
Modelling the Complexity of Human Skin In Vitro

 ,
,  and
and
Abstract
1. Introduction
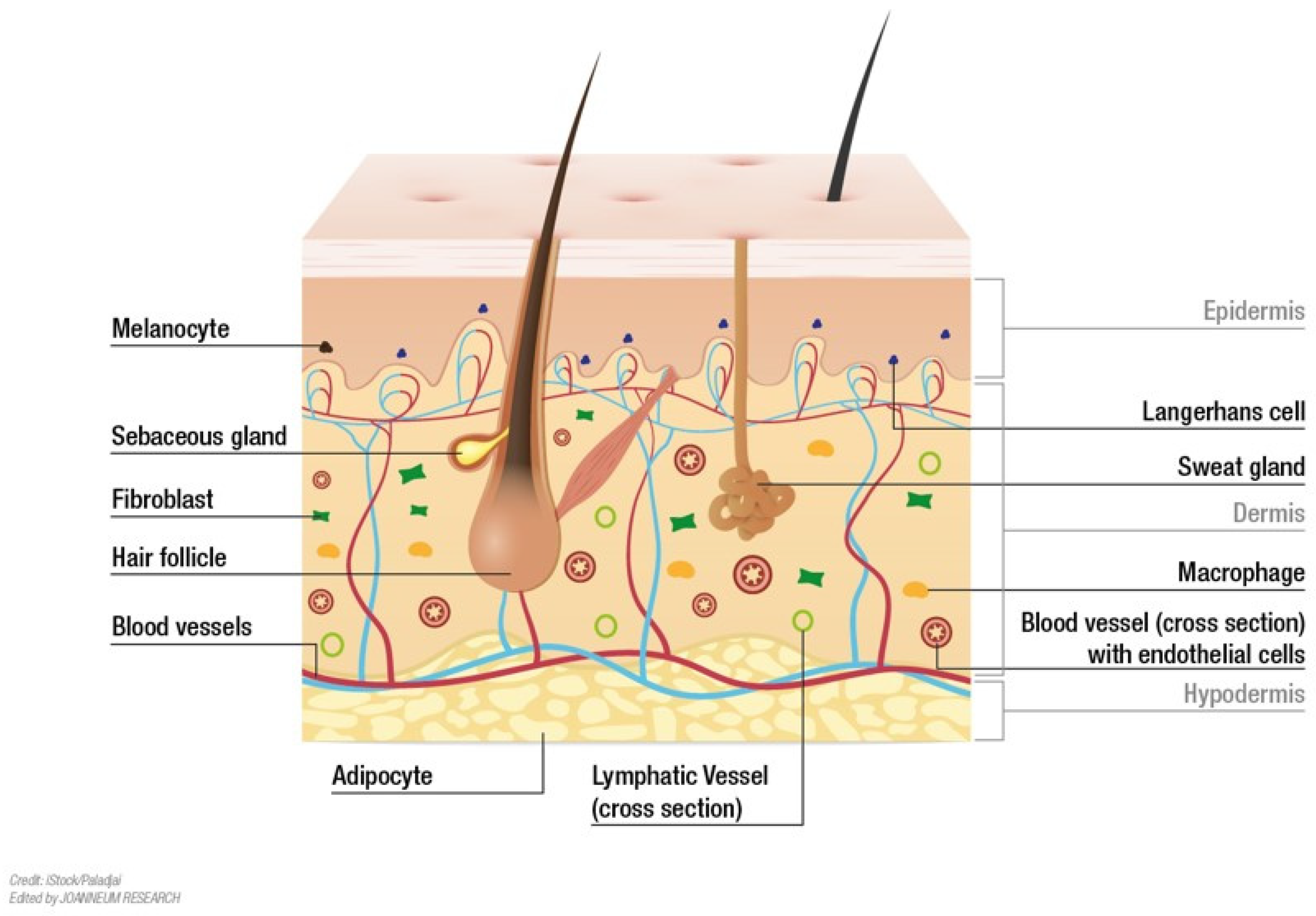
2. Skin structure
2.1. Epidermis
2.2. Basement Membrane
2.3. Dermis
2.4. Hypodermis
3. Reconstructed Human Epidermis
4. Human Skin Equivalents
4.1. Immunocompetence
4.2. Vascularization
4.3. Nervous System
4.4. Adipose Tissue
4.5. Skin Appendages
4.6. 3D Bioprinting of Skin Constructs
5. Limitations of Current In Vitro Skin Models
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Avci, P.; Sadasivam, M.; Gupta, A.; Melo, W.; Huang, Y.-Y.; Yin, R.; Chandran, R.; Kumar, R.; Otufowora, A.; Nyame, T.; et al. Animal models of skin disease for drug discovery. Expert Opin. Drug Discov. 2013, 8, 331–355. [Google Scholar] [CrossRef]
- Jung, E.C.; Maibach, H.I. Animal models for percutaneous absorption. J. Appl. Toxicol. 2015, 35, 1–10. [Google Scholar] [CrossRef]
- Dellambra, E.; Odorisio, T.; D’Arcangelo, D.; Failla, C.M.; Facchiano, A. Non-animal models in dermatological research. ALTEX 2019, 36, 177–202. [Google Scholar] [CrossRef]
- Todo, H. Transdermal Permeation of Drugs in Various Animal Species. Pharmaceutics 2017, 9, 33. [Google Scholar] [CrossRef]
- Sullivan, T.P.; Eaglstein, W.H.; Davis, S.C.; Mertz, P. The pig as a model for human wound healing. Wound Repair Regen. 2001, 9, 66–76. [Google Scholar] [CrossRef]
- Middelkoop, E.; Van Den Bogaerdt, A.J.; Lamme, E.N.; Hoekstra, M.J.; Brandsma, K.; Ulrich, M.M.W. Porcine wound models for skin substitution and burn treatment. Biomaterials 2004, 25, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Summerfield, A.; Meurens, F.; Ricklin, M.E. The immunology of the porcine skin and its value as a model for human skin. Mol. Immunol. 2015, 66, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Leist, M.; Hasiwa, N.; Rovida, C.; Daneshian, M.; Basketter, D.; Kimber, I.; Clewell, H.; Gocht, T.; Goldberg, A.; Busquet, F.; et al. Consensus report on the future of animal-free systemic toxicity testing. Altern. Anim. Exp. ALTEX 2014, 31, 341–356. [Google Scholar] [CrossRef]
- Cosmetics Europe: Cosmetics Directive 76/768/ECC Consolidated Version. Available online: https://cosmeticseurope.eu/files/7714/6400/4401/EU_Cosmetics_Directive_-_Consolidated_version_2004.pdf (accessed on 9 April 2021).
- Proksch, E.; Brandner, J.M.; Jensen, J.-M. The skin: An indispensable barrier. Exp. Dermatol. 2008, 17, 1063–1072. [Google Scholar] [CrossRef]
- Wertz, P.W. Current Understanding of Skin Biology Pertinent to Skin Penetration: Skin Biochemistry. Skin Pharmacol. Physiol. 2013, 26, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Hwang, K. Skin thickness of Korean adults. Surg. Radiol. Anat. 2002, 24, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Honari, G.; Maibach, H. Chapter 1—Skin Structure and Function. In Applied Dermatotoxicology; Maibach, H., Honari, G.B.T.-A.D., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 1–10. ISBN 978-0-12-420130-9. [Google Scholar]
- Nafisi, S.; Maibach, H.I. Chapter 3—Skin penetration of nanoparticles. In Micro and Nano Technologies; Shegokar, R., Souto, E.B.B.T.-E.N., Eds.; Elsevier: Boston, MA, USA, 2018; pp. 47–88. ISBN 978-0-323-40016-9. [Google Scholar]
- Rousselle, P.; Braye, F.; Dayan, G. Re-epithelialization of adult skin wounds: Cellular mechanisms and therapeutic strategies. Adv. Drug Deliv. Rev. 2019, 146, 344–365. [Google Scholar] [CrossRef] [PubMed]
- Houben, E.; De Paepe, K.; Rogiers, V. A Keratinocyte’s Course of Life. Skin Pharmacol. Physiol. 2007, 20, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.S.; Alam, H.; Patil, J.S.; Shrinivasan, R.; Raikundalia, S.; Chaudhari Rajeev, P.; Vaidya, M.M. Keratin 5/14-mediated cell differentiation and transformation are regulated by TAp63 and Notch-1 in oral squamous cell carcinoma-derived cells. Oncol. Rep. 2018, 39, 2393–2401. [Google Scholar] [CrossRef]
- Barbieri, J.S.; Wanat, K.; Seykora, J. Skin: Basic Structure and Function. In Pathobiology of Human Disease; McManus, L.M., Mitchell, R.N.B.T.-P., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 1134–1144. ISBN 978-0-12-386457-4. [Google Scholar]
- Yousef, H.; Alhajj, M.; Sharma, S. Anatomy, Skin (Integument), Epidermis. In StatPearls [Internet]; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Tong, L.; Corrales, R.M.; Chen, Z.; Villarreal, A.L.; De Paiva, C.S.; Beuerman, R.; Li, D.-Q.; Pflugfelder, S.C. Expression and regulation of cornified envelope proteins in human corneal epithelium. Invest. Ophthalmol. Vis. Sci. 2006, 47, 1938–1946. [Google Scholar] [CrossRef] [PubMed]
- Jackson, B.; Tilli, C.M.L.J.; Hardman, M.J.; Avilion, A.A.; MacLeod, M.C.; Ashcroft, G.S.; Byrne, C. Late Cornified Envelope Family in Differentiating Epithelia—Response to Calcium and Ultraviolet Irradiation. J. Investig. Dermatol. 2005, 124, 1062–1070. [Google Scholar] [CrossRef]
- Elias, P.M.; Gruber, R.; Crumrine, D.; Menon, G.; Williams, M.L.; Wakefield, J.S.; Holleran, W.M.; Uchida, Y. Formation and functions of the corneocyte lipid envelope (CLE). Biochim. Biophys. Acta 2014, 1841, 314–318. [Google Scholar] [CrossRef]
- Nemes, Z.; Steinert, P.M. Bricks and mortar of the epidermal barrier. Exp. Mol. Med. 1999, 31, 5–19. [Google Scholar] [CrossRef]
- Cichorek, M.; Wachulska, M.; Stasiewicz, A.; Tymińska, A. Skin melanocytes: Biology and development. Postep. Dermatol. Alergol. 2013, 30, 30–41. [Google Scholar] [CrossRef]
- Abraham, J.; Mathew, S. Merkel Cells: A Collective Review of Current Concepts. Int. J. Appl. basic Med. Res. 2019, 9, 9–13. [Google Scholar] [CrossRef]
- Mathes, S.H.; Ruffner, H.; Graf-Hausner, U. The use of skin models in drug development. Adv. Drug Deliv. Rev. 2014, 69–70, 81–102. [Google Scholar] [CrossRef] [PubMed]
- Stoitzner, P.; Stössel, H.; Romani, N.; Pfaller, K. A Close-Up View of Migrating Langerhans Cells in the Skin. J. Investig. Dermatol. 2002, 118, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, M.; Egawa, G.; Kabashima, K. Uncovering the Mysteries of Langerhans Cells, Inflammatory Dendritic Epidermal Cells, and Monocyte-Derived Langerhans Cell-Like Cells in the Epidermis. Front. Immunol. 2018, 9, 1768. [Google Scholar] [CrossRef]
- Doebel, T.; Voisin, B.; Nagao, K. Langerhans Cells—The Macrophage in Dendritic Cell Clothing. Trends Immunol. 2017, 38, 817–828. [Google Scholar] [CrossRef]
- Fukunaga, A.; Khaskhely, N.M.; Sreevidya, C.S.; Byrne, S.N.; Ullrich, S.E. Dermal dendritic cells, and not Langerhans cells, play an essential role in inducing an immune response. J. Immunol. 2008, 180, 3057–3064. [Google Scholar] [CrossRef] [PubMed]
- Shpichka, A.; Butnaru, D.; Bezrukov, E.A.; Sukhanov, R.B.; Atala, A.; Burdukovskii, V.; Zhang, Y.; Timashev, P. Skin tissue regeneration for burn injury. Stem Cell Res. Ther. 2019, 10, 94. [Google Scholar] [CrossRef]
- Kruegel, J.; Miosge, N. Basement membrane components are key players in specialized extracellular matrices. Cell. Mol. Life Sci. 2010, 67, 2879–2895. [Google Scholar] [CrossRef]
- Borradori, L.; Sonnenberg, A. Structure and Function of Hemidesmosomes: More Than Simple Adhesion Complexes. J. Investig. Dermatol. 1999, 112, 411–418. [Google Scholar] [CrossRef]
- Dos Santos, M.; Michopoulou, A.; André-Frei, V.; Boulesteix, S.; Guicher, C.; Dayan, G.; Whitelock, J.; Damour, O.; Rousselle, P. Perlecan expression influences the keratin 15-positive cell population fate in the epidermis of aging skin. Aging 2016, 8, 751–768. [Google Scholar] [CrossRef]
- Sugawara, K.; Tsuruta, D.; Ishii, M.; Jones, J.C.R.; Kobayashi, H. Laminin-332 and -511 in skin. Exp. Dermatol. 2008, 17, 473–480. [Google Scholar] [CrossRef]
- Abreu-Velez, A.M.; Howard, M.S. Collagen IV in Normal Skin and in Pathological Processes. N. Am. J. Med. Sci. 2012, 4, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Breitkreutz, D.; Koxholt, I.; Thiemann, K.; Nischt, R. Skin basement membrane: The foundation of epidermal integrity—BM functions and diverse roles of bridging molecules nidogen and perlecan. Biomed Res. Int. 2013, 2013, 179784. [Google Scholar] [CrossRef]
- Mortensen, J.H.; Karsdal, M.A. Chapter 7—Type VII Collagen. In Biochemistry of Collagens, Laminins and Elastin; Karsdal, M.A., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 57–60. ISBN 978-0-12-809847-9. [Google Scholar]
- Villone, D.; Fritsch, A.; Koch, M.; Bruckner-Tuderman, L.; Hansen, U.; Bruckner, P. Supramolecular interactions in the dermo-epidermal junction zone: Anchoring fibril-collagen VII tightly binds to banded collagen fibrils. J. Biol. Chem. 2008, 283, 24506–24513. [Google Scholar] [CrossRef] [PubMed]
- Weedon, D. 6—The vesiculobullous reaction pattern. In Weedon’s Skin Pathology Weedon, 3rd ed.; Churchill Livingstone: London, UK, 2010; pp. 123–168.e54. ISBN 978-0-7020-3485-5. [Google Scholar]
- Marinkovich, M.P.; Keene, D.R.; Rimberg, C.S.; Burgeson, R.E. Cellular origin of the dermal-epidermal basement membrane. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1993, 197, 255–267. [Google Scholar] [CrossRef]
- Fleischmajer, R.; Schechter, A.; Bruns, M.; Perlish, J.S.; Macdonald, E.D.; Pan, T.C.; Timpl, R.; Chu, M.L. Skin fibroblasts are the only source of nidogen during early basal lamina formation in vitro. J. Investig. Dermatol. 1995, 105, 597–601. [Google Scholar] [CrossRef]
- Oltulu, P.; Ince, B.; Kökbudak, N.; Fındık, S.; Kiliç, F. Measurement of epidermis, dermis, and total skin thicknesses from six different body regions with a new ethical histometric technique. Turk Plast. Rekonstr. Ve Estet. Cerrahi Derg. 2018, 26, 56–61. [Google Scholar] [CrossRef]
- Wong, R.; Geyer, S.; Weninger, W.; Guimberteau, J.-C.; Wong, J.K. The dynamic anatomy and patterning of skin. Exp. Dermatol. 2016, 25, 92–98. [Google Scholar] [CrossRef]
- Zeng, Q.; Macri, L.K.; Prasad, A.; Clark, R.A.F.; Zeugolis, D.I.; Hanley, C.; Garcia, Y.; Pandit, A.; Leavesley, D.I.; Stupar, D.; et al. 6.20 Skin Tissue Engineering. In Comprehensive Biomaterials II; Ducheyne, P., Ed.; Elsevier: Oxford, UK, 2017; pp. 334–382. ISBN 978-0-08-100692-4. [Google Scholar]
- Reihsner, R.; Balogh, B.; Menzel, E.J. Two-dimensional elastic properties of human skin in terms of an incremental model at the in vivo configuration. Med. Eng. Phys. 1995, 17, 304–313. [Google Scholar] [CrossRef]
- Chermnykh, E.; Kalabusheva, E.; Vorotelyak, E. Extracellular Matrix as a Regulator of Epidermal Stem Cell Fate. Int. J. Mol. Sci. 2018, 19, 1003. [Google Scholar] [CrossRef]
- Nguyen, A.V.; Soulika, A.M. The dynamics of the skin’s immune system. Int. J. Mol. Sci. 2019, 20, 1811. [Google Scholar] [CrossRef]
- Haniffa, M.; Gunawan, M.; Jardine, L. Human skin dendritic cells in health and disease. J. Dermatol. Sci. 2015, 77, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast cell: A multi-functional master cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef] [PubMed]
- Benyon, R.C. The human skin mast cell. Clin. Exp. Allergy 1989, 19, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.S.; Diamond, A.; Russell, A.; Jameson, J.M. Human αβ and γδ T cells in skin immunity and disease. Front. Immunol. 2018, 9, 1304. [Google Scholar] [CrossRef]
- Weng, T.; Wu, P.; Zhang, W.; Zheng, Y.; Li, Q.; Jin, R.; Chen, H.; You, C.; Guo, S.; Han, C.; et al. Regeneration of skin appendages and nerves: Current status and further challenges. J. Transl. Med. 2020, 18, 53. [Google Scholar] [CrossRef] [PubMed]
- Klar, A.S.; Zimoch, J.; Biedermann, T. Skin Tissue Engineering: Application of Adipose-Derived Stem Cells. Biomed Res. Int. 2017, 2017, 9747010. [Google Scholar] [CrossRef]
- Ramakrishnan, V.M.; Boyd, N.L. The Adipose Stromal Vascular Fraction as a Complex Cellular Source for Tissue Engineering Applications. Tissue Eng. Part B Rev. 2018, 24, 289–299. [Google Scholar] [CrossRef]
- Diegel, K.L.; Danilenko, D.M.; Wojcinski, Z.W. Chapter 24—The Integumentary System. In Fundamentals of Toxicologic Pathology, 3rd ed.; Wallig, M.A., Haschek, W.M., Rousseaux, C.G., Bolon, B., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 791–822. ISBN 978-0-12-809841-7. [Google Scholar]
- Kershaw, E.E.; Flier, J.S. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef]
- Prunieras, M.; Regnier, M.; Woodley, D. Methods for Cultivation of Keratinocytes with an Air-Liquid Interface. J. Investig. Dermatol. 1983, 81, S28–S33. [Google Scholar] [CrossRef]
- Asselineau, D.; Bernard, B.A.; Bailly, C.; Darmon, M.; Pruniéras, M. Human Epidermis Reconstructed by Culture: Is It “Normal”? J. Investig. Dermatol. 1986, 86, 181–186. [Google Scholar] [CrossRef]
- Rosdy, M.; Pisani, A.; Ortonne, J.P. Production of basement membrane components by a reconstructed epidermis cultured in the absence of serum and dermal factors. Br. J. Dermatol. 1993, 129, 227–234. [Google Scholar] [CrossRef]
- Rosdy, M.; Clauss, L.C. Terminal epidermal differentiation of human keratinocytes grown in chemically defined medium on inert filter substrates at the air-liquid interface. J. Investig. Dermatol. 1990, 95, 409–414. [Google Scholar] [CrossRef]
- Ponec, M.; Boelsma, E.; Weerheim, A.; Mulder, A.; Bouwstra, J.; Mommaas, M. Lipid and Ultrastructural Characterization of Reconstructed Skin Models; Elsevier: Amsterdam, The Netherlands, 2000; Volume 203, pp. 211–225. [Google Scholar]
- Gazel, A.; Ramphal, P.; Rosdy, M.; De Wever, B.; Tornier, C.; Hosein, N.; Lee, B.; Tomic-Canic, M.; Blumenberg, M. Transcriptional Profiling of Epidermal Keratinocytes: Comparison of Genes Expressed in Skin, Cultured Keratinocytes, and Reconstituted Epidermis, Using Large DNA Microarrays. J. Investig. Dermatol. 2003, 121, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Sriram, G.; Lorenz Bigliardi, P.; Bigliardi-Qi, M. Fibroblast heterogeneity and its implications for engineering organotypic skin models in vitro. Eur. J. Cell Biol. 2015, 94, 483–512. [Google Scholar] [CrossRef] [PubMed]
- Boehnke, K.; Mirancea, N.; Pavesio, A.; Fusenig, N.E.; Boukamp, P.; Stark, H.-J.J. Effects of fibroblasts and microenvironment on epidermal regeneration and tissue function in long-term skin equivalents. Eur. J. Cell Biol. 2007, 86, 731–746. [Google Scholar] [CrossRef] [PubMed]
- El Ghalbzouri, A.; Lamme, E.; Ponec, M. Crucial role of fibroblasts in regulating epidermal morphogenesis. Cell Tissue Res. 2002, 310, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Michel, M.; Uheureux, N.; Pouliot, R.; Xu, W.; Auger, F.A.; Germain, L. Characterization of a new tissue-engineered human skin equivalent with hair. Vitro Cell. Dev. Biol. Anim. 1999, 35, 318–326. [Google Scholar] [CrossRef]
- Alépée, N.; Bahinski, A.; Daneshian, M.; De Wever, B.; Fritsche, E.; Goldberg, A.; Hansmann, J.; Hartung, T.; Haycock, J.; Hogberg, H.T.; et al. State-of-the-art of 3D cultures (organs-on-a-chip) in safety testing and pathophysiology. ALTEX 2014, 31, 441–477. [Google Scholar] [CrossRef]
- Mewes, K.R.; Fischer, A.; Zöller, N.N.; Laubach, V.; Bernd, A.; Jacobs, A.; van Rompay, A.; Liebsch, M.; Pirow, R.; Petersohn, D. Catch-up validation study of an in vitro skin irritation test method based on an open source reconstructed epidermis (phase I). Toxicol. Vitr. 2016, 36, 238–253. [Google Scholar] [CrossRef]
- Groeber, F.; Schober, L.; Schmid, F.F.; Traube, A.; Kolbus-Hernandez, S.; Daton, K.; Hoffmann, S.; Petersohn, D.; Schäfer-Korting, M.; Walles, H.; et al. Catch-up validation study of an in vitro skin irritation test method based on an open source reconstructed epidermis (phase II). Toxicol. Vitr. 2016, 36, 254–261. [Google Scholar] [CrossRef]
- Poumay, Y.; Dupont, F.; Marcoux, S.; Leclercq-Smekens, M.; Hérin, M.; Coquette, A. A simple reconstructed human epidermis: Preparation of the culture model and utilization in in vitro studies. Arch. Dermatol. Res. 2004, 296, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Suwa, F.; Wang, X.; Takemura, A.; Fang, Y.R.; Li, Y.; Zhao, Y.; Jin, Y. Reconstruction of a tissue-engineered skin containing melanocytes. Cell Biol. Int. 2007, 31, 985–990. [Google Scholar] [CrossRef]
- Schmid, F.; Groeber-Becker, F.; Schwab, S.; Thude, S.; Goebeler, M.; Walles, H.; Hansmann, J. A standardized method based on pigmented epidermal models evaluates sensitivity against UV-irradiation. ALTEX 2018, 35, 390–396. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, J.E.; Kim, B.J.; Cho, K.H. In vitro phototoxicity test using artificial skin with melanocytes. Photodermatol. Photoimmunol. Photomed. 2007, 23, 73–80. [Google Scholar] [CrossRef]
- Contard, P.; Bartel, R.L.; Jacobs, L.; Perlish, J.S.; MacDonald, E.D.; Handler, L.; Cone, D.; Fleischmajer, R. Culturing Keratinocytes and Fibroblasts in a Three-Dimensional Mesh Results in Epidermal Differentiation and Formation of a Basal Lamina-Anchoring Zone. J. Investig. Dermatol. 1993, 100, 35–39. [Google Scholar] [CrossRef]
- El Ghalbzouri, A.; Jonkman, M.F.; Dijkman, R.; Ponec, M. Basement Membrane Reconstruction in Human Skin Equivalents Is Regulated by Fibroblasts and/or Exogenously Activated Keratinocytes. J. Investig. Dermatol. 2005, 124, 79–86. [Google Scholar] [CrossRef]
- Parenteau-Bareil, R.; Gauvin, R.; Berthod, F. Collagen-based biomaterials for tissue engineering applications. Materials 2010, 3, 1863–1887. [Google Scholar] [CrossRef]
- Sarrigiannidis, S.O.; Rey, J.M.; Dobre, O.; González-García, C.; Dalby, M.J.; Salmeron-Sanchez, M. A tough act to follow: Collagen hydrogel modifications to improve mechanical and growth factor loading capabilities. Mater. Today Bio 2021, 10, 100098. [Google Scholar] [CrossRef]
- Randall, M.J.; Jüngel, A.; Rimann, M.; Wuertz-Kozak, K. Advances in the Biofabrication of 3D Skin in vitro: Healthy and Pathological Models. Front. Bioeng. Biotechnol. 2018, 6, 154. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.E.L.; Tamamoto, K.A.; Nguyen, H.; Abbott, R.D.; Cairns, D.M.; Kaplan, D.L. 3D biomaterial matrix to support long term, full thickness, immuno-competent human skin equivalents with nervous system components. Biomaterials 2019, 198, 194–203. [Google Scholar] [CrossRef]
- Bellas, E.; Seiberg, M.; Garlick, J.; Kaplan, D.L. In Vitro 3D Full-Thickness Skin-Equivalent Tissue Model Using Silk and Collagen Biomaterials. Macromol. Biosci. 2012, 12, 1627–1636. [Google Scholar] [CrossRef]
- Gil, E.S.; Panilaitis, B.; Bellas, E.; Kaplan, D.L. Functionalized Silk Biomaterials for Wound Healing. Adv. Healthc. Mater. 2013, 2, 206–217. [Google Scholar] [CrossRef]
- Rosa, E.; Diaferia, C.; Gianolio, E.; Sibillano, T.; Gallo, E.; Smaldone, G.; Stornaiuolo, M.; Giannini, C.; Morelli, G.; Accardo, A. Multicomponent Hydrogel Matrices of Fmoc-FF and Cationic Peptides for Application in Tissue Engineering. Macromol. Biosci. 2022, 22, 2200128. [Google Scholar] [CrossRef]
- Arab, W.T.; Susapto, H.H.; Alhattab, D.; Hauser, C.A.E. Peptide nanogels as a scaffold for fabricating dermal grafts and 3D vascularized skin models. J. Tissue Eng. 2022, 13, 20417314221111868. [Google Scholar] [CrossRef]
- Bergers, L.I.J.C.; Reijnders, C.M.A.; Van Den Broek, L.J.; Spiekstra, S.W.; De Gruijl, T.D.; Weijers, E.M.; Gibbs, S. Immune-competent human skin disease models. Drug Discov. Today 2016, 21, 1479–1488. [Google Scholar] [CrossRef]
- Pupovac, A.; Senturk, B.; Griffoni, C.; Maniura-Weber, K.; Rottmar, M.; McArthur, S.L. Toward Immunocompetent 3D Skin Models. Adv. Healthc. Mater. 2018, 7, 1701405. [Google Scholar] [CrossRef]
- Ouwehand, K.; Spiekstra, S.W.; Waaijman, T.; Scheper, R.J.; de Gruijl, T.D.; Gibbs, S. Technical Advance: Langerhans cells derived from a human cell line in a full-thickness skin equivalent undergo allergen-induced maturation and migration. J. Leukoc. Biol. 2011, 90, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Kühbacher, A.; Henkel, H.; Stevens, P.; Grumaz, C.; Finkelmeier, D.; Burger-Kentischer, A.; Sohn, K.; Rupp, S. Central Role for Dermal Fibroblasts in Skin Model Protection against Candida albicans. J. Infect. Dis. 2017, 215, 1742–1752. [Google Scholar] [CrossRef] [PubMed]
- Chau, D.Y.S.; Johnson, C.; Macneil, S.; Haycock, J.W.; Ghaemmaghami, A.M. The development of a 3D immunocompetent model of human skin. Biofabrication 2013, 5, 035011. [Google Scholar] [CrossRef] [PubMed]
- Bechetoille, N.; Dezutter-Dambuyant, C.; Damour, O.; André, V.; Orly, I.; Perrier, E. Effects of Solar Ultraviolet Radiation on Engineered Human Skin Equivalent Containing Both Langerhans Cells and Dermal Dendritic Cells. Tissue Eng. 2007, 13, 2667–2679. [Google Scholar] [CrossRef]
- Schellenberger, M.T.; Bock, U.; Hennen, J.; Groeber-Becker, F.; Walles, H.; Blömeke, B. A coculture system composed of THP-1 cells and 3D reconstructed human epidermis to assess activation of dendritic cells by sensitizing chemicals after topical exposure. Toxicol. Vitr. 2019, 57, 62–66. [Google Scholar] [CrossRef]
- Griffoni, C.; Neidhart, B.; Yang, K.; Groeber-Becker, F.; Maniura-Weber, K.; Dandekar, T.; Walles, H.; Rottmar, M. In vitro skin culture media influence the viability and inflammatory response of primary macrophages. Sci. Rep. 2021, 11, 7070. [Google Scholar] [CrossRef]
- Black, A.F.; Berthod, F.; L’Heureux, N.; Germain, L.; Auger, F.A. In vitro reconstruction of a human capillary-like network in a tissue-engineered skin equivalent. FASEB J. 1998, 12, 1331–1340. [Google Scholar] [CrossRef]
- Ponec, M.; El Ghalbzouri, A.; Dijkman, R.; Kempenaar, J.; Van Der Pluijm, G.; Koolwijk, P. Endothelial network formed with human dermal microvascular endothelial cells in autologous multicellular skin substitutes. Angiogenesis 2004, 7, 295–305. [Google Scholar] [CrossRef]
- Frueh, F.S.; Später, T.; Lindenblatt, N.; Calcagni, M.; Giovanoli, P.; Scheuer, C.; Menger, M.D.; Laschke, M.W. Adipose Tissue-Derived Microvascular Fragments Improve Vascularization, Lymphangiogenesis, and Integration of Dermal Skin Substitutes. J. Investig. Dermatol. 2017, 137, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Frueh, F.S.; Später, T.; Körbel, C.; Scheuer, C.; Simson, A.C.; Lindenblatt, N.; Giovanoli, P.; Menger, M.D.; Laschke, M.W. Prevascularization of dermal substitutes with adipose tissue-derived microvascular fragments enhances early skin grafting. Sci. Rep. 2018, 8, 10977. [Google Scholar] [CrossRef] [PubMed]
- Matei, A.E.; Chen, C.W.; Kiesewetter, L.; Györfi, A.H.; Li, Y.N.; Trinh-Minh, T.; Xu, X.; Tran Manh, C.; Van Kuppevelt, T.; Hansmann, J.; et al. Vascularised human skin equivalents as a novel in vitro model of skin fibrosis and platform for testing of antifibrotic drugs. Ann. Rheum. Dis. 2019, 78, 1686–1692. [Google Scholar] [CrossRef] [PubMed]
- Abaci, H.E.; Guo, Z.; Coffman, A.; Gillette, B.; Lee, W.H.; Sia, S.K.; Christiano, A.M. Human Skin Constructs with Spatially Controlled Vasculature Using Primary and iPSC-Derived Endothelial Cells. Adv. Healthc. Mater. 2016, 5, 1800–1807. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.L.; Wang, S.; Yeong, W.Y.; Naing, M.W. Skin Bioprinting: Impending Reality or Fantasy? Trends Biotechnol. 2016, 34, 689–699. [Google Scholar] [CrossRef]
- Roggenkamp, D.; Köpnick, S.; Stäb, F.; Wenck, H.; Schmelz, M.; Neufang, G. Epidermal Nerve Fibers Modulate Keratinocyte Growth via Neuropeptide Signaling in an Innervated Skin Model. J. Investig. Dermatol. 2013, 133, 1620–1628. [Google Scholar] [CrossRef]
- Blais, M.; Mottier, L.; Germain, M.-A.; Bellenfant, S.; Cadau, S.; Berthod, F. Sensory Neurons Accelerate Skin Reepithelialization via Substance P in an Innervated Tissue-Engineered Wound Healing Model. Tissue Eng. Part A 2014, 20, 2180–2188. [Google Scholar] [CrossRef]
- Cairns, D.M.; Chwalek, K.; Moore, Y.E.; Kelley, M.R.; Abbott, R.D.; Moss, S.; Kaplan, D.L. Expandable and Rapidly Differentiating Human Induced Neural Stem Cell Lines for Multiple Tissue Engineering Applications. Stem. Cell Rep. 2016, 7, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Gimble, J.M.; Lee, K.; Marra, K.G.; Rubin, J.P.; Yoo, J.J.; Vunjak-Novakovic, G.; Kaplan, D.L. Adipose tissue engineering for soft tissue regeneration. Tissue Eng. Part B Rev. 2010, 16, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Asuku, M.; Yu, T.-C.; Yan, Q.; Böing, E.; Hahn, H.; Hovland, S.; Donelan, M.B. Split-thickness skin graft donor-site morbidity: A systematic literature review. Burns 2021, 47, 1525–1546. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.F.; Sousa, A.; Barrias, C.C.; Bayat, A.; Granja, P.L.; Bártolo, P.J. Advances in bioprinted cell-laden hydrogels for skin tissue engineering. Biomanuf. Rev. 2017, 2, 1–26. [Google Scholar] [CrossRef]
- Trottier, V.; Marceau-Fortier, G.; Germain, L.; Vincent, C.; Fradette, J. IFATS Collection: Using Human Adipose-Derived Stem/Stromal Cells for the Production of New Skin Substitutes. Stem Cells 2008, 26, 2713–2723. [Google Scholar] [CrossRef]
- Monfort, A.; Soriano-Navarro, M.; García-Verdugo, J.M.; Izeta, A. Production of human tissue-engineered skin trilayer on a plasma-based hypodermis. J. Tissue Eng. Regen. Med. 2013, 7, 479–490. [Google Scholar] [CrossRef] [PubMed]
- von Heimburg, D.; Hemmrich, K.; Zachariah, S.; Staiger, H.; Pallua, N. Oxygen consumption in undifferentiated versus differentiated adipogenic mesenchymal precursor cells. Respir. Physiol. Neurobiol. 2005, 146, 107–116. [Google Scholar] [CrossRef]
- Gimble, J.M.; Katz, A.J.; Bunnell, B.A. Adipose-derived stem cells for regenerative medicine. Circ. Res. 2007, 100, 1249–1260. [Google Scholar] [CrossRef]
- Satija, N.K.; Gurudutta, G.U.; Sharma, S.; Afrin, F.; Gupta, P.; Verma, Y.K.; Singh, V.K.; Tripathi, R.P. Mesenchymal stem cells: Molecular targets for tissue engineering. Stem Cells Dev. 2007, 16, 7–23. [Google Scholar] [CrossRef]
- Huber, B.; Link, A.; Linke, K.; Gehrke, S.A.; Winnefeld, M.; Kluger, P.J. Integration of Mature Adipocytes to Build-Up a Functional Three-Layered Full-Skin Equivalent. Tissue Eng.—Part C Methods 2016, 22, 756–764. [Google Scholar] [CrossRef]
- Huber, B.; Kluger, P.J. Decelerating Mature Adipocyte Dedifferentiation by Media Composition. Tissue Eng. Part C Methods 2015, 21, 1237–1245. [Google Scholar] [CrossRef]
- Huber, B.; Borchers, K.; Tovar, G.E.M.; Kluger, P.J. Methacrylated gelatin and mature adipocytes are promising components for adipose tissue engineering. J. Biomater. Appl. 2015, 30, 699–710. [Google Scholar] [CrossRef]
- Kober, J.; Gugerell, A.; Schmid, M.; Kamolz, L.P.; Keck, M. Generation of a Fibrin Based Three-Layered Skin Substitute. Biomed Res. Int. 2015, 2015, 170427. [Google Scholar] [CrossRef]
- Ataç, B.; Wagner, I.; Horland, R.; Lauster, R.; Marx, U.; Tonevitsky, A.G.; Azar, R.P.; Lindner, G. Skin and hair on-a-chip: In vitro skin models versus ex vivo tissue maintenance with dynamic perfusion. Lab. Chip 2013, 13, 3555–3561. [Google Scholar] [CrossRef]
- Ojeh, N.; Pastar, I.; Tomic-Canic, M.; Stojadinovic, O. Stem cells in skin regeneration, wound healing, and their clinical applications. Int. J. Mol. Sci. 2015, 16, 25476–25501. [Google Scholar] [CrossRef]
- Langan, E.A.; Philpott, M.P.; Kloepper, J.E.; Paus, R. Human hair follicle organ culture: Theory, application and perspectiDes. Exp. Dermatol. 2015, 24, 200–215. [Google Scholar] [CrossRef]
- Ghetti, M.; Topouzi, H.; Theocharidis, G.; Papa, V.; Williams, G.; Bondioli, E.; Cenacchi, G.; Connelly, J.T.; Higgins, C.A. Subpopulations of dermal skin fibroblasts secrete distinct extracellular matrix: Implications for using skin substitutes in the clinic. Br. J. Dermatol. 2018, 179, 381–393. [Google Scholar] [CrossRef]
- Higgins, C.A.; Chen, J.C.; Cerise, J.E.; Jahoda, C.A.B.; Christiano, A.M. Microenvironmental reprogramming by threedimensional culture enables dermal papilla cells to induce de novo human hair-follicle growth. Proc. Natl. Acad. Sci. USA 2013, 110, 19679–19688. [Google Scholar] [CrossRef]
- Topouzi, H.; Logan, N.J.; Williams, G.; Higgins, C.A.; Claire Higgins, C.A. Methods for the isolation and 3D culture of dermal papilla cells from human hair follicles. Exp. Dermatol. 2017, 26, 491–496. [Google Scholar] [CrossRef]
- Abaci, H.E.; Guo, Z.; Doucet, Y.; Jacków, J.; Christiano, A. Next generation human skin constructs as advanced tools for drug development. Exp. Biol. Med. 2017, 242, 1657–1668. [Google Scholar] [CrossRef]
- Abaci, H.E.; Coffman, A.; Doucet, Y.; Chen, J.; Jacków, J.; Wang, E.; Guo, Z.; Shin, J.U.; Jahoda, C.A.; Christiano, A.M. Tissue engineering of human hair follicles using a biomimetic developmental approach. Nat. Commun. 2018, 9, 5301. [Google Scholar] [CrossRef]
- Matai, I.; Kaur, G.; Seyedsalehi, A.; McClinton, A.; Laurencin, C.T. Progress in 3D bioprinting technology for tissue/organ regenerative engineering. Biomaterials 2020, 226. [Google Scholar] [CrossRef]
- Murphy, S.V.; Atala, A. 3D bioprinting of tissues and organs. Nat. Biotechnol. 2014, 32, 773–785. [Google Scholar] [CrossRef]
- Lee, W.; Debasitis, J.C.; Lee, V.K.; Lee, J.H.; Fischer, K.; Edminster, K.; Park, J.K.; Yoo, S.S. Multi-layered culture of human skin fibroblasts and keratinocytes through three-dimensional freeform fabrication. Biomaterials 2009, 30, 1587–1595. [Google Scholar] [CrossRef]
- Koch, L.; Kuhn, S.; Sorg, H.; Gruene, M.; Schlie, S.; Gaebel, R.; Polchow, B.; Reimers, K.; Stoelting, S.; Ma, N.; et al. Laser Printing of Skin Cells and Human Stem Cells. Tissue Eng. Part C Methods 2009, 16, 847–854. [Google Scholar] [CrossRef]
- Olejnik, A.; Semba, J.A.; Kulpa, A.; Dańczak-Pazdrowska, A.; Rybka, J.D.; Gornowicz-Porowska, J. 3D Bioprinting in Skin Related Research: Recent Achievements and Application Perspectives. ACS Synth. Biol. 2022, 11, 26–38. [Google Scholar] [CrossRef]
- Manita, P.G.; Garcia-orue, I.; Santos-vizcaino, E.; Hernandez, R.M.; Igartua, M. 3D Bioprinting of Functional Skin Substitutes: From Current Achievements to Future Goals. Pharmaceuticals 2021, 14, 362. [Google Scholar] [CrossRef]
- Pourchet, L.J.; Thepot, A.; Albouy, M.; Courtial, E.J.; Boher, A.; Blum, L.J.; Marquette, C.A. Human Skin 3D Bioprinting Using Scaffold-Free Approach. Adv. Healthc. Mater. 2017, 6, 1601101. [Google Scholar] [CrossRef]
- Lee, V.; Singh, G.; Trasatti, J.P.; Bjornsson, C.; Xu, X.; Tran, T.N.; Yoo, S.S.; Dai, G.; Karande, P. Design and Fabrication of Human Skin by Three-Dimensional Bioprinting. Tissue Eng. Part C Methods 2014, 20, 473. [Google Scholar] [CrossRef]
- Ashammakhi, N.; Ahadian, S.; Xu, C.; Montazerian, H.; Ko, H.; Nasiri, R.; Barros, N.; Khademhosseini, A. Bioinks and bioprinting technologies to make heterogeneous and biomimetic tissue constructs. Mater. Today Bio. 2019, 1, 100008. [Google Scholar] [CrossRef]
- Velasco, D.; Quílez, C.; Garcia, M.; del Cañizo, J.F.; Jorcano, J.L. 3D human skin bioprinting: A view from the bio side. J. 3D Print. Med. 2018, 2, 141–162. [Google Scholar] [CrossRef]
- Parak, A.; Pradeep, P.; du Toit, L.C.; Kumar, P.; Choonara, Y.E.; Pillay, V. Functionalizing bioinks for 3D bioprinting applications. Drug Discov. Today 2019, 24, 198–205. [Google Scholar] [CrossRef]
- Williams, D.; Thayer, P.; Martinez, H.; Gatenholm, E.; Khademhosseini, A. A perspective on the physical, mechanical and biological specifications of bioinks and the development of functional tissues in 3D bioprinting. Bioprinting 2018, 9, 19–36. [Google Scholar] [CrossRef]
- Gopinathan, J.; Noh, I. Recent trends in bioinks for 3D printing. Biomater. Res. 2018, 22, 11. [Google Scholar] [CrossRef]
- Perez-Valle, A.; Del Amo, C.; Andia, I. Overview of Current Advances in Extrusion Bioprinting for Skin Applications. Int. J. Mol. Sci. 2020, 21, 6679. [Google Scholar] [CrossRef]
- Kim, B.S.; Gao, G.; Kim, J.Y.; Cho, D. 3D Cell Printing of Perfusable Vascularized Human Skin Equivalent Composed of Epidermis, Dermis, and Hypodermis for Better Structural Recapitulation of Native Skin. Adv. Healthc. Mater. 2019, 8, 1801019. [Google Scholar] [CrossRef]
- Baltazar, T.; Merola, J.; Catarino, C.; Xie, C.B.; Kirkiles-Smith, N.C.; Lee, V.; Hotta, S.; Dai, G.; Xu, X.; Ferreira, F.C.; et al. Three Dimensional Bioprinting of a Vascularized and Perfusable Skin Graft Using Human Keratinocytes, Fibroblasts, Pericytes, and Endothelial Cells. Tissue Eng. Part A 2020, 26, 227. [Google Scholar] [CrossRef]
- Liu, X.; Michael, S.; Bharti, K.; Ferrer, M.; Song, M.J. A biofabricated vascularized skin model of atopic dermatitis for preclinical studies. Biofabrication 2020, 12, 035002. [Google Scholar] [CrossRef]
- Groeber, F.; Engelhardt, L.; Lange, J.; Kurdyn, S.; Schmid, F.F.; Rücker, C.; Mielke, S.; Walles, H.; Hansmann, J. A first vascularized skin equivalent for as an alternative to animal experimentation. ALTEX 2016, 33, 415–422. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type | Brand Name (Company) | Cell Types | Scaffold | Validated for |
|---|---|---|---|---|
| RHE | SkinEthic™ RHE (Episkin, Lyon, France) | NHEK | Polycarbonate filter | Skin irritation test (EC TMR B.46; OECD TGL 439) |
| EpiSkin™ (Episkin, Lyon, France) | NHEK | Collagen | Skin irritation test (EU TMR 440/2008/EC; OECD TGL 439) | |
| EpiDerm™ (MatTek Corporation, Ashland, MA, USA) | NHEK | Collagen coated polycarbonate membrane | Skin Corrosion (OECD TGL 431), Skin Irritation (OECD TGL 439), Phototoxicity (OECD TGL 498) | |
| EpiCS® (Phenion, Düsseldorf, Germany) | NHEK | Polycarbonate membrane | Skin Corrosion (OECD TGL 431), Skin Irritation (OECD TGL 439) | |
| ZenSkin RHE model (Zen-Bio, Inc., Durham, NC, USA) | NHEK | Polycarbonate filter | - | |
| LabCyte EPI-MODEL (J-TEC, Ltd., Osaka, Japan) | NHEK | Filter insert | - | |
| KeraSkin™ (Biosolution Co., Ltd., Seoul, Republic of Korea) | NHEK | Unknown | - | |
| Skin+™ (Sterlab, Saint Bernard, France) | NHEK | Unknown | - | |
| pigmented RHE | SkinEthic™ (Episkin, Lyon, France) | NHEK; Melanocytes of different phototypes | Polycarbonate filter | - |
| MelanoDerm™ (MatTek Corporation, Ashland, USA) | NHEK; Melanocytes of different phototypes | Filter insert | - | |
| EpiCS®-M (Phenion, RHPE, Düsseldorf, Germany) | NHEK; Melanocytes of different phototypes | Polycarbonate membrane | - | |
| MEL/001 (StratiCELL, Gembloux, Belgium) | NHEK; Melanocytes of different phototypes | Polycarbonate filter | - | |
| KeraSkin-M™ (Biosolution Co., Ltd., Seoul, South Korea) | NHEK; Melanocytes | Unknown | - | |
| RHEP (Sterlab, Saint Bernard, France) | NHEK; Melanocytes of different phototypes | Unknown | - | |
| HSE | T-Skin™ (Episkin, Lyon, France) | NHEK; human fibroblasts | Dermal equivalent | - |
| Full Thickness model (Sterlab, Saint Bernard, France) | NHEK; human fibroblast | Collagen | - | |
| EpiDermFT™ (MatTek Corporation, Ashland, USA) | NHEK; NHDF | Collagen | - | |
| Phenion® FT (Henkel AG & Co. KgaA, Düsseldorf, Germany) | NHEK; human fibroblasts | ECM proteins | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hofmann, E.; Schwarz, A.; Fink, J.; Kamolz, L.-P.; Kotzbeck, P. Modelling the Complexity of Human Skin In Vitro. Biomedicines 2023, 11, 794. https://doi.org/10.3390/biomedicines11030794
Hofmann E, Schwarz A, Fink J, Kamolz L-P, Kotzbeck P. Modelling the Complexity of Human Skin In Vitro. Biomedicines. 2023; 11(3):794. https://doi.org/10.3390/biomedicines11030794
Chicago/Turabian StyleHofmann, Elisabeth, Anna Schwarz, Julia Fink, Lars-Peter Kamolz, and Petra Kotzbeck. 2023. "Modelling the Complexity of Human Skin In Vitro" Biomedicines 11, no. 3: 794. https://doi.org/10.3390/biomedicines11030794
APA StyleHofmann, E., Schwarz, A., Fink, J., Kamolz, L.-P., & Kotzbeck, P. (2023). Modelling the Complexity of Human Skin In Vitro. Biomedicines, 11(3), 794. https://doi.org/10.3390/biomedicines11030794