
Obesity Correlates with Chronic Inflammation of the Innate Immune System in Preeclampsia and HELLP Syndrome during Pregnancy
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
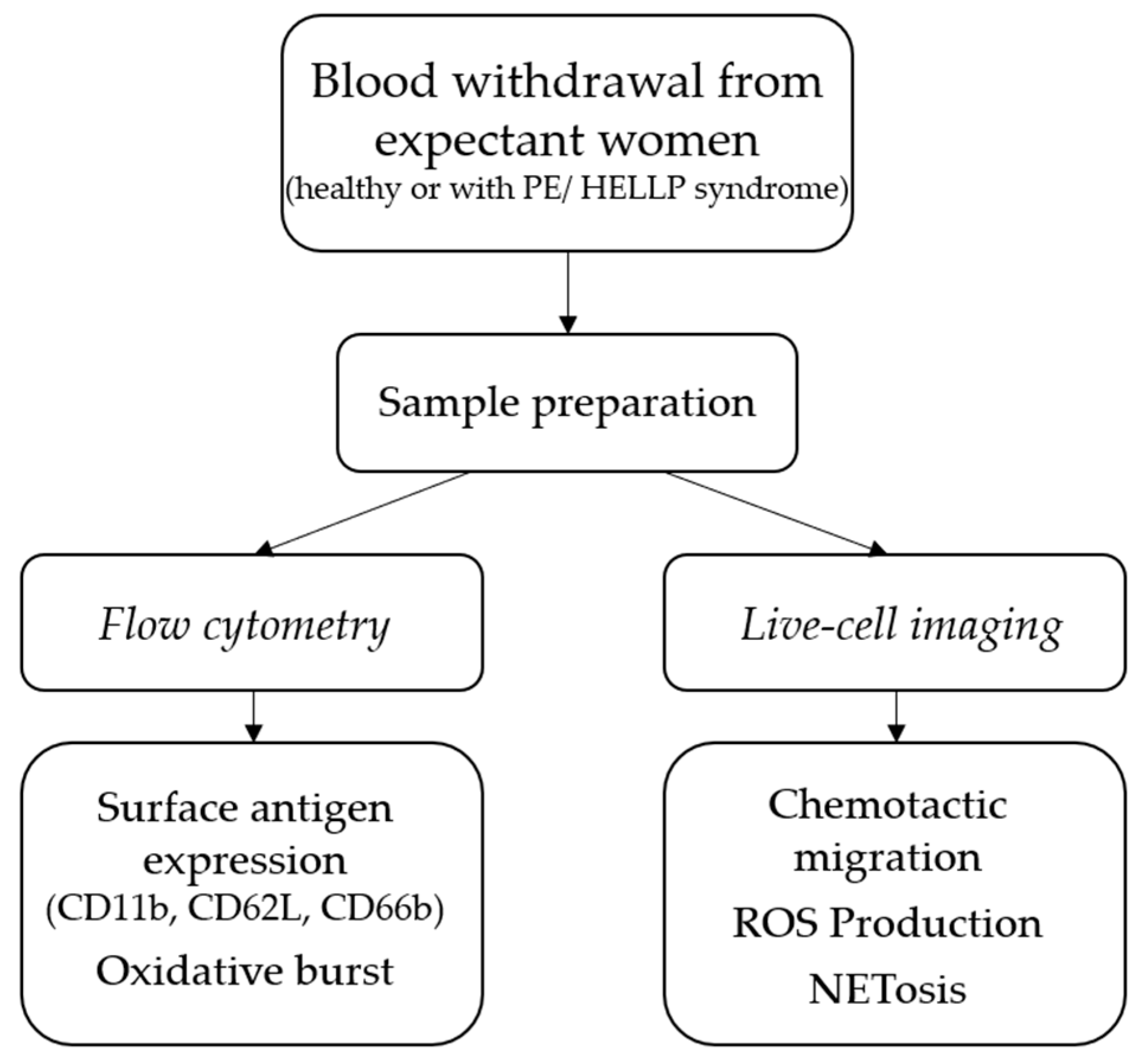
2.1. Study Plan
2.2. Blood Withdrawal and PMN Isolation
2.3. Flow Cytometry
2.3.1. Surface Antigen Expression
2.3.2. Oxidative Burst
2.3.3. Flow Cytometric Measurement and Software Data Analysis
2.4. Live-Cell Imaging
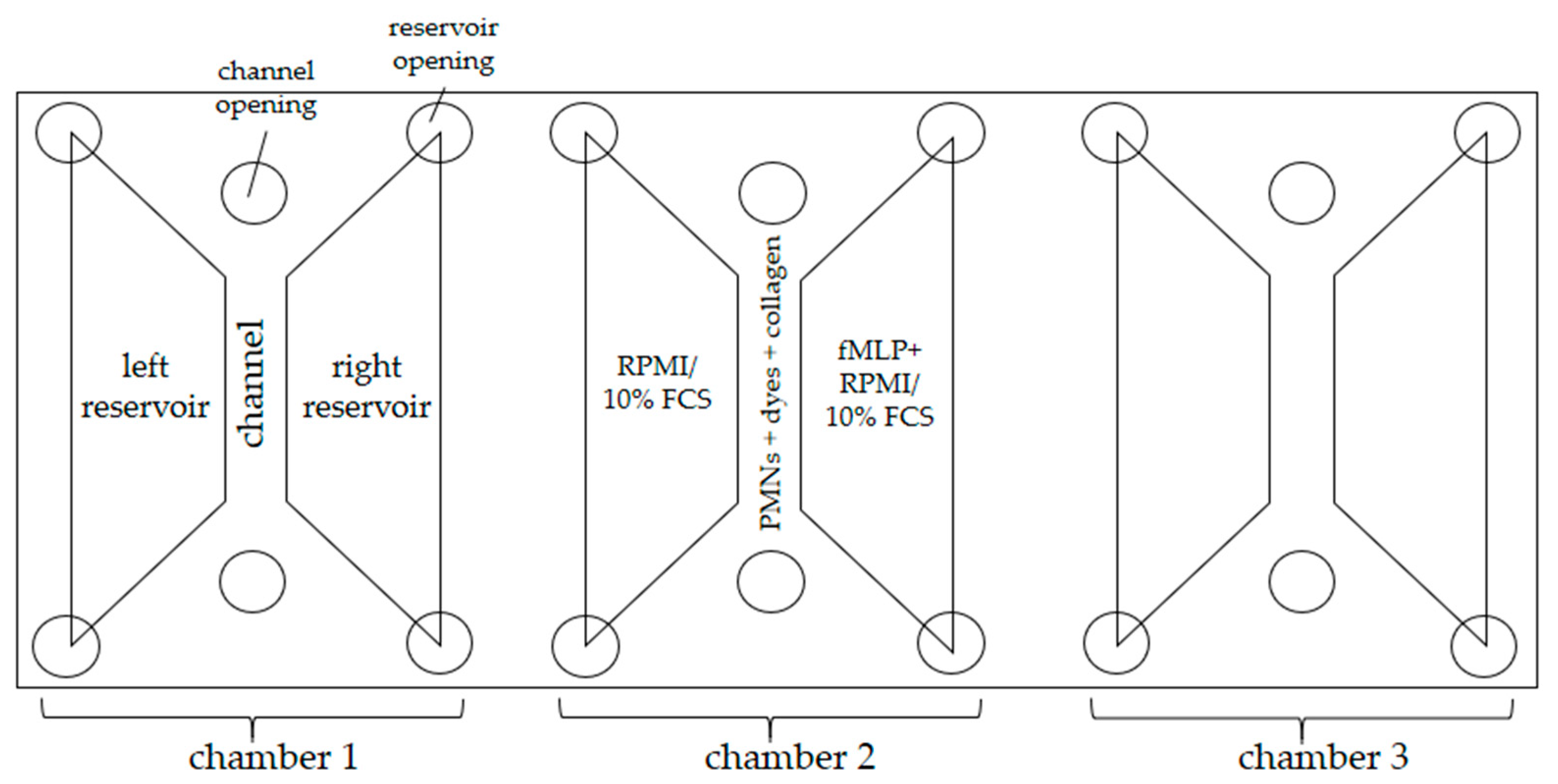
2.4.1. Experimental Setup
2.4.2. Chemotactic Migration
2.4.3. ROS Production, NET Formation, and MPO Release
2.5. Statistical Evaluation
3. Results
3.1. Demographical and Clinical Characteristics
3.2. Platelets, Leukocytes, CRP, GPT, GOT, and Creatinine
3.3. sFlt-1/PlGF Ratio
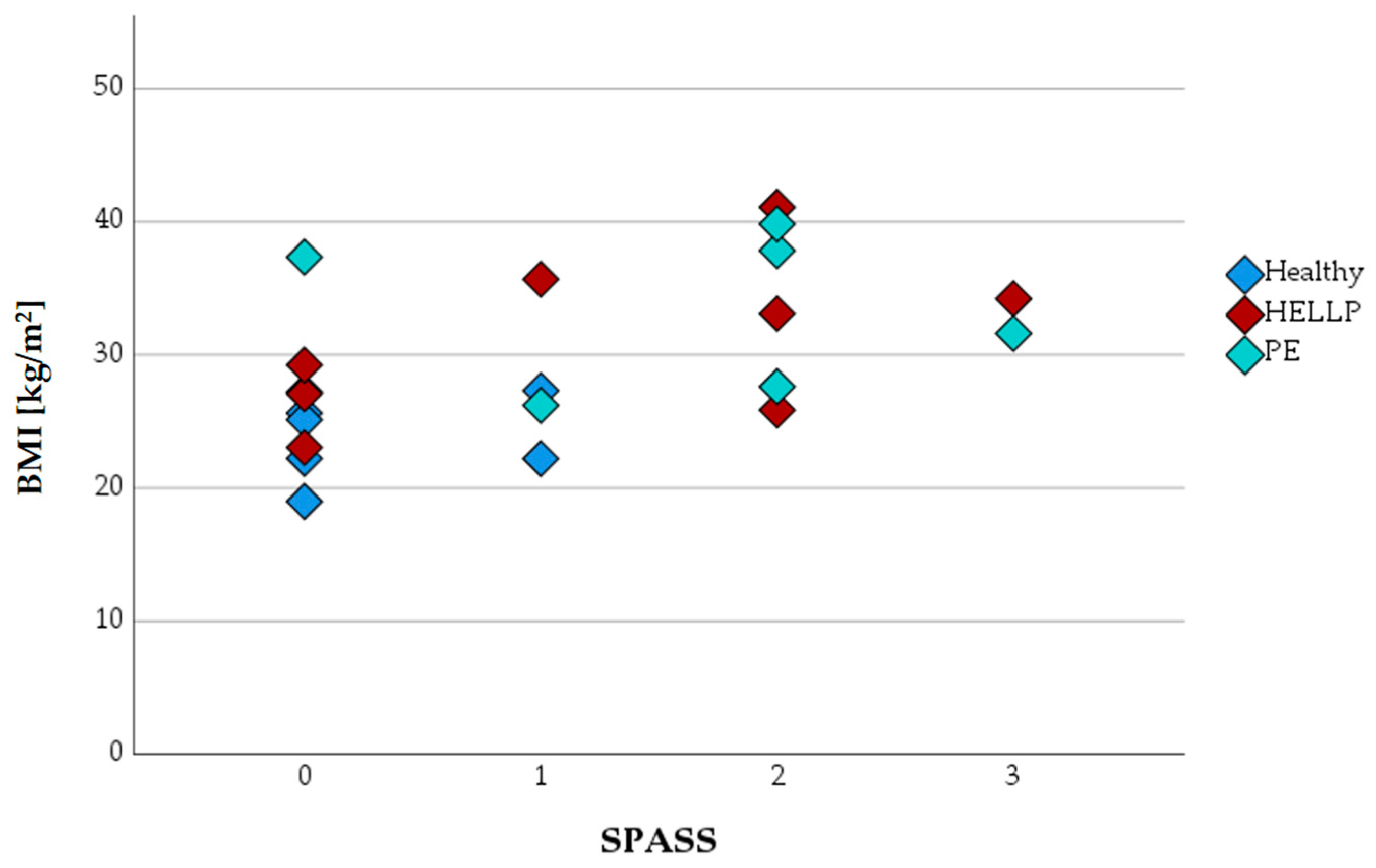
3.4. Expression of Surface Antigens CD11b, CD62L, CD66b, and SPASS Index
Introduction of Surface Protein Activation Sum Scale (SPASS)
3.5. Oxidative Burst
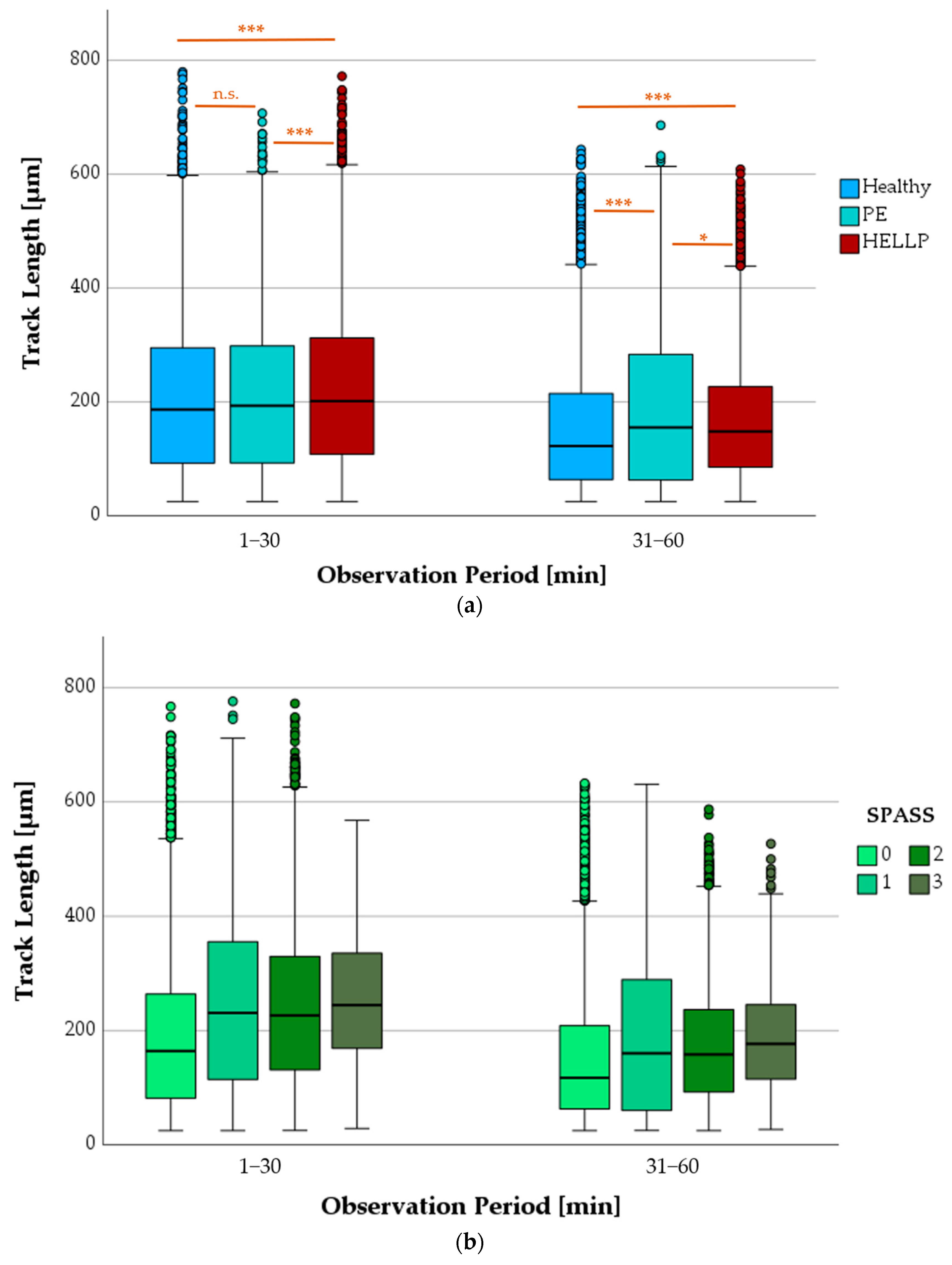
3.6. Chemotactic Migration
3.6.1. Progression of TL as a Function of Health Status or SPASS Values
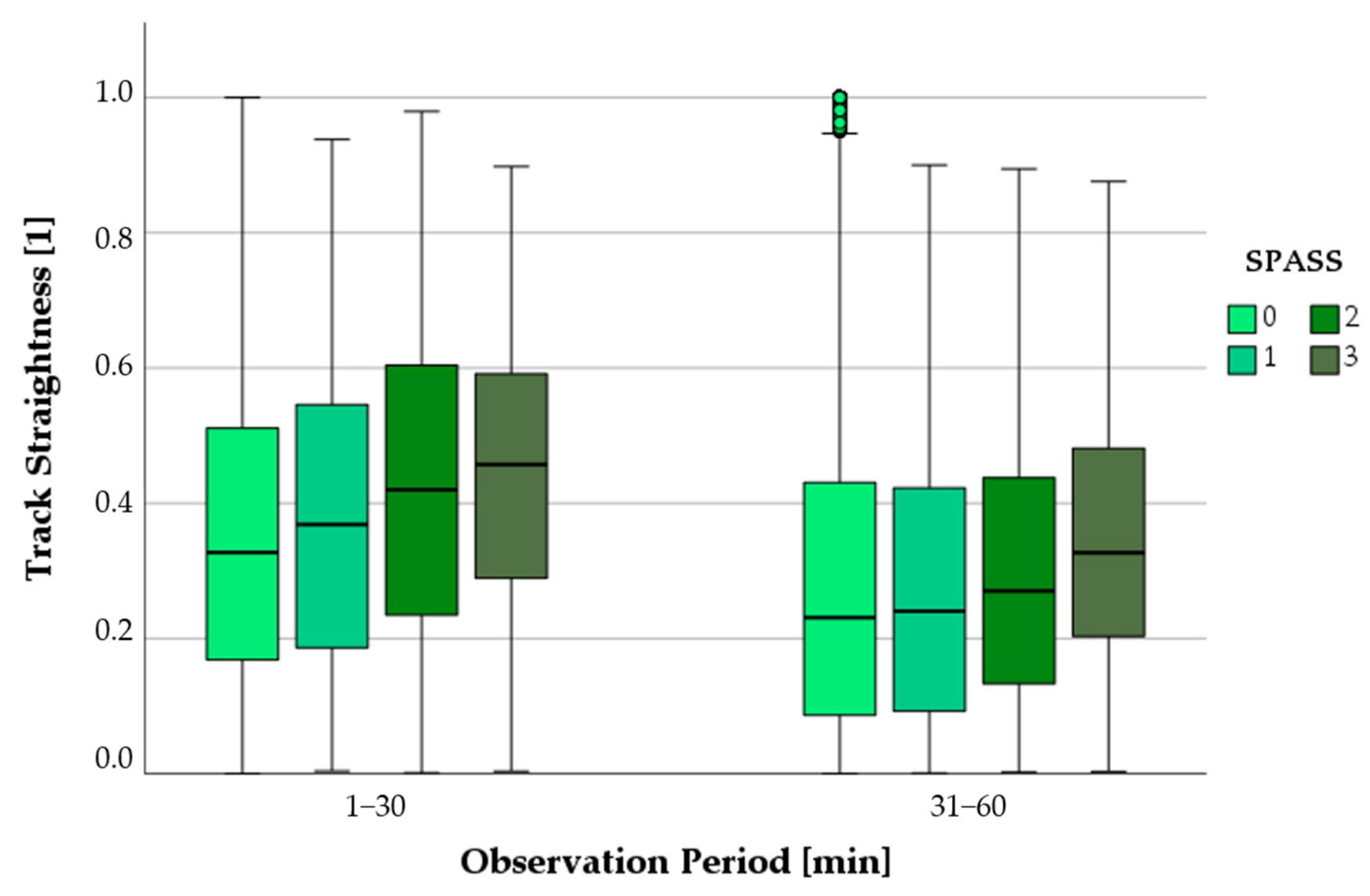
3.6.2. Progression of Track Straightness as a Function of SPASS Values
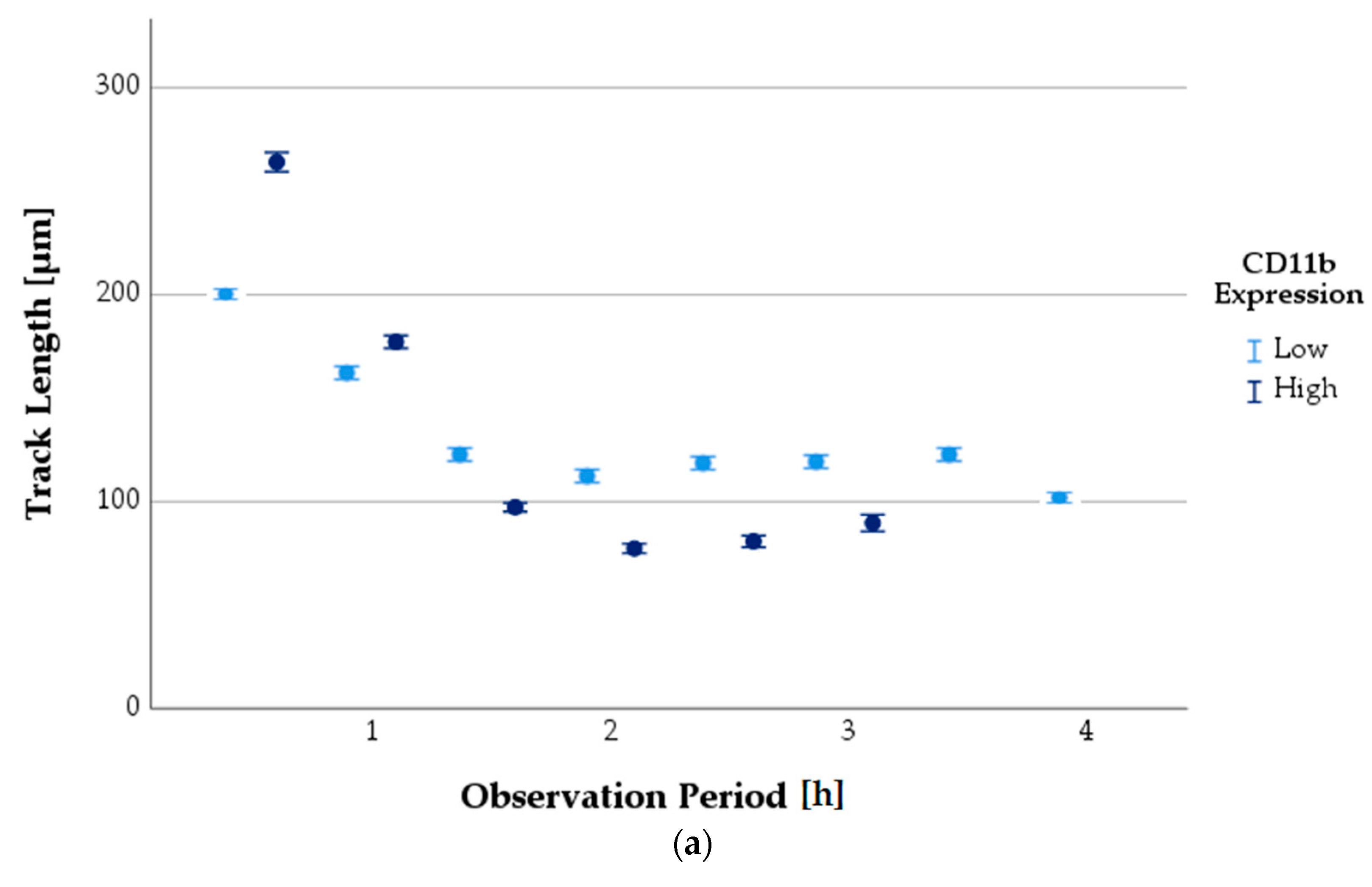
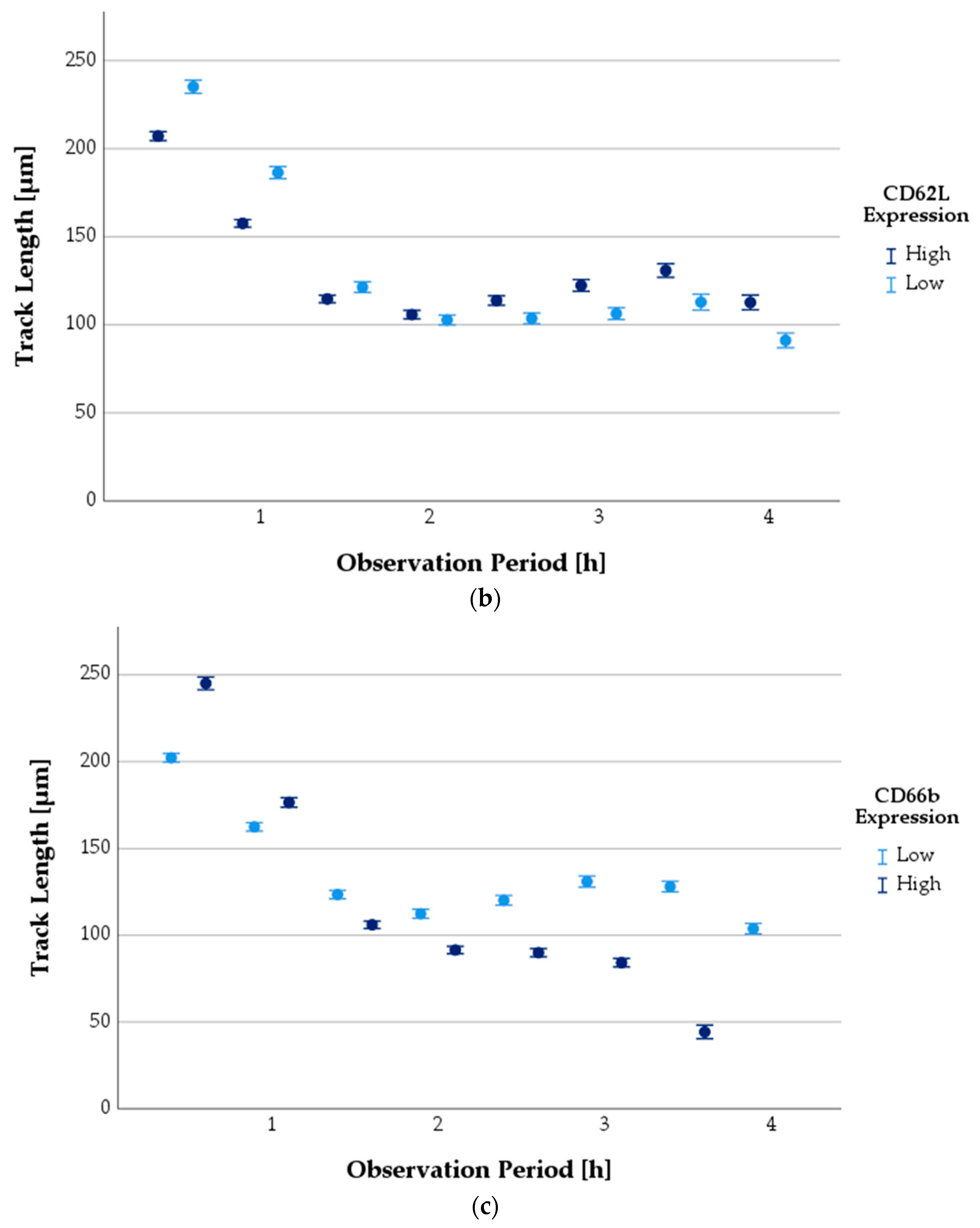
3.6.3. Progression of TL as a Function of Antigen Expression State
3.7. ROS Production
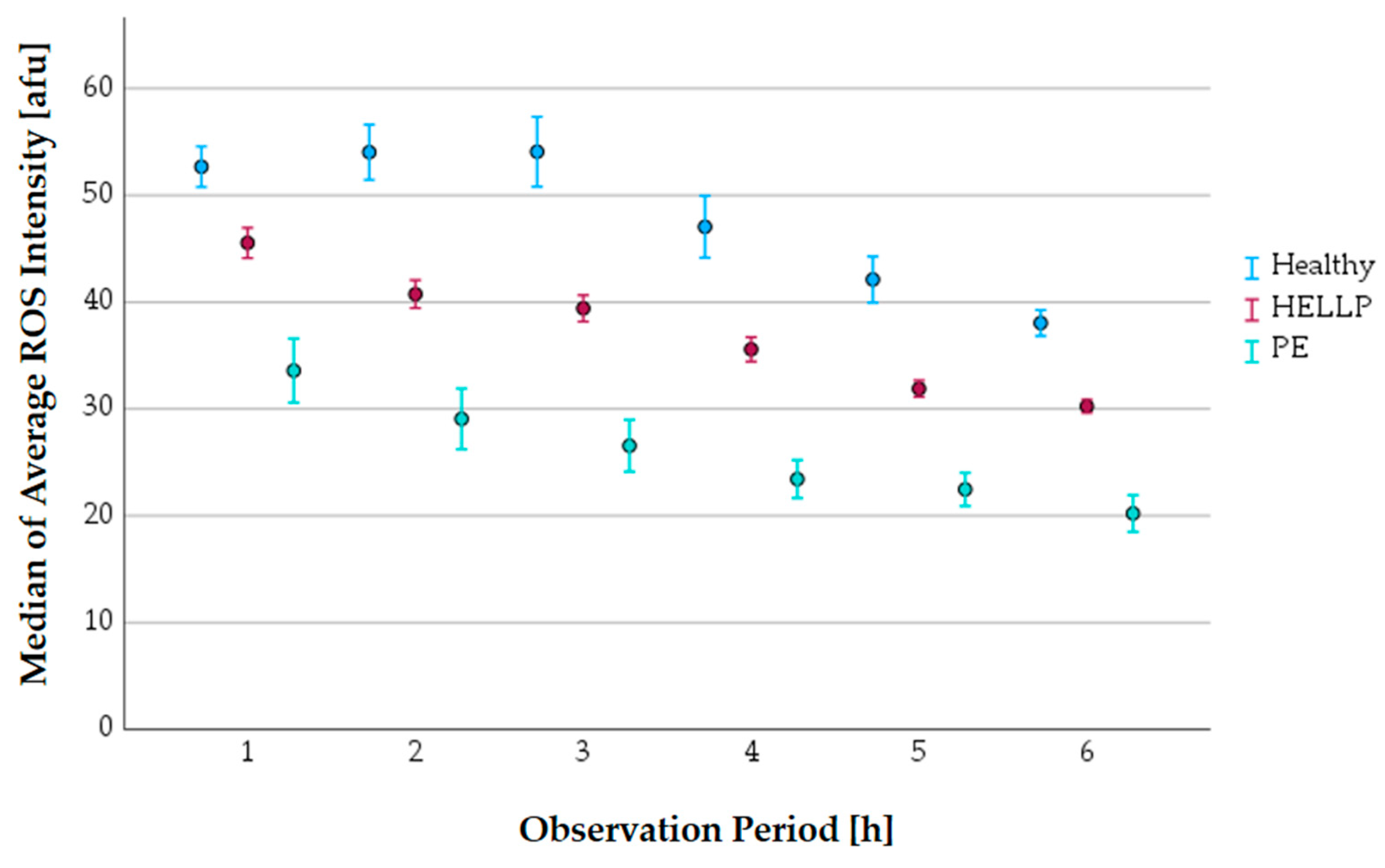
3.7.1. The Progress of the Average ROS Intensity along the Observation Period
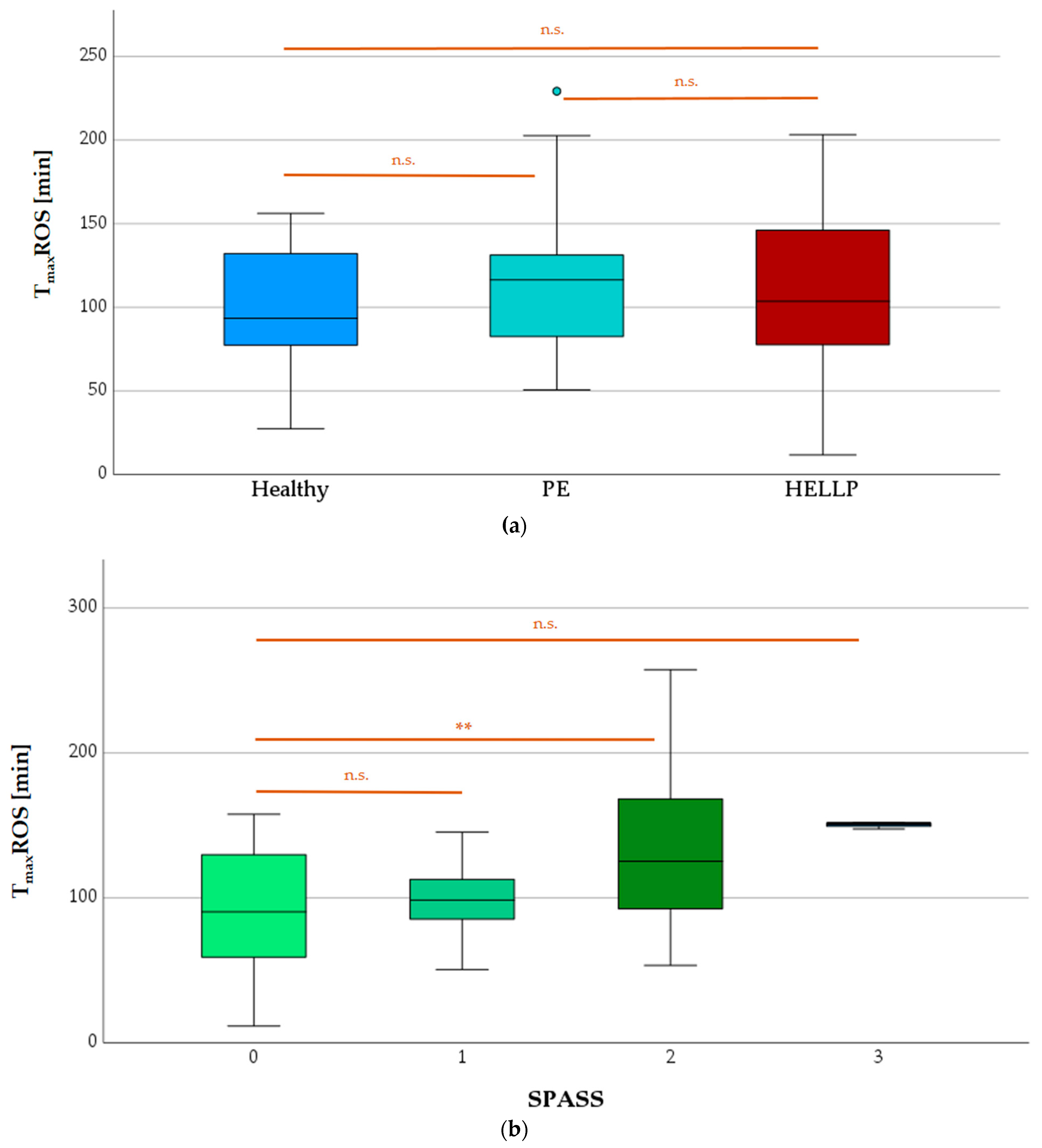
3.7.2. TmaxROS Depending on Health Status or SPASS Value
3.8. NETs and MPO Release
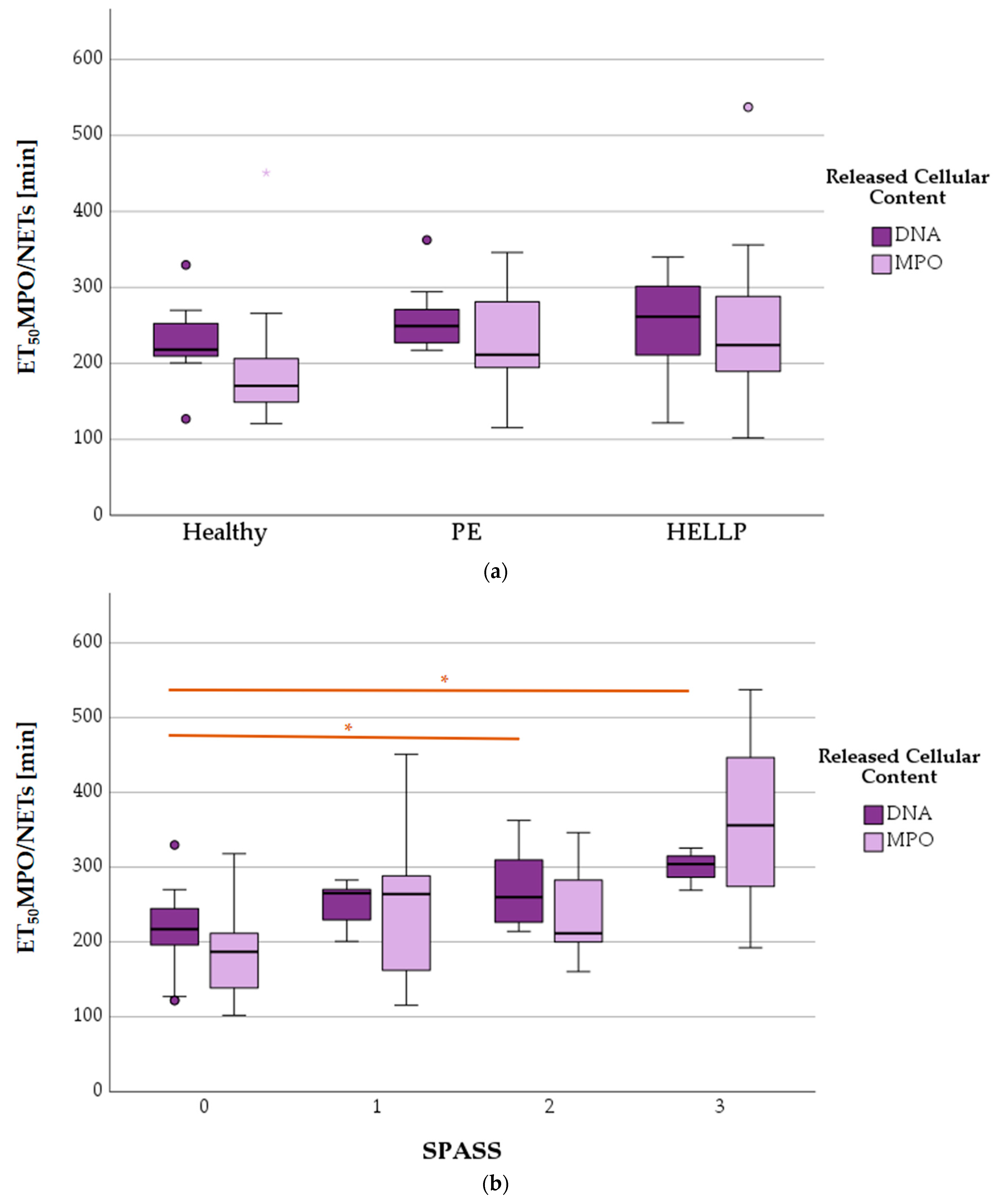
3.8.1. ET50NETosis in Terms of Health Status and SPASS Value
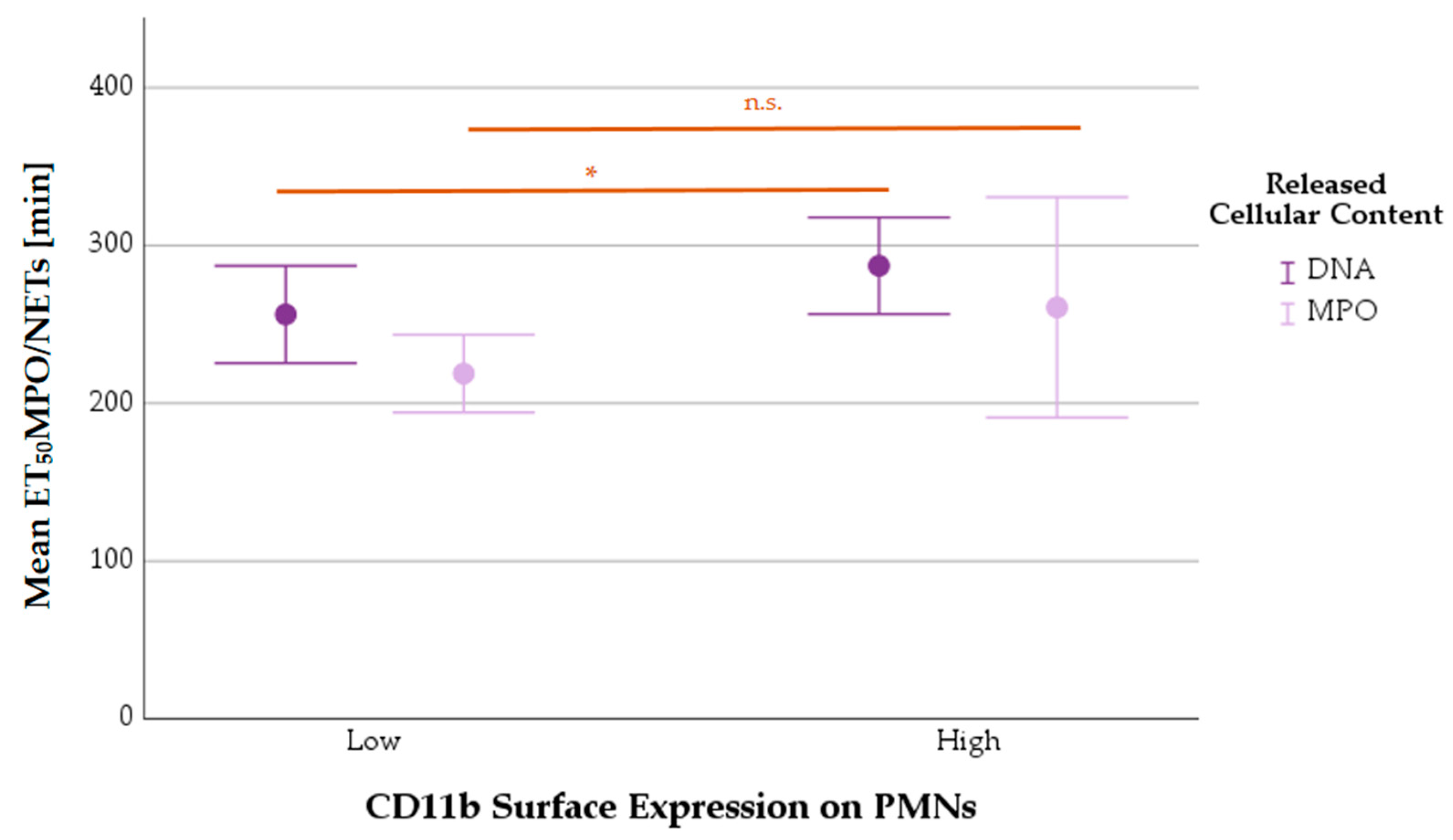
3.8.2. Mean ET50NETosis Depending on PMNs’ CD11b Surface Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petca, A.; Miron, B.C.; Pacu, I.; Dumitrașcu, M.C.; Mehedințu, C.; Șandru, F.; Petca, R.-C.; Rotar, I.C. HELLP Syndrome-Holistic Insight into Pathophysiology. Medicina 2022, 58, 326. [Google Scholar] [CrossRef] [PubMed]
- Várkonyi, T.; Nagy, B.; Füle, T.; Tarca, A.L.; Karászi, K.; Schönléber, J.; Hupuczi, P.; Mihalik, N.; Kovalszky, I.; Rigó, J.; et al. Microarray profiling reveals that placental transcriptomes of early-onset HELLP syndrome and preeclampsia are similar. Placenta 2011, 32, S21–S29. [Google Scholar] [CrossRef]
- Aloizos, S.; Seretis, C.; Liakos, N.; Aravosita, P.; Mystakelli, C.; Kanna, E.; Gourgiotis, S. HELLP syndrome: Understanding and management of a pregnancy-specific disease. J. Obstet. Gynaecol. 2013, 33, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Abildgaard, U.; Heimdal, K. Pathogenesis of the syndrome of hemolysis, elevated liver enzymes, and low platelet count (HELLP): A review. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 166, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Andraweera, P.H.; Dekker, G.A.; Roberts, C.T. The vascular endothelial growth factor family in adverse pregnancy outcomes. Hum. Reprod. Update 2012, 18, 436–457. [Google Scholar] [CrossRef]
- Whitley, G.S.J.; Cartwright, J.E. Trophoblast-mediated spiral artery remodelling: A role for apoptosis. J. Anat. 2009, 215, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; von Dadelszen, P.; Nadeau, J. CMV infection, TLR-2 and -4 expression, and cytokine profiles in early-onset preeclampsia with HELLP syndrome. Am. J. Reprod. Immunol. 2014, 71, 379–386. [Google Scholar] [CrossRef]
- Kraus, R.F.; Gruber, M.A. Neutrophils-From Bone Marrow to First-Line Defense of the Innate Immune System. Front. Immunol. 2021, 12, 767175. [Google Scholar] [CrossRef]
- De Gaetano, G.; Cerletti, C.; Evangelista, V. Recent advances in platelet-polymorphonuclear leukocyte interaction. Haemostasis 1999, 29, 41–49. [Google Scholar] [CrossRef]
- Barnard, M.R.; Linden, M.D.; Frelinger, A.L.; Li, Y.; Fox, M.L.; Furman, M.I.; Michelson, A.D. Effects of platelet binding on whole blood flow cytometry assays of monocyte and neutrophil procoagulant activity. J. Thromb. Haemost. 2005, 3, 2563–2570. [Google Scholar] [CrossRef]
- Aziz, K.A.; Cawley, J.C.; Kamiguti, A.S.; Zuzel, M. Degradation of platelet glycoprotein Ib by elastase released from primed neutrophils. Br. J. Haematol. 1995, 91, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Aziz, K.A.; Cawley, J.C.; Zuzel, M. Platelets prime PMN via released PF4: Mechanism of priming and synergy with GM-CSF. Br. J. Haematol. 1995, 91, 846–853. [Google Scholar] [CrossRef]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.-C.; Gougerot-Pocidalo, M.-A.; Dang, P.M.-C. Priming of the neutrophil respiratory burst: Role in host defense and inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Mincham, K.T.; Bruno, N.; Singanayagam, A.; Snelgrove, R.J. Our evolving view of neutrophils in defining the pathology of chronic lung disease. Immunology 2021, 164, 701–721. [Google Scholar] [CrossRef]
- Ma, Y.; Zhang, Y.; Zhu, L. Role of neutrophils in acute viral infection. Immun. Inflamm. Dis. 2021, 9, 1186–1196. [Google Scholar] [CrossRef]
- Kraus, R.F.; Gruber, M.A.; Kieninger, M. The influence of extracellular tissue on neutrophil function and its possible linkage to inflammatory diseases. Immun. Inflamm. Dis. 2021, 9, 1237–1251. [Google Scholar] [CrossRef] [PubMed]
- Lokki, A.I.; Teirilä, L.; Triebwasser, M.; Daly, E.; Bhattacharjee, A.; Uotila, L.; Llort Asens, M.; Kurki, M.I.; Perola, M.; Auro, K.; et al. Dysfunction of complement receptors CR3 (CD11b/18) and CR4 (CD11c/18) in pre-eclampsia: A genetic and functional study. BJOG Int. J. Obstet. Gynaecol. 2021, 128, 1282–1291. [Google Scholar] [CrossRef]
- Liu, D.; Li, Q.; Ding, H.; Zhao, G.; Wang, Z.; Cao, C.; Dai, Y.; Zheng, M.; Zhu, X.; Wu, Q.; et al. Placenta-derived IL-32β activates neutrophils to promote preeclampsia development. Cell Mol. Immunol. 2021, 18, 979–991. [Google Scholar] [CrossRef]
- Wang, Y.; Adair, C.D.; Weeks, J.W.; Lewis, D.F.; Alexander, J.S. Increased neutrophil-endothelial adhesion induced by placental factors is mediated by platelet-activating factor in preeclampsia. J. Soc. Gynecol. Investig. 1999, 6, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.S.; Khandelwal, M.; Zhao, J.; Mehmet, A.H.; Sammel, M.D.; Parry, S. Neutrophils are stimulated by syncytiotrophoblast microvillous membranes to generate superoxide radicals in women with preeclampsia. Am. J. Obstet. Gynecol. 2004, 190, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, H.; Yan, R.; Wang, C.; Wang, Y.; Zhang, C.; Liu, M.; Zhou, T.; Zhu, W.; Zhang, H.; et al. Increased Neutrophil Activation and Plasma DNA Levels in Patients with Pre-Eclampsia. Thromb. Haemost. 2018, 118, 2064–2073. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.; Motomura, K.; Galaz, J.; Gershater, M.; Lee, E.D.; Romero, R.; Gomez-Lopez, N. Cellular immune responses in the pathophysiology of preeclampsia. J. Leukoc. Biol. 2022, 111, 237–260. [Google Scholar] [CrossRef]
- Mazzitelli, I.; Bleichmar, L.; Ludueña, M.G.; Pisarevsky, A.; Labato, M.; Chiaradia, V.; Finocchieto, P.; Paulin, F.; Hormanstorfer, M.; Baretto, M.C.; et al. Immunoglobulin G Immune Complexes May Contribute to Neutrophil Activation in the Course of Severe Coronavirus Disease 2019. J. Infect. Dis. 2021, 224, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Bongers, S.H.; Chen, N.; van Grinsven, E.; van Staveren, S.; Hassani, M.; Spijkerman, R.; Hesselink, L.; Lo Tam Loi, A.T.; van Aalst, C.; Leijte, G.P.; et al. Kinetics of Neutrophil Subsets in Acute, Subacute, and Chronic Inflammation. Front. Immunol. 2021, 12, 674079. [Google Scholar] [CrossRef]
- Pai, D.; Gruber, M.; Pfaehler, S.-M.; Bredthauer, A.; Lehle, K.; Trabold, B. Polymorphonuclear Cell Chemotaxis and Suicidal NETosis: Simultaneous Observation Using fMLP, PMA, H7, and Live Cell Imaging. J. Immunol. Res. 2020, 2020, 1415947. [Google Scholar] [CrossRef]
- Doblinger, N.; Bredthauer, A.; Mohrez, M.; Hähnel, V.; Graf, B.; Gruber, M.; Ahrens, N. Impact of hydroxyethyl starch and modified fluid gelatin on granulocyte phenotype and function. Transfusion 2019, 59, 2121–2130. [Google Scholar] [CrossRef] [PubMed]
- Bredthauer, A.; Kopfmueller, M.; Gruber, M.; Pfaehler, S.-M.; Lehle, K.; Petermichl, W.; Seyfried, T.; Bitzinger, D.; Redel, A. Therapeutic Anticoagulation with Argatroban and Heparins Reduces Granulocyte Migration: Possible Impact on ECLS-Therapy? Cardiovasc. Ther. 2020, 2020, 9783630. [Google Scholar] [CrossRef]
- Zeisler, H.; Llurba, E.; Chantraine, F.; Vatish, M.; Staff, A.C.; Sennström, M.; Olovsson, M.; Brennecke, S.P.; Stepan, H.; Allegranza, D.; et al. Predictive Value of the sFlt-1:PlGF Ratio in Women with Suspected Preeclampsia. N. Engl. J. Med. 2016, 374, 13–22. [Google Scholar] [CrossRef]
- Verlohren, S.; Brennecke, S.P.; Galindo, A.; Karumanchi, S.A.; Mirkovic, L.B.; Schlembach, D.; Stepan, H.; Vatish, M.; Zeisler, H.; Rana, S. Clinical interpretation and implementation of the sFlt-1/PlGF ratio in the prediction, diagnosis and management of preeclampsia. Pregnancy Hypertens. 2022, 27, 42–50. [Google Scholar] [CrossRef]
- Jeon, H.R.; Da Jeong, H.; Lee, J.Y.; Woo, E.Y.; Shin, G.T.; Kim, S.-Y. sFlt-1/PlGF ratio as a predictive and prognostic marker for preeclampsia. J. Obstet. Gynaecol. Res. 2021, 47, 2318–2323. [Google Scholar] [CrossRef]
- Gervasi, M.T.; Chaiworapongsa, T.; Pacora, P.; Naccasha, N.; Yoon, B.H.; Maymon, E.; Romero, R. Phenotypic and metabolic characteristics of monocytes and granulocytes in preeclampsia. Am. J. Obstet. Gynecol. 2001, 185, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Sacks, G.P.; Studena, K.; Sargent, K.; Redman, C.W. Normal pregnancy and preeclampsia both produce inflammatory changes in peripheral blood leukocytes akin to those of sepsis. Am. J. Obstet. Gynecol. 1998, 179, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Sabatier, F.; Bretelle, F.; D’ercole, C.; Boubli, L.; Sampol, J.; Dignat-George, F. Neutrophil activation in preeclampsia and isolated intrauterine growth restriction. Am. J. Obstet. Gynecol. 2000, 183, 1558–1563. [Google Scholar] [CrossRef] [PubMed]
- Luppi, P.; Tse, H.; Lain, K.Y.; Markovic, N.; Piganelli, J.D.; DeLoia, J.A. Preeclampsia activates circulating immune cells with engagement of the NF-kappaB pathway. Am. J. Reprod. Immunol. 2006, 56, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Zhang, X.; Wang, C.; Wang, H.; Wang, T.; Qian, Z. Hydrogen peroxide promotes the activation of preeclampsia peripheral T cells. Innate Immun. 2018, 24, 203–209. [Google Scholar] [CrossRef]
- Husse, S.; Gottschlich, A.; Schrey, S.; Stepan, H.; Hoffmann, J. Predictive value of the sFlt1/PlGF ratio for the diagnosis of preeclampsia in high-risk patients. Z. Geburtshilfe Neonatol. 2014, 218, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Shah, T.J.; Walsh, S.W. Activation of NF-kappaB and expression of COX-2 in association with neutrophil infiltration in systemic vascular tissue of women with preeclampsia. Am. J. Obstet. Gynecol. 2007, 196, 48.e1–48.e8. [Google Scholar] [CrossRef]
- Cadden, K.A.; Walsh, S.W. Neutrophils, but not lymphocytes or monocytes, infiltrate maternal systemic vasculature in women with preeclampsia. Hypertens. Pregnancy 2008, 27, 396–405. [Google Scholar] [CrossRef]
- Von Dadelszen, P.; Watson, R.W.; Noorwali, F.; Marshall, J.C.; Parodo, J.; Farine, D.; Lye, S.J.; Ritchie, J.W.; Rotstein, O.D. Maternal neutrophil apoptosis in normal pregnancy, preeclampsia, and normotensive intrauterine growth restriction. Am. J. Obstet. Gynecol. 1999, 181, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Walsh, S.W.; Nugent, W.H.; Al Dulaimi, M.; Washington, S.L.; Dacha, P.; Strauss, J.F. Proteases Activate Pregnancy Neutrophils by a Protease-Activated Receptor 1 Pathway: Epigenetic Implications for Preeclampsia. Reprod. Sci. 2020, 27, 2115–2127. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Left Reservoir (65 µL) | Channel (6 µL) | Right Reservoir (65 µL) |
|---|---|---|
| RPMI/10% FCS | PMNs in medium (15.7%) DAPI [0.5 µg/mL] DHR 123 [1 µM] | fMLP [10 nM] RPMI/10% FCS |
| Anti-MPO antibody [0.5 µg/mL] | ||
| Collagen Type 1/1.67% FCS [1.5 mg/mL] |
| Parameter (Abbreviation) | Unit | Description |
|---|---|---|
| Track Length (TL) | [µm] | Total length of the migration route of each individual tracked cell |
| Track Straightness (TS) | [1] | Fraction of Euclidean track length and total track length showing the cell’s tendency to migrate directly; higher factors refer to straighter lines |
| Healthy | PE | p-Value | |
|---|---|---|---|
| Age (years) | 33 (28;34) | 32 (30;32) | n.s. |
| Time of pregnancy (weeks) | 33 (31;38) | 35 (29;36) | n.s. |
| Body height (cm) | 167 (158;171) | 163 (161;170) | n.s. |
| Body weight (kg) | 65 (59;71) | 84 (73;107) | 0.021 |
| BMI (kg/m2) | 22 (22;27) | 32 (27;39) | 0.016 |
| Healthy | HELLP | p-Value | |
| Age (years) | 33 (28;34) | 31 (29;38) | n.s. |
| Time of pregnancy (weeks) | 33 (31;38) | 33 (28;36) | n.s. |
| Body height (cm) | 167 (158;171) | 166 (163;169) | n.s. |
| Body weight (kg) | 65 (59;71) | 85 (72;100) | 0.028 |
| BMI (kg/m2) | 22 (22;27) | 31 (26;35) | 0.019 |
| Healthy | PE | HELLP | |
|---|---|---|---|
| Platelets (103/µL) | 232 ± 49 p < 0.001 vs. HELLP | 182 ± 32 p < 0.001 vs. HELLP | 80 ± 28 |
| Leukocytes (103/µL) | 10.8 ± 1.9 p = 0.045 vs. PE | 8.8 ± 1.1 p = 0.010 vs. PE | 13.6 ± 5.5 |
| CRP (mg/L) | 4.2 ± 4.5 p = 0.035 vs. HELLP | 3.4 ± 4.5 p = 0.031 vs. HELLP | 32.7 ± 21.7 |
| GPT (U/L) | 16.7 ± 9.9 p = 0.003 vs. HELLP | 19.8 ± 6.4 p = 0.005 vs. HELLP | 191.3 ± 133 |
| GOT (U/L) | 24.3 ± 8.6 p < 0.001 vs. HELLP | 28.7 ± 6.6 p = 0.001 vs. HELLP | 232 ± 229 |
| Creatinine (mg/dL) | 0.70 ± 0.08 p = 0.009 vs. HELLP | 0.70 ± 0.08 p = 0.012 vs. HELLP | 0.95 ± 0.16 |
| PE | HELLP | |
|---|---|---|
| sFlt-1 [ng/mL] | 12.2 ± 4.5 | 17.3 ± 5.4 |
| PlGF [pg/mL] | 40 ± 4 | 68 ± 62 |
| sFlt-1/PlGF ratio | 308 ± 98 | 442 ± 385 |
| Healthy | PE | HELLP | |
|---|---|---|---|
| CD11b | 372 ± 140 | 398 ± 174 p = 1.000 | 586 ± 358 p = 0.217 |
| CD62L | 137 ± 31.5 | 94.9 ± 10.6 p = 0.024 | 99.0 ± 28.8 p = 0.116 |
| CD66b | 176 ± 47.5 | 220 ± 45.4 p = 0.472 | 253 ± 84.1 p = 0.174 |
| SPASS | Healthy (n) | PE (n) | HELLP (n) | Total (n) |
|---|---|---|---|---|
| 0 | 5 | 1 | 3 | 9 |
| 1 | 2 | 1 | 1 | 4 |
| 2 | 0 | 3 | 3 | 6 |
| 3 | 0 | 1 | 1 | 2 |
| Healthy | PE | HELLP | ||
|---|---|---|---|---|
| fMLP + TNFα | 23.4 ± 20.2 | 25.8 ± 18.6 | 34.7 ± 4.9 | |
| PMA | 569 ± 182 | 553 ± 152 | 592 ± 129 | |
| SPASS | 0 | 1 | 2 | 3 |
| fMLP + TNFα | 30.1 ± 15.5 | 26.2 ± 22.2 | 20.2 ± 15.2 | 40.0 ± 1.4 |
| PMA | 620 ± 164 | 492 ± 93.2 | 570 ± 171 | 555 ± 123 |
| Time [h] | Healthy | PE | HELLP |
|---|---|---|---|
| 1 | 52.7 | 33.6 *** | 45.5 *** |
| 2 | 54.0 | 29.0 *** | 40.7 *** |
| 3 | 54.1 | 26.6 *** | 39.4 *** |
| 4 | 47.1 | 23.4 *** | 35.6 *** |
| 5 | 42.1 | 22.5 *** | 31.9 *** |
| 6 | 38.0 | 20.2 *** | 30.3 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rimboeck, J.; Gruber, M.; Weigl, M.; Huber, P.; Lunz, D.; Petermichl, W. Obesity Correlates with Chronic Inflammation of the Innate Immune System in Preeclampsia and HELLP Syndrome during Pregnancy. Biomedicines 2023, 11, 2851. https://doi.org/10.3390/biomedicines11102851
Rimboeck J, Gruber M, Weigl M, Huber P, Lunz D, Petermichl W. Obesity Correlates with Chronic Inflammation of the Innate Immune System in Preeclampsia and HELLP Syndrome during Pregnancy. Biomedicines. 2023; 11(10):2851. https://doi.org/10.3390/biomedicines11102851
Chicago/Turabian StyleRimboeck, Julia, Michael Gruber, Marco Weigl, Pia Huber, Dirk Lunz, and Walter Petermichl. 2023. "Obesity Correlates with Chronic Inflammation of the Innate Immune System in Preeclampsia and HELLP Syndrome during Pregnancy" Biomedicines 11, no. 10: 2851. https://doi.org/10.3390/biomedicines11102851
APA StyleRimboeck, J., Gruber, M., Weigl, M., Huber, P., Lunz, D., & Petermichl, W. (2023). Obesity Correlates with Chronic Inflammation of the Innate Immune System in Preeclampsia and HELLP Syndrome during Pregnancy. Biomedicines, 11(10), 2851. https://doi.org/10.3390/biomedicines11102851