In particular, the compound LQM755 showed greater efficacy in the prevention of acute hepatocellular damage induced by CCl4. Additionally, bioinformatic analysis revealed several possible protein targets, which opens a wide range of opportunities to explore and elucidate the mechanisms of action and the signaling pathways responsible for the observed hepatoprotective effects. However, these compounds must be evaluated in subsequent studies for their potential anti-inflammatory, antifibrotic, and antisteatotic effects to determine their hepatoprotective potential and action mechanism. Therefore, these early findings are expected to promote further research on the hepatoprotective activity of both compounds using others acute and chronic models of liver damage. Further studies could ultimately contribute to the development of effective drug treatments for liver diseases.
Author Contributions
Conceptualization, J.R.M.-P. and L.R.A.-M.; methodology B.E.-H., A.Y.A.-L., P.A.M.-S., E.A., J.R.M.-P. and L.R.A.-M.; software, B.E.-H., J.R.M.-P., and L.R.A.-M.; validation, J.R.M.-P., E.A. and L.R.A.-M.; formal analysis, J.R.M.-P., E.A. and L.R.A.-M.; investigation, J.R.M.-P., E.A., and L.R.A.-M.; resources, J.R.M.-P. and E.A.; data curation, E.A., J.R.M.-P. and L.R.A.-M.; writing—original draft preparation, L.R.A.-M.; writing—review and editing, E.A., J.R.M.-P., A.Y.A.-L., P.A.M.-S., B.E.-H. and L.R.A.-M.; visualization, J.R.M.-P. and L.R.A.-M.; supervision, E.A., J.R.M.-P. and L.R.A.-M.; project administration, J.R.M.-P.; funding acquisition, J.R.M.-P. and E.A. All authors have read and agreed to the published version of the manuscript.
Funding
The authors wish to thank Secretaría De Ciencias Humanidades Tecnología E Innovación (SECIHTI No. 320331 “Ciencia Básica y/o de Frontera Modalidad: Paradigmas y Controversias de la Ciencia 2022”) for their financial support. Furthermore, this work was partially supported by SECIHTI under grants No. CF-2023-I-195; SECIHTI 321878; PAPIIT Program UNAM Project IN206023; and PIAPI FESC-UNAM CI2406.
Institutional Review Board Statement
All experimental protocols using rats were approved by the Research Ethics Committee, Faculty of Professional Studies Huasteca Zone, Autonomous University of San Luis Potosí, and were conducted according to institutional guidelines (date of approval 5 October 2020). All animals received care according to international guidelines and in adherence to the technical specifications established in the official Mexican standard NOM-062-ZOO-1999.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author J.R.M.-P.
Acknowledgments
Technical assistance was provided by Draucin Jiménez and Moises Hernández.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Baviskar, K.; Kshirsagar, A.; Raut, H.; Shaikh, M.R.N. Overview: Global burden of liver disease. Int. J. Pharm. Chem. Anal. 2024, 11, 1–10. [Google Scholar] [CrossRef]
- Devarbhavi, H.; Asrani, S.K.; Arab, J.P.; Nartey, Y.A.; Pose, E.; Kamath, P.S. Global burden of liver disease: 2023 update. J. Hepatol. 2023, 79, 516–537. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yu, L. Role and therapeutic perspectives of extracellular vesicles derived from liver and adipose tissue in metabolic dysfunction-associated steatotic liver disease. Artif Cells Nanomed. Biotechnol. 2024, 52, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Zannad, F.; Sanyal, A.J.; Butler, J.; Ferreira, J.P.; Girerd, N.; Miller, V.; Pandey, A.; Parikh, C.R.; Ratziu, V.; Younossi, Z.M.; et al. MASLD and MASH at the crossroads of hepatology trials and cardiorenal metabolic trials. J. Intern. Med. 2024, 296, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Wong, G.; Anstee, Q.M.; Henry, L. The Global Burden of Liver Disease. Clin. Gastroenterol. Hepatol. 2023, 21, 1978–1991. [Google Scholar] [CrossRef]
- Cheemerla, S.; Balakrishnan, M. Global Epidemiology of Chronic Liver Disease. Clin. Liver Dis. 2021, 17, 365–370. [Google Scholar] [CrossRef]
- Bittermann, T.; Shaked, A.; Goldberg, D.S. When Living Donor Liver Allografts Fail: Exploring the Outcomes of Retransplantation Using Deceased Donors. Am. J. Transplant. 2017, 17, 1097–1102. [Google Scholar] [CrossRef]
- Rumgay, H.; Ferlay, J.; de Martel, C.; Georges, D.; Ibrahim, A.S.; Zheng, R.; Wei, W.; Lemmens, V.E.P.P.; Soerjomataram, I. Global, regional and national burden of primary liver cancer by subtype. Eur. J. Cancer. 2022, 161, 108–118. [Google Scholar] [CrossRef]
- Kim, G.A.; Jeong, S.; Jang, H.; Lee, D.H.; Joo, S.K.; Kim, W. Metabolic Dysfunction-Associated Steatotic Liver Disease and Metabolic Dysfunction-Associated Steatotic Liver Disease with Increased Alcohol Intake Increase the Risk of Developing Hepatocellular Carcinoma and Incident or Decompensated Cirrhosis: A Korean Nationwide Study. Liver Cancer 2023, 13, 426–437. [Google Scholar] [CrossRef]
- Adhoute, X.; Pietri, O.; Pénaranda, G.; Wolf, T.; Beaurain, P.; Monnet, O.; Laquière, A.; Bonomini, J.; Neumann, F.; Levrel, O.; et al. Intrahepatic Cholangiocarcinoma and Hepatocellular Carcinoma: Real-life Data on Liver Disease, Treatment and Prognosis. J. Clin. Transl. Hepatol. 2023, 11, 1106–1117. [Google Scholar] [CrossRef]
- Gordan, J.D.; Kennedy, E.B.; Abou-Alfa, G.K.; Beal, E.; Finn, R.S.; Gade, T.P.; Goff, L.; Gupta, S.; Guy, J.; Hoang, H.T.; et al. Systemic Therapy for Advanced Hepatocellular Carcinoma: ASCO Guideline Update. J. Clin. Oncol. 2024, 42, 1830–1850. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, W.; Jiang, L.; Chen, Y. Recent advances in systemic therapy for hepatocellular carcinoma. Biomark. Res. 2022, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Kudo, M. Recent Advances in Systemic Therapy for Hepatocellular Carcinoma in an Aging Society: 2020 Update. Liver Cancer 2020, 9, 640–662. [Google Scholar] [CrossRef]
- Danish, F.A.; Koul, S.S.; Subhani, F.R.; Rabbani, A.E.; Yasmin, S. Antiviral therapy in HCV-infected decompensated cirrhotics. Saudi J. Gastroenterol. 2010, 16, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Xu, Q.; Guo, H.Y.; Huang, X.; Chen, F.; Jin, L.; Quan, Z.S.; Shen, Q.K. Application of cinnamic acid in the structural modification of natural products: A review. Phytochemistry 2023, 206, 113532. [Google Scholar] [CrossRef]
- Contardi, M.; Lenzuni, M.; Fiorentini, F.; Summa, M.; Bertorelli, R.; Suarato, G.; Athanassiou, A. Hydroxycinnamic Acids and Derivatives Formulations for Skin Damages and Disorders: A Review. Pharmaceutics 2021, 13, 999. [Google Scholar] [CrossRef]
- Sun, W.; Shahrajabian, M.H. Therapeutic Potential of Phenolic Compounds in Medicinal Plants-Natural Health Products for Human Health. Molecules 2023, 28, 1845. [Google Scholar] [CrossRef]
- Alarcón-López, A.; Hernández, M.; Aguirre-Vidal, P.; Granados, L.A.; Vazquez Valadez, V.; Martínez-Soriano, P.; Briseño-Lugo, P.; Velázquez, A.; Rul-Ramírez, E.; Jiménez-Jiménez, M.; et al. Computational Insight and Anticancer Effect of Cinnamic Acid-Derivative Amide Compounds. J. Braz. Chem. Soc. 2024, 35, e20230195. [Google Scholar] [CrossRef]
- Macías-Pérez, J.R.; Beltrán-Ramírez, O.; Vásquez-Garzón, V.R.; Salcido-Neyoy, M.E.; Martínez-Soriano, P.A.; Ruiz-Sánchez, M.B.; Angeles, E.; Villa-Treviño, S. The effect of caffeic acid phenethyl ester analogues in a modified resistant hepatocyte model. Anti-Cancer Drugs 2013, 24, 394–405. [Google Scholar] [CrossRef]
- Clifford, M.N.; King, L.J.; Kerimi, A.; Pereira-Caro, M.G.; Williamson, G. Metabolism of phenolics in coffee and plant-based foods by canonical pathways: An assessment of the role of fatty acid β-oxidation to generate biologically-active and -inactive intermediates. Crit. Rev. Food Sci. Nutr. 2024, 64, 3326–3383. [Google Scholar] [CrossRef]
- Sova, M.; Saso, L. Natural Sources, Pharmacokinetics, Biological Activities and Health Benefits of Hydroxycinnamic Acids and Their Metabolites. Nutrients 2020, 12, 2190. [Google Scholar] [CrossRef] [PubMed]
- Ketnawa, S.; Reginio, F.C., Jr.; Thuengtung, S.; Ogawa, Y. Changes in bioactive compounds and antioxidant activity of plant-based foods by gastrointestinal digestion: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 4684–4705. [Google Scholar] [CrossRef] [PubMed]
- Babaeenezhad, E.; Nouryazdan, N.; Nasri, M.; Ahmadvand, H.; Moradi Sarabi, M. Cinnamic acid ameliorate gentamicin-induced liver dysfunctions and nephrotoxicity in rats through induction of antioxidant activities. Heliyon 2021, 7, e07465. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Ashafaq, M.; Alshahrani, S.; Siddiqui, R.; Ahmed, R.A.; Khuwaja, G.; Islam, F. Cinnamon oil against acetaminophen-induced acute liver toxicity by attenuating inflammation, oxidative stress and apoptosis. Toxicol. Rep. 2020, 7, 1296–1304. [Google Scholar] [CrossRef]
- Pérez-Alvarez, V.; Bobadilla, R.A.; Muriel, P. Structure-hepatoprotective activity relationship of 3,4-dihydroxycinnamic acid (caffeic acid) derivatives. J. Appl. Toxicol. 2001, 21, 527–531. [Google Scholar] [CrossRef]
- Li, X.; Hu, Y.; He, B.; Li, L.; Tian, Y.; Xiao, Y.; Shang, H.; Zou, Z. Design, synthesis and evaluation of ursodeoxycholic acid-cinnamic acid hybrids as potential anti-inflammatory agents by inhibiting Akt/NF-κB and MAPK signaling pathways. Eur. J. Med. Chem. 2023, 260, 115785. [Google Scholar] [CrossRef]
- Martínez-Soriano, P.; Macías-Pérez, J.; Velázquez, A.; Camacho-Enriquez, B.; Pretelín-Castillo, G.; Ruiz-Sánchez, M.; Abrego-Reyes, V.; Villa-Treviño, S.; Angeles, E. Solvent-Free Synthesis of Carboxylic Acids and Amide Analogs of CAPE (Caffeic Acid Phenethyl Ester) under Infrared Irradiation Conditions. Green Sustain. Chem. 2015, 5, 81–91. [Google Scholar] [CrossRef]
- NOM-062-ZOO-1999; Especificaciones Técnicas Para La Producción, Cuidado Y Uso De Los Animales De Laboratorio. Diario Oficial. 1999, p. 107. Available online: https://www.gob.mx/cms/uploads/attachment/file/203498/NOM-062-ZOO-1999_220801.pdf (accessed on 12 May 2024).
- Syabani, D.M.; Eliyani, H.; Suharsono, S.; Rantam, F.A.; Ma’ruf, A. Postmortem Interval Estimation Time from Algormortis Temperature of Rats Expressed by MARS Model Approach. KnE Life Sci. 2017, 3, 404–412. [Google Scholar] [CrossRef]
- Luna, L.G. Manual of Histologic Staining Methods of the Armeed Forces Institute of Pathology, 3rd ed.; Blakiston Division, McGraw Hill: New York, NY, USA, 1968. [Google Scholar]
- Zakout, Y.M.; Abdellah, M.A.; Abdallah, M.A.; Batran, S.A. Optimization of PAS stain and similar Schiff’s based methods for glycogen demonstration in liver tissue. Histochem. Cell Biol. 2024, 161, 359–364. [Google Scholar] [CrossRef]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef]
- Bergmeyer, H.U.; Grassl, M.; Walter, H.E. Methods of Enzymatic Analysis, 3rd ed.; Bergmeyer, H.U.M., Ed.; Verlag Chemie: Deerfield Beach, FL, USA, 1983; Volume 2, pp. 269–270. [Google Scholar]
- Seifter, S.; Dayton, S. The estimation of glycogen with the anthrone reagent. Arch. Biochem. 1950, 25, 191–200. [Google Scholar] [PubMed]
- Zheng, S.; Yan, X.; Yang, Y.; Xu, J. Identifying Structure-Property Relationships through SMILES Syntax Analysis with Self-Attention Mechanism. J. Chem. Inf. Model. 2019, 59, 914–923. [Google Scholar] [CrossRef]
- Gfeller, D.; Grosdidier, A.; Wirth, M.; Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: A web server for target prediction of bioactive small molecules. Nucleic Acids Res. 2014, 42, W32–W38. [Google Scholar] [CrossRef] [PubMed]
- Gallo, K.; Goede, A.; Preissner, R.; Gohlke, B.O. SuperPred 3.0: Drug classification and target prediction—A machine learning approach. Nucleic Acids Res. 2022, 50, W726–W731. [Google Scholar] [CrossRef] [PubMed]
- Gaulton, A.; Bellis, L.J.; Bento, A.P.; Chambers, J.; Davies, M.; Hersey, A.; Light, Y.; McGlinchey, S.; Michalovich, D.; Al-Lazikani, B.; et al. ChEMBL: A large-scale bioactivity database for drug discovery. Nucleic Acids Res. 2012, 40, D1100–D1107. [Google Scholar] [CrossRef]
- Wang, X.; Shen, Y.; Wang, S.; Li, S.; Zhang, W.; Liu, X.; Lai, L.; Pei, J.; Li, H. PharmMapper 2017 update: A web server for potential drug target identification with a comprehensive target pharmacophore database. Nucleic Acids Res. 2017, 45, W356–W360. [Google Scholar] [CrossRef]
- Kelleher, K.J.; Sheils, T.K.; Mathias, S.L.; Yang, J.J.; Metzger, V.T.; Siramshetty, V.B.; Nguyen, D.T.; Jensen, L.J.; Vidović, D.; Schürer, S.C.; et al. Pharos 2023: An integrated resource for the understudied human proteome. Nucleic Acids Res. 2023, 51, D1405–D1416. [Google Scholar] [CrossRef]
- Wang, Z.; Liang, L.; Yin, Z.; Lin, J. Improving chemical similarity ensemble approach in target prediction. J. Cheminform. 2016, 8, 20. [Google Scholar] [CrossRef]
- Min, S.; Lee, B.; Yoon, S. TargetNet: Functional microRNA Target Prediction with Deep Neural Networks. Bioinformatics 2022, 38, 671–677. [Google Scholar] [CrossRef]
- Gilson, M.K.; Liu, T.; Baitaluk, M.; Nicola, G.; Hwang, L.; Chong, J. BindingDB in 2015: A public database for medicinal chemistry, computational chemistry and systems pharmacology. Nucleic Acids Res. 2016, 44, D1045–D1053. [Google Scholar] [CrossRef]
- Aldaba-Muruato, L.R.; Moreno, M.G.; Shibayama, M.; Tsutsumi, V.; Muriel, P. Protective effects of allopurinol against acute liver damage and cirrhosis induced by carbon tetrachloride: Modulation of NF-kappaB, cytokine production and oxidative stress. Biochim. Biophys. Acta 2012, 1820, 65–75. [Google Scholar] [CrossRef]
- Zhai, H.; Zhang, J.; Shang, D.; Zhu, C.; Xiang, X. The progress to establish optimal animal models for the study of acute-on-chronic liver failure. Front. Med. 2023, 10, 1087274. [Google Scholar] [CrossRef]
- Al Amin, A.S.M.; Menezes, R.G. Carbon Tetrachloride Toxicity. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Varela, M.L.; Mogildea, M.; Moreno, I.; Lopes, A. Acute Inflammation and Metabolism. Inflammation 2018, 41, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Tohamy, A.A.; Aref, A.M.; Moneim, A.E.A.; Sayed, R.H.; Esmat, A.; Moneim, A. Cinnamic acid Attenuates Cisplatin-Induced Hepatotoxicity and Nephrotoxicity. J. Basic Environ. Sci. 2016, 3, 1–9. [Google Scholar] [CrossRef]
- Wu, N.; Liu, T.; Tian, M.; Liu, C.; Ma, S.; Cao, H.; Bian, H.; Wang, L.; Feng, Y.; Qi, J. Albumin, an interesting and functionally diverse protein, varies from ‘native’ to ‘effective’ (Review). Mol. Med. Rep. 2024, 29, 24. [Google Scholar] [CrossRef]
- Martínez-Rosas, J.R.; Díaz-Torres, R.; Ramírez-Noguera, P.; López-Barrera, L.D.; Escobar-Chavez, J.J.; Ángeles, E.R. PLGA nanoparticles of a new cinnamic acid derivative inhibits cellular proliferation on breast cancer cell line MCF-7 in a PPARγ dependent way. Die Pharm. 2020, 75, 324–328. [Google Scholar]
- Wang, D.D.; Wang, Z.Z.; Liu, W.C.; Qian, X.K.; Zhu, Y.D.; Wang, T.G.; Pan, S.M.; Zou, L.W. Pyrazolone compounds could inhibit CES1 and ameliorates fat accumulation during adipocyte differentiation. Bioorg. Chem. 2024, 150, 107536. [Google Scholar] [CrossRef]
- Chen, J.; Zhao, K.N.; Chen, C. The role of CYP3A4 in the biotransformation of bile acids and therapeutic implication for cholestasis. Ann. Transl. Med. 2014, 2, 7. [Google Scholar] [CrossRef]
- Lentz, F.; Tran, A.; Rey, E.; Pons, G.; Tréluyer, J.M. Pharmacogenomics of fluorouracil, irinotecan, and oxaliplatin in hepatic metastases of colorectal cancer: Clinical implications. Am. J. Pharmacogenomics 2005, 5, 21–33. [Google Scholar] [CrossRef]
- Chen, L.; Guo, W.; Mao, C.; Shen, J.; Wan, M. Liver fibrosis: Pathological features, clinical treatment and application of therapeutic nanoagents. J. Mater Chem. B 2024, 12, 1446–1466. [Google Scholar] [CrossRef]
- Sabir, U.; Gu, H.M.; Zhang, D.W. Extracellular matrix turnover: Phytochemicals target and modulate the dual role of matrix metalloproteinases (MMPs) in liver fibrosis. Phytother. Res. 2023, 37, 4932–4962. [Google Scholar] [CrossRef] [PubMed]
- Taru, V.; Szabo, G.; Mehal, W.; Reiberger, T. Inflammasomes in chronic liver disease: Hepatic injury, fibrosis progression and systemic inflammation. J. Hepatol. 2024, 5, 895–910. [Google Scholar] [CrossRef]
- Wang, J.; Cao, Y.; Lu, Y.; Zhu, H.; Zhang, J.; Che, J.; Zhuang, R.; Shao, J. Recent progress and applications of small molecule inhibitors of Keap1-Nrf2 axis for neurodegenerative diseases. Eur. J. Med. Chem. 2024, 264, 115998. [Google Scholar] [CrossRef] [PubMed]
- Yi, M.; Toribio, A.J.; Salem, Y.M.; Alexander, M.; Ferrey, A.; Swentek, L.; Tantisattamo, E.; Ichii, H. Nrf2 Signaling Pathway as a Key to Treatment for Diabetic Dyslipidemia and Atherosclerosis. Int. J. Mol. Sci. 2024, 25, 5831. [Google Scholar] [CrossRef]
- Park, J.S.; Rustamov, N.; Roh, Y.S. The Roles of NFR2-Regulated Oxidative Stress and Mitochondrial Quality Control in Chronic Liver Diseases. Antioxidants 2023, 12, 1928. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Xiao, M.; Hu, S.; Wang, M. Keap1-Nrf2 pathway: A key mechanism in the occurrence and development of cancer. Front. Oncol. 2024, 14, 1381467. [Google Scholar] [CrossRef]
- Guo, Q.; Jin, Y.; Chen, X.; Ye, X.; Shen, X.; Lin, M.; Zeng, C.; Zhou, T.; Zhang, J. NF-κB in biology and targeted therapy: New insights and translational implications. Signal Transduct. Target. Ther. 2024, 9, 53. [Google Scholar] [CrossRef]
- O’Rourke, S.A.; Shanley, L.C.; Dunne, A. The Nrf2-HO-1 system and inflammaging. Front. Immunol. 2024, 15, 1457010. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).