
Characterization of Mesothelin Glycosylation in Pancreatic Cancer: Decreased Core Fucosylated Glycoforms in Pancreatic Cancer Patients’ Sera

 , , , , , , and
, , , , , , and 
Abstract
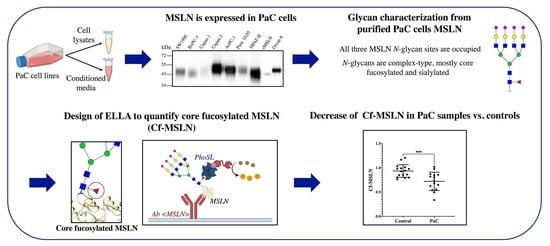
1. Introduction
2. Materials and Methods
2.1. Samples
2.1.1. Cell Lines
2.1.2. Human Samples
2.2. In Solution Mesothelin Glycosidase Digestion
2.3. Western Blotting (WB)
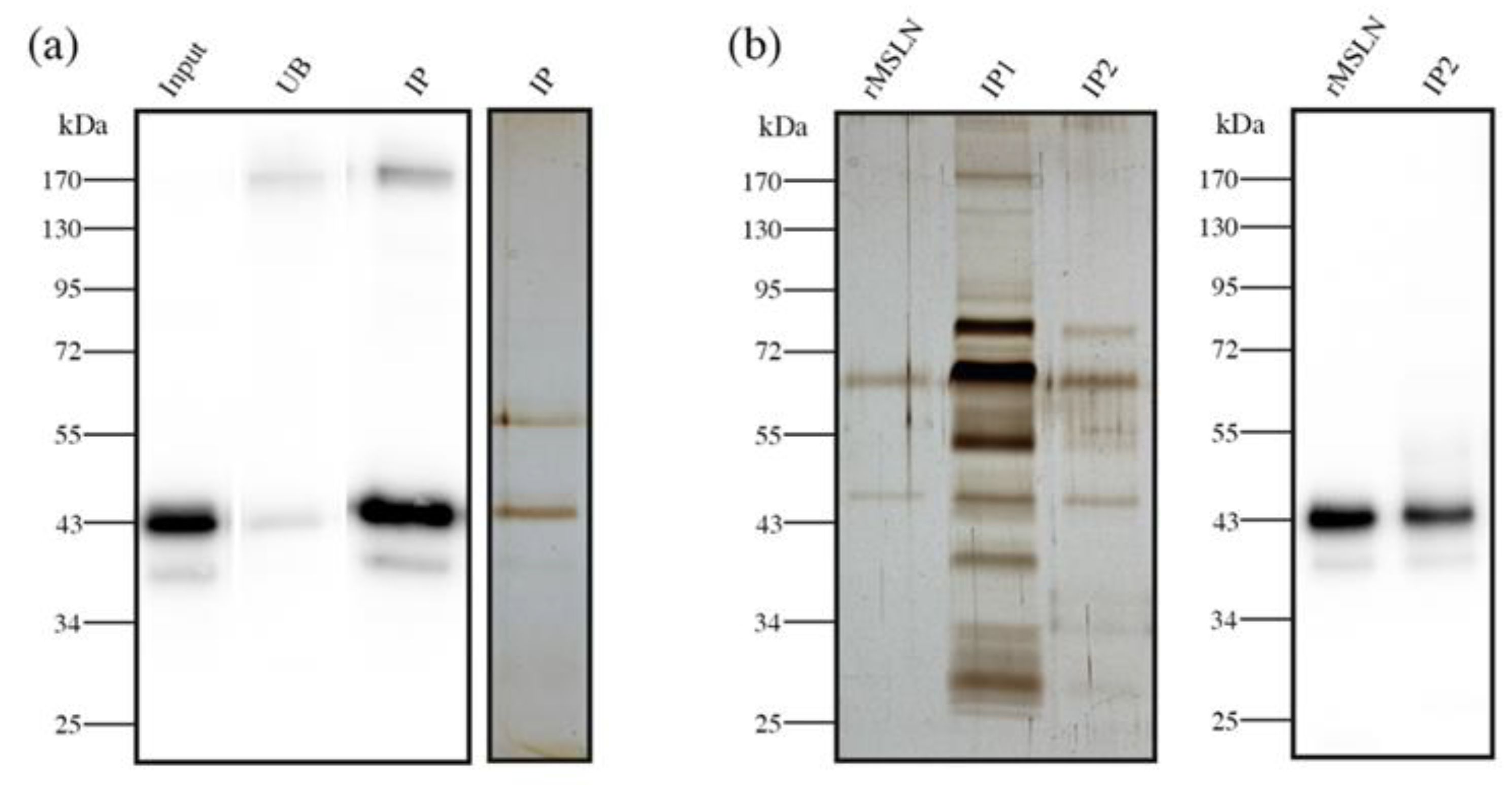
2.4. Mesothelin Immunopurification
2.5. Silver Staining
2.6. N-Glycan Sequencing of MSLN Glycans
2.7. UPLC-ESI-QTof Analyses
2.7.1. Intact Protein Analysis
2.7.2. Peptide Analysis
2.8. Enzyme-Linked Immunosorbent Assay (ELISA) to Quantify Mesothelin
2.9. Enzyme-Linked Lectin Assay (ELLA) to Quantify Core Fucosylated Mesothelin
2.10. Statistics
3. Results
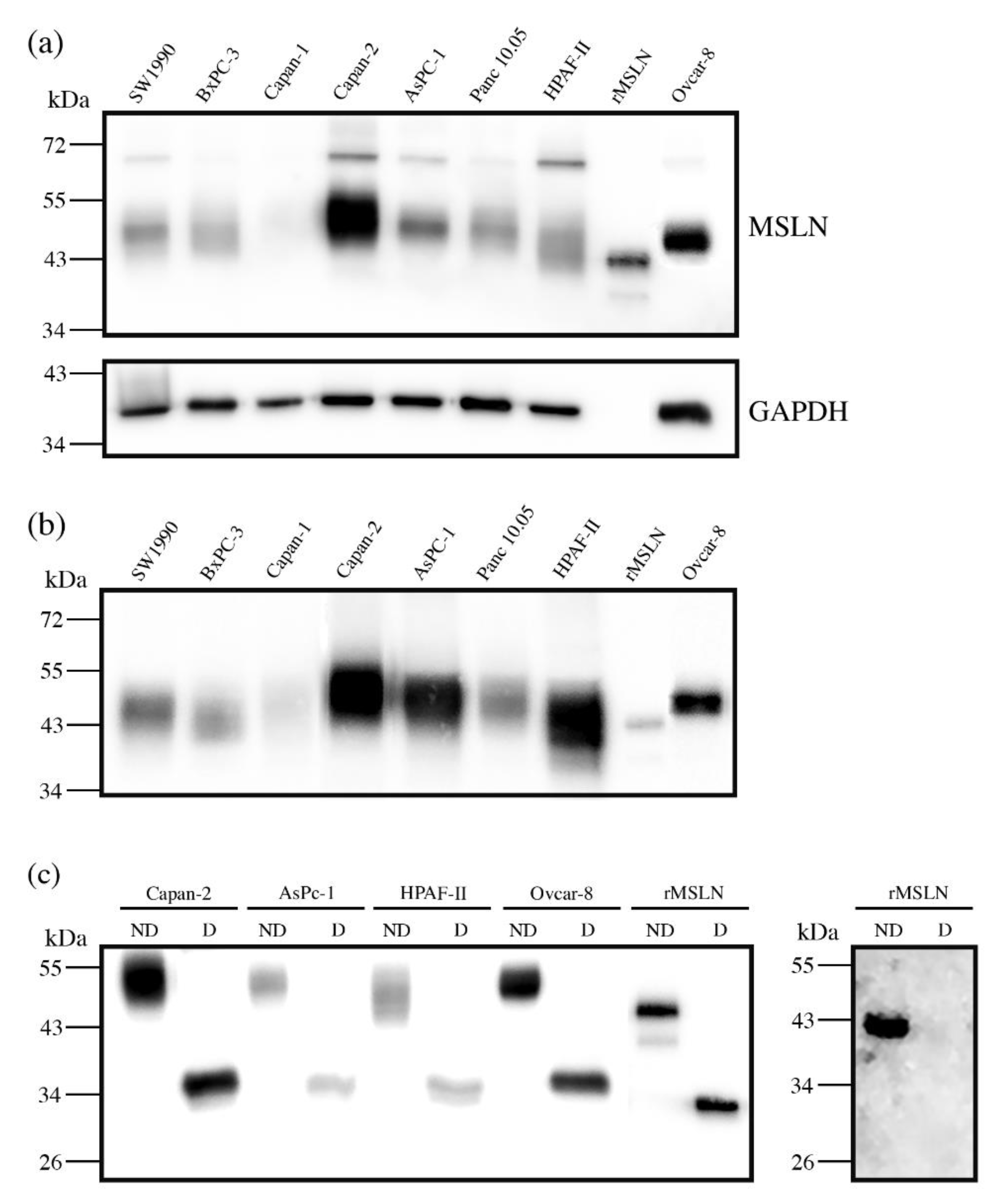
3.1. Mesothelin Expression in Cell Lines
3.2. Mesothelin N-Glycosylation: Site Occupancy
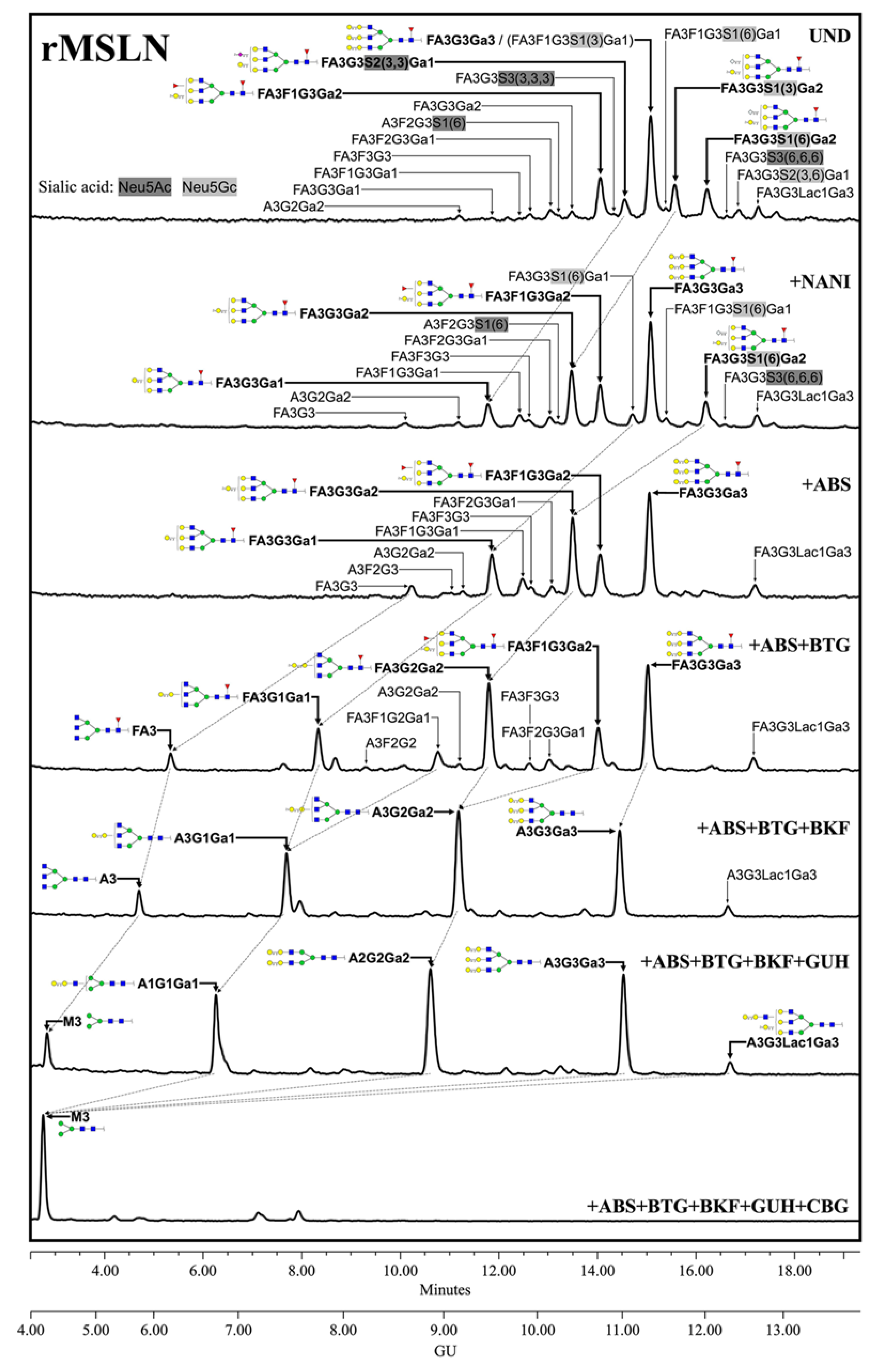
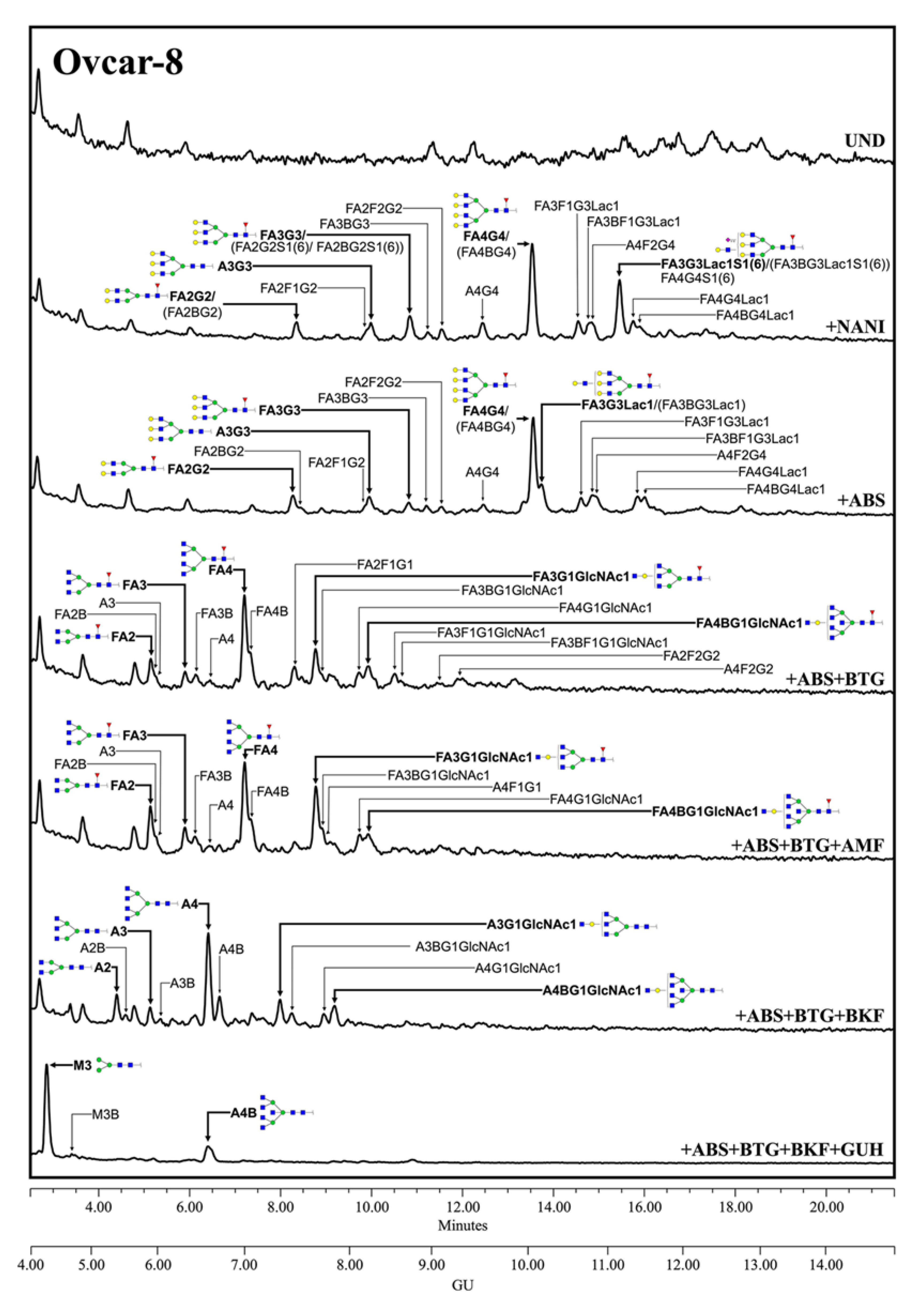
3.3. Characterization of Mesothelin N-Glycans
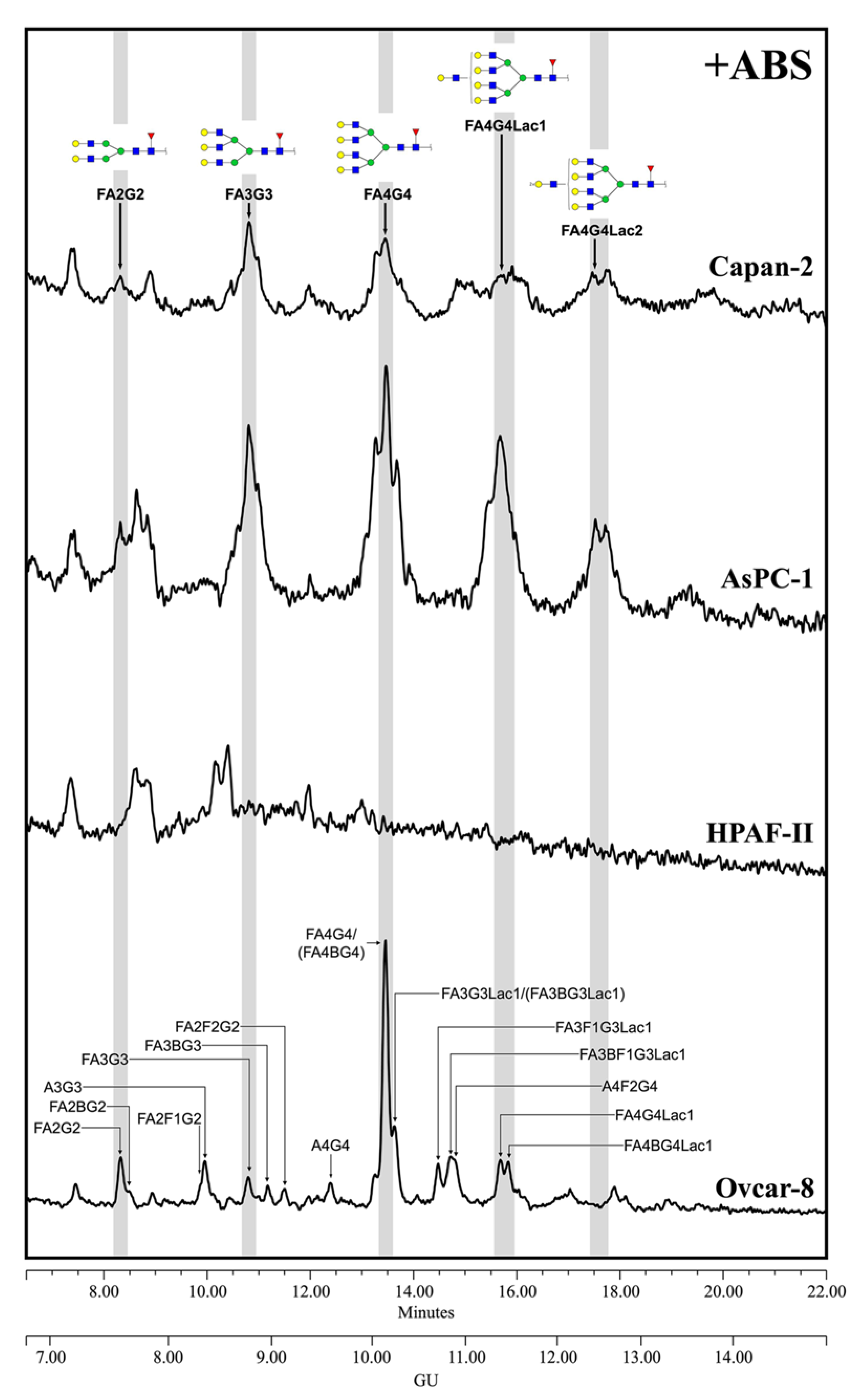
3.3.1. N-Glycan Sequencing by HILIC-UPLC
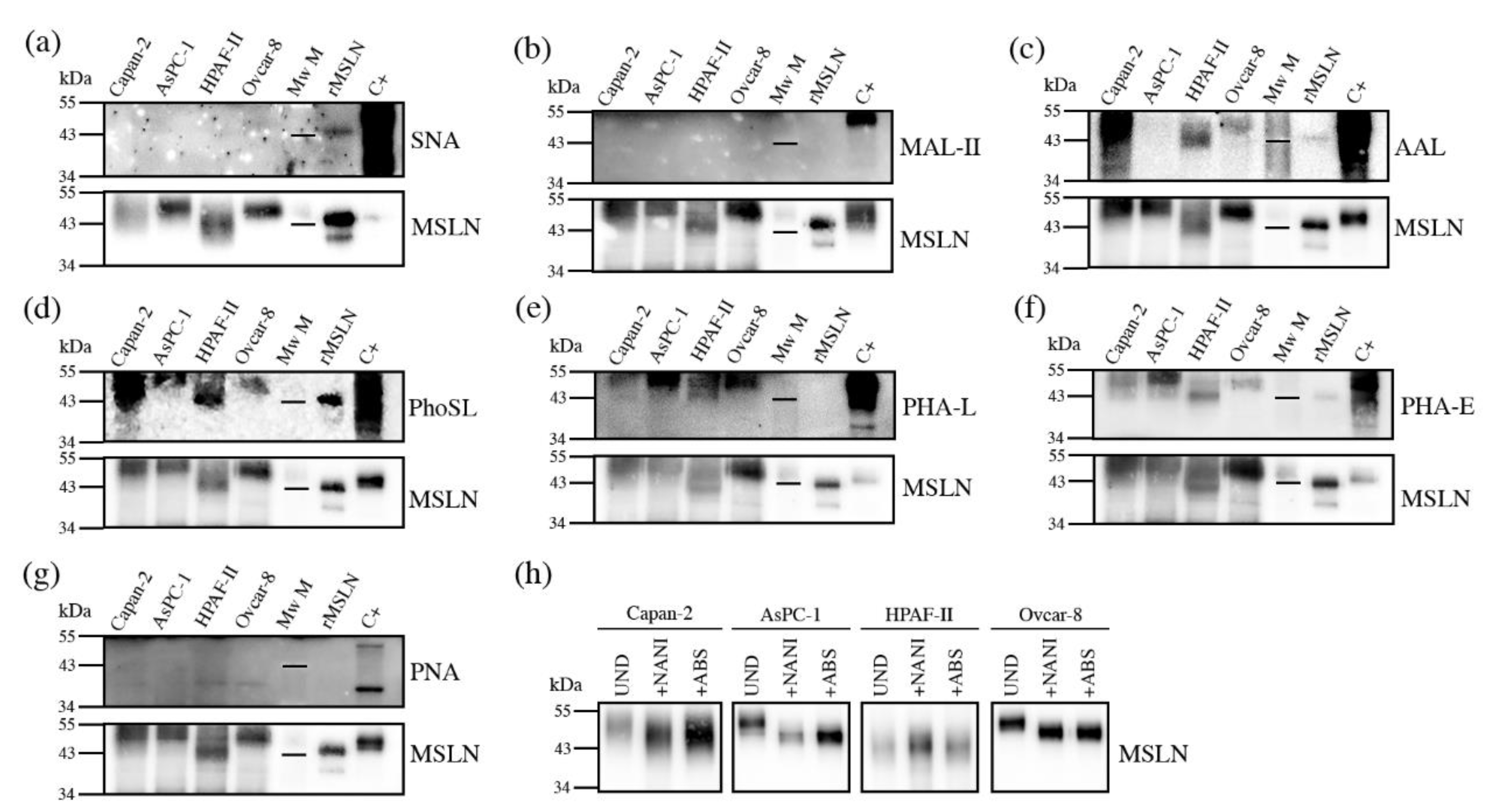
3.3.2. Glycan Determinants’ Expression by WB with Lectins and Sialidase Digestion
3.4. Mesothelin Expression in Pancreatic Tissues
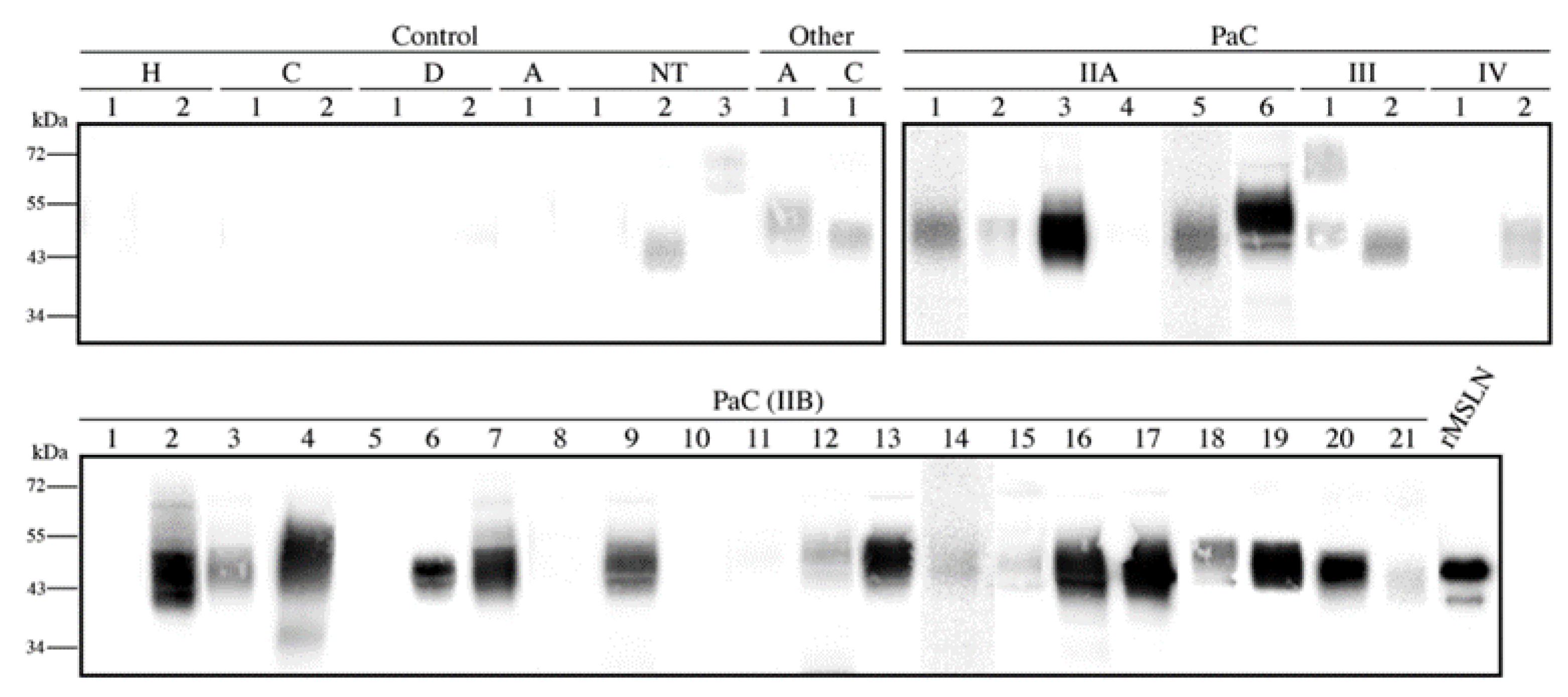
3.5. Core Fucosylated Mesothelin Abundance in Pancreatic Tissues
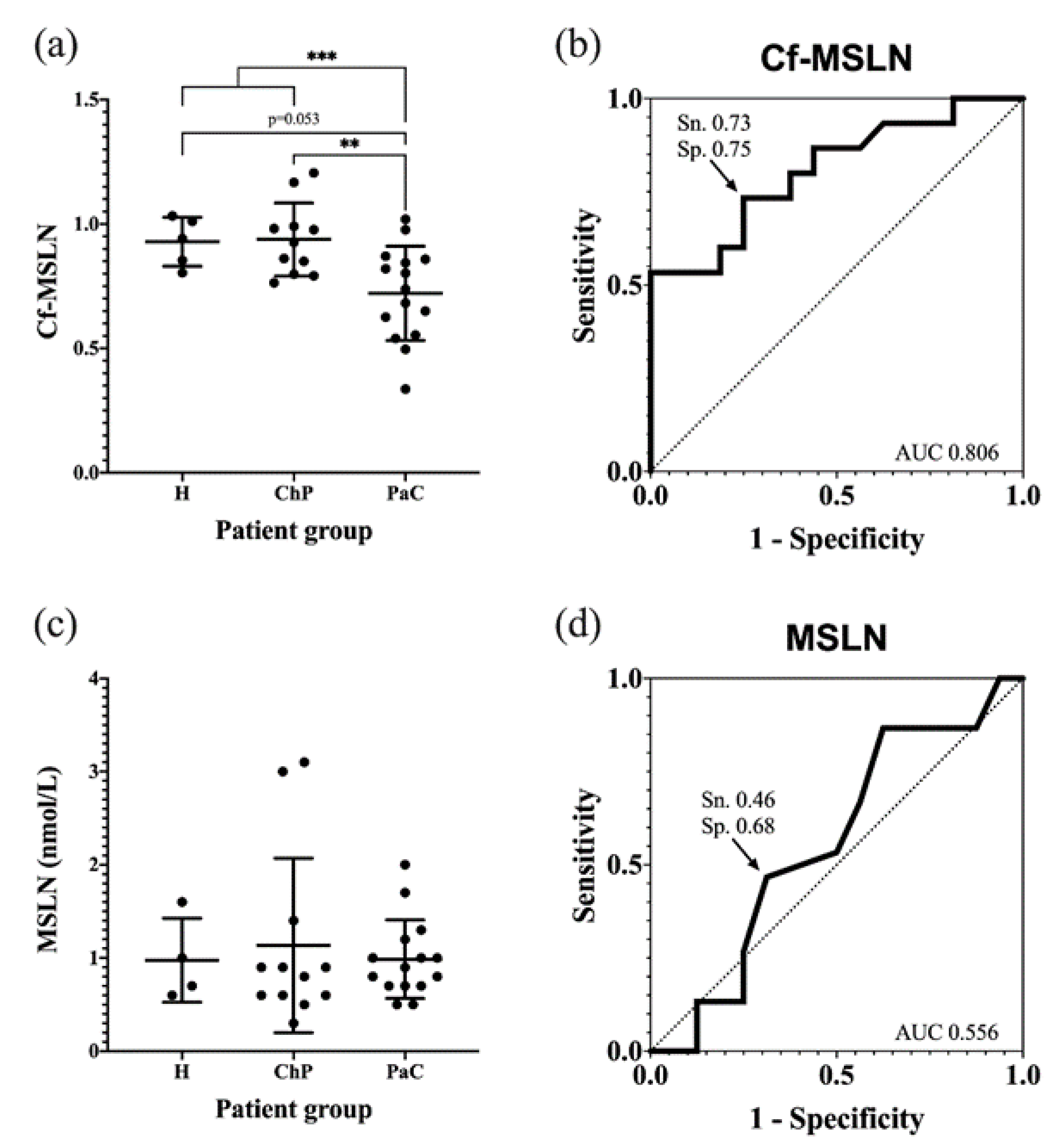
3.6. Serum Core Fucosylated Mesothelin as PaC Biomarker
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Upadhyay, G.; Srivastava, R.K.; Shankar, S. Recent Advances in Pancreatic Cancer: Biology, Treatment, and Prevention. Biochim. Biophys. Acta 2015, 1856, 13–27. [Google Scholar] [CrossRef]
- Roth, M.T.; Cardin, D.B.; Berlin, J.D. Recent Advances in the Treatment of Pancreatic Cancer. F1000Research 2020, 9, 131. [Google Scholar] [CrossRef] [PubMed]
- Kirwan, A.; Utratna, M.; O’Dwyer, M.E.; Joshi, L.; Kilcoyne, M. Glycosylation-Based Serum Biomarkers for Cancer Diagnostics and Prognostics. BioMed Res. Int. 2015, 2015, 490531. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, J.; Li, H.; Wu, Y.; Zhang, H.; Chen, W. Tumor Markers CA19-9, CA242 and CEA in the Diagnosis of Pancreatic Cancer: A Meta-Analysis. Int. J. Clin. Exp. Med. 2015, 8, 11683–11691. [Google Scholar]
- Ferri, M.J.; Saez, M.; Figueras, J.; Fort, E.; Sabat, M.; López-Ben, S.; de Llorens, R.; Aleixandre, R.N.; Peracaula, R. Improved Pancreatic Adenocarcinoma Diagnosis in Jaundiced and Non-Jaundiced Pancreatic Adenocarcinoma Patients through the Combination of Routine Clinical Markers Associated to Pancreatic Adenocarcinoma Pathophysiology. PLoS ONE 2016, 11, e0147214. [Google Scholar] [CrossRef] [PubMed]
- Molina, V.; Visa, L.; Conill, C.; Navarro, S.; Escudero, J.M.; Auge, J.M.; Filella, X.; Lopez-Boado, M.A.; Ferrer, J.; Fernandez-Cruz, L.; et al. CA 19-9 in Pancreatic Cancer: Retrospective Evaluation of Patients with Suspicion of Pancreatic Cancer. Tumor Biol. 2012, 33, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Tsen, A.; Barbara, M.; Rosenkranz, L. Dilemma of Elevated CA 19-9 in Biliary Pathology. Pancreatology 2018, 18, 862–867. [Google Scholar] [CrossRef]
- Ballehaninna, U.K.; Chamberlain, R.S. The Clinical Utility of Serum CA 19-9 in the Diagnosis, Prognosis and Management of Pancreatic Adenocarcinoma: An Evidence Based Appraisal. J. Gastrointest. Oncol. 2012, 3, 15. [Google Scholar] [CrossRef]
- Chan, A.; Diamandis, E.P.; Blasutig, I.M. Strategies for Discovering Novel Pancreatic Cancer Biomarkers. J. Proteom. 2013, 81, 126–134. [Google Scholar] [CrossRef]
- Pereira, S.P.; Oldfield, L.; Ney, A.; Hart, P.A.; Keane, M.G.; Pandol, S.J.; Li, D.; Greenhalf, W.; Jeon, C.Y.; Koay, E.J.; et al. Early Detection of Pancreatic Cancer. Lancet Gastroenterol. Hepatol. 2020, 5, 698–710. [Google Scholar] [CrossRef]
- Peracaula, R.; Barrabés, S.; Sarrats, A.; Rudd, P.M.; Llorens, R.D. Altered Glycosylation in Tumours Focused to Cancer Diagnosis. Dis. Markers 2008, 25, 207–218. [Google Scholar] [CrossRef]
- Munkley, J.; Elliott, D.J. Hallmarks of Glycosylation in Cancer. Oncotarget 2016, 7, 35478–35489. [Google Scholar] [CrossRef]
- Stowell, S.R.; Ju, T.; Cummings, R.D. Protein Glycosylation in Cancer. Annu. Rev. Pathol. 2015, 10, 473–510. [Google Scholar] [CrossRef] [PubMed]
- Pinho, S.S.; Reis, C.A. Glycosylation in Cancer: Mechanisms and Clinical Implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef] [PubMed]
- Munkley, J. The Glycosylation Landscape of Pancreatic Cancer. Oncol. Lett. 2019, 17, 2569–2575. [Google Scholar] [CrossRef]
- Wagatsuma, T.; Nagai-Okatani, C.; Matsuda, A.; Masugi, Y.; Imaoka, M.; Yamazaki, K.; Sakamoto, M.; Kuno, A. Discovery of Pancreatic Ductal Adenocarcinoma-Related Aberrant Glycosylations: A Multilateral Approach of Lectin Microarray-Based Tissue Glycomic Profiling With Public Transcriptomic Datasets. Front. Oncol. 2020, 10, 338. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Garay, M.; Arteta, B.; Pagés, L.; de Llorens, R.; de Bolós, C.; Vidal-Vanaclocha, F.; Peracaula, R. A2,3-Sialyltransferase ST3Gal III Modulates Pancreatic Cancer Cell Motility and Adhesion in Vitro and Enhances Its Metastatic Potential in Vivo. PLoS ONE 2010, 5, e12524. [Google Scholar] [CrossRef]
- Pérez-Garay, M.; Arteta, B.; Llop, E.; Cobler, L.; Pagès, L.; Ortiz, R.; Ferri, M.J.; De Bolós, C.; Figueras, J.; De Llorens, R.; et al. A2,3-Sialyltransferase ST3Gal IV Promotes Migration and Metastasis in Pancreatic Adenocarcinoma Cells and Tends to Be Highly Expressed in Pancreatic Adenocarcinoma Tissues. Int. J. Biochem. Cell Biol. 2013, 45, 1748–1757. [Google Scholar] [CrossRef]
- Schultz, M.J.; Holdbrooks, A.T.; Chakraborty, A.; Grizzle, W.E.; Landen, C.N.; Buchsbaum, D.J.; Conner, M.G.; Arend, R.C.; Yoon, K.J.; Klug, C.A.; et al. The Tumor-Associated Glycosyltransferase ST6Gal-I Regulates Stem Cell Transcription Factors and Confers a Cancer Stem Cell Phenotype. Cancer Res. 2016, 76, 3978–3988. [Google Scholar] [CrossRef]
- Guerrero, P.E.; Miró, L.; Wong, B.S.; Massaguer, A.; Martínez-Bosch, N.; de Llorens, R.; Navarro, P.; Konstantopoulos, K.; Llop, E.; Peracaula, R. Knockdown of A2,3-Sialyltransferases Impairs Pancreatic Cancer Cell Migration, Invasion and E-Selectin-Dependent Adhesion. Int. J. Mol. Sci. 2020, 21, 6239. [Google Scholar] [CrossRef] [PubMed]
- Vreeker, G.C.M.; Hanna-Sawires, R.G.; Mohammed, Y.; Bladergroen, M.R.; Nicolardi, S.; Dotz, V.; Nouta, J.; Bonsing, B.A.; Mesker, W.E.; Burgt, Y.E.M.; et al. Serum N -Glycome Analysis Reveals Pancreatic Cancer Disease Signatures. Cancer Med. 2020, 9, 8519–8529. [Google Scholar] [CrossRef] [PubMed]
- Balmaña, M.; Giménez, E.; Puerta, A.; Llop, E.; Figueras, J. Increased A1-3 Fucosylation of α-1-Acid Glycoprotein (AGP) in Pancreatic Cancer. J. Proteom. 2016, 132, 144–154. [Google Scholar] [CrossRef]
- Barrabés, S.; Pagès-Pons, L.; Radcliffe, C.M.; Tabarés, G.; Fort, E.; Royle, L.; Harvey, D.J.; Moenner, M.; Dwek, R.A.; Rudd, P.M.; et al. Glycosylation of Serum Ribonuclease 1 Indicates a Major Endothelial Origin and Reveals an Increase in Core Fucosylation in Pancreatic Cancer. Glycobiology 2007, 17, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Kusama, K.; Okamoto, Y.; Saito, K.; Kasahara, T.; Murata, T.; Ueno, Y.; Kobayashi, Y.; Kamada, Y.; Miyoshi, E. Reevaluation of Pholiota Squarrosa Lectin-Reactive Haptoglobin as a Pancreatic Cancer Biomarker Using an Improved ELISA System. Glycoconj. J. 2017, 34, 537–544. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Llop, E.; Guerrero, P.E.; Duran, A.; Barrabés, S.; Massaguer, A.; Ferri, M.J.; Albiol-Quer, M.; De Llorens, R.; Peracaula, R. Glycoprotein Biomarkers for the Detection of Pancreatic Ductal Adenocarcinoma. World J. Gastroenterol. 2018, 24, 2537–2554. [Google Scholar] [CrossRef]
- Silva, M.L.S. Cancer Serum Biomarkers Based on Aberrant Post-Translational Modifications of Glycoproteins: Clinical Value and Discovery Strategies. Biochim. Biophys. Acta—Rev. Cancer 2015, 1856, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Nichetti, F.; Marra, A.; Corti, F.; Guidi, A.; Raimondi, A.; Prinzi, N.; de Braud, F.; Pusceddu, S. The Role of Mesothelin as a Diagnostic and Therapeutic Target in Pancreatic Ductal Adenocarcinoma: A Comprehensive Review. Target. Oncol. 2018, 13, 333–351. [Google Scholar] [CrossRef]
- Hassan, R.; Bera, T.; Pastan, I. Mesothelin: A New Target for Immunotherapy. Clin. Cancer Res. 2004, 10, 3937–3942. [Google Scholar] [CrossRef]
- Scholler, N.; Fu, N.; Yang, Y.; Ye, Z.; Goodman, G.E.; Hellstrom, K.E.; Hellstrom, I. Soluble Member(s) of the Mesothelin/Megakaryocyte Potentiating Factor Family Are Detectable in Sera from Patients with Ovarian Carcinoma. Proc. Natl. Acad. Sci. USA 1999, 96, 11531–11536. [Google Scholar] [CrossRef]
- Rump, A.; Morikawa, Y.; Tanaka, M.; Minami, S.; Umesaki, N.; Takeuchi, M.; Miyajima, A. Binding of Ovarian Cancer Antigen CA125/MUC16 to Mesothelin Mediates Cell Adhesion. J. Biol. Chem. 2004, 279, 9190–9198. [Google Scholar] [CrossRef] [PubMed]
- Gubbels, J.A.A.; Belisle, J.; Onda, M.; Rancourt, C.; Migneault, M.; Ho, M.; Bera, T.K.; Connor, J.P.; Sathyanarayana, B.K.; Lee, B.; et al. Mesothelin-MUC16 Binding Is a High Affinity, N-Glycan Dependent Interaction That Facilitates Peritoneal Metastasis of Ovarian Tumors. Mol. Cancer 2006, 5, 50. [Google Scholar] [CrossRef]
- Hassan, R.; Thomas, A.; Alewine, C.; Le, D.T.; Jaffee, E.M.; Pastan, I. Mesothelin Immunotherapy for Cancer: Ready for Prime Time? J. Clin. Oncol. 2016, 34, 4171–4179. [Google Scholar] [CrossRef] [PubMed]
- Montemagno, C.; Cassim, S.; Pouyssegur, J.; Broisat, A.; Pagès, G. From Malignant Progression to Therapeutic Targeting: Current Insights of Mesothelin in Pancreatic Ductal Adenocarcinoma. Int. J. Mol. Sci. 2020, 21, 4067. [Google Scholar] [CrossRef]
- Zheng, C.; Jia, W.; Tang, Y.; Zhao, H.; Jiang, Y.; Sun, S. Mesothelin Regulates Growth and Apoptosis in Pancreatic Cancer Cells through P53-Dependent and -Independent Signal Pathway. J. Exp. Clin. Cancer Res. CR 2012, 31, 84. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, U.; Marin-Muller, C.; Li, M.; Chen, C.; Yao, Q. Mesothelin Confers Pancreatic Cancer Cell Resistance to TNF-α-Induced Apoptosis through Akt/PI3K/NF-ΚB Activation and IL-6/Mcl-1 Overexpression. Mol. Cancer 2011, 10, 106. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wang, L.; Riedel, H.; Wang, K.; Yang, Y.; Dinu, C.Z.; Rojanasakul, Y. Mesothelin Promotes Epithelial-to-Mesenchymal Transition and Tumorigenicity of Human Lung Cancer and Mesothelioma Cells. Mol. Cancer 2017, 16, 63. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.; Pastan, I. Molecular Cloning of Mesothelin, a Differentiation Antigen Present on Mesothelium, Mesotheliomas, and Ovarian Cancers. Proc. Natl. Acad. Sci. USA 1996, 93, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Remaley, A.T.; Sampson, M.L.; Zhang, J.; Cox, D.D.; Pingpank, J.; Alexander, R.; Willingham, M.; Pastan, I.; Onda, M. Detection and Quantitation of Serum Mesothelin, a Tumor Marker for Patients with Mesothelioma and Ovarian Cancer. Clin. Cancer Res. 2006, 12, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Hanaoka, T.; Hasegawa, K.; Kato, T.; Sato, S.; Kurosaki, A.; Miyara, A.; Nagao, S.; Seki, H.; Yasuda, M.; Fujiwara, K. Correlation Between Tumor Mesothelin Expression and Serum Mesothelin in Patients with Epithelial Ovarian Carcinoma: A Potential Noninvasive Biomarker for Mesothelin-Targeted Therapy. Mol. Diagn. Ther. 2017, 21, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Li, P. Mesothelin as a Biomarker for Targeted Therapy. Biomark. Res. 2019, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Argani, P.; Iacobuzio-Donahue, C.; Ryu, B.; Rosty, C.; Goggins, M.; Wilentz, R.E.; Murugesan, S.R.; Leach, S.D.; Jaffee, E.; Yeo, C.J.; et al. Mesothelin Is Overexpressed in the Vast Majority of Ductal Adenocarcinomas of the Pancreas: Identification of a New Pancreatic Cancer Marker by Serial Analysis of Gene Expression (SAGE). Clin. Cancer Res. 2001, 7, 3862–3868. [Google Scholar] [PubMed]
- Hassan, R.; Laszik, Z.G.; Lerner, M.; Raffeld, M.; Postier, R.; Brackett, D. Mesothelin Is Overexpressed in Pancreaticobiliary Adenocarcinomas But Not in Normal Pancreas and Chronic Pancreatitis. Am. J. Clin. Pathol. 2005, 124, 838–845. [Google Scholar] [CrossRef] [PubMed]
- Frank, R.; Li, S.; Ahmad, N.A.; Sepulveda, A.R.; Jhala, N.C. Mesothelin Expression in Pancreatic Mucinous Cysts. Am. J. Clin. Pathol. 2014, 142, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Einama, T.; Kamachi, H.; Nishihara, H.; Homma, S.; Kanno, H.; Ishikawa, M.; Kawamata, F.; Konishi, Y.; Sato, M.; Tahara, M.; et al. Importance of Luminal Membrane Mesothelin Expression in Intraductal Papillary Mucinous Neoplasms. Oncol. Lett. 2015, 9, 1583–1589. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez, N.G. Application of Mesothelin Immunostaining in Tumor Diagnosis. Am. J. Surg. Pathol. 2003, 27, 1418–1428. [Google Scholar] [CrossRef]
- Weidemann, S.; Perez, D.; Izbicki, J.R.; Neipp, M.; Mofid, H.; Daniels, T.; Nahrstedt, U.; Jacobsen, F.; Bernreuther, C.; Simon, R.; et al. Mesothelin Is Commonly Expressed in Pancreatic Adenocarcinoma but Unrelated to Cancer Aggressiveness. Cancer Investig. 2021, 39, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Jhala, N.; Jhala, D.; Vickers, S.M.; Eltoum, I.; Batra, S.K.; Manne, U.; Eloubeidi, M.; Jones, J.J.; Grizzle, W.E. Biomarkers in Diagnosis of Pancreatic Carcinoma in Fine-Needle Aspirates: A Translational Research Application. Am. J. Clin. Pathol. 2006, 126, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Fukushima, N.; Maitra, A.; Iacobuzio-Donahue, C.A.; van Heek, N.T.; Cameron, J.L.; Yeo, C.J.; Hruban, R.H.; Goggins, M. Gene Expression Profiling Identifies Genes Associated with Invasive Intraductal Papillary Mucinous Neoplasms of the Pancreas. Am. J. Pathol. 2004, 164, 903–914. [Google Scholar] [CrossRef]
- Kendrick, Z.W.; Firpo, M.A.; Repko, R.C.; Scaife, C.L.; Adler, D.G.; Boucher, K.M.; Mulvihill, S.J. Serum IGFBP2 and MSLN as Diagnostic and Prognostic Biomarkers for Pancreatic Cancer. Hpb 2014, 16, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Beyer, H.L.; Geschwindt, R.D.; Glover, C.L.; Tran, L.; Hellstrom, I.; Hellstrom, K.E.; Miller, M.C.; Verch, T.; Allard, W.J.; Pass, H.I.; et al. MESOMARK: A Potential Test for Malignant Pleural Mesothelioma. Clin. Chem. 2007, 53, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Sharon, E.; Zhang, J.; Hollevoet, K.; Steinberg, S.M.; Pastan, I.; Onda, M.; Gaedcke, J.; Ghadimi, B.M.; Ried, T.; Hassan, R. Serum Mesothelin and Megakaryocyte Potentiating Factor in Pancreatic and Biliary Cancers. Clin. Chem. Lab. Med. 2012, 50, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.; Bhimani, N.; Gill, A.J.; Samra, J.S.; Sahni, S.; Mittal, A. Serum Biomarker Panel for Diagnosis and Prognosis of Pancreatic Ductal Adenocarcinomas. Front. Oncol. 2021, 11, 708963. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.; Onda, M.; Wang, Q.C.; Hassan, R.; Pastan, I.; Lively, M.O. Mesothelin Is Shed from Tumor Cells. Cancer Epidemiol. Biomark. Prev. 2006, 15, 1751. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Tateno, H.; Dohra, H.; Moriwaki, K.; Miyoshi, E.; Hirabayashi, J.; Kawagishi, H. A Novel Core Fucose-Specific Lectin from the Mushroom Pholiota Squarrosa. J. Biol. Chem. 2012, 287, 33973–33982. [Google Scholar] [CrossRef] [PubMed]
- Royle, L.; Campbell, M.P.; Radcliffe, C.M.; White, D.M.; Harvey, D.J.; Abrahams, J.L.; Kim, Y.-G.; Henry, G.W.; Shadick, N.A.; Weinblatt, M.E.; et al. HPLC-Based Analysis of Serum N-Glycans on a 96-Well Plate Platform with Dedicated Database Software. Anal. Biochem. 2008, 376, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rebelo, A.L.; Gubinelli, F.; Roost, P.; Jan, C.; Brouillet, E.; Van Camp, N.; Drake, R.R.; Saldova, R.; Pandit, A. Complete Spatial Characterisation of N-Glycosylation upon Striatal Neuroinflammation in the Rodent Brain. J. Neuroinflamm. 2021, 18, 116. [Google Scholar] [CrossRef]
- Campbell, M.P.; Royle, L.; Radcliffe, C.M.; Dwek, R.A.; Rudd, P.M. GlycoBase and AutoGU: Tools for HPLC-Based Glycan Analysis. Bioinformatics 2008, 24, 1214–1216. [Google Scholar] [CrossRef]
- Abrahams, J.L.; Campbell, M.P.; Packer, N.H. Building a PGC-LC-MS N-Glycan Retention Library and Elution Mapping Resource. Glycoconj. J. 2018, 35, 15–29. [Google Scholar] [CrossRef]
- Le, D.T.; Brockstedt, D.G.; Nir-Paz, R.; Hampl, J.; Mathur, S.; Nemunaitis, J.; Sterman, D.H.; Hassan, R.; Lutz, E.; Moyer, B.; et al. A Live-Attenuated Listeria Vaccine (ANZ-100) and a Live-Attenuated Listeria Vaccine Expressing Mesothelin (CRS-207) for Advanced Cancers: Phase I Studies of Safety and Immune Induction. Clin. Cancer Res. 2012, 18, 858–868. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Sharon, E.; Thomas, A.; Zhang, J.; Ling, A.; Miettinen, M.; Kreitman, R.J.; Steinberg, S.M.; Hollevoet, K.; Pastan, I. Phase 1 Study of the Antimesothelin Immunotoxin SS1P in Combination with Pemetrexed and Cisplatin for Front-Line Therapy of Pleural Mesothelioma and Correlation of Tumor Response with Serum Mesothelin, Megakaryocyte Potentiating Factor, and Cancer Antigen: SS1P Plus Chemotherapy for Mesothelioma. Cancer 2014, 120, 3311–3319. [Google Scholar] [CrossRef] [PubMed]
- Kreitman, R.J.; Hassan, R.; FitzGerald, D.J.; Pastan, I. Phase I Trial of Continuous Infusion Anti-Mesothelin Recombinant Immunotoxin SS1P. Clin. Cancer Res. 2009, 15, 5274–5279. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Alewine, C.; Pastan, I. New Life for Immunotoxin Cancer Therapy. Clin. Cancer Res. 2016, 22, 1055–1058. [Google Scholar] [CrossRef] [PubMed]
- Hollevoet, K.; Mason-Osann, E.; Liu, X.; Imhof-Jung, S.; Niederfellner, G.; Pastan, I. In Vitro and In Vivo Activity of the Low-Immunogenic Antimesothelin Immunotoxin RG7787 in Pancreatic Cancer. Mol. Cancer Ther. 2014, 13, 2040–2049. [Google Scholar] [CrossRef]
- Matsuzawa, F.; Kamachi, H.; Mizukami, T.; Einama, T.; Kawamata, F.; Fujii, Y.; Fukai, M.; Kobayashi, N.; Hatanaka, Y.; Taketomi, A. Mesothelin Blockage by Amatuximab Suppresses Cell Invasiveness, Enhances Gemcitabine Sensitivity and Regulates Cancer Cell Stemness in Mesothelin-Positive Pancreatic Cancer Cells. BMC Cancer 2021, 21, 200. [Google Scholar] [CrossRef] [PubMed]
- Hassan, R.; Ebel, W.; Routhier, E.L.; Patel, R.; Kline, J.B.; Zhang, J.; Chao, Q.; Jacob, S.; Turchin, H.; Gibbs, L.; et al. Preclinical Evaluation of MORAb-009, a Chimeric Antibody Targeting Tumor-Associated Mesothelin. Cancer Immun. 2007, 7, 20. [Google Scholar] [PubMed]
- Golfier, S.; Kopitz, C.; Kahnert, A.; Heisler, I.; Schatz, C.A.; Stelte-Ludwig, B.; Mayer-Bartschmid, A.; Unterschemmann, K.; Bruder, S.; Linden, L.; et al. Anetumab Ravtansine: A Novel Mesothelin-Targeting Antibody–Drug Conjugate Cures Tumors with Heterogeneous Target Expression Favored by Bystander Effect. Mol. Cancer Ther. 2014, 13, 1537–1548. [Google Scholar] [CrossRef] [PubMed]
- Weekes, C.D.; Lamberts, L.E.; Borad, M.J.; Voortman, J.; McWilliams, R.R.; Diamond, J.R.; de Vries, E.G.E.; Verheul, H.M.; Lieu, C.H.; Kim, G.P.; et al. Phase I Study of DMOT4039A, an Antibody–Drug Conjugate Targeting Mesothelin, in Patients with Unresectable Pancreatic or Platinum-Resistant Ovarian Cancer. Mol. Cancer Ther. 2016, 15, 439–447. [Google Scholar] [CrossRef]
- Morello, A.; Sadelain, M.; Adusumilli, P.S. Mesothelin-Targeted CARs: Driving T Cells to Solid Tumors. Cancer Discov. 2016, 6, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Hollevoet, K.; Reitsma, J.B.; Creaney, J.; Grigoriu, B.D.; Robinson, B.W.; Sherpereel, A.; Cristaudo, A.; Pass, H.I.; Nackaerts, K.; Rodríguez Portal, J.A.; et al. Serum mesothelin for diagnosing malignant pleural mesothelioma: An individual patient data meta-analysis. J. Clin. Oncol. 2012, 30, 1541–1549. [Google Scholar] [CrossRef]
- Ibrahim, D.A.; Abouhashem, N.S. Diagnostic Value of IMP3 and Mesothelin in Differentiating Pancreatic Ductal Adenocarcinoma from Chronic Pancreatitis. Pathol. Res. Pract. 2016, 212, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Llop, E.; Ferrer-Batallé, M.; Barrabés, S.; Guerrero, P.E.; Ramírez, M.; Saldova, R.; Rudd, P.M.; Aleixandre, R.N.; Comet, J.; de Llorens, R.; et al. Improvement of Prostate Cancer Diagnosis by Detecting PSA Glycosylation-Specific Changes. Theranostics 2016, 6, 1190–1204. [Google Scholar] [CrossRef]
- Qi, F.; Zhou, A.; Yan, L.; Yuan, X.; Wang, D.; Chang, R.; Zhang, Y.; Shi, F.; Han, X.; Hou, J.; et al. The Diagnostic Value of PIVKA-II, AFP, AFP-L3, CEA, and Their Combinations in Primary and Metastatic Hepatocellular Carcinoma. J. Clin. Lab. Anal. 2020, 34, e23158. [Google Scholar] [CrossRef]
- Salminen, L.; Nadeem, N.; Jain, S.; Grènman, S.; Carpén, O.; Hietanen, S.; Oksa, S.; Lamminmäki, U.; Pettersson, K.; Gidwani, K.; et al. A Longitudinal Analysis of CA125 Glycoforms in the Monitoring and Follow up of High Grade Serous Ovarian Cancer. Gynecol. Oncol. 2020, 156, 689–694. [Google Scholar] [CrossRef]
- Wang, Y.-N.; Lee, H.-H.; Hsu, J.L.; Yu, D.; Hung, M.-C. The Impact of PD-L1 N-Linked Glycosylation on Cancer Therapy and Clinical Diagnosis. J. Biomed. Sci. 2020, 27, 77. [Google Scholar] [CrossRef] [PubMed]
- Uray, K.; Mizuno, M.; Inazu, T.; Goto, K.; Hudecz, F. The Effect of Glycosylation on the Antibody Recognition of a MUC2 Mucin Epitope: Effect of Glycosylation on MUC2 Mucin Epitope. Biopolymers 2014, 102, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Inami, K.; Kajino, K.; Abe, M.; Hagiwara, Y.; Maeda, M.; Suyama, M.; Watanabe, S.; Hino, O. Secretion of N-ERC/Mesothelin and Expression of C-ERC/Mesothelin in Human Pancreatic Ductal Carcinoma. Oncol. Rep. 2008, 20, 1375–1380. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Liu, Y.; Chen, G. Diagnostic Value of Mesothelin in Pancreatic Cancer: A Meta-Analysis. Int. J. Clin. Exp. Med. 2014, 7, 4000–4007. [Google Scholar] [PubMed]
- Le, K.; Wang, J.; Zhang, T.; Guo, Y.; Chang, H.; Wang, S.; Zhu, B. Overexpression of Mesothelin in Pancreatic Ductal Adenocarcinoma (PDAC). Int. J. Med. Sci. 2020, 17, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Mann, B.F.; Goetz, J.A.; House, M.G.; Schmidt, C.M.; Novotny, M.V. Glycomic and Proteomic Profiling of Pancreatic Cyst Fluids Identifies Hyperfucosylated Lactosamines on the N-Linked Glycans of Overexpressed Glycoproteins. Mol. Cell. Proteom. 2012, 11, M111-015792. [Google Scholar] [CrossRef] [PubMed]
- Kamada, Y.; Kinoshita, N.; Tsuchiya, Y.; Kobayashi, K.; Fujii, H.; Terao, N.; Kamihagi, K.; Koyama, N.; Yamada, S.; Daigo, Y.; et al. Reevaluation of a Lectin Antibody ELISA Kit for Measuring Fucosylated Haptoglobin in Various Conditions. Clin. Chim. Acta 2013, 417, 48–53. [Google Scholar] [CrossRef]
- Zhao, J.; Qiu, W.; Simeone, D.M.; Lubman, D.M. N-Linked Glycosylation Profiling of Pancreatic Cancer Serum Using Capillary Liquid Phase Separation Coupled with Mass Spectrometric Analysis. J. Proteome Res. 2007, 6, 1126–1138. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-M.; Hwang, M.P.; Kim, Y.-W.; Kim, K.-J.; Jin, J.M.; Kim, Y.H.; Yang, Y.-H.; Lee, K.H.; Kim, Y.-G. Mass Spectrometry-Based N-Linked Glycomic Profiling as a Means for Tracking Pancreatic Cancer Metastasis. Carbohydr. Res. 2015, 413, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, E.; Boelaars, K.; Brown, K.; Eveline Li, R.J.; Kruijssen, L.; Bruijns, S.C.M.; van Ee, T.; Schetters, S.T.T.; Crommentuijn, M.H.W.; van der Horst, J.C.; et al. Sialic Acids in Pancreatic Cancer Cells Drive Tumour-Associated Macrophage Differentiation via the Siglec Receptors Siglec-7 and Siglec-9. Nat. Commun. 2021, 12, 1270. [Google Scholar] [CrossRef]
- Ghaderi, D.; Zhang, M.; Hurtado-Ziola, N.; Varki, A. Production Platforms for Biotherapeutic Glycoproteins. Occurrence, Impact, and Challenges of Non-Human Sialylation. Biotechnol. Genet. Eng. Rev. 2012, 28, 147–176. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Franc, V.; Heck, A.J.R. Glycoproteomics: A Balance between High-Throughput and In-Depth Analysis. Trends Biotechnol. 2017, 35, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Tabang, D.N.; Ford, M.; Li, L. Recent Advances in Mass Spectrometry-Based Glycomic and Glycoproteomic Studies of Pancreatic Diseases. Front. Chem. 2021, 9, 707387. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathology | Cases | N | Male/Female | Age Average | Range |
|---|---|---|---|---|---|
| Control | Healthy | 2 | 1/1 | 59 | 50–68 |
| NT cholangiocarcinoma | 2 | 1/1 | 74.5 | 71–78 | |
| NT duodenum adenocarcinoma | 2 | 0/2 | 75 | 72–78 | |
| NT Ampullary carcinoma | 1 | 1/0 | 67 | - | |
| NT PaC | 3 | 2/1 | 68.3 | 64–77 | |
| PaC | IIA | 6 | 4/2 | 69.5 | 64–78 |
| IIB | 21 | 10/1 | 64.2 | 49–79 | |
| III | 2 | 2/0 | 58 | 52–64 | |
| IV | 2 | 2/0 | 70.5 | 65–76 | |
| Other gastrointestinal malignancies | Ampullary carcinoma | 1 | 0/1 | 77 | - |
| Cholangiocarcinoma | 1 | 1/0 | 78 | - |
| Pathology | Cases | N | Male/Female | Age Average | Range |
|---|---|---|---|---|---|
| Control | Healthy | 5 | 3/2 | 59.4 | 44–75 |
| ChP | 11 | 8/3 | 51.8 | 42–72 | |
| PaC | IIA | 4 | 2/2 | 68.5 | 61–78 |
| IIB | 3 | 2/1 | 73 | 64–81 | |
| III | 3 | 3/0 | 63 | 52–73 | |
| IV | 5 | 3/2 | 62 | 30–76 |
| Lectin | Glycan Determinant | Concentration | Reference |
|---|---|---|---|
| Biotinylated Sambucus nigra Agglutinin (SNA) | Neu5Acα6Gal/GalNAc | 2 µg/mL | B-1305 (Vector Laboratories) |
| Biotinylated Maackia amurensis Lectin II (MAL-II) | Neu5Acα3Galβ3GalNAc | 2 µg/mL | B-1265 (Vector Laboratories) |
| Biotinylated Aleuria aurantia Lectin (AAL) | Fucα3/6GlcNAc | 2 µg/mL | B-1395 (Vector Laboratories) |
| Biotinylated Pholiota squarrosa Lectin (PhoSL) | Core fucose (α1,6) | 2 µg/mL | [55] |
| Fluorescein labelled Phaseolus vulgaris Leucoagglutinin (PHA-L) | Galβ4GlcNAcβ6 (GlcNAcβ2Manα3) Manα3 | 2 µg/mL | FL-1111 (Vector Laboratories) |
| Fluorescein labelled Phaseolus vulgaris Erythroagglutinin (PHA-E) | Galβ4GlcNAcβ2Manα6 (GlcNAcβ4) (GlcNAcβ4Manα3) Manβ4 | 2 µg/mL | FL-1121 (Vector Laboratories) |
| Digoxigenin labelled Peanut Agglutinin (PNA) | Galβ3GalNAc | 10 µg/mL | 11 210 238 001 (Roche) |
| Peptide | Sequence | N Mass (Da) | D Mass (Da) | Identification | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | D | ||||||||||||
| C | A | H | O | r | C | A | H | O | r | ||||
| Peptide 92-101 | WNVTSLETLK | 1189.64 | 1190.62 | X | X | X | X | X | |||||
| Peptide 187-200 | LAFQNMNGSEYFVK | 1646.78 | 1647.76 | X | X | ||||||||
| Peptide 215-230 | ALSQQNVSMDLATFMK | 1782.87 | 1783.85 | X | |||||||||
| Peptide ID | Sequence | Retention Time (min) | Theoretical Mass (Da) | Experimental Mass (Da) | Mass Deviation | Identification | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| C | A | H | O | rM | dM | ||||||
| 84-90 | MSPEDIR | 7.06 | 846.3905 | 846.4038 (424.2097 + 2H+) | +0.0133 | X | X | ||||
| 102-108 | ALLEVNK | 8.94 | 785.4647 | 785.4818 (393.7487 + 2H+) | +0.0171 | X | X | X | X | X | X |
| 247-256 | LLGPHVEGLK | 12.11 | 1061.6233 | 1061.6517 (354.8917 + 3H+) | +0.0284 | X | X | X | X | X | X |
| 177-184 | QLDVLYPK | 15.16 | 974.5437 | 974.5490 (488.2823 + 2H+) | +0.0053 | X | X | X | X | X | X |
| 109-122 | GHEMSPQVATLIDR | 17.05 | 1552.7667 | 1552.7793 (518.6009 + 3H+) | +0.0126 | X | X | ||||
| 266-270 | DWILR | 17.57 | 701.3860 | 701.4058 (351.7107 + 2H+) | +0.0198 | X | X | X | X | X | X |
| 201-214 | IQSFLGGAPTEDLK | 22.15 | 1474.7667 | 1474.7676 (738.3916 + 2H+) | +0.0009 | X | X | X | X | X | X |
| 233-246 | TDAVLPLTVAEVQK | 24.67 | 1482.8293 | 1482.8290 (742.4223 + 2H+) | −0.0003 | X | X | X | X | X | X |
| 187-200 | LAFQNMNGSEYFVK | 26.14 | 1646.7763 | 1647.7828 (824.8922 + 2H+) | +1.0065 | X | X | ||||
| 92-101 | WNVTSLETLK | 27.01 | 1189.6343 | 1190.6230 (596.3193 + 2H+) | +0.9887 | X | X | X | X | X | |
| 15-24 | EIDESLIFYK | 27.12 | 1255.6336 | 1255.6336 (628.8246 + 2H+) | ±0.0000 | X | X | X | X | ||
| 13-24 * | APEIDESLIFYK | 28.40 | 1423.7235 | 1423.7188 (712.8672 + 2H+) | −0.0047 | X | X | ||||
| 215-230 | ALSQQNVSMDLATFMK | 34.27 | 1782.8644 | 1783.8636 (595.6290 + 3H+) | +0.9992 | X | |||||
| 44-58 | VNAIPFTYEQLDVLK | 35.96 | 1748.9348 | 1748.9320 (875.4738 + 2H+) | −0.0028 | X | X | X | X | X | X |
| 26-43 | WELEACVDAALLATQMDR | 42.51 | 2090.9765 | 2090.9950 (698.0062 + 3H+) | +0.0185 | X | X | ||||
| 61-83 | LDELYPQGYPESVIQHLGYLFLK | 46.30 | 2721.4053 | 2721.3921 (908.1385 + 3H+) | −0.0132 | X | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duran, A.; Guerrero, P.E.; Ortiz, M.R.; Pérez del Campo, D.; Castro, E.; Garcia-Velasco, A.; Fort, E.; de Llorens, R.; Saldova, R.; Llop, E.; et al. Characterization of Mesothelin Glycosylation in Pancreatic Cancer: Decreased Core Fucosylated Glycoforms in Pancreatic Cancer Patients’ Sera. Biomedicines 2022, 10, 1942. https://doi.org/10.3390/biomedicines10081942
Duran A, Guerrero PE, Ortiz MR, Pérez del Campo D, Castro E, Garcia-Velasco A, Fort E, de Llorens R, Saldova R, Llop E, et al. Characterization of Mesothelin Glycosylation in Pancreatic Cancer: Decreased Core Fucosylated Glycoforms in Pancreatic Cancer Patients’ Sera. Biomedicines. 2022; 10(8):1942. https://doi.org/10.3390/biomedicines10081942
Chicago/Turabian StyleDuran, Adrià, Pedro E. Guerrero, Maria Rosa Ortiz, Dúnia Pérez del Campo, Ernesto Castro, Adelaida Garcia-Velasco, Esther Fort, Rafael de Llorens, Radka Saldova, Esther Llop, and et al. 2022. "Characterization of Mesothelin Glycosylation in Pancreatic Cancer: Decreased Core Fucosylated Glycoforms in Pancreatic Cancer Patients’ Sera" Biomedicines 10, no. 8: 1942. https://doi.org/10.3390/biomedicines10081942
APA StyleDuran, A., Guerrero, P. E., Ortiz, M. R., Pérez del Campo, D., Castro, E., Garcia-Velasco, A., Fort, E., de Llorens, R., Saldova, R., Llop, E., & Peracaula, R. (2022). Characterization of Mesothelin Glycosylation in Pancreatic Cancer: Decreased Core Fucosylated Glycoforms in Pancreatic Cancer Patients’ Sera. Biomedicines, 10(8), 1942. https://doi.org/10.3390/biomedicines10081942