
Genetic Polymorphisms and Diversity in Nonalcoholic Fatty Liver Disease (NAFLD): A Mini Review



Abstract
1. Introduction
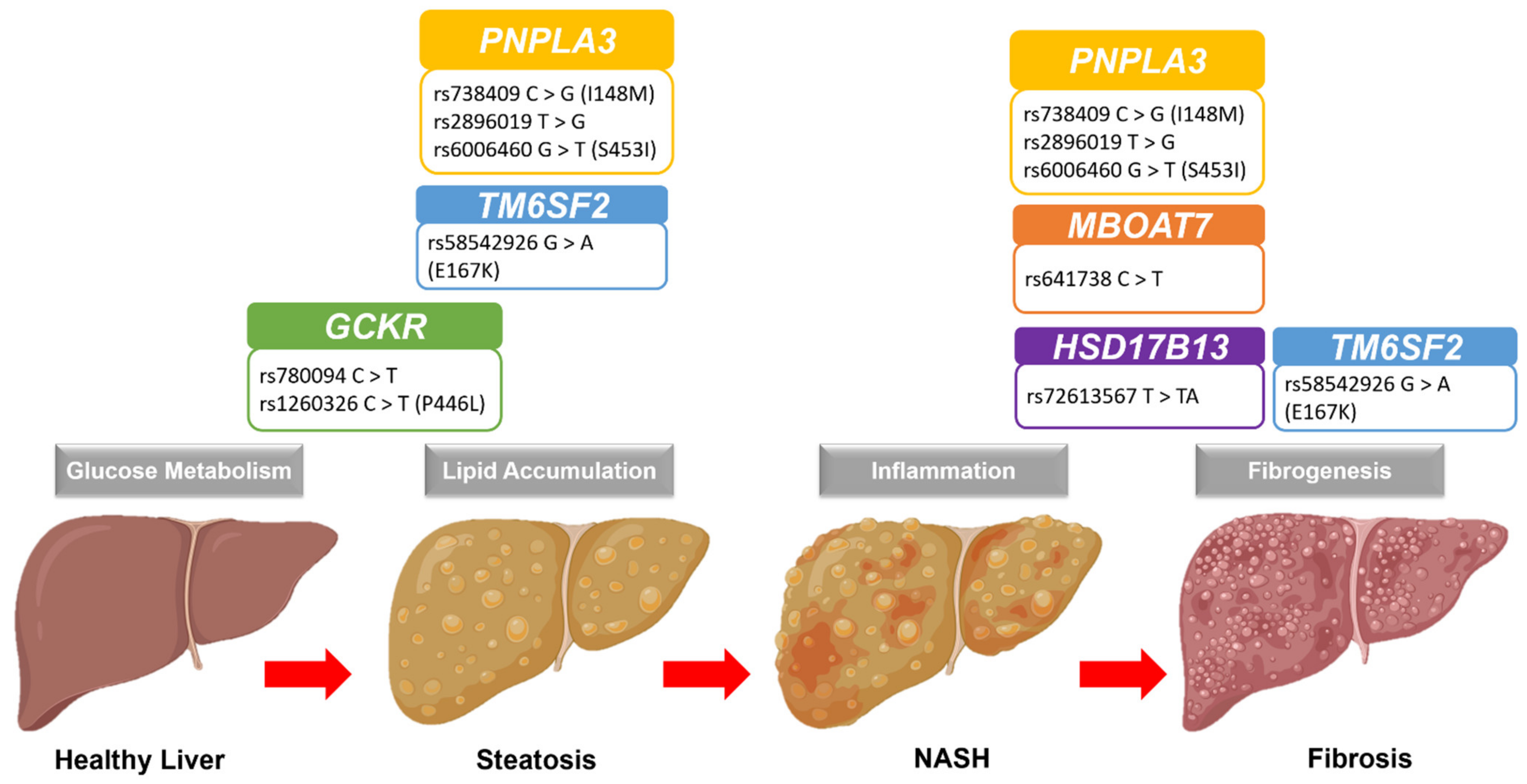
2. NAFLD Pathogenesis and Progression
3. Genetic Polymorphisms of NAFLD
3.1. PNPLA3 Loci
3.2. TM6SF2 Loci
3.3. GCKR Loci
3.4. MBOAT7 Loci
3.5. HSD17B13 Loci
3.6. Other Genetic Loci
3.6.1. Loci in Energy Metabolism

{kind=link}
| Gene | SNP | Population Study | SNP Effects | Reference(s) |
|---|---|---|---|---|
| PPP1R3B | rs4240624 G > A/C | Amish, Family Heart Study, and Framingham Heart Study | Associated with NAFLD risk | [39] |
| Caucasian, African, and Mexican Americans | Associated with steatosis | [92] | ||
| rs61756425 G > T | Italian | Associated with NAFLD severity | [49] | |
| PPARGC1A | rs8192678 G > A (G482S) | Iranian, Chinese | Associated with NAFLD and NASH risk | [152,153,154] |
| rs2290602 G > T | Japanese | Associated with NAFLD and NASH risk | [156] | |
| SAMM50 | rs3761472 A > G | Chinese, Korean, and Japanese | Associated with NAFLD risk | [48,157,158,159] |
| Indian | Associated with NASH | [72] | ||
| rs738491 C > T | Chinese, Japanese | Associated with NAFLD risk | [48,157] | |
| rs2143571 G > A | Chinese, Korean, Japanese | Associated with NAFLD risk | [48,157,159] | |
| APOC3 | rs2070666 T > A | Chinese | Associated with hepatic lipid content | [160] |
| rs2854116 (T-455C) | European | No association with NAFLD risk | [161] | |
| Indian | Associated with NAFLD and lipid content | [162,163] | ||
| rs2854117 (C-482T) | European | No association with NAFLD risk | [161] | |
| Indian | Associated with NAFLD and lipid content | [162,163] | ||
| ATGR1 | rs2276736 A > G/T rs3772630 T > A/C rs3772627 A > G | Malaysians (Malays, Chinese, and Indians) | Associated with NAFLD risk in the presence of PNPLA3 I148M variant | [164] |
| Japanese | Associated with NAFLD risk and fibrosis | [165] | ||
| GATAD2A | rs4808199 G > A | Japanese | Associated with NAFLD | [74] |
| IL27 | rs4788084 C > T | Indian | Associated with hepatic lipid content | [72] |
| LPIN1 | rs13412852 C > T | Italian | Tended to associate with lower NASH | [166] |
| LYPLAL1 | rs12137855 C > T | Amish, Family Heart Study, Framingham Heart Study, Finnish | Associated with NAFLD risk and steatosis | [39,101] |
| PEMT | rs7946 C > T (V175M) | Chinese, Caucasian | Associated with NAFLD risk | [167,168,169] |
| Indian | Associated with NAFLD risk in lean individuals | [170] | ||
| Korean | No association with NAFLD risk | [171] | ||
| MTTP | -493C/G | Japanese | Associated with NASH | [172] |
| Italian, Brazilian | No association with NAFLD risk | [173,174,175] | ||
| PARVB | rs5764455 A > C/G | Japanese | Associated with NASH | [48] |
| rs2073080 C > A/T | Indian, Finnish | Associated with steatosis | [66,101] | |
| SOD | rs4880 A > G (C47T) | Italian and European | Associated with hepatic steatosis and fibrosis | [176] |
| Japanese | Associated with NASH | [172] | ||
| REPIN1 | 12 bp deletion | Germany | Associated with lower NAFLD severity | [177] |
| UCP3 | rs1800849 C > T (-55CT) | Spaniards | Associated with NASH | [178] |
3.6.2. Loci in Mitochondrial Regulation
3.6.3. Loci in Cholesterol Polymorphism
3.6.4. Copy Number of Variants (CNV)
3.7. Polygenic Risk Scores
4. Genetic Diversity in NAFLD Risk
4.1. Genetic Diversity of PNPLA3 I148M
4.2. Genetic Diversity of Other NAFLD Variants
4.3. Discrepancies between the Same Populations or Ethnicities
4.4. Challenges in NAFLD Genetic Diversity Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Cotter, T.G.; Rinella, M. Nonalcoholic fatty liver disease 2020: The state of the disease. Gastroenterology 2020, 158, 1851–1864. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, J.; Nazratun Nafizah, A.H.; Zariyantey, A.H.; Budin, S.B. Mechanisms of diabetes-induced liver damage: The role of oxidative stress and inflammation. Sultan Qaboos Univ. Med. J. 2016, 16, e132–e141. [Google Scholar] [CrossRef] [PubMed]
- Henry, L.; Paik, J.; Younossi, Z.M. Review article: The epidemiologic burden of non-alcoholic fatty liver disease across the world. Aliment. Pharmacol. Ther. 2022, 56, 942–956. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Yilmaz, Y.; Yu, M.L.; Wai-Sun Wong, V.; Fernandez, M.C.; Isakov, V.A.; Duseja, A.K.; Mendez-Sanchez, N.; Eguchi, Y.; Bugianesi, E.; et al. Clinical and patient-reported outcomes from patients with nonalcoholic fatty liver disease across the world: Data from the global non-alcoholic steatohepatitis (nash)/non-alcoholic fatty liver disease (nafld) registry. Clin. Gastroenterol. Hepatol. 2022, 20, 2296–2306.e6. [Google Scholar] [CrossRef] [PubMed]
- Han, M.A.T.; Yu, Q.; Tafesh, Z.; Pyrsopoulos, N. Diversity in nafld: A review of manifestations of nonalcoholic fatty liver disease in different ethnicities globally. J. Clin. Transl. Hepatol. 2021, 9, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Riazi, K.; Swain, M.G.; Congly, S.E.; Kaplan, G.G.; Shaheen, A.A. Race and ethnicity in non-alcoholic fatty liver disease (nafld): A narrative review. Nutrients 2022, 14, 4556. [Google Scholar] [CrossRef]
- Yip, T.C.-F.; Vilar-Gomez, E.; Petta, S.; Yilmaz, Y.; Wong, G.L.-H.; Adams, L.A.; De Lédinghen, V.; Sookoian, S.; Wong, V.W.-S. Geographical similarity and differences in the burden and genetic predisposition of nafld. Hepatology 2022. online ahead of print. [Google Scholar] [CrossRef]
- Jonas, W.; Schürmann, A. Genetic and epigenetic factors determining nafld risk. Mol. Metab. 2021, 50, 101111. [Google Scholar] [CrossRef]
- Del Campo, J.A.; Gallego-Durán, R.; Gallego, P.; Grande, L. Genetic and epigenetic regulation in nonalcoholic fatty liver disease (nafld). Int. J. Mol. Sci. 2018, 19, 911. [Google Scholar] [CrossRef]
- Zhu, X.; Xia, M.; Gao, X. Update on genetics and epigenetics in metabolic associated fatty liver disease. Ther. Adv. Endocrinol. Metab. 2022, 13, 20420188221132138. [Google Scholar] [CrossRef]
- Wajsbrot, N.B.; Leite, N.C.; Salles, G.F.; Villela-Nogueira, C.A. Non-alcoholic fatty liver disease and the impact of genetic, epigenetic and environmental factors in the offspring. World J. Gastroenterol. 2022, 28, 2890–2899. [Google Scholar] [CrossRef]
- Sulaiman, S.A.; Muhsin, N.I.A.; Jamal, R. Regulatory non-coding rnas network in non-alcoholic fatty liver disease. Front. Physiol. 2019, 10, 279. [Google Scholar] [CrossRef]
- Dorairaj, V.; Sulaiman, S.A.; Abu, N.; Abdul Murad, N.A. Extracellular vesicles in the development of the non-alcoholic fatty liver disease: An update. Biomolecules 2020, 10, 1494. [Google Scholar] [CrossRef]
- Pafili, K.; Roden, M. Nonalcoholic fatty liver disease (nafld) from pathogenesis to treatment concepts in humans. Mol. Metab. 2021, 50, 101122. [Google Scholar] [CrossRef]
- Dorairaj, V.; Sulaiman, S.A.; Abu, N.; Abdul Murad, N.A. Nonalcoholic fatty liver disease (nafld): Pathogenesis and noninvasive diagnosis. Biomedicines 2021, 10, 15. [Google Scholar] [CrossRef]
- Zhao, J.; Wu, Y.; Rong, X.; Zheng, C.; Guo, J. Anti-lipolysis induced by insulin in diverse pathophysiologic conditions of adipose tissue. Diabetes Metab. Syndr. Obes. 2020, 13, 1575–1585. [Google Scholar] [CrossRef]
- Bence, K.K.; Birnbaum, M.J. Metabolic drivers of non-alcoholic fatty liver disease. Mol. Metab. 2021, 50, 101143. [Google Scholar] [CrossRef]
- Pais, R.; Maurel, T. Natural history of nafld. J. Clin. Med. 2021, 10, 1161. [Google Scholar] [CrossRef]
- Koliaki, C.; Szendroedi, J.; Kaul, K.; Jelenik, T.; Nowotny, P.; Jankowiak, F.; Herder, C.; Carstensen, M.; Krausch, M.; Knoefel, W.T.; et al. Adaptation of hepatic mitochondrial function in humans with non-alcoholic fatty liver is lost in steatohepatitis. Cell Metab. 2015, 21, 739–746. [Google Scholar] [CrossRef]
- Ma, X.; Cheng, F.; Yuan, K.; Jiang, K.; Zhu, T. Lipid storage droplet protein 5 reduces sodium palmitate-induced lipotoxicity in human normal liver cells by regulating lipid metabolism-related factors. Mol. Med. Rep. 2019, 20, 879–886. [Google Scholar] [CrossRef]
- Li, J.L.; Wang, Q.Y.; Luan, H.Y.; Kang, Z.C.; Wang, C.B. Effects of l-carnitine against oxidative stress in human hepatocytes: Involvement of peroxisome proliferator-activated receptor alpha. J. Biomed. Sci. 2012, 19, 32. [Google Scholar] [CrossRef] [PubMed]
- Engin, A. Non-alcoholic fatty liver disease. Adv. Exp. Med. Biol. 2017, 960, 443–467. [Google Scholar] [PubMed]
- Zhu, B.; Chan, S.L.; Li, J.; Li, K.; Wu, H.; Cui, K.; Chen, H. Non-alcoholic steatohepatitis pathogenesis, diagnosis, and treatment. Front. Cardiovasc. Med. 2021, 8, 1059. [Google Scholar] [CrossRef] [PubMed]
- Locati, M.; Curtale, G.; Mantovani, A. Diversity, mechanisms, and significance of macrophage plasticity. Annu. Rev. Pathol. 2020, 15, 123–147. [Google Scholar] [CrossRef] [PubMed]
- Kazankov, K.; Jørgensen, S.M.D.; Thomsen, K.L.; Møller, H.J.; Vilstrup, H.; George, J.; Schuppan, D.; Grønbæk, H. The role of macrophages in nonalcoholic fatty liver disease and nonalcoholic steatohepatitis. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 145–159. [Google Scholar] [CrossRef]
- Smith, K. Liver disease: Kupffer cells regulate the progression of ald and nafld. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 503. [Google Scholar] [CrossRef]
- Sulaiman, S.A.; Dorairaj, V.; Abdul Ghafar, K.N.; Abdul Murad, N.A. Noncoding rnas interactions in hepatic stellate cells during hepatic fibrosis. Livers 2021, 1, 263–285. [Google Scholar] [CrossRef]
- Yan, Y.; Zeng, J.; Xing, L.; Li, C. Extra- and intra-cellular mechanisms of hepatic stellate cell activation. Biomedicines 2021, 9, 1014. [Google Scholar] [CrossRef]
- Yang, S.J.; IglayReger, H.B.; Kadouh, H.C.; Bodary, P.F. Inhibition of the chemokine (c-c motif) ligand 2/chemokine (c-c motif) receptor 2 pathway attenuates hyperglycaemia and inflammation in a mouse model of hepatic steatosis and lipoatrophy. Diabetologia 2009, 52, 972–981. [Google Scholar] [CrossRef]
- Baeck, C.; Wehr, A.; Karlmark, K.R.; Heymann, F.; Vucur, M.; Gassler, N.; Huss, S.; Klussmann, S.; Eulberg, D.; Luedde, T.; et al. Pharmacological inhibition of the chemokine ccl2 (mcp-1) diminishes liver macrophage infiltration and steatohepatitis in chronic hepatic injury. Gut 2012, 61, 416–426. [Google Scholar] [CrossRef]
- Nio, Y.; Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Funata, M.; Yamaguchi, M.; Ueki, K.; Kadowaki, T. Monocyte chemoattractant protein-1 (mcp-1) deficiency enhances alternatively activated m2 macrophages and ameliorates insulin resistance and fatty liver in lipoatrophic diabetic a-zip transgenic mice. Diabetologia 2012, 55, 3350–3358. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Cervera, A.; Vinué, Á.; Burks, D.J.; González-Navarro, H. Genetic inactivation of the light (tnfsf14) cytokine in mice restores glucose homeostasis and diminishes hepatic steatosis. Diabetologia 2019, 62, 2143–2157. [Google Scholar] [CrossRef]
- Ambade, A.; Lowe, P.; Kodys, K.; Catalano, D.; Gyongyosi, B.; Cho, Y.; Iracheta-Vellve, A.; Adejumo, A.; Saha, B.; Calenda, C.; et al. Pharmacological inhibition of ccr2/5 signaling prevents and reverses alcohol-induced liver damage, steatosis, and inflammation in mice. Hepatology 2019, 69, 1105–1121. [Google Scholar] [CrossRef] [PubMed]
- Romeo, S.; Kozlitina, J.; Xing, C.; Pertsemlidis, A.; Cox, D.; Pennacchio, L.A.; Boerwinkle, E.; Cohen, J.C.; Hobbs, H.H. Genetic variation in pnpla3 confers susceptibility to nonalcoholic fatty liver disease. Nat. Genet. 2008, 40, 1461–1465. [Google Scholar] [CrossRef] [PubMed]
- Rotman, Y.; Koh, C.; Zmuda, J.M.; Kleiner, D.E.; Liang, T.J. The association of genetic variability in patatin-like phospholipase domain-containing protein 3 (pnpla3) with histological severity of nonalcoholic fatty liver disease. Hepatology 2010, 52, 894–903. [Google Scholar] [CrossRef]
- Gorden, A.; Yang, R.; Yerges-Armstrong, L.M.; Ryan, K.A.; Speliotes, E.; Borecki, I.B.; Harris, T.B.; Chu, X.; Wood, G.C.; Still, C.D.; et al. Genetic variation at ncan locus is associated with inflammation and fibrosis in non-alcoholic fatty liver disease in morbid obesity. Hum. Hered. 2013, 75, 34–43. [Google Scholar] [CrossRef]
- Mansoor, S.; Maheshwari, A.; Di Guglielmo, M.; Furuya, K.; Wang, M.; Crowgey, E.; Molle-Rios, Z.; He, Z. The pnpla3 rs738409 variant but not mboat7 rs641738 is a risk factor for nonalcoholic fatty liver disease in obese U.S. Children of hispanic ethnicity. Pediatr. Gastroenterol. Hepatol. Nutr. 2021, 24, 455–469. [Google Scholar] [CrossRef]
- Park, S.L.; Li, Y.; Sheng, X.; Hom, V.; Xia, L.; Zhao, K.; Pooler, L.; Setiawan, V.W.; Lim, U.; Monroe, K.R.; et al. Genome-wide association study of liver fat: The multiethnic cohort adiposity phenotype study. Hepatol. Commun. 2020, 4, 1112–1123. [Google Scholar] [CrossRef]
- Speliotes, E.K.; Yerges-Armstrong, L.M.; Wu, J.; Hernaez, R.; Kim, L.J.; Palmer, C.D.; Gudnason, V.; Eiriksdottir, G.; Garcia, M.E.; Launer, L.J.; et al. Genome-wide association analysis identifies variants associated with nonalcoholic fatty liver disease that have distinct effects on metabolic traits. PLoS Genet. 2011, 7, e1001324. [Google Scholar] [CrossRef]
- Hyysalo, J.; Stojkovic, I.; Kotronen, A.; Hakkarainen, A.; Sevastianova, K.; Makkonen, J.; Lundbom, N.; Rissanen, A.; Krauss, R.M.; Melander, O.; et al. Genetic variation in pnpla3 but not apoc3 influences liver fat in non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2012, 27, 951–956. [Google Scholar] [CrossRef]
- Li, Y.; Xing, C.; Tian, Z.; Ku, H.C. Genetic variant i148m in pnpla3 is associated with the ultrasonography-determined steatosis degree in a chinese population. BMC. Med. Genet. 2012, 13, 113. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.-R.; Song, J.-Y.; Liu, F.-H.; Ma, J.; Wang, H.-J. Gwas-identified common variants with nonalcoholic fatty liver disease in chinese children. J. Pediatr. Gastroenterol. Nutr. 2015, 60, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Z.; Wang, K.; Wang, Z.; Sun, X.; Zhong, L.; Deng, G.; Song, G.; Sun, B.; Peng, Z.; et al. Additive effects of the risk alleles of pnpla3 and tm6sf2 on non-alcoholic fatty liver disease (nafld) in a chinese population. Front. Genet. 2016, 7, 140. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.F.; Ling, Y.; Bian, H.; Lin, H.D.; Yan, H.M.; Chang, X.X.; Li, X.M.; Ma, H.; Wang, D.; Zhang, L.S.; et al. I148m variant of pnpla3 increases the susceptibility to non-alcoholic fatty liver disease caused by obesity and metabolic disorders. Aliment. Pharmacol. Ther. 2016, 43, 631–642. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.W.; Lin, H.Y.; Shin, S.J.; Yu, M.L.; Lin, Z.Y.; Dai, C.Y.; Huang, J.F.; Chen, S.C.; Li, S.S.; Chuang, W.L. The pnpla3 i148m polymorphism is associated with insulin resistance and nonalcoholic fatty liver disease in a normoglycaemic population. Liver Int. 2011, 31, 1326–1331. [Google Scholar] [CrossRef]
- Hudert, C.A.; Selinski, S.; Rudolph, B.; Bläker, H.; Loddenkemper, C.; Thielhorn, R.; Berndt, N.; Golka, K.; Cadenas, C.; Reinders, J.; et al. Genetic determinants of steatosis and fibrosis progression in paediatric non-alcoholic fatty liver disease. Liver Int. 2019, 39, 540–556. [Google Scholar] [CrossRef]
- Krawczyk, M.; Rau, M.; Schattenberg, J.M.; Bantel, H.; Pathil, A.; Demir, M.; Kluwe, J.; Boettler, T.; Lammert, F.; Geier, A. Combined effects of the pnpla3 rs738409, tm6sf2 rs58542926, and mboat7 rs641738 variants on nafld severity: A multicenter biopsy-based study. J. Lipid Res. 2017, 58, 247–255. [Google Scholar] [CrossRef]
- Kitamoto, T.; Kitamoto, A.; Yoneda, M.; Hyogo, H.; Ochi, H.; Nakamura, T.; Teranishi, H.; Mizusawa, S.; Ueno, T.; Chayama, K.; et al. Genome-wide scan revealed that polymorphisms in the pnpla3, samm50, and parvb genes are associated with development and progression of nonalcoholic fatty liver disease in japan. Hum. Genet. 2013, 132, 783–792. [Google Scholar] [CrossRef]
- Di Costanzo, A.; Belardinilli, F.; Bailetti, D.; Sponziello, M.; D’Erasmo, L.; Polimeni, L.; Baratta, F.; Pastori, D.; Ceci, F.; Montali, A.; et al. Evaluation of polygenic determinants of non-alcoholic fatty liver disease (nafld) by a candidate genes resequencing strategy. Sci. Rep. 2018, 8, 3702. [Google Scholar] [CrossRef]
- Valenti, L.; Al-Serri, A.; Daly, A.K.; Galmozzi, E.; Rametta, R.; Dongiovanni, P.; Nobili, V.; Mozzi, E.; Roviaro, G.; Vanni, E.; et al. Homozygosity for the patatin-like phospholipase-3/adiponutrin i148m polymorphism influences liver fibrosis in patients with nonalcoholic fatty liver disease. Hepatology 2010, 51, 1209–1217. [Google Scholar] [CrossRef]
- Vespasiani-Gentilucci, U.; Gallo, P.; Porcari, A.; Carotti, S.; Galati, G.; Piccioni, L.; De Vincentis, A.; Dell’Unto, C.; Vorini, F.; Morini, S.; et al. The pnpla3 rs738409 c > g polymorphism is associated with the risk of progression to cirrhosis in nafld patients. Scand. J. Gastroenterol. 2016, 51, 967–973. [Google Scholar] [CrossRef]
- Longo, M.; Meroni, M.; Paolini, E.; Erconi, V.; Carli, F.; Fortunato, F.; Ronchi, D.; Piciotti, R.; Sabatini, S.; Macchi, C.; et al. Tm6sf2/pnpla3/mboat7 loss-of-function genetic variants impact on nafld development and progression both in patients and in in vitro models. Cell Mol. Gastroenterol. Hepatol. 2022, 13, 759–788. [Google Scholar] [CrossRef]
- Verrijken, A.; Beckers, S.; Francque, S.; Hilden, H.; Caron, S.; Zegers, D.; Ruppert, M.; Hubens, G.; Van Marck, E.; Michielsen, P.; et al. A gene variant of pnpla3, but not of apoc3, is associated with histological parameters of nafld in an obese population. Obesity 2013, 21, 2138–2145. [Google Scholar] [CrossRef]
- Lisboa, Q.C.; Nardelli, M.J.; Pereira, P.A.; Miranda, D.M.; Ribeiro, S.N.; Costa, R.S.N.; Versiani, C.A.; Vidigal, P.V.T.; Ferrari, T.C.A.; Couto, C.A. Pnpla3 and tm6sf2 polymorphisms in brazilian patients with nonalcoholic fatty liver disease. World J. Hepatol. 2020, 12, 792–806. [Google Scholar] [CrossRef]
- Zain, S.M.; Mohamed, R.; Mahadeva, S.; Cheah, P.L.; Rampal, S.; Basu, R.C.; Mohamed, Z. A multi-ethnic study of a pnpla3 gene variant and its association with disease severity in non-alcoholic fatty liver disease. Hum. Genet. 2012, 131, 1145–1152. [Google Scholar] [CrossRef]
- Uygun, A.; Ozturk, K.; Demirci, H.; Oztuna, A.; Eren, F.; Kozan, S.; Yilmaz, Y.; Kurt, O.; Turker, T.; Vatansever, S.; et al. The association of nonalcoholic fatty liver disease with genetic polymorphisms: A multicenter study. Eur. J. Gastroenterol. Hepatol. 2017, 29, 441–447. [Google Scholar] [CrossRef]
- Idilman, R.; Karatayli, S.C.; Kabacam, G.; Savas, B.; Elhan, A.H.; Bozdayi, A.M. The role of pnpla3 (rs738409) c>g variant on histological progression of non-alcoholic fatty liver disease. Hepatol. Forum 2020, 1, 82–87. [Google Scholar] [CrossRef]
- Li, X.; Zhao, Q.; Wu, K.; Fan, D. I148m variant of pnpla3 confer increased risk for nonalcoholic fatty liver disease not only in european population, but also in chinese population. Hepatology 2011, 54, 2276. [Google Scholar] [CrossRef]
- Lin, H.; Wong, G.L.; Whatling, C.; Chan, A.W.; Leung, H.H.; Tse, C.H.; Shu, S.S.; Chim, A.M.; Lai, J.C.; Yip, T.C.; et al. Association of genetic variations with nafld in lean individuals. Liver Int. 2022, 42, 149–160. [Google Scholar] [CrossRef]
- Hotta, K.; Yoneda, M.; Hyogo, H.; Ochi, H.; Mizusawa, S.; Ueno, T.; Chayama, K.; Nakajima, A.; Nakao, K.; Sekine, A. Association of the rs738409 polymorphism in pnpla3 with liver damage and the development of nonalcoholic fatty liver disease. BMC Med. Genet. 2010, 11, 172. [Google Scholar] [CrossRef]
- Kawaguchi, T.; Sumida, Y.; Umemura, A.; Matsuo, K.; Takahashi, M.; Takamura, T.; Yasui, K.; Saibara, T.; Hashimoto, E.; Kawanaka, M.; et al. Genetic polymorphisms of the human pnpla3 gene are strongly associated with severity of non-alcoholic fatty liver disease in japanese. PLoS ONE 2012, 7, e38322. [Google Scholar] [CrossRef]
- Lee, S.S.; Byoun, Y.S.; Jeong, S.H.; Woo, B.H.; Jang, E.S.; Kim, J.W.; Kim, H.Y. Role of the pnpla3 i148m polymorphism in nonalcoholic fatty liver disease and fibrosis in korea. Dig. Dis. Sci. 2014, 59, 2967–2974. [Google Scholar] [CrossRef]
- Park, J.H.; Cho, B.; Kwon, H.; Prilutsky, D.; Yun, J.M.; Choi, H.C.; Hwang, K.B.; Lee, I.H.; Kim, J.I.; Kong, S.W. I148m variant in pnpla3 reduces central adiposity and metabolic disease risks while increasing nonalcoholic fatty liver disease. Liver Int. 2015, 35, 2537–2546. [Google Scholar] [CrossRef]
- Bhatt, S.P.; Nigam, P.; Misra, A.; Guleria, R.; Pandey, R.M.; Pasha, M.A. Genetic variation in the patatin-like phospholipase domain-containing protein-3 (pnpla-3) gene in asian indians with nonalcoholic fatty liver disease. Metab. Syndr. Relat. Disord. 2013, 11, 329–335. [Google Scholar] [CrossRef]
- Karoli, R.; Fatima, J.; Singh, P.S.; Siddiqi, Z.; Varshney, S.; Beg, M.S.; Khan, M.A. Association of genetic non-alcoholic fatty liver disease with insulin resistance-are we different? J. Assoc. Physicians India 2019, 67, 34–38. [Google Scholar]
- Kanth, V.V.; Sasikala, M.; Rao, P.N.; Steffie Avanthi, U.; Rao, K.R.; Nageshwar Reddy, D. Pooled genetic analysis in ultrasound measured non-alcoholic fatty liver disease in indian subjects: A pilot study. World J. Hepatol. 2014, 6, 435–442. [Google Scholar] [CrossRef]
- Narayanasamy, K.; Karthick, R.; Panneerselvam, P.; Mohan, N.; Ramachandran, A.; Prakash, R.; Rajaram, M. Association of metabolic syndrome and patatin-like phospholipase 3–rs738409 gene variant in non-alcoholic fatty liver disease among a chennai-based south indian population. J. Gene Med. 2020, 22, e3160. [Google Scholar] [CrossRef]
- Niriella, M.A.; Pathmeswaran, A.; De Silva, S.T.; Kasturiratna, A.; Perera, R.; Subasinghe, C.E.; Kodisinghe, K.; Piyaratna, C.; Rishikesawan, V.; Dassanayaka, A.S.; et al. Incidence and risk factors for non-alcoholic fatty liver disease: A 7-year follow-up study among urban, adult sri lankans. Liver Int. 2017, 37, 1715–1722. [Google Scholar] [CrossRef]
- Alam, S.; Islam, M.S.; Islam, S.; Mustafa, G.; Saleh, A.A.; Ahmad, N. Association of single nucleotide polymorphism at pnpla3 with fatty liver, steatohepatitis, and cirrhosis of liver. Indian J. Gastroenterol. 2017, 36, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Choobini, N.; Azarpira, N.; Mashayekhi, M.R. Association of pnpla3 gene polymorphism (rs738409) and nonalcoholic fatty liver disease in southern iranian population. JABS 2016, 6, 60–68. [Google Scholar]
- Lee, G.H.; Phyo, W.W.; Loo, W.M.; Kwok, R.; Ahmed, T.; Shabbir, A.; So, J.; Koh, C.J.; Hartono, J.L.; Muthiah, M.; et al. Validation of genetic variants associated with metabolic dysfunction-associated fatty liver disease in an ethnic chinese population. World J. Hepatol. 2020, 12, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Basu, A.; Das, K.; Chowdhury, A.; Basu, P. Exome-wide scan identifies significant association of rs4788084 in il27 promoter with increase in hepatic fat content among indians. Gene 2021, 775, 145431. [Google Scholar] [CrossRef]
- Baclig, M.O.; Lozano-Kühne, J.P.; Mapua, C.A.; Gopez-Cervantes, J.; Natividad, F.F. Genetic variation i148m in patatin-like phospholipase 3 gene and risk of non-alcoholic fatty liver disease among filipinos. Int. J. Clin. Exp. Med. 2014, 7, 2129–2136. [Google Scholar]
- Kawaguchi, T.; Shima, T.; Mizuno, M.; Mitsumoto, Y.; Umemura, A.; Kanbara, Y.; Tanaka, S.; Sumida, Y.; Yasui, K.; Takahashi, M.; et al. Risk estimation model for nonalcoholic fatty liver disease in the japanese using multiple genetic markers. PLoS ONE 2018, 13, e0185490. [Google Scholar] [CrossRef]
- Kozlitina, J.; Smagris, E.; Stender, S.; Nordestgaard, B.G.; Zhou, H.H.; Tybjærg-Hansen, A.; Vogt, T.F.; Hobbs, H.H.; Cohen, J.C. Exome-wide association study identifies a tm6sf2 variant that confers susceptibility to nonalcoholic fatty liver disease. Nat. Genet. 2014, 46, 352–356. [Google Scholar] [CrossRef]
- Eslam, M.; Mangia, A.; Berg, T.; Chan, H.L.; Irving, W.L.; Dore, G.J.; Abate, M.L.; Bugianesi, E.; Adams, L.A.; Najim, M.A.; et al. Diverse impacts of the rs58542926 e167k variant in tm6sf2 on viral and metabolic liver disease phenotypes. Hepatology 2016, 64, 34–46. [Google Scholar] [CrossRef]
- Sookoian, S.; Castaño, G.O.; Scian, R.; Mallardi, P.; Fernández Gianotti, T.; Burgueño, A.L.; San Martino, J.; Pirola, C.J. Genetic variation in transmembrane 6 superfamily member 2 and the risk of nonalcoholic fatty liver disease and histological disease severity. Hepatology 2015, 61, 515–525. [Google Scholar] [CrossRef]
- Zhou, Y.; Llauradó, G.; Orešič, M.; Hyötyläinen, T.; Orho-Melander, M.; Yki-Järvinen, H. Circulating triacylglycerol signatures and insulin sensitivity in nafld associated with the e167k variant in tm6sf2. J. Hepatol. 2015, 62, 657–663. [Google Scholar] [CrossRef]
- Krawczyk, M.; Stachowska, E.; Milkiewicz, P.; Lammert, F.; Milkiewicz, M. Reduction of caloric intake might override the prosteatotic effects of the pnpla3 p.I148m and tm6sf2 p.E167k variants in patients with fatty liver: Ultrasound-based prospective study. Digestion 2016, 93, 139–148. [Google Scholar] [CrossRef]
- Grandone, A.; Cozzolino, D.; Marzuillo, P.; Cirillo, G.; Di Sessa, A.; Ruggiero, L.; Di Palma, M.R.; Perrone, L.; Miraglia Del Giudice, E. Tm6sf2 glu167lys polymorphism is associated with low levels of ldl-cholesterol and increased liver injury in obese children. Pediatr. Obes. 2016, 11, 115–119. [Google Scholar] [CrossRef]
- Mancina, R.M.; Sentinelli, F.; Incani, M.; Bertoccini, L.; Russo, C.; Romeo, S.; Baroni, M.G. Transmembrane-6 superfamily member 2 (tm6sf2) e167k variant increases susceptibility to hepatic steatosis in obese children. Dig. Liver Dis. 2016, 48, 100–101. [Google Scholar] [CrossRef]
- Musso, G.; Cipolla, U.; Cassader, M.; Pinach, S.; Saba, F.; De Michieli, F.; Paschetta, E.; Bongiovanni, D.; Framarin, L.; Leone, N.; et al. Tm6sf2 rs58542926 variant affects postprandial lipoprotein metabolism and glucose homeostasis in nafld. J. Lipid Res. 2017, 58, 1221–1229. [Google Scholar] [CrossRef]
- Goffredo, M.; Caprio, S.; Feldstein, A.E.; D’Adamo, E.; Shaw, M.M.; Pierpont, B.; Savoye, M.; Zhao, H.; Bale, A.E.; Santoro, N. Role of tm6sf2 rs58542926 in the pathogenesis of nonalcoholic pediatric fatty liver disease: A multiethnic study. Hepatology 2016, 63, 117–125. [Google Scholar] [CrossRef]
- Liu, Y.L.; Reeves, H.L.; Burt, A.D.; Tiniakos, D.; McPherson, S.; Leathart, J.B.; Allison, M.E.; Alexander, G.J.; Piguet, A.C.; Anty, R.; et al. Tm6sf2 rs58542926 influences hepatic fibrosis progression in patients with non-alcoholic fatty liver disease. Nat. Commun. 2014, 5, 4309. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Z.; Luan, G.P.; Qun, L.I.U.; Hui, G.A.O.; Xin, Y.N.; Xuan, S.Y. Association between tm6sf2 rs58542926 polymorphism and non-alcoholic fatty liver disease in qingdao han population and molecular mechanism. PLAMJ 2019, 44, 127–131. [Google Scholar]
- Xu, M.; Li, Y.; Zhang, S.; Wang, X.; Shen, J.; Zhang, S. Interaction of tm6sf2 e167k and pnpla3 i148m variants in nafld in northeast china. Ann. Hepatol. 2019, 18, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, Z.; Peng, Z.; Liu, W. The tm6sf2 rs58542926 t allele is significantly associated with non-alcoholic fatty liver disease in chinese. J. Hepatol. 2015, 62, 1438–1439. [Google Scholar] [CrossRef]
- Dongiovanni, P.; Petta, S.; Maglio, C.; Fracanzani, A.L.; Pipitone, R.; Mozzi, E.; Motta, B.M.; Kaminska, D.; Rametta, R.; Grimaudo, S.; et al. Transmembrane 6 superfamily member 2 gene variant disentangles nonalcoholic steatohepatitis from cardiovascular disease. Hepatology 2015, 61, 506–514. [Google Scholar] [CrossRef]
- Koo, B.K.; Joo, S.K.; Kim, D.; Bae, J.M.; Park, J.H.; Kim, J.H.; Kim, W. Additive effects of pnpla3 and tm6sf2 on the histological severity of non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2018, 33, 1277–1285. [Google Scholar] [CrossRef]
- Bale, G.; Steffie, A.U.; Ravi Kanth, V.V.; Rao, P.N.; Sharma, M.; Sasikala, M.; Reddy, D.N. Regional differences in genetic susceptibility to non-alcoholic liver disease in two distinct indian ethnicities. World J. Hepatol. 2017, 9, 1101–1107. [Google Scholar] [CrossRef]
- Akuta, N.; Kawamura, Y.; Arase, Y.; Suzuki, F.; Sezaki, H.; Hosaka, T.; Kobayashi, M.; Kobayashi, M.; Saitoh, S.; Suzuki, Y.; et al. Relationships between genetic variations of pnpla3, tm6sf2 and histological features of nonalcoholic fatty liver disease in japan. Gut Liver 2016, 10, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Hernaez, R.; McLean, J.; Lazo, M.; Brancati, F.L.; Hirschhorn, J.N.; Borecki, I.B.; Harris, T.B.; Nguyen, T.; Kamel, I.R.; Bonekamp, S.; et al. Association between variants in or near pnpla3, gckr, and ppp1r3b with ultrasound-defined steatosis based on data from the third national health and nutrition examination survey. Clin. Gastroenterol. Hepatol. 2013, 11, 1183–1190.e2. [Google Scholar] [CrossRef] [PubMed]
- Petta, S.; Miele, L.; Bugianesi, E.; Cammà, C.; Rosso, C.; Boccia, S.; Cabibi, D.; Di Marco, V.; Grimaudo, S.; Grieco, A.; et al. Glucokinase regulatory protein gene polymorphism affects liver fibrosis in non-alcoholic fatty liver disease. PLoS ONE 2014, 9, e87523. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Chang, P.F.; Chang, M.H.; Ni, Y.H. Genetic variants in gckr and pnpla3 confer susceptibility to nonalcoholic fatty liver disease in obese individuals. Am. J. Clin. Nutr. 2014, 99, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.; Weng, D.H.; Yan, P.; Lin, Y.T.; Dong, Z.H.; Mailamuguli; Yao, H. Genetic polymorphisms associated with nonalcoholic fatty liver disease in uyghur population: A case-control study and meta-analysis. Lipids Health Dis. 2019, 18, 14. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.L.; Zain, S.M.; Mohamed, R.; Rampal, S.; Chin, K.F.; Basu, R.C.; Cheah, P.L.; Mahadeva, S.; Mohamed, Z. Association of glucokinase regulatory gene polymorphisms with risk and severity of non-alcoholic fatty liver disease: An interaction study with adiponutrin gene. J. Gastroenterol. 2014, 49, 1056–1064. [Google Scholar] [CrossRef]
- Mohammadi, S.; Farajnia, S.; Shadmand, M.; Mohseni, F.; Baghban, R. Association of rs780094 polymorphism of glucokinase regulatory protein with non-alcoholic fatty liver disease. BMC Res. Notes 2020, 13, 26. [Google Scholar] [CrossRef]
- Gao, H.; Liu, S.; Zhao, Z.; Yu, X.; Liu, Q.; Xin, Y.; Xuan, S. Association of gckr gene polymorphisms with the risk of nonalcoholic fatty liver disease and coronary artery disease in a chinese northern han population. J. Clin. Transl. Hepatol. 2019, 7, 297–303. [Google Scholar] [CrossRef]
- Yuan, F.; Gu, Z.; Bi, Y.; Yuan, R.; Niu, W.; Ren, D.; Zhang, L.; He, G.; Liu, B.C. The association between rs1260326 with the risk of nafld and the mediation effect of triglyceride on nafld in the elderly chinese han population. Aging 2022, 14, 2736–2747. [Google Scholar] [CrossRef]
- Yang, Z.; Wen, J.; Tao, X.; Lu, B.; Du, Y.; Wang, M.; Wang, X.; Zhang, W.; Gong, W.; Ling, C.; et al. Genetic variation in the gckr gene is associated with non-alcoholic fatty liver disease in chinese people. Mol. Biol. Rep. 2011, 38, 1145–1150. [Google Scholar] [CrossRef]
- Sliz, E.; Sebert, S.; Würtz, P.; Kangas, A.J.; Soininen, P.; Lehtimäki, T.; Kähönen, M.; Viikari, J.; Männikkö, M.; Ala-Korpela, M.; et al. Nafld risk alleles in pnpla3, tm6sf2, gckr and lyplal1 show divergent metabolic effects. Hum. Mol. Genet. 2018, 27, 2214–2223. [Google Scholar] [CrossRef]
- Liao, S.; An, K.; Liu, Z.; He, H.; An, Z.; Su, Q.; Li, S. Genetic variants associated with metabolic dysfunction-associated fatty liver disease in western china. J. Clin. Lab. Anal. 2022, 36, e24626. [Google Scholar] [CrossRef]
- Mancina, R.M.; Dongiovanni, P.; Petta, S.; Pingitore, P.; Meroni, M.; Rametta, R.; Borén, J.; Montalcini, T.; Pujia, A.; Wiklund, O.; et al. The mboat7-tmc4 variant rs641738 increases risk of nonalcoholic fatty liver disease in individuals of european descent. Gastroenterology 2016, 150, 1219–1230.e6. [Google Scholar] [CrossRef]
- Luukkonen, P.K.; Zhou, Y.; Hyötyläinen, T.; Leivonen, M.; Arola, J.; Orho-Melander, M.; Orešič, M.; Yki-Järvinen, H. The mboat7 variant rs641738 alters hepatic phosphatidylinositols and increases severity of non-alcoholic fatty liver disease in humans. J. Hepatol. 2016, 65, 1263–1265. [Google Scholar] [CrossRef]
- Basyte-Bacevice, V.; Skieceviciene, J.; Valantiene, I.; Sumskiene, J.; Petrenkiene, V.; Kondrackiene, J.; Petrauskas, D.; Lammert, F.; Kupcinskas, J. Tm6sf2 and mboat7 gene variants in liver fibrosis and cirrhosis. Int. J. Mol. Sci. 2019, 20, 1277. [Google Scholar] [CrossRef]
- Zusi, C.; Morandi, A.; Maguolo, A.; Corradi, M.; Costantini, S.; Mosca, A.; Crudele, A.; Mantovani, A.; Alisi, A.; Miraglia Del Giudice, E.; et al. Association between mboat7 rs641738 polymorphism and non-alcoholic fatty liver in overweight or obese children. Nutr. Metab. Cardiovasc. Dis 2021, 31, 1548–1555. [Google Scholar] [CrossRef]
- Sookoian, S.; Flichman, D.; Garaycoechea, M.E.; Gazzi, C.; Martino, J.S.; Castaño, G.O.; Pirola, C.J. Lack of evidence supporting a role of tmc4-rs641738 missense variant-mboat7- intergenic downstream variant-in the susceptibility to nonalcoholic fatty liver disease. Sci. Rep. 2018, 8, 5097. [Google Scholar] [CrossRef]
- Lin, Y.C.; Chang, P.F.; Chang, M.H.; Ni, Y.H. Genetic determinants of hepatic steatosis and serum cytokeratin-18 fragment levels in taiwanese children. Liver Int. 2018, 38, 1300–1307. [Google Scholar] [CrossRef]
- Ma, Y.; Belyaeva, O.V.; Brown, P.M.; Fujita, K.; Valles, K.; Karki, S.; De Boer, Y.S.; Koh, C.; Chen, Y.; Du, X.; et al. 17-beta hydroxysteroid dehydrogenase 13 is a hepatic retinol dehydrogenase associated with histological features of nonalcoholic fatty liver disease. Hepatology 2019, 69, 1504–1519. [Google Scholar] [CrossRef]
- Vilar-Gomez, E.; Pirola, C.J.; Sookoian, S.; Wilson, L.A.; Liang, T.; Chalasani, N. The protection conferred by hsd17b13 rs72613567 polymorphism on risk of steatohepatitis and fibrosis may be limited to selected subgroups of patients with nafld. Clin. Transl. Gastroenterol. 2021, 12, e00400. [Google Scholar] [CrossRef]
- Abul-Husn, N.S.; Cheng, X.; Li, A.H.; Xin, Y.; Schurmann, C.; Stevis, P.; Liu, Y.; Kozlitina, J.; Stender, S.; Wood, G.C.; et al. A protein-truncating hsd17b13 variant and protection from chronic liver disease. N. Engl. J. Med. 2018, 378, 1096–1106. [Google Scholar] [CrossRef] [PubMed]
- Luukkonen, P.K.; Tukiainen, T.; Juuti, A.; Sammalkorpi, H.; Haridas, P.A.N.; Niemelä, O.; Arola, J.; Orho-Melander, M.; Hakkarainen, A.; Kovanen, P.T.; et al. Hydroxysteroid 17-β dehydrogenase 13 variant increases phospholipids and protects against fibrosis in nonalcoholic fatty liver disease. JCI Insight 2020, 5, e132158. [Google Scholar] [CrossRef] [PubMed]
- Pirola, C.J.; Garaycoechea, M.; Flichman, D.; Arrese, M.; San Martino, J.; Gazzi, C.; Castaño, G.O.; Sookoian, S. Splice variant rs72613567 prevents worst histologic outcomes in patients with nonalcoholic fatty liver disease. J. Lipid Res. 2019, 60, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Seko, Y.; Yamaguchi, K.; Tochiki, N.; Yano, K.; Takahashi, A.; Okishio, S.; Kataoka, S.; Okuda, K.; Umemura, A.; Moriguchi, M.; et al. Attenuated effect of pnpla3 on hepatic fibrosis by hsd17b13 in japanese patients with non-alcoholic fatty liver disease. Liver Int. 2020, 40, 1686–1692. [Google Scholar] [CrossRef] [PubMed]
- Koo, B.K.; Joo, S.K.; Kim, D.; Lee, S.; Bae, J.M.; Park, J.H.; Kim, J.H.; Chang, M.S.; Kim, W. Development and validation of a scoring system, based on genetic and clinical factors, to determine risk of steatohepatitis in asian patients with nonalcoholic fatty liver disease. Clin. Gastroenterol. Hepatol. 2020, 18, 2592–2599.e10. [Google Scholar] [CrossRef]
- Pirazzi, C.; Adiels, M.; Burza, M.A.; Mancina, R.M.; Levin, M.; Ståhlman, M.; Taskinen, M.R.; Orho-Melander, M.; Perman, J.; Pujia, A.; et al. Patatin-like phospholipase domain-containing 3 (pnpla3) i148m (rs738409) affects hepatic vldl secretion in humans and in vitro. J. Hepatol. 2012, 57, 1276–1282. [Google Scholar] [CrossRef]
- Salari, N.; Darvishi, N.; Mansouri, K.; Ghasemi, H.; Hosseinian-Far, M.; Darvishi, F.; Mohammadi, M. Association between pnpla3 rs738409 polymorphism and nonalcoholic fatty liver disease: A systematic review and meta-analysis. BMC Endocr. Disord. 2021, 21, 125. [Google Scholar] [CrossRef]
- Kantartzis, K.; Peter, A.; Machicao, F.; Machann, J.; Wagner, S.; Königsrainer, I.; Königsrainer, A.; Schick, F.; Fritsche, A.; Häring, H.U.; et al. Dissociation between fatty liver and insulin resistance in humans carrying a variant of the patatin-like phospholipase 3 gene. Diabetes 2009, 58, 2616–2623. [Google Scholar] [CrossRef]
- Davis, J.N.; Lê, K.A.; Walker, R.W.; Vikman, S.; Spruijt-Metz, D.; Weigensberg, M.J.; Allayee, H.; Goran, M.I. Increased hepatic fat in overweight hispanic youth influenced by interaction between genetic variation in pnpla3 and high dietary carbohydrate and sugar consumption. Am. J. Clin. Nutr. 2010, 92, 1522–1527. [Google Scholar] [CrossRef]
- Dubuquoy, C.; Robichon, C.; Lasnier, F.; Langlois, C.; Dugail, I.; Foufelle, F.; Girard, J.; Burnol, A.F.; Postic, C.; Moldes, M. Distinct regulation of adiponutrin/pnpla3 gene expression by the transcription factors chrebp and srebp1c in mouse and human hepatocytes. J. Hepatol. 2011, 55, 145–153. [Google Scholar] [CrossRef]
- Perttilä, J.; Huaman-Samanez, C.; Caron, S.; Tanhuanpää, K.; Staels, B.; Yki-Järvinen, H.; Olkkonen, V.M. Pnpla3 is regulated by glucose in human hepatocytes, and its i148m mutant slows down triglyceride hydrolysis. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1063–E1069. [Google Scholar] [CrossRef]
- Sookoian, S.; Pirola, C.J. Meta-analysis of the influence of i148m variant of patatin-like phospholipase domain containing 3 gene (pnpla3) on the susceptibility and histological severity of nonalcoholic fatty liver disease. Hepatology 2011, 53, 1883–1894. [Google Scholar] [CrossRef]
- Salameh, H.; Hanayneh, M.A.; Masadeh, M.; Naseemuddin, M.; Matin, T.; Erwin, A.; Singal, A.K. Pnpla3 as a genetic determinant of risk for and severity of non-alcoholic fatty liver disease spectrum. J. Clin. Transl. Hepatol. 2016, 4, 175–191. [Google Scholar]
- Zhou, Y.; Orešič, M.; Leivonen, M.; Gopalacharyulu, P.; Hyysalo, J.; Arola, J.; Verrijken, A.; Francque, S.; Van Gaal, L.; Hyötyläinen, T.; et al. Noninvasive detection of nonalcoholic steatohepatitis using clinical markers and circulating levels of lipids and metabolites. Clin. Gastroenterol. Hepatol. 2016, 14, 1463–1472.e6. [Google Scholar] [CrossRef]
- Hyysalo, J.; Männistö, V.T.; Zhou, Y.; Arola, J.; Kärjä, V.; Leivonen, M.; Juuti, A.; Jaser, N.; Lallukka, S.; Käkelä, P.; et al. A population-based study on the prevalence of nash using scores validated against liver histology. J. Hepatol. 2014, 60, 839–846. [Google Scholar] [CrossRef]
- Segura-Azuara, N.; Varela-Chinchilla, C.D.; Trinidad-Calderón, P.A. Mafld/nafld biopsy-free scoring systems for hepatic steatosis, nash, and fibrosis diagnosis. Front. Med. 2021, 8, 774079. [Google Scholar] [CrossRef]
- Liu, Z.; Que, S.; Zhou, L.; Zheng, S.; Romeo, S.; Mardinoglu, A.; Valenti, L. The effect of the tm6sf2 e167k variant on liver steatosis and fibrosis in patients with chronic hepatitis c: A meta-analysis. Sci. Rep. 2017, 7, 9273. [Google Scholar] [CrossRef]
- Li, X.Y.; Liu, Z.; Li, L.; Wang, H.J.; Wang, H. Tm6sf2 rs58542926 is related to hepatic steatosis, fibrosis and serum lipids both in adults and children: A meta-analysis. Front. Endocrinol. 2022, 13, 1026901. [Google Scholar] [CrossRef]
- Sookoian, S.; Pirola, C.J. Meta-analysis of the influence of tm6sf2 e167k variant on plasma concentration of aminotransferases across different populations and diverse liver phenotypes. Sci. Rep. 2016, 6, 27718. [Google Scholar] [CrossRef]
- Chen, F.; Esmaili, S.; Rogers, G.B.; Bugianesi, E.; Petta, S.; Marchesini, G.; Bayoumi, A.; Metwally, M.; Azardaryany, M.K.; Coulter, S.; et al. Lean nafld: A distinct entity shaped by differential metabolic adaptation. Hepatology 2020, 71, 1213–1227. [Google Scholar] [CrossRef]
- Young, S.; Tariq, R.; Provenza, J.; Satapathy, S.K.; Faisal, K.; Choudhry, A.; Friedman, S.L.; Singal, A.K. Prevalence and profile of nonalcoholic fatty liver disease in lean adults: Systematic review and meta-analysis. Hepatol. Commun. 2020, 4, 953–972. [Google Scholar] [CrossRef] [PubMed]
- Kozlitina, J. Genetic risk factors and disease modifiers of nonalcoholic steatohepatitis. Gastroenterol. Clin. N. Am. 2020, 49, 25–44. [Google Scholar] [CrossRef] [PubMed]
- Holmen, O.L.; Zhang, H.; Fan, Y.; Hovelson, D.H.; Schmidt, E.M.; Zhou, W.; Guo, Y.; Zhang, J.; Langhammer, A.; Løchen, M.L.; et al. Systematic evaluation of coding variation identifies a candidate causal variant in tm6sf2 influencing total cholesterol and myocardial infarction risk. Nat. Genet. 2014, 46, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Pirola, C.J.; Sookoian, S. The dual and opposite role of the tm6sf2-rs58542926 variant in protecting against cardiovascular disease and conferring risk for nonalcoholic fatty liver: A meta-analysis. Hepatology 2015, 62, 1742–1756. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Oldoni, F.; Das, A. Tm6sf2: A novel genetic player in nonalcoholic fatty liver and cardiovascular disease. Hepatol. Commun. 2022, 6, 448–460. [Google Scholar] [CrossRef]
- Smagris, E.; Gilyard, S.; BasuRay, S.; Cohen, J.C.; Hobbs, H.H. Inactivation of tm6sf2, a gene defective in fatty liver disease, impairs lipidation but not secretion of very low density lipoproteins. J. Biol. Chem. 2016, 291, 10659–10676. [Google Scholar] [CrossRef]
- Simons, N.; Isaacs, A.; Koek, G.H.; Kuč, S.; Schaper, N.C.; Brouwers, M. Pnpla3, tm6sf2, and mboat7 genotypes and coronary artery disease. Gastroenterology 2017, 152, 912–913. [Google Scholar] [CrossRef]
- Liu, D.J.; Peloso, G.M.; Yu, H.; Butterworth, A.S.; Wang, X.; Mahajan, A.; Saleheen, D.; Emdin, C.; Alam, D.; Alves, A.C.; et al. Exome-wide association study of plasma lipids in >300,000 individuals. Nat. Genet. 2017, 49, 1758–1766. [Google Scholar] [CrossRef]
- Fang, Y.L.; Chen, H.; Wang, C.L.; Liang, L. Pathogenesis of non-alcoholic fatty liver disease in children and adolescence: From “two hit theory” to “multiple hit model”. World J. Gastroenterol. 2018, 24, 2974–2983. [Google Scholar] [CrossRef]
- Eslam, M.; Valenti, L.; Romeo, S. Genetics and epigenetics of nafld and nash: Clinical impact. J. Hepatol. 2018, 68, 268–279. [Google Scholar] [CrossRef]
- Zain, S.M.; Mohamed, Z.; Mohamed, R. Common variant in the glucokinase regulatory gene rs780094 and risk of nonalcoholic fatty liver disease: A meta-analysis. J. Gastroenterol. Hepatol. 2015, 30, 21–27. [Google Scholar] [CrossRef]
- Li, J.; Zhao, Y.; Zhang, H.; Hua, W.; Jiao, W.; Du, X.; Rui, J.; Li, S.; Teng, H.; Shi, B.; et al. Contribution of rs780094 and rs1260326 polymorphisms in gckr gene to non-alcoholic fatty liver disease: A meta-analysis involving 26,552 participants. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 1696–1708. [Google Scholar] [CrossRef]
- Sparsø, T.; Andersen, G.; Nielsen, T.; Burgdorf, K.S.; Gjesing, A.P.; Nielsen, A.L.; Albrechtsen, A.; Rasmussen, S.S.; Jørgensen, T.; Borch-Johnsen, K.; et al. The gckr rs780094 polymorphism is associated with elevated fasting serum triacylglycerol, reduced fasting and ogtt-related insulinaemia, and reduced risk of type 2 diabetes. Diabetologia 2008, 51, 70–75. [Google Scholar] [CrossRef]
- Beer, N.L.; Tribble, N.D.; McCulloch, L.J.; Roos, C.; Johnson, P.R.; Orho-Melander, M.; Gloyn, A.L. The p446l variant in gckr associated with fasting plasma glucose and triglyceride levels exerts its effect through increased glucokinase activity in liver. Hum. Mol. Genet 2009, 18, 4081–4088. [Google Scholar] [CrossRef]
- Santoro, N.; Zhang, C.K.; Zhao, H.; Pakstis, A.J.; Kim, G.; Kursawe, R.; Dykas, D.J.; Bale, A.E.; Giannini, C.; Pierpont, B.; et al. Variant in the glucokinase regulatory protein (gckr) gene is associated with fatty liver in obese children and adolescents. Hepatology 2012, 55, 781–789. [Google Scholar] [CrossRef]
- Thabet, K.; Chan, H.L.Y.; Petta, S.; Mangia, A.; Berg, T.; Boonstra, A.; Brouwer, W.P.; Abate, M.L.; Wong, V.W.; Nazmy, M.; et al. The membrane-bound o-acyltransferase domain-containing 7 variant rs641738 increases inflammation and fibrosis in chronic hepatitis b. Hepatology 2017, 65, 1840–1850. [Google Scholar] [CrossRef]
- Viitasalo, A.; Eloranta, A.M.; Atalay, M.; Romeo, S.; Pihlajamäki, J.; Lakka, T.A. Association of mboat7 gene variant with plasma alt levels in children: The panic study. Pediatr. Res. 2016, 80, 651–655. [Google Scholar] [CrossRef]
- Lee, H.C.; Inoue, T.; Imae, R.; Kono, N.; Shirae, S.; Matsuda, S.; Gengyo-Ando, K.; Mitani, S.; Arai, H. Caenorhabditis elegans mboa-7, a member of the mboat family, is required for selective incorporation of polyunsaturated fatty acids into phosphatidylinositol. Mol. Biol. Cell 2008, 19, 1174–1184. [Google Scholar] [CrossRef]
- Thangapandi, V.R.; Knittelfelder, O.; Brosch, M.; Patsenker, E.; Vvedenskaya, O.; Buch, S.; Hinz, S.; Hendricks, A.; Nati, M.; Herrmann, A.; et al. Loss of hepatic mboat7 leads to liver fibrosis. Gut 2021, 70, 940–950. [Google Scholar] [CrossRef]
- Wang, P.; Wu, C.X.; Li, Y.; Shen, N. Hsd17b13 rs72613567 protects against liver diseases and histological progression of nonalcoholic fatty liver disease: A systematic review and meta-analysis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 8997–9007. [Google Scholar]
- Stender, S.; Smagris, E.; Lauridsen, B.K.; Kofoed, K.F.; Nordestgaard, B.G.; Tybjaerg-Hansen, A.; Pennacchio, L.A.; Dickel, D.E.; Cohen, J.C.; Hobbs, H.H. Relationship between genetic variation at ppp1r3b and levels of liver glycogen and triglyceride. Hepatology 2018, 67, 2182–2195. [Google Scholar] [CrossRef] [PubMed]
- Saremi, L.; Lotfıpanah, S.; Mohammadi, M.; Hosseinzadeh, H.; Hosseini-Khah, Z.; Johari, B.; Saltanatpour, Z. Association between ppargc1a single nucleotide polymorphisms and increased risk of nonalcoholic fatty liver disease among iranian patients with type 2 diabetes mellitus. Turk. J. Med. Sci. 2019, 49, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Chang, P.F.; Chang, M.H.; Ni, Y.H. A common variant in the peroxisome proliferator-activated receptor-γ coactivator-1α gene is associated with nonalcoholic fatty liver disease in obese children. Am. J. Clin. Nutr. 2013, 97, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.N.; Shen, F.; Pan, Q.; Cao, H.X.; Chen, G.Y.; Fan, J.G. Ppargc1a rs8192678 g>a polymorphism affects the severity of hepatic histological features and nonalcoholic steatohepatitis in patients with nonalcoholic fatty liver disease. World J. Gastroenterol. 2021, 27, 3863–3876. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, P.; Bermano, G.; Williams, H.C.; Knott, R.M. Meta-analysis demonstrates gly482ser variant of ppargc1a is associated with components of metabolic syndrome within asian populations. Genomics 2020, 112, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, M.; Hotta, K.; Nozaki, Y.; Endo, H.; Uchiyama, T.; Mawatari, H.; Iida, H.; Kato, S.; Hosono, K.; Fujita, K.; et al. Association between ppargc1a polymorphisms and the occurrence of nonalcoholic fatty liver disease (nafld). BMC Gastroenterol. 2008, 8, 27. [Google Scholar] [CrossRef]
- Chen, L.; Lin, Z.; Jiang, M.; Lu, L.; Zhang, H.; Xin, Y.; Jiang, X.; Xuan, S. Genetic variants in the samm50 gene create susceptibility to nonalcoholic fatty liver disease in a chinese han population. Hepat. Mon. 2015, 15, e31076. [Google Scholar] [CrossRef]
- Li, Z.; Shen, W.; Wu, G.; Qin, C.; Zhang, Y.; Wang, Y.; Song, G.; Xiao, C.; Zhang, X.; Deng, G.; et al. The role of samm50 in non-alcoholic fatty liver disease: From genetics to mechanisms. FEBS Open Bio 2021, 11, 1893–1906. [Google Scholar] [CrossRef]
- Chung, G.E.; Lee, Y.; Yim, J.Y.; Choe, E.K.; Kwak, M.S.; Yang, J.I.; Park, B.; Lee, J.E.; Kim, J.A.; Kim, J.S. Genetic polymorphisms of pnpla3 and samm50 are associated with nonalcoholic fatty liver disease in a korean population. Gut Liver 2018, 12, 316–323. [Google Scholar] [CrossRef]
- Zhang, R.N.; Zheng, R.D.; Mi, Y.Q.; Zhou, D.; Shen, F.; Chen, G.Y.; Zhu, C.Y.; Pan, Q.; Fan, J.G. Apoc3 rs2070666 is associated with the hepatic steatosis independently of pnpla3 rs738409 in chinese han patients with nonalcoholic fatty liver diseases. Dig. Dis. Sci. 2016, 61, 2284–2293. [Google Scholar] [CrossRef]
- Valenti, L.; Nobili, V.; Al-Serri, A.; Rametta, R.; Leathart, J.B.; Zappa, M.A.; Dongiovanni, P.; Fracanzani, A.L.; Alterio, A.; Roviaro, G.; et al. The apoc3 t-455c and c-482t promoter region polymorphisms are not associated with the severity of liver damage independently of pnpla3 i148m genotype in patients with nonalcoholic fatty liver. J. Hepatol. 2011, 55, 1409–1414. [Google Scholar] [CrossRef]
- Petersen, K.F.; Dufour, S.; Hariri, A.; Nelson-Williams, C.; Foo, J.N.; Zhang, X.M.; Dziura, J.; Lifton, R.P.; Shulman, G.I. Apolipoprotein c3 gene variants in nonalcoholic fatty liver disease. N. Engl. J. Med. 2010, 362, 1082–1089. [Google Scholar] [CrossRef]
- Jain, V.; Kumar, A.; Ahmad, N.; Jana, M.; Kalaivani, M.; Kumar, B.; Shastri, S.; Jain, O.; Kabra, M. Genetic polymorphisms associated with obesity and non-alcoholic fatty liver disease in asian indian adolescents. J. Pediatr. Endocrinol. Metab. 2019, 32, 749–758. [Google Scholar] [CrossRef]
- Zain, S.M.; Mohamed, Z.; Mahadeva, S.; Rampal, S.; Basu, R.C.; Cheah, P.L.; Salim, A.; Mohamed, R. Susceptibility and gene interaction study of the angiotensin ii type 1 receptor (agtr1) gene polymorphisms with non-alcoholic fatty liver disease in a multi-ethnic population. PLoS ONE 2013, 8, e58538. [Google Scholar] [CrossRef]
- Yoneda, M.; Hotta, K.; Nozaki, Y.; Endo, H.; Uchiyama, T.; Mawatari, H.; Iida, H.; Kato, S.; Fujita, K.; Takahashi, H.; et al. Association between angiotensin ii type 1 receptor polymorphisms and the occurrence of nonalcoholic fatty liver disease. Liver Int. 2009, 29, 1078–1085. [Google Scholar] [CrossRef]
- Valenti, L.; Motta, B.M.; Alisi, A.; Sartorelli, R.; Buonaiuto, G.; Dongiovanni, P.; Rametta, R.; Pelusi, S.; Fargion, S.; Nobili, V. Lpin1 rs13412852 polymorphism in pediatric nonalcoholic fatty liver disease. J. Pediatr. Gastroenterol. Nutr. 2012, 54, 588–593. [Google Scholar] [CrossRef]
- Zhou, Y.-J.; Li, Y.-Y.; Nie, Y.-Q.; Yang, H.; Zhan, Q.; Huang, J.; Shi, S.-L.; Lai, X.-B.; Huang, H.-L. Influence of polygenetic polymorphisms on the susceptibility to non-alcoholic fatty liver disease of chinese people. J. Gastroenterol. Hepatol. 2010, 25, 772–777. [Google Scholar] [CrossRef]
- Dong, H.; Wang, J.; Li, C.; Hirose, A.; Nozaki, Y.; Takahashi, M.; Ono, M.; Akisawa, N.; Iwasaki, S.; Saibara, T.; et al. The phosphatidylethanolamine n-methyltransferase gene v175m single nucleotide polymorphism confers the susceptibility to nash in japanese population. J. Hepatol. 2007, 46, 915–920. [Google Scholar] [CrossRef]
- Song, J.; Da Costa, K.A.; Fischer, L.M.; Kohlmeier, M.; Kwock, L.; Wang, S.; Zeisel, S.H. Polymorphism of the pemt gene and susceptibility to nonalcoholic fatty liver disease (nafld). FASEB J. 2005, 19, 1266–1271. [Google Scholar] [CrossRef]
- Bale, G.; Vishnubhotla, R.V.; Mitnala, S.; Sharma, M.; Padaki, R.N.; Pawar, S.C.; Duvvur, R.N. Whole-exome sequencing identifies a variant in phosphatidylethanolamine n-methyltransferase gene to be associated with lean-nonalcoholic fatty liver disease. J. Clin. Exp. Hepatol. 2019, 9, 561–568. [Google Scholar] [CrossRef]
- Jun, D.W.; Han, J.H.; Jang, E.C.; Kim, S.H.; Kim, S.H.; Jo, Y.J.; Park, Y.S.; Chae, J.D. Polymorphisms of microsomal triglyceride transfer protein gene and phosphatidylethanolamine n-methyltransferase gene in alcoholic and nonalcoholic fatty liver disease in koreans. Eur. J. Gastroenterol. Hepatol. 2009, 21, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Namikawa, C.; Shu-Ping, Z.; Vyselaar, J.R.; Nozaki, Y.; Nemoto, Y.; Ono, M.; Akisawa, N.; Saibara, T.; Hiroi, M.; Enzan, H.; et al. Polymorphisms of microsomal triglyceride transfer protein gene and manganese superoxide dismutase gene in non-alcoholic steatohepatitis. J. Hepatol. 2004, 40, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Gambino, R.; Cassader, M.; Pagano, G.; Durazzo, M.; Musso, G. Polymorphism in microsomal triglyceride transfer protein: A link between liver disease and atherogenic postprandial lipid profile in nash? Hepatology 2007, 45, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Carulli, L.; Canedi, I.; Rondinella, S.; Lombardini, S.; Ganazzi, D.; Fargion, S.; De Palma, M.; Lonardo, A.; Ricchi, M.; Bertolotti, M.; et al. Genetic polymorphisms in non-alcoholic fatty liver disease: Interleukin-6-174g/c polymorphism is associated with non-alcoholic steatohepatitis. Dig. Liver Dis. 2009, 41, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.P.; Stefano, J.T.; Cavaleiro, A.M.; Zanella Fortes, M.A.; Vieira, S.M.; Rodrigues Lima, V.M.; Santos, T.E.; Santos, V.N.; De Azevedo Salgado, A.L.; Parise, E.R.; et al. Association of polymorphisms of glutamate-cystein ligase and microsomal triglyceride transfer protein genes in non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2010, 25, 357–361. [Google Scholar] [CrossRef]
- Al-Serri, A.; Anstee, Q.M.; Valenti, L.; Nobili, V.; Leathart, J.B.; Dongiovanni, P.; Patch, J.; Fracanzani, A.; Fargion, S.; Day, C.P.; et al. The sod2 c47t polymorphism influences nafld fibrosis severity: Evidence from case-control and intra-familial allele association studies. J. Hepatol. 2012, 56, 448–454. [Google Scholar] [CrossRef]
- Abshagen, K.; Berger, C.; Dietrich, A.; Schütz, T.; Wittekind, C.; Stumvoll, M.; Blüher, M.; Klöting, N. A human repin1 gene variant: Genetic risk factor for the development of nonalcoholic fatty liver disease. Clin. Transl. Gastroenterol 2020, 11, e00114. [Google Scholar] [CrossRef]
- Aller, R.; De Luis, D.A.; Izaola, O.; González Sagrado, M.; Conde, R.; Alvarez, T.; Pacheco, D.; Velasco, M.C. Role of -55ct polymorphism of ucp3 gene on non alcoholic fatty liver disease and insulin resistance in patients with obesity. Nutr. Hosp. 2010, 25, 572–576. [Google Scholar]
- Borén, J.; Packard, C.J.; Taskinen, M.-R. The roles of apoc-iii on the metabolism of triglyceride-rich lipoproteins in humans. Front. Endocrinol. 2020, 11, 474. [Google Scholar] [CrossRef]
- Hooper, A.J.; Burnett, J.R.; Watts, G.F. Contemporary aspects of the biology and therapeutic regulation of the microsomal triglyceride transfer protein. Circ. Res. 2015, 116, 193–205. [Google Scholar] [CrossRef]
- Zain, S.M.; Mohamed, R.; Cooper, D.N.; Razali, R.; Rampal, S.; Mahadeva, S.; Chan, W.K.; Anwar, A.; Rosli, N.S.; Mahfudz, A.S.; et al. Genome-wide analysis of copy number variation identifies candidate gene loci associated with the progression of non-alcoholic fatty liver disease. PLoS ONE 2014, 9, e95604. [Google Scholar] [CrossRef]
- Zain, S.M.; Mohamed, Z.; Pirmohamed, M.; Tan, H.L.; Alshawsh, M.A.; Mahadeva, S.; Chan, W.K.; Mustapha, N.R.; Mohamed, R. Copy number variation in exportin-4 (xpo4) gene and its association with histological severity of non-alcoholic fatty liver disease. Sci. Rep. 2015, 5, 13306. [Google Scholar] [CrossRef]
- Metwally, M.; Bayoumi, A.; Khan, A.; Adams, L.A.; Aller, R.; García-Monzón, C.; Arias-Loste, M.T.; Bugianesi, E.; Miele, L.; Alisi, A.; et al. Copy number variation and expression of exportin-4 associates with severity of fibrosis in metabolic associated fatty liver disease. eBioMedicine 2021, 70, 103521. [Google Scholar] [CrossRef]
- Li, Y.F.; Zheng, J.; Peng, H.W.; Cai, X.L.; Pan, X.T.; Li, H.Q.; Hong, Q.Z.; Hu, Z.J.; Wu, Y.L.; Peng, X.-E. Identifying potential biomarkers of nonalcoholic fatty liver disease via genome-wide analysis of copy number variation. BMC Gastroenterol. 2021, 21, 171. [Google Scholar] [CrossRef]
- Chen, B.B.; Yan, J.H.; Zheng, J.; Peng, H.W.; Cai, X.L.; Pan, X.T.; Li, H.Q.; Hong, Q.Z.; Peng, X.-E. Copy number variation in the ces1 gene and the risk of non-alcoholic fatty liver in a chinese han population. Sci. Rep. 2021, 11, 13984. [Google Scholar] [CrossRef]
- Dongiovanni, P.; Stender, S.; Pietrelli, A.; Mancina, R.M.; Cespiati, A.; Petta, S.; Pelusi, S.; Pingitore, P.; Badiali, S.; Maggioni, M.; et al. Causal relationship of hepatic fat with liver damage and insulin resistance in nonalcoholic fatty liver. J. Intern. Med. 2018, 283, 356–370. [Google Scholar] [CrossRef]
- De Vincentis, A.; Tavaglione, F.; Jamialahmadi, O.; Picardi, A.; Antonelli Incalzi, R.; Valenti, L.; Romeo, S.; Vespasiani-Gentilucci, U. A polygenic risk score to refine risk stratification and prediction for severe liver disease by clinical fibrosis scores. Clin. Gastroenterol. Hepatol. 2022, 20, 658–673. [Google Scholar] [CrossRef]
- Di Costanzo, A.; Pacifico, L.; Chiesa, C.; Perla, F.M.; Ceci, F.; Angeloni, A.; D’Erasmo, L.; Di Martino, M.; Arca, M. Genetic and metabolic predictors of hepatic fat content in a cohort of italian children with obesity. Pediatr. Res. 2019, 85, 671–677. [Google Scholar] [CrossRef]
- Thomas, C.E.; Diergaarde, B.; Kuipers, A.L.; Adibi, J.J.; Luu, H.N.; Chang, X.; Dorajoo, R.; Heng, C.K.; Khor, C.C.; Wang, R.; et al. Nafld polygenic risk score and risk of hepatocellular carcinoma in an east asian population. Hepatol. Commun. 2022, 6, 2310–2321. [Google Scholar] [CrossRef]
- Donati, B.; Dongiovanni, P.; Romeo, S.; Meroni, M.; McCain, M.; Miele, L.; Petta, S.; Maier, S.; Rosso, C.; De Luca, L.; et al. Mboat7 rs641738 variant and hepatocellular carcinoma in non-cirrhotic individuals. Sci. Rep. 2017, 7, 4492. [Google Scholar] [CrossRef]
- Wang, Y.; Kory, N.; BasuRay, S.; Cohen, J.C.; Hobbs, H.H. Pnpla3, cgi-58, and inhibition of hepatic triglyceride hydrolysis in mice. Hepatology 2019, 69, 2427–2441. [Google Scholar] [CrossRef]
- Gellert-Kristensen, H.; Richardson, T.G.; Davey Smith, G.; Nordestgaard, B.G.; Tybjaerg-Hansen, A.; Stender, S. Combined effect of pnpla3, tm6sf2, and hsd17b13 variants on risk of cirrhosis and hepatocellular carcinoma in the general population. Hepatology 2020, 72, 845–856. [Google Scholar] [CrossRef]
- Bianco, C.; Jamialahmadi, O.; Pelusi, S.; Baselli, G.; Dongiovanni, P.; Zanoni, I.; Santoro, L.; Maier, S.; Liguori, A.; Meroni, M.; et al. Non-invasive stratification of hepatocellular carcinoma risk in non-alcoholic fatty liver using polygenic risk scores. J. Hepatol. 2021, 74, 775–782. [Google Scholar] [CrossRef]
- Chen, X.Y.; Wang, C.; Huang, Y.Z.; Zhang, L.L. Nonalcoholic fatty liver disease shows significant sex dimorphism. World J. Clin. Cases 2022, 10, 1457–1472. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M. Non-alcoholic fatty liver disease—A global public health perspective. J. Hepatol. 2019, 70, 531–544. [Google Scholar] [CrossRef] [PubMed]
- Torres, D.M.; Williams, C.D.; Harrison, S.A. Features, diagnosis, and treatment of nonalcoholic fatty liver disease. Clin. Gastroenterol. Hepatol. 2012, 10, 837–858. [Google Scholar] [CrossRef] [PubMed]
- Bjune, J.I.; Strømland, P.P.; Jersin, R.; Mellgren, G.; Dankel, S.N. Metabolic and epigenetic regulation by estrogen in adipocytes. Front. Endocrinol. 2022, 13, 828780. [Google Scholar] [CrossRef] [PubMed]
- Ismaiel, A.; Dumitrascu, D.L. Genetic predisposition in metabolic-dysfunction-associated fatty liver disease and cardiovascular outcomes-systematic review. Eur. J. Clin. Investig. 2020, 50, e13331. [Google Scholar] [CrossRef]
- Wang, J.; Ye, C.; Fei, S. Association between apoc3 polymorphisms and non-alcoholic fatty liver disease risk: A meta-analysis. Afr. Health Sci. 2020, 20, 1800–1808. [Google Scholar] [CrossRef]
- Bamshad, M.; Kivisild, T.; Watkins, W.S.; Dixon, M.E.; Ricker, C.E.; Rao, B.B.; Naidu, J.M.; Prasad, B.V.; Reddy, P.G.; Rasanayagam, A.; et al. Genetic evidence on the origins of indian caste populations. Genome Res. 2001, 11, 994–1004. [Google Scholar] [CrossRef]
- Hernaez, R.; Lazo, M.; Bonekamp, S.; Kamel, I.; Brancati, F.L.; Guallar, E.; Clark, J.M. Diagnostic accuracy and reliability of ultrasonography for the detection of fatty liver: A meta-analysis. Hepatology 2011, 54, 1082–1090. [Google Scholar] [CrossRef]
- Menon, D.K.; Rosand, J. Finding a place for candidate gene studies in a genome-wide association study world. JAMA Netw. Open 2021, 4, e2118594. [Google Scholar] [CrossRef]
- Wong, V.W.; Chan, W.K.; Chitturi, S.; Chawla, Y.; Dan, Y.Y.; Duseja, A.; Fan, J.; Goh, K.L.; Hamaguchi, M.; Hashimoto, E.; et al. Asia-pacific working party on non-alcoholic fatty liver disease guidelines 2017-part 1: Definition, risk factors and assessment. J. Gastroenterol. Hepatol. 2018, 33, 70–85. [Google Scholar] [CrossRef]
- Li, J.; Zou, B.; Yeo, Y.H.; Feng, Y.; Xie, X.; Lee, D.H.; Fujii, H.; Wu, Y.; Kam, L.Y.; Ji, F.; et al. Prevalence, incidence, and outcome of non-alcoholic fatty liver disease in asia, 1999–2019: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2019, 4, 389–398. [Google Scholar] [CrossRef]
- Fleischman, M.W.; Budoff, M.; Zeb, I.; Li, D.; Foster, T. Nafld prevalence differs among hispanic subgroups: The multi-ethnic study of atherosclerosis. World J. Gastroenterol. 2014, 20, 4987–4993. [Google Scholar] [CrossRef]
- Kumar, A.; Walia, G.K.; Gupta, V.; Sachdeva, M.P. Genetics of nonalcoholic fatty liver disease in asian populations. J. Genet 2019, 98, 29. [Google Scholar] [CrossRef]
- Turner, R.M.; Bird, S.M.; Higgins, J.P. The impact of study size on meta-analyses: Examination of underpowered studies in cochrane reviews. PLoS ONE 2013, 8, e59202. [Google Scholar] [CrossRef]
- Robinson, M.R.; Wray, N.R.; Visscher, P.M. Explaining additional genetic variation in complex traits. Trends Genet. 2014, 30, 124–132. [Google Scholar] [CrossRef]
- Flannick, J.; Fuchsberger, C.; Mahajan, A.; Teslovich, T.M.; Agarwala, V.; Gaulton, K.J.; Caulkins, L.; Koesterer, R.; Ma, C.; Moutsianas, L.; et al. Sequence data and association statistics from 12,940 type 2 diabetes cases and controls. Sci. Data 2017, 4, 170179. [Google Scholar] [CrossRef]
- Gershon, E.S.; Pearlson, G.; Keshavan, M.S.; Tamminga, C.; Clementz, B.; Buckley, P.F.; Alliey-Rodriguez, N.; Liu, C.; Sweeney, J.A.; Keedy, S.; et al. Genetic analysis of deep phenotyping projects in common disorders. Schizophr. Res. 2018, 195, 51–57. [Google Scholar] [CrossRef]

| Gene | SNP | Population Study | SNP Effects | Reference(s) |
|---|---|---|---|---|
| PNPLA3 | rs738409 C > G (I148M) | Caucasians, African, Japanese, Latinos, Native Hawaiians, and European Americans, Amish, Family Heart Study, Framingham Heart Study, Finnish, Chinese, Taiwanese | Associated with NAFLD risk and steatosis | [34,35,36,37,38,39,40,41,42,43,44,45] |
| German, Japanese | Associated with NAFLD risk, steatosis, and fibrosis | [46,47,48] | ||
| Italian | Associated with NAFLD risk, steatosis, fibrosis, and cirrhosis | [49,50,51,52] | ||
| Belgians | Associated with NAFLD and NASH risk | [53] | ||
| Brazilian, Malaysians (Malays, Chinese, and Indians) | Associated with NAFLD and NASH risk, but not steatosis | [54,55] | ||
| Turkish | Associated with NAFLD and NASH risk, and fibrosis | [56,57] | ||
| Chinese | Associated with NAFLD risk but not steatosis | [58] | ||
| Chinese | Associated with NAFLD risk in lean individuals | [59] | ||
| Japanese | Associated with NAFLD risk and fibrosis, but not steatosis | [60,61] | ||
| Korean | Associated with NAFLD risk and fibrosis | [62,63] | ||
| Indian, Sri Lankans, Bangladeshi, Iranian, Singaporean (Chinese) | Associated with NAFLD risk | [64,65,66,67,68,69,70,71] | ||
| Indian | Associated with NAFLD and NASH risk, and steatosis | [72] | ||
| Filipinos | No association with NAFLD risk | [73] | ||
| rs2896019 T > G | Japanese | Associated with NAFLD risk | [74] | |
| rs6006460 G > T (S453I) | African American | Associated with lower hepatic fat content | [34] | |
| TM6SF2 | rs58542926 G > A (E167K) | Caucasian, African, Hispanic, and European Americans, Swedish, European, Finnish, Polish, and Italian | Associated with NAFLD risk and steatosis | [49,52,75,76,77,78,79,80,81,82] |
| German | Associated with NAFLD risk and steatosis but not fibrosis | [47] | ||
| Caucasians, African, and Hispanic Americans, Chinese | Associated with NAFLD risk, steatosis, and fibrosis | [43,83,84,85,86,87] | ||
| Italian, Finnish, and Swedish | Associated with NASH progression | [88] | ||
| Korean | Associated with NASH and fibrosis | [89] | ||
| Indian, Argentinian | Associated with NAFLD risk | [66,77,90] | ||
| Brazilian, Japanese, Singaporean (Chinese) | No association with NAFLD | [54,71,91] | ||
| GCKR | rs780094 C > T | Amish, Family Heart Study, and Framingham Heart Study, Caucasian, African, and Mexican Americans | Associated with NAFLD risk and steatosis | [39,92] |
| German | Associated with NAFLD and fibrosis severity | [46] | ||
| Italian | Associated with liver fibrosis severity | [93] | ||
| Taiwanese, Uygur | Associated with NAFLD risk | [94,95] | ||
| Malaysians (Malays, Chinese, and Indians) | Associated with NAFLD, NASH, and fibrosis | [96] | ||
| Iranian, Chinese, Singaporean (Chinese) | No association with NAFLD | [71,97,98] | ||
| rs1260326 C > T (P446L) | Italian, Japanese, Uygur, and Chinese | Associated with NAFLD risk | [49,74,95,99,100] | |
| Finnish | Associated with steatosis | [101] | ||
| Malaysians (Malays, Chinese, and Indians) | Associated with NAFLD, NASH, and fibrosis | [96] | ||
| Chinese | No association with NAFLD | [98,102] | ||
| MBOAT7 | rs641738 C > T | Dallas Heart Study and Europeans, Italian | Associated with NAFLD risk and steatosis | [49,52,103] |
| Finnish | Associated with NASH and fibrosis | [104] | ||
| German | Associated with fibrosis only | [47] | ||
| Eastern European | No association with fibrosis or cirrhosis | [105] | ||
| Americans, Italian, Argentinian, Chinese, Taiwanese, and Korean | No association with NAFLD risk | [37,89,102,106,107,108] | ||
| HSD17B13 | rs72613567 T > TA | Caucasian Americans | Associated with steatosis and lower risk of inflammation and NASH | [109,110] |
| European, Finnish | Associated with lower NASH and fibrosis risk | [111,112] | ||
| Argentinian, Japanese | Associated with lower NASH risk | [113,114] | ||
| Korean | Associated with lower NAFLD risk | [115] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulaiman, S.A.; Dorairaj, V.; Adrus, M.N.H. Genetic Polymorphisms and Diversity in Nonalcoholic Fatty Liver Disease (NAFLD): A Mini Review. Biomedicines 2023, 11, 106. https://doi.org/10.3390/biomedicines11010106
Sulaiman SA, Dorairaj V, Adrus MNH. Genetic Polymorphisms and Diversity in Nonalcoholic Fatty Liver Disease (NAFLD): A Mini Review. Biomedicines. 2023; 11(1):106. https://doi.org/10.3390/biomedicines11010106
Chicago/Turabian StyleSulaiman, Siti Aishah, Vicneswarry Dorairaj, and Muhammad Nafiz Haidi Adrus. 2023. "Genetic Polymorphisms and Diversity in Nonalcoholic Fatty Liver Disease (NAFLD): A Mini Review" Biomedicines 11, no. 1: 106. https://doi.org/10.3390/biomedicines11010106
APA StyleSulaiman, S. A., Dorairaj, V., & Adrus, M. N. H. (2023). Genetic Polymorphisms and Diversity in Nonalcoholic Fatty Liver Disease (NAFLD): A Mini Review. Biomedicines, 11(1), 106. https://doi.org/10.3390/biomedicines11010106