
Therapeutic Potential of BMP7 in the Treatment of Osteoporosis Caused by the Interaction between Inflammation and Corticosteroids in Inflammatory Bowel Disease

Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
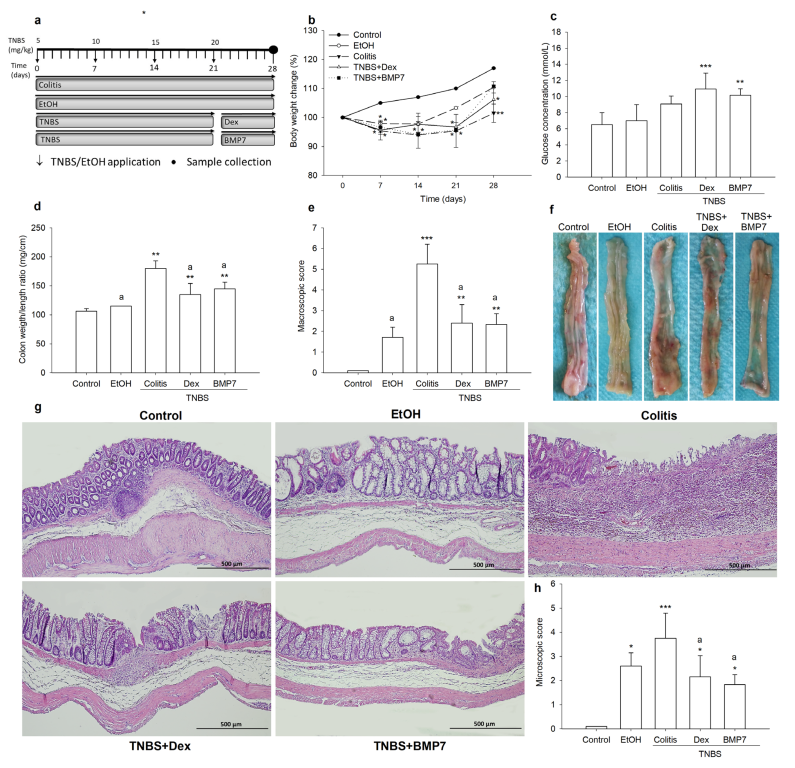
2.2. Chronic Colitis Induction and Assessment of Severity
2.3. Evaluation of Colonic Damage
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Real-Time Polymerase Chain Reaction Analysis of Gene Expression
2.6. Immunohistochemical Analysis
2.7. Bone Morphometric Analysis
2.8. Statistical Analysis
3. Results
3.1. BMP7 Treatment Diminished Clinical Symptoms and Reduced the Damage of Colonic Tissue
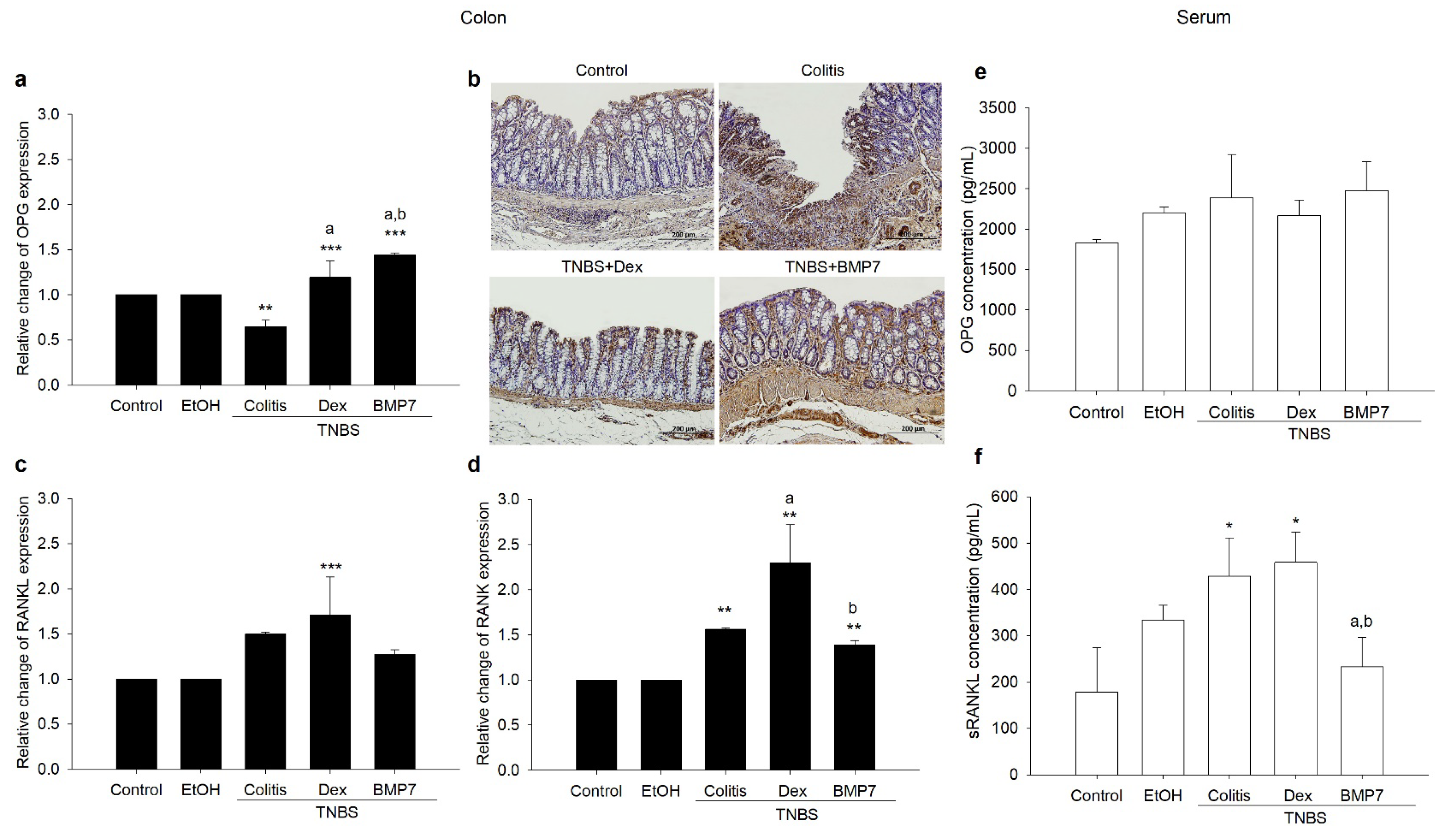
3.2. BMP7 Treatment Affected the Gene Expressions and Protein Concentrations of OPG/RANKL/RANK
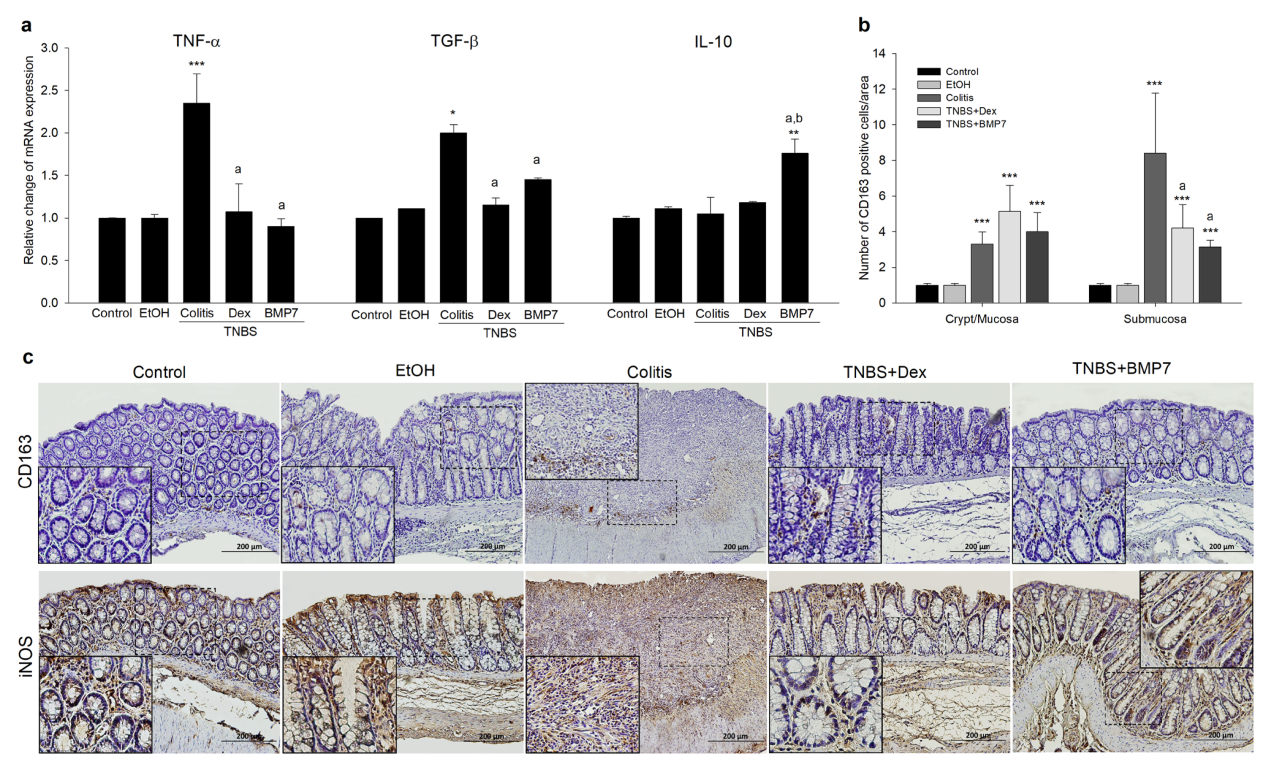
3.3. BMP7 Suppressed the Inflammatory Response in Chronic TNBS Colitis
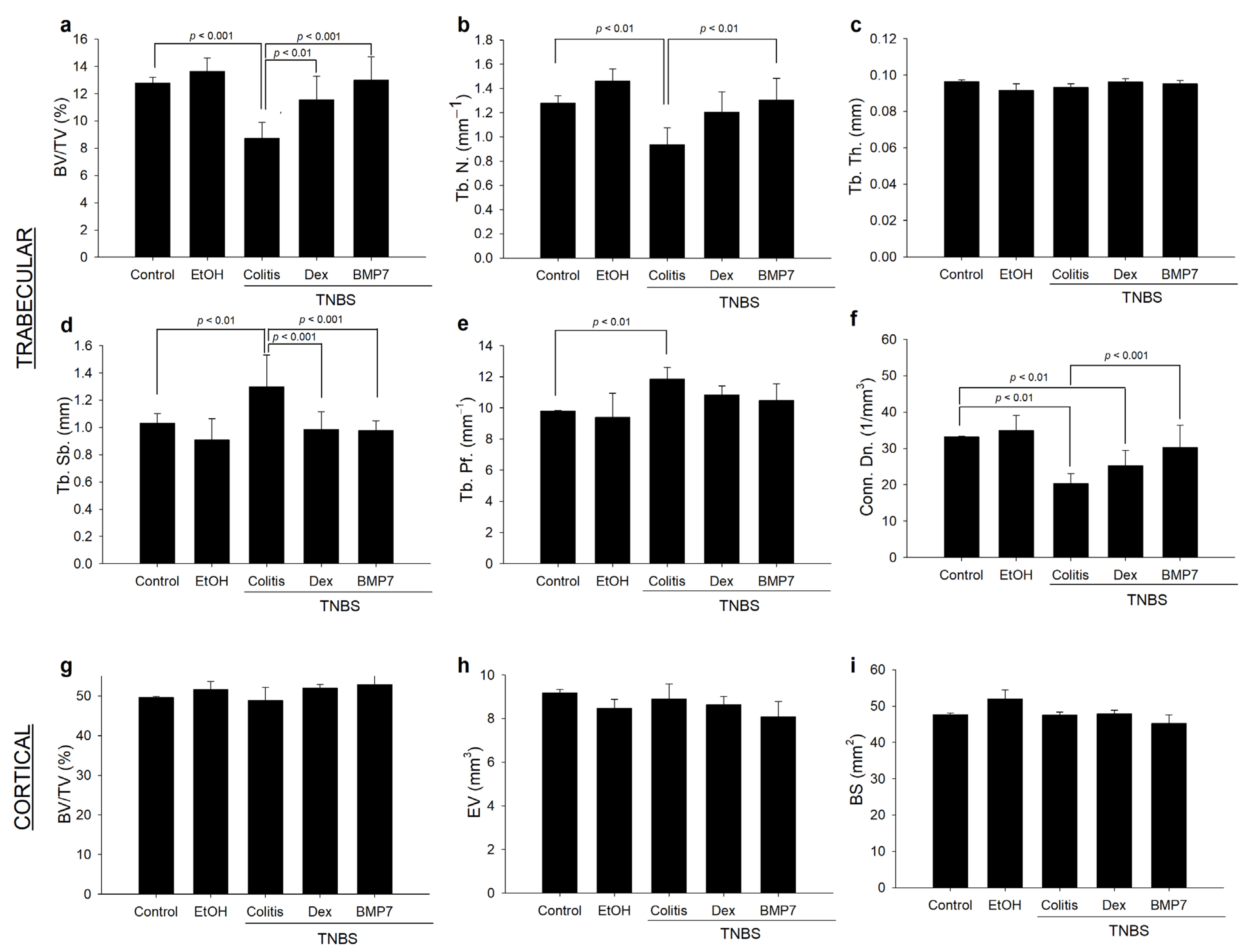
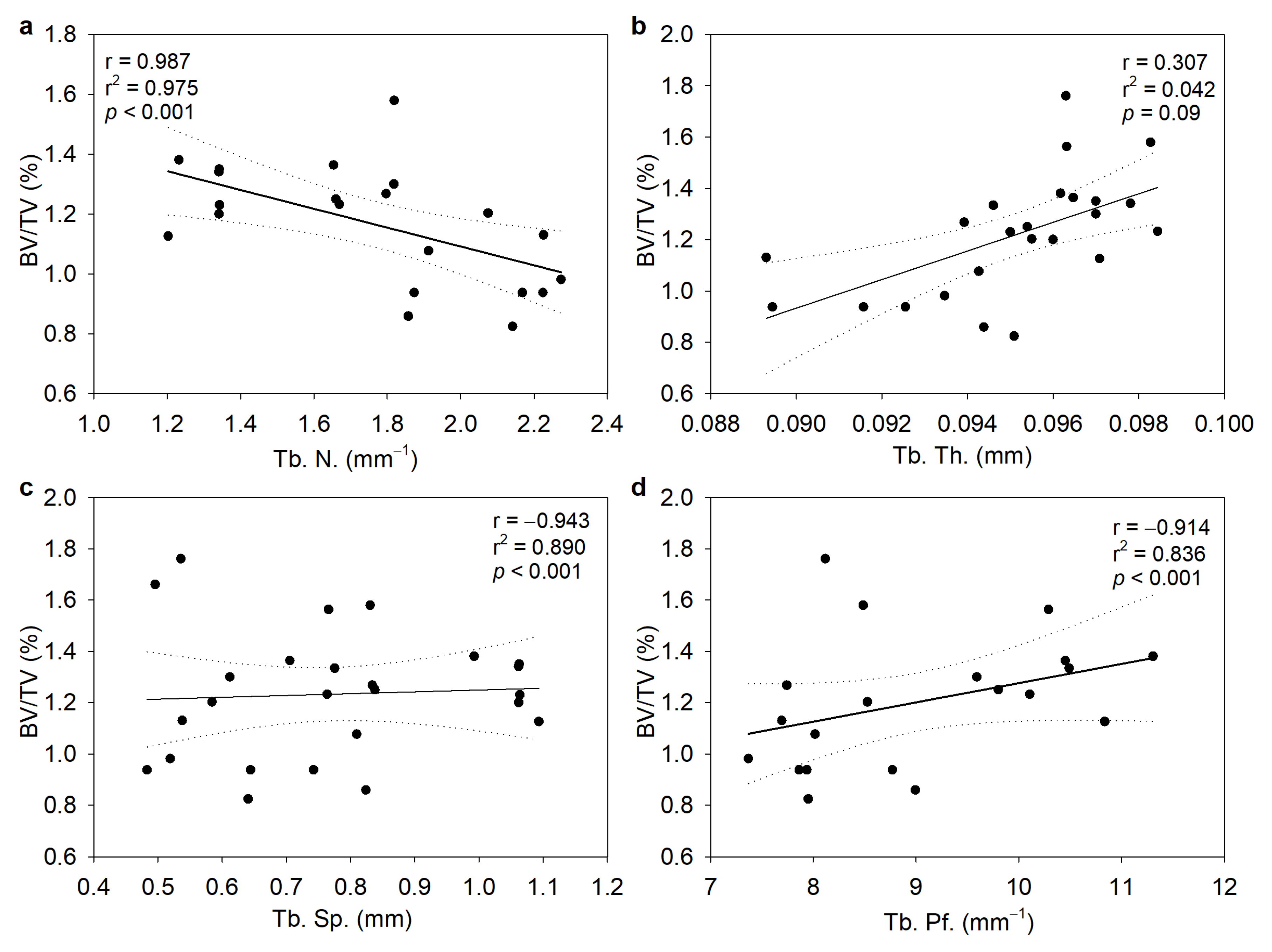
3.4. Alterations in the Trabecular and Cortical Long Bone Parameters in Chronic Colitis
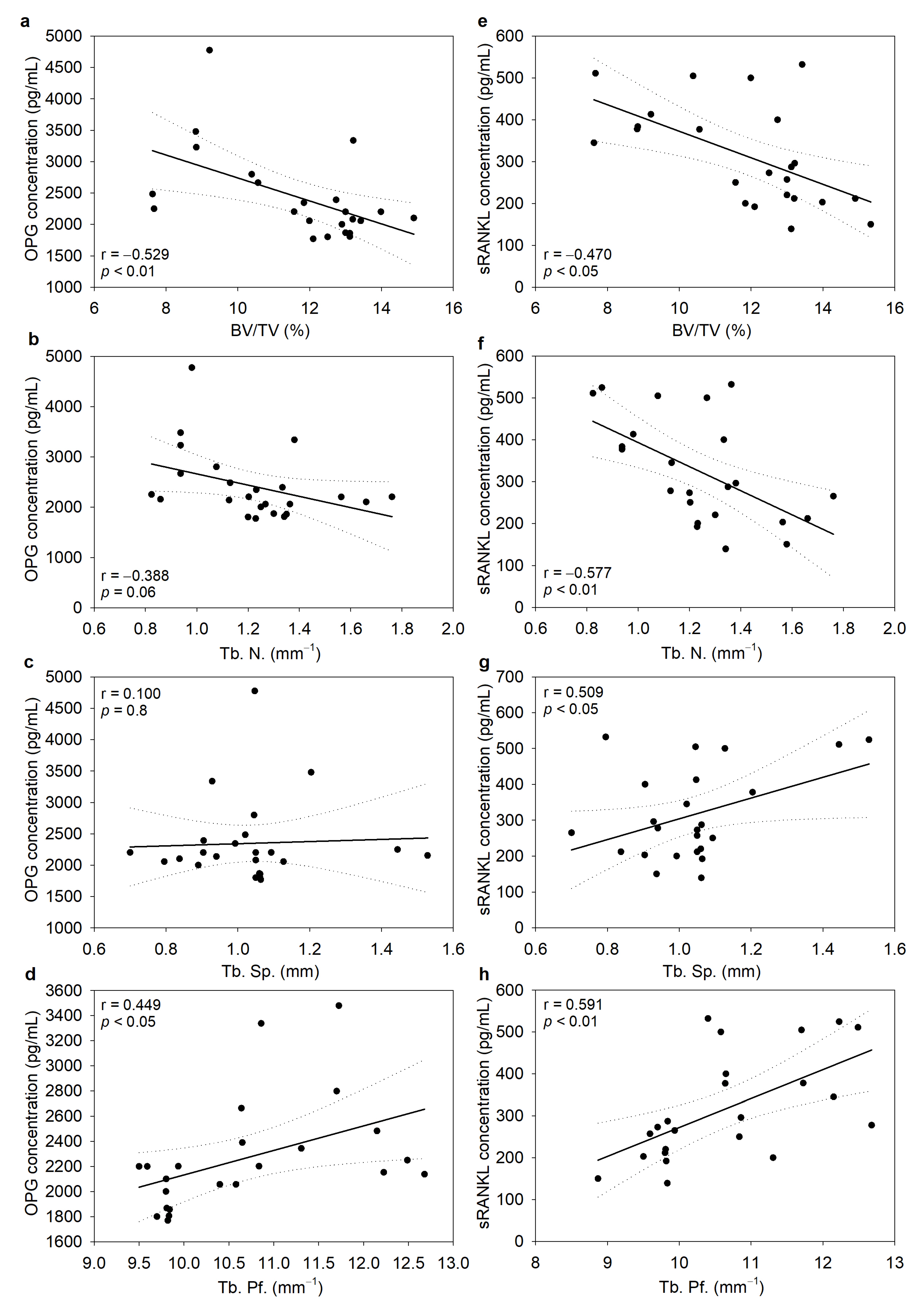
3.5. Serum Concentrations of OPG and sRANKL Were Inversely Correlated with Bone Morphometric Parameters
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alatab, S.; Sepanlou, S.G.; Ikuta, K.; Vahedi, H.; Bisignano, C.; Safiri, S.; Sadeghi, A.; Nixon, M.R.; Abdoli, A.; Abolhassani, H.; et al. The Global, Regional, and National Burden of Inflammatory Bowel Disease in 195 Countries and Territories, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2020, 5, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Caviglia, G.P.; Garrone, A.; Bertolino, C.; Vanni, R.; Bretto, E.; Poshnjari, A.; Tribocco, E.; Frara, S.; Armandi, A.; Astegiano, M.; et al. Epidemiology of Inflammatory Bowel Diseases: A Population Study in a Healthcare District of North-West Italy. J. Clin. Med. 2023, 12, 641. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q. A Comprehensive Review and Update on the Pathogenesis of Inflammatory Bowel Disease. J. Immunol. Res. 2019, 2019, 7247238. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, S.; Li, J. Treatment of Inflammatory Bowel Disease: A Comprehensive Review. Front. Med. 2021, 8, 765474. [Google Scholar] [CrossRef] [PubMed]
- Harbord, M.; Annese, V.; Vavricka, S.R.; Allez, M.; Barreiro-de Acosta, M.; Boberg, K.M.; Burisch, J.; De Vos, M.; De Vries, A.-M.; Dick, A.D.; et al. The First European Evidence-Based Consensus on Extra-Intestinal Manifestations in Inflammatory Bowel Disease. J. Crohn’s Colitis 2016, 10, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Scott, F.I.; Brensinger, C.M.; Roy, J.A.; Osterman, M.T.; Mamtani, R.; Bewtra, M.; Chen, L.; Yun, H.; Xie, F.; et al. Increased Mortality Rates with Prolonged Corticosteroid Therapy When Compared with Antitumor Necrosis Factor-α-Directed Therapy for Inflammatory Bowel Disease. Am. J. Gastroenterol. 2018, 113, 405–417. [Google Scholar] [CrossRef]
- Lewandowski, K.; Kaniewska, M.; Więcek, M.; Szwarc, P.; Panufnik, P.; Tulewicz-Marti, E.; Walicka, M.; Franek, E.; Rydzewska, G. Risk Factors for Osteoporosis among Patients with Inflammatory Bowel Disease—Do We Already Know Everything? Nutrients 2023, 15, 1151. [Google Scholar] [CrossRef]
- Flynn, S.; Eisenstein, S. Inflammatory Bowel Disease Presentation and Diagnosis. Surg. Clin. N. Am. 2019, 99, 1051–1062. [Google Scholar] [CrossRef]
- Seibel, M.J.; Cooper, M.S.; Zhou, H. Glucocorticoid-Induced Osteoporosis: Mechanisms, Management, and Future Perspectives. Lancet Diabetes Endocrinol. 2013, 1, 59–70. [Google Scholar] [CrossRef]
- Hardy, R.S.; Zhou, H.; Seibel, M.J.; Cooper, M.S. Glucocorticoids and Bone: Consequences of Endogenous and Exogenous Excess and Replacement Therapy. Endocr. Rev. 2018, 39, 519–548. [Google Scholar] [CrossRef]
- Coutinho, A.E.; Chapman, K.E. The Anti-Inflammatory and Immunosuppressive Effects of Glucocorticoids, Recent Developments and Mechanistic Insights. Mol. Cell. Endocrinol. 2011, 335, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Shi, S.; Acosta, L.; Li, W.; Lu, J.; Bao, S.; Chen, Z.; Yang, Z.; Schneider, M.D.; Chien, K.R.; et al. BMP10 Is Essential for Maintaining Cardiac Growth during Murine Cardiogenesis. Development 2004, 131, 2219–2231. [Google Scholar] [CrossRef] [PubMed]
- Simic, P.; Vukicevic, S. Bone Morphogenetic Proteins in Development and Homeostasis of Kidney. Cytokine Growth Factor Rev. 2005, 16, 299–308. [Google Scholar] [CrossRef]
- Wu, M.; Chen, G.; Li, Y.-P. TGF-β and BMP Signaling in Osteoblast, Skeletal Development, and Bone Formation, Homeostasis and Disease. Bone Res. 2016, 4, 16009. [Google Scholar] [CrossRef]
- Wu, D.H.; Hatzopoulos, A.K. Bone Morphogenetic Protein Signaling in Inflammation. Exp. Biol. Med. 2019, 244, 147–156. [Google Scholar] [CrossRef]
- Kim, B.Y.; Choi, S.H.; Kim, J.-Y.; Ko, J.; Yook, J.I.; Kim, H.S.; Lee, E.J.; Kikkawa, D.O.; Yoon, J.S. Potential Therapeutic Role of Bone Morphogenic Protein 7 (BMP7) in the Pathogenesis of Graves’ Orbitopathy. Invest. Ophthalmol. Vis. Sci. 2022, 63, 7. [Google Scholar] [CrossRef] [PubMed]
- Gould, S.E.; Day, M.; Jones, S.S.; Dorai, H. BMP-7 Regulates Chemokine, Cytokine, and Hemodynamic Gene Expression in Proximal Tubule Cells1. Kidney Int. 2002, 61, 51–60. [Google Scholar] [CrossRef]
- Singla, D.K.; Singla, R.; Wang, J. BMP-7 Treatment Increases M2 Macrophage Differentiation and Reduces Inflammation and Plaque Formation in Apo E-/- Mice. PLoS ONE 2016, 11, e0147897. [Google Scholar] [CrossRef] [PubMed]
- Mowat, A.M.; Bain, C.C. Mucosal Macrophages in Intestinal Homeostasis and Inflammation. J. Innate Immun. 2011, 3, 550–564. [Google Scholar] [CrossRef]
- Han, X.; Ding, S.; Jiang, H.; Liu, G. Roles of Macrophages in the Development and Treatment of Gut Inflammation. Front. Cell Dev. Biol. 2021, 9, 625423. [Google Scholar] [CrossRef]
- Rocher, C.; Singla, R.; Singal, P.K.; Parthasarathy, S.; Singla, D.K. Bone Morphogenetic Protein 7 Polarizes THP-1 Cells into M2 Macrophages. Can. J. Physiol. Pharmacol. 2012, 90, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.P.; Beck, P.L.; Herridge, M.S.; Depew, W.T.; Szewczuk, M.R.; Wallace, J.L. Hapten-Induced Model of Chronic Inflammation and Ulceration in the Rat Colon. Gastroenterology 1989, 96, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.Y.; Lu, Y.M.; Ou, Y.X.; Zhang, H.Z.; Chen, W.X. Dynamic Progress of 2,4,6-Trinitrobenzene Sulfonic Acid Induced Chronic Colitis and Fibrosis in Rat Model: Progress of Chronic Colitis in Rat Model. J. Dig. Dis. 2012, 13, 421–429. [Google Scholar] [CrossRef]
- Appleyard, C.B.; Wallace, J.L. Reactivation of Hapten-Induced Colitis and Its Prevention by Anti-Inflammatory Drugs. Am. J. Physiol. Gastrointest. Liver Physiol. 1995, 269, G119–G125. [Google Scholar] [CrossRef]
- Ali, T.; Lam, D.; Bronze, M.S.; Humphrey, M.B. Osteoporosis in Inflammatory Bowel Disease. Am. J. Med. 2009, 122, 599–604. [Google Scholar] [CrossRef]
- Ballester, I.; Daddaoua, A.; López-Posadas, R.; Nieto, A.; Suárez, M.D.; Zarzuelo, A.; Martínez-Augustin, O.; De Medina, F.S. The Bisphosphonate Alendronate Improves the Damage Associated with Trinitrobenzenesulfonic Acid-Induced Colitis in Rats: Alendronate and Experimental Colitis. Br. J. Pharmacol. 2007, 151, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Bobin-Dubigeon, C.; Collin, X.; Grimaud, N.; Robert, J.M.; Le Baut, G.; Petit, J.Y. Effects of Tumour Necrosis Factor-Alpha Synthesis Inhibitors on Rat Trinitrobenzene Sulphonic Acid-Induced Chronic Colitis. Eur. J. Pharmacol. 2001, 431, 103–110. [Google Scholar] [CrossRef]
- Maric, I.; Poljak, L.; Zoricic, S.; Bobinac, D.; Bosukonda, D.; Sampath, K.T.; Vukicevic, S. Bone Morphogenetic Protein-7 Reduces the Severity of Colon Tissue Damage and Accelerates the Healing of Inflammatory Bowel Disease in Rats. J. Cell. Physiol. 2003, 196, 258–264. [Google Scholar] [CrossRef]
- Rashidian, A.; Muhammadnejad, A.; Dehpour, A.-R.; Mehr, S.E.; Akhavan, M.M.; Shirkoohi, R.; Chamanara, M.; Mousavi, S.-E.; Rezayat, S.-M. Atorvastatin Attenuates TNBS-Induced Rat Colitis: The Involvement of the TLR4/NF-KB Signaling Pathway. Inflammopharmacology 2016, 24, 109–118. [Google Scholar] [CrossRef]
- Vukelić, I.; Detel, D.; Pučar, L.B.; Potočnjak, I.; Buljević, S.; Domitrović, R. Chlorogenic Acid Ameliorates Experimental Colitis in Mice by Suppressing Signaling Pathways Involved in Inflammatory Response and Apoptosis. Food Chem. Toxicol. 2018, 121, 140–150. [Google Scholar] [CrossRef]
- Erjavec, I.; Bordukalo-Niksic, T.; Brkljacic, J.; Grcevic, D.; Mokrovic, G.; Kesic, M.; Rogic, D.; Zavadoski, W.; Paralkar, V.M.; Grgurevic, L.; et al. Constitutively Elevated Blood Serotonin Is Associated with Bone Loss and Type 2 Diabetes in Rats. PLoS ONE 2016, 11, e0150102. [Google Scholar] [CrossRef] [PubMed]
- Haschka, J.; Hirschmann, S.; Kleyer, A.; Englbrecht, M.; Faustini, F.; Simon, D.; Figueiredo, C.P.; Schuster, L.; Muschitz, C.; Kocijan, R.; et al. High-Resolution Quantitative Computed Tomography Demonstrates Structural Defects in Cortical and Trabecular Bone in IBD Patients. ECCOJC 2016, 10, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, B.; Roberts, J.A.; Haag, M.M.; Spohn, S.N.; Margolis, K.G.; Sharkey, K.A.; Lian, J.B.; Mawe, G.M. Gut-Derived Serotonin Contributes to Bone Deficits in Colitis. Pharmacol. Res. 2019, 140, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Moschen, A.R.; Kaser, A.; Stadlmann, S.; Millonig, G.; Kaser, S.; Mühllechner, P.; Habior, A.; Graziadei, I.; Vogel, W.; Tilg, H. The RANKL/OPG System and Bone Mineral Density in Patients with Chronic Liver Disease. J. Hepatol. 2005, 43, 973–983. [Google Scholar] [CrossRef]
- Ashcroft, A.J.; Cruickshank, S.M.; Croucher, P.I.; Perry, M.J.; Rollinson, S.; Lippitt, J.M.; Child, J.A.; Dunstan, C.; Felsburg, P.J.; Morgan, G.J.; et al. Colonic Dendritic Cells, Intestinal Inflammation, and T Cell-Mediated Bone Destruction Are Modulated by Recombinant Osteoprotegerin. Immunity 2003, 19, 849–861. [Google Scholar] [CrossRef]
- Rocher, C.; Singla, D.K. SMAD-PI3K-Akt-MTOR Pathway Mediates BMP-7 Polarization of Monocytes into M2 Macrophages. PLoS ONE 2013, 8, e84009. [Google Scholar] [CrossRef]
- Fuller, K.; Murphy, C.; Kirstein, B.; Fox, S.W.; Chambers, T.J. TNFα Potently Activates Osteoclasts, through a Direct Action Independent of and Strongly Synergistic with RANKL. Endocrinology 2002, 143, 1108–1118. [Google Scholar] [CrossRef]
- Maric, I.; Kucic, N.; Turk Wensveen, T.; Smoljan, I.; Grahovac, B.; Zoricic Cvek, S.; Celic, T.; Bobinac, D.; Vukicevic, S. BMP Signaling in Rats with TNBS-Induced Colitis Following BMP7 Therapy. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G1151–G1162. [Google Scholar] [CrossRef]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and Therapeutic Potential of Interleukin-10. J. Exp. Med. 2020, 217, e20190418. [Google Scholar] [CrossRef]
- Ihara, S.; Hirata, Y.; Koike, K. TGF-β in Inflammatory Bowel Disease: A Key Regulator of Immune Cells, Epithelium, and the Intestinal Microbiota. J. Gastroenterol. 2017, 52, 777–787. [Google Scholar] [CrossRef]
- Shoulders, H.; Garner, K.H.; Singla, D.K. Macrophage Depletion by Clodronate Attenuates Bone Morphogenetic Protein-7 Induced M2 Macrophage Differentiation and Improved Systolic Blood Velocity in Atherosclerosis. Transl. Res. 2019, 203, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Katsandegwaza, B.; Horsnell, W.; Smith, K. Inflammatory Bowel Disease: A Review of Pre-Clinical Murine Models of Human Disease. Int. J. Mol. Sci. 2022, 23, 9344. [Google Scholar] [CrossRef]
- Wei, H.-X.; Wang, B.; Li, B. IL-10 and IL-22 in Mucosal Immunity: Driving Protection and Pathology. Front. Immunol. 2020, 11, 1315. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Chen, Z.; Xu, Z.; Lv, F.; Zhang, K.; Yang, Y. The Effects of Dihydroartemisinin on Inflammatory Bowel Disease-Related Bone Loss in a Rat Model. Exp. Biol. Med. 2018, 243, 715–724. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Criteria | Grade |
|---|---|
| Diarrhea | |
| No | 0 |
| Yes, with or without blood | 1 |
| Hyperemia/Ulceration | |
| No hyperemia (mucosa was similar to uninflamed intestine) | 0 |
| Localized hyperemia (mild and intense) without ulcers | 1 |
| Extended hyperemia or hemorrhage without ulcers | 2 |
| Ulceration with hyperemia or bowel wall thickening (1–2 mm) | 3 |
| Ulcerations at two or more sites with or without bowel wall thickening > 2 mm | 4 |
| Major sites of damage extended > 1 cm along the colon length | 5 |
| If the damage extends > 2 cm along the colon length, the score was increased by 1 grade for each additional centimeter | 6–10 |
| Adhesion | |
| No adhesions | 0 |
| One adhesion without adhesion to other tissues or organs (colon could be separated from other tissues) | 1 |
| Two or more adhesions or adhesion to tissue or an organ | 2 |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| OPG | 5′TGTGGAATAGATGTCACCCTGTGC3′ | 5′CACAGAGGTCAATGTCTTGGATGATC3′ |
| RANKL | 5′GCTTCTCAGTTCCAGCTATGTT3′ | 5′CGTTGCTTAACGTCATGTTAGAGATCT3′ |
| RANK | 5′GTCTCATCGTCCTGCTCCTCTT′ | 5′CAGCGTTTTCCCTCCCTTC′ |
| TNF-α | 5′ACTCCCAGAAAAGCAAGCAA3′ | 5′TGGAAGACTCCTCCCAGGTA3′ |
| TGF-β | 5′CTCAACACCTGCACAGCTCC3′ | 5′ACGATCATGTTGGACAACTGCT3′ |
| IL-10 | 5′CTTACTGGCTGGAGTGAAGACC3′ | 5′AATCATTCTTCACCTGCTCC3′ |
| GAPDH | 5′GGACCAGGTTGTCTCCTGTG3′ | 5′CATTGAGAGCAATGCCAGC3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Smoljan, I.; Detel, D.; Buljevic, S.; Erjavec, I.; Marić, I. Therapeutic Potential of BMP7 in the Treatment of Osteoporosis Caused by the Interaction between Inflammation and Corticosteroids in Inflammatory Bowel Disease. Biomedicines 2023, 11, 2161. https://doi.org/10.3390/biomedicines11082161
Smoljan I, Detel D, Buljevic S, Erjavec I, Marić I. Therapeutic Potential of BMP7 in the Treatment of Osteoporosis Caused by the Interaction between Inflammation and Corticosteroids in Inflammatory Bowel Disease. Biomedicines. 2023; 11(8):2161. https://doi.org/10.3390/biomedicines11082161
Chicago/Turabian StyleSmoljan, Ivana, Dijana Detel, Suncica Buljevic, Igor Erjavec, and Ivana Marić. 2023. "Therapeutic Potential of BMP7 in the Treatment of Osteoporosis Caused by the Interaction between Inflammation and Corticosteroids in Inflammatory Bowel Disease" Biomedicines 11, no. 8: 2161. https://doi.org/10.3390/biomedicines11082161
APA StyleSmoljan, I., Detel, D., Buljevic, S., Erjavec, I., & Marić, I. (2023). Therapeutic Potential of BMP7 in the Treatment of Osteoporosis Caused by the Interaction between Inflammation and Corticosteroids in Inflammatory Bowel Disease. Biomedicines, 11(8), 2161. https://doi.org/10.3390/biomedicines11082161