
The Functional Properties and Physiological Roles of Signal-Transducing Adaptor Protein-2 in the Pathogenesis of Inflammatory and Immune Disorders


{kind=link}
{kind=link}
Abstract
1. Introduction
Contribution of Adaptor Proteins in T Cell Development and TCR-Mediated T Cell Activation
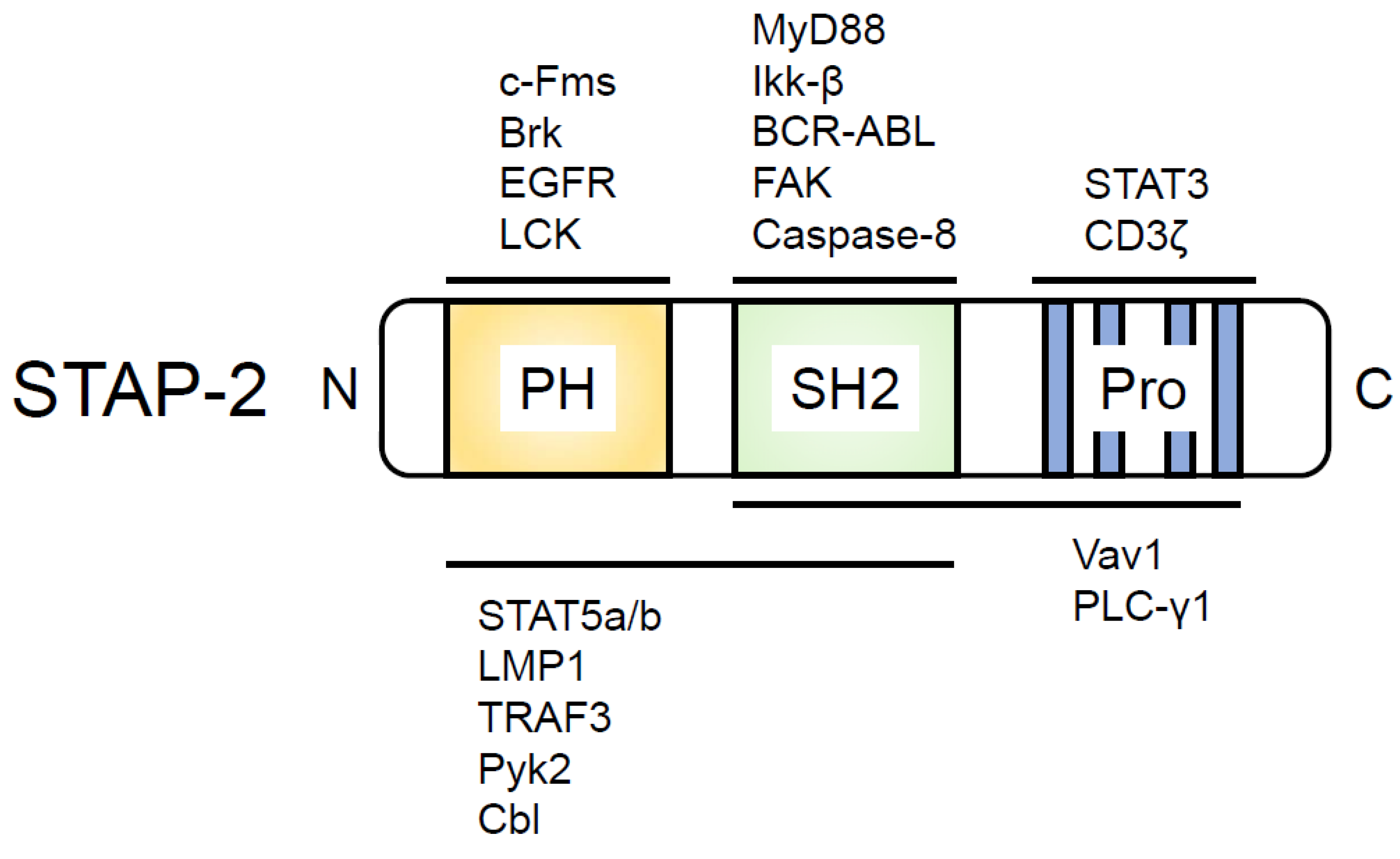
2. Signal-Transducing Adaptor Protein Family of Proteins
3. Functional Role of STAP-2 in T Cells
3.1. STAP-2 in Non-TCR-Mediated Signal Transduction
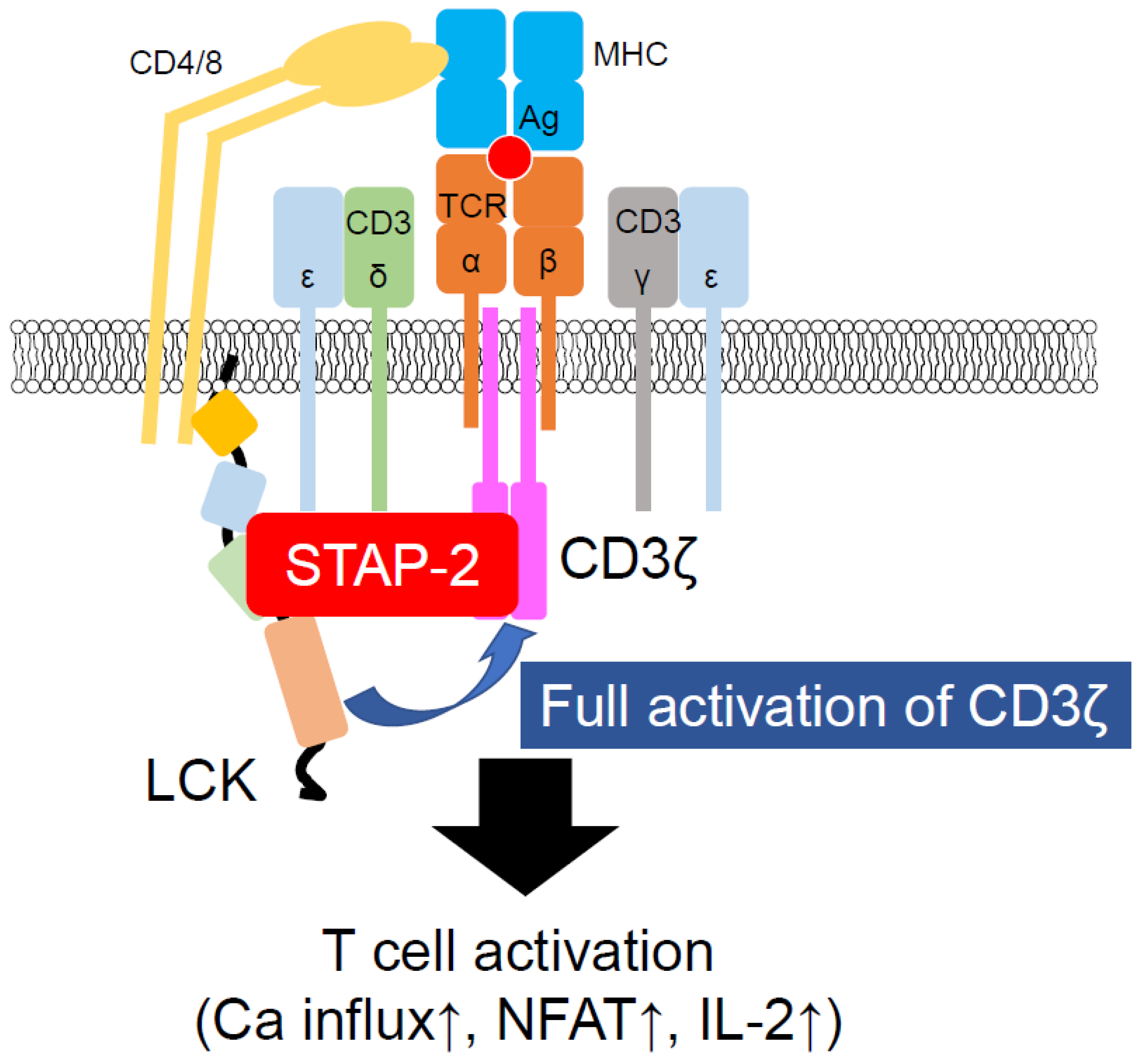
3.2. STAP-2 in TCR-Mediated Signal Transduction
4. Pathophysiological Role of STAP-2 in T Cells
4.1. STAP-2 in Infection
4.2. STAP-2 in Autoimmune Diseases
4.3. STAP-2 in Tumorigenesis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- van der Merwe, P.A.; Dushek, O. Mechanisms for T cell receptor triggering. Nat. Rev. Immunol. 2011, 11, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Gorentla, B.K.; Zhong, X.P. T cell Receptor Signal Transduction in T lymphocytes. J. Clin. Cell. Immunol. 2012, 2012, 5. [Google Scholar] [CrossRef] [PubMed]
- Gaud, G.; Lesourne, R.; Love, P.E. Regulatory mechanisms in T cell receptor signalling. Nat. Rev. Immunol. 2018, 18, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.R.; Byeon, Y.; Kim, D.; Park, S.G. Recent insights of T cell receptor-mediated signaling pathways for T cell activation and development. Exp. Mol. Med. 2020, 52, 750–761. [Google Scholar] [CrossRef]
- Shah, K.; Al-Haidari, A.; Sun, J.; Kazi, J.U. T cell receptor (TCR) signaling in health and disease. Signal Transduct. Target. Ther. 2021, 6, 412. [Google Scholar] [CrossRef]
- Wange, R.L. LAT, the linker for activation of T cells: A bridge between T cell-specific and general signaling pathways. Sci. STKE 2000, 2000, re1. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.K.; Berry, D.M.; McGlade, C.J. The role of Gads in hematopoietic cell signalling. Oncogene 2001, 20, 6284–6290. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Heyeck, S.D.; Wilcox, H.M.; Bunnell, S.C.; Berg, L.J. Lck phosphorylates the activation loop tyrosine of the Itk kinase domain and activates Itk kinase activity. J. Biol. Chem. 1997, 272, 25401–25408. [Google Scholar] [CrossRef] [PubMed]
- Andreotti, A.H.; Schwartzberg, P.L.; Joseph, R.E.; Berg, L.J. T-cell signaling regulated by the Tec family kinase, Itk. Cold Spring Harb. Perspect. Biol. 2010, 2, a002287. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.R.; Rao, A. NFAT, immunity and cancer: A transcription factor comes of age. Nat. Rev. Immunol. 2010, 10, 645–656. [Google Scholar] [CrossRef]
- Lo, W.L.; Shah, N.H.; Ahsan, N.; Horkova, V.; Stepanek, O.; Salomon, A.R.; Kuriyan, J.; Weiss, A. Lck promotes Zap70-dependent LAT phosphorylation by bridging Zap70 to LAT. Nat. Immunol. 2018, 19, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Sommers, C.L.; Burshtyn, D.N.; Stebbins, C.C.; DeJarnette, J.B.; Trible, R.P.; Grinberg, A.; Tsay, H.C.; Jacobs, H.M.; Kessler, C.M.; et al. Essential role of LAT in T cell development. Immunity 1999, 10, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, M.A.; Dazin, P.F.; Weiss, A. At least two non-antigen-binding molecules are required for signal transduction by the T-cell antigen receptor. Proc. Natl. Acad. Sci. USA 1988, 85, 8613–8617. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Irvin, B.J.; Trible, R.P.; Abraham, R.T.; Samelson, L.E. Functional analysis of LAT in TCR-mediated signaling pathways using a LAT-deficient Jurkat cell line. Int. Immunol. 1999, 11, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Pivniouk, V.; Tsitsikov, E.; Swinton, P.; Rathbun, G.; Alt, F.W.; Geha, R.S. Impaired viability and profound block in thymocyte development in mice lacking the adaptor protein SLP-76. Cell 1998, 94, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Yablonski, D.; Kuhne, M.R.; Kadlecek, T.; Weiss, A. Uncoupling of nonreceptor tyrosine kinases from PLC-gamma1 in an SLP-76-deficient T cell. Science 1998, 281, 413–416. [Google Scholar] [CrossRef]
- Nishida, K.; Wang, L.; Morii, E.; Park, S.J.; Narimatsu, M.; Itoh, S.; Yamasaki, S.; Fujishima, M.; Ishihara, K.; Hibi, M.; et al. Requirement of Gab2 for mast cell development and KitL/c-Kit signaling. Blood 2002, 99, 1866–1869. [Google Scholar] [CrossRef]
- Yamasaki, S.; Nishida, K.; Hibi, M.; Sakuma, M.; Shiina, R.; Takeuchi, A.; Ohnishi, H.; Hirano, T.; Saito, T. Docking protein Gab2 is phosphorylated by ZAP-70 and negatively regulates T cell receptor signaling by recruitment of inhibitory molecules. J. Biol. Chem. 2001, 276, 45175–45183. [Google Scholar] [CrossRef]
- Yamasaki, S.; Nishida, K.; Sakuma, M.; Berry, D.; McGlade, C.J.; Hirano, T.; Saito, T. Gads/Grb2-mediated association with LAT is critical for the inhibitory function of Gab2 in T cells. Mol. Cell. Biol. 2003, 23, 2515–2529. [Google Scholar] [CrossRef]
- Matsuda, T.; Oritani, K. STAP-2 Adaptor Protein Regulates Multiple Steps of Immune and Inflammatory Responses. Biol. Pharm. Bull. 2021, 44, 895–901. [Google Scholar] [CrossRef]
- Minoguchi, M.; Minoguchi, S.; Aki, D.; Joo, A.; Yamamoto, T.; Yumioka, T.; Matsuda, T.; Yoshimura, A. STAP-2/BKS, an adaptor/docking protein, modulates STAT3 activation in acute-phase response through its YXXQ motif. J. Biol. Chem. 2003, 278, 11182–11189. [Google Scholar] [CrossRef] [PubMed]
- Ohya, K.; Kajigaya, S.; Kitanaka, A.; Yoshida, K.; Miyazato, A.; Yamashita, Y.; Yamanaka, T.; Ikeda, U.; Shimada, K.; Ozawa, K.; et al. Molecular cloning of a docking protein, BRDG1, that acts downstream of the Tec tyrosine kinase. Proc. Natl. Acad. Sci. USA 1999, 96, 11976–11981. [Google Scholar] [CrossRef] [PubMed]
- Masuhara, M.; Nagao, K.; Nishikawa, M.; Sasaki, M.; Yoshimura, A.; Osawa, M. Molecular cloning of murine STAP-1, the stem-cell-specific adaptor protein containing PH and SH2 domains. Biochem. Biophys. Res. Commun. 2000, 268, 697–703. [Google Scholar] [CrossRef]
- Yokohari, K.; Yamashita, Y.; Okada, S.; Ohya, K.; Oda, S.; Hatano, M.; Mano, H.; Hirasawa, H.; Tokuhisa, T. Isoform-dependent interaction of BRDG1 with Tec kinase. Biochem. Biophys. Res. Commun. 2001, 289, 414–420. [Google Scholar] [CrossRef]
- Toda, J.; Ichii, M.; Oritani, K.; Shibayama, H.; Tanimura, A.; Saito, H.; Yokota, T.; Motooka, D.; Okuzaki, D.; Kitai, Y.; et al. Signal-transducing adapter protein-1 is required for maintenance of leukemic stem cells in CML. Oncogene 2020, 39, 5601–5615. [Google Scholar] [CrossRef] [PubMed]
- Ishiura, M.; Kitai, Y.; Kashiwakura, J.I.; Muromoto, R.; Toda, J.; Ichii, M.; Oritani, K.; Matsuda, T. Positive interactions between STAP-1 and BCR-ABL influence chronic myeloid leukemia cell proliferation and survival. Biochem. Biophys. Res. Commun. 2021, 556, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.J.; Sara, E.A.; Crompton, M.R. A novel adaptor-like protein which is a substrate for the non-receptor tyrosine kinase, BRK. Oncogene 2000, 19, 4273–4282. [Google Scholar] [CrossRef][Green Version]
- Ikeda, O.; Mizushima, A.; Sekine, Y.; Yamamoto, C.; Muromoto, R.; Nanbo, A.; Oritani, K.; Yoshimura, A.; Matsuda, T. Involvement of STAP-2 in Brk-mediated phosphorylation and activation of STAT5 in breast cancer cells. Cancer Sci. 2011, 102, 756–761. [Google Scholar] [CrossRef]
- Sekine, Y.; Yamamoto, T.; Yumioka, T.; Sugiyama, K.; Tsuji, S.; Oritani, K.; Shimoda, K.; Minoguchi, M.; Yoshimura, A.; Matsuda, T. Physical and functional interactions between STAP-2/BKS and STAT5. J. Biol. Chem. 2005, 280, 8188–8196. [Google Scholar] [CrossRef]
- Ikeda, O.; Sekine, Y.; Kakisaka, M.; Tsuji, S.; Muromoto, R.; Ohbayashi, N.; Oritani, K.; Yoshimura, A.; Matsuda, T. STAP-2 regulates c-Fms/M-CSF receptor signaling in murine macrophage Raw 264.7 cells. Biochem. Biophys. Res. Commun. 2007, 358, 931–937. [Google Scholar] [CrossRef]
- Ikeda, O.; Sekine, Y.; Muromoto, R.; Ohbayashi, N.; Yoshimura, A.; Matsuda, T. Enhanced c-Fms/M-CSF receptor signaling and wound-healing process in bone marrow-derived macrophages of signal-transducing adaptor protein-2 (STAP-2) deficient mice. Biol. Pharm. Bull. 2008, 31, 1790–1793. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ikeda, O.; Miyasaka, Y.; Sekine, Y.; Mizushima, A.; Muromoto, R.; Nanbo, A.; Yoshimura, A.; Matsuda, T. STAP-2 is phosphorylated at tyrosine-250 by Brk and modulates Brk-mediated STAT3 activation. Biochem. Biophys. Res. Commun. 2009, 384, 71–75. [Google Scholar] [CrossRef]
- Sekine, Y.; Tsuji, S.; Ikeda, O.; Kakisaka, M.; Sugiyama, K.; Yoshimura, A.; Matsuda, T. Leukemia inhibitory factor-induced phosphorylation of STAP-2 on tyrosine-250 is involved in its STAT3-enhancing activity. Biochem. Biophys. Res. Commun. 2007, 356, 517–522. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sekine, Y.; Ikeda, O.; Tsuji, S.; Yamamoto, C.; Muromoto, R.; Nanbo, A.; Oritani, K.; Yoshimura, A.; Matsuda, T. Signal-transducing adaptor protein-2 regulates stromal cell-derived factor-1 alpha-induced chemotaxis in T cells. J. Immunol. 2009, 183, 7966–7974. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, K.; Tsuchiya, T.; Kashiwakura, J.I.; Muromoto, R.; Kitai, Y.; Sekine, Y.; Oritani, K.; Matsuda, T. STAP-2 interacts with Pyk2 and enhances Pyk2 activity in T-cells. Biochem. Biophys. Res. Commun. 2017, 488, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. Integrins: Bidirectional, allosteric signaling machines. Cell 2002, 110, 673–687. [Google Scholar] [CrossRef] [PubMed]
- Sekine, Y.; Tsuji, S.; Ikeda, O.; Sugiyma, K.; Oritani, K.; Shimoda, K.; Muromoto, R.; Ohbayashi, N.; Yoshimura, A.; Matsuda, T. Signal-transducing adaptor protein-2 regulates integrin-mediated T cell adhesion through protein degradation of focal adhesion kinase. J. Immunol. 2007, 179, 2397–2407. [Google Scholar] [CrossRef]
- Sekine, Y.; Yamamoto, C.; Kakisaka, M.; Muromoto, R.; Kon, S.; Ashitomi, D.; Fujita, N.; Yoshimura, A.; Oritani, K.; Matsuda, T. Signal-transducing adaptor protein-2 modulates Fas-mediated T cell apoptosis by interacting with caspase-8. J. Immunol. 2012, 188, 6194–6204. [Google Scholar] [CrossRef]
- Saitoh, K.; Kashiwakura, J.I.; Kagohashi, K.; Sasaki, Y.; Kawahara, S.; Sekine, Y.; Kitai, Y.; Muromoto, R.; Ichii, M.; Nakatsukasa, H.; et al. STAP-2 Is a Novel Positive Regulator of TCR-Proximal Signals. J. Immunol. 2022, 209, 57–68. [Google Scholar] [CrossRef]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-like receptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [CrossRef]
- Weighardt, H.; Kaiser-Moore, S.; Vabulas, R.M.; Kirschning, C.J.; Wagner, H.; Holzmann, B. Cutting edge: Myeloid differentiation factor 88 deficiency improves resistance against sepsis caused by polymicrobial infection. J. Immunol. 2002, 169, 2823–2827. [Google Scholar] [CrossRef] [PubMed]
- Sekine, Y.; Yumioka, T.; Yamamoto, T.; Muromoto, R.; Imoto, S.; Sugiyma, K.; Oritani, K.; Shimoda, K.; Minoguchi, M.; Akira, S.; et al. Modulation of TLR4 signaling by a novel adaptor protein signal-transducing adaptor protein-2 in macrophages. J. Immunol. 2006, 176, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Umemura, M.; Yahagi, A.; Hamada, S.; Begum, M.D.; Watanabe, H.; Kawakami, K.; Suda, T.; Sudo, K.; Nakae, S.; Iwakura, Y.; et al. IL-17-mediated regulation of innate and acquired immune response against pulmonary Mycobacterium bovis bacille Calmette-Guerin infection. J. Immunol. 2007, 178, 3786–3796. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Yamamoto, Y.; Muromoto, R.; Ikeda, O.; Sekine, Y.; Grusby, M.J.; Kaisho, T.; Matsuda, T. PDLIM2 inhibits T helper 17 cell development and granulomatous inflammation through degradation of STAT3. Sci. Signal. 2011, 4, ra85. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Li, Z.; Yang, X.O.; Chang, S.H.; Nurieva, R.; Wang, Y.H.; Wang, Y.; Hood, L.; Zhu, Z.; Tian, Q.; et al. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nat. Immunol. 2005, 6, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Sekine, Y.; Ikeda, O.; Mizushima, A.; Ueno, Y.; Muromoto, R.; Yoshimura, A.; Kanakura, Y.; Oritani, K.; Matsuda, T. STAP-2 interacts with and modulates BCR-ABL-mediated tumorigenesis. Oncogene 2012, 31, 4384–4396. [Google Scholar] [CrossRef] [PubMed]
- Kitai, Y.; Iwakami, M.; Saitoh, K.; Togi, S.; Isayama, S.; Sekine, Y.; Muromoto, R.; Kashiwakura, J.I.; Yoshimura, A.; Oritani, K.; et al. STAP-2 protein promotes prostate cancer growth by enhancing epidermal growth factor receptor stabilization. J. Biol. Chem. 2017, 292, 19392–19399. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, D.; Seo, N.; Hayashi, T.; Hyuga-Amaike, C.; Okamori, K.; Tawara, I.; Harada, N.; Shiku, H. Signal-transducing adaptor protein-2 promotes generation of functional long-term memory CD8+ T cells by preventing terminal effector differentiation. Oncotarget 2017, 8, 30766–30780. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kashiwakura, J.-i.; Oritani, K.; Matsuda, T. The Functional Properties and Physiological Roles of Signal-Transducing Adaptor Protein-2 in the Pathogenesis of Inflammatory and Immune Disorders. Biomedicines 2022, 10, 3079. https://doi.org/10.3390/biomedicines10123079
Kashiwakura J-i, Oritani K, Matsuda T. The Functional Properties and Physiological Roles of Signal-Transducing Adaptor Protein-2 in the Pathogenesis of Inflammatory and Immune Disorders. Biomedicines. 2022; 10(12):3079. https://doi.org/10.3390/biomedicines10123079
Chicago/Turabian StyleKashiwakura, Jun-ichi, Kenji Oritani, and Tadashi Matsuda. 2022. "The Functional Properties and Physiological Roles of Signal-Transducing Adaptor Protein-2 in the Pathogenesis of Inflammatory and Immune Disorders" Biomedicines 10, no. 12: 3079. https://doi.org/10.3390/biomedicines10123079
APA StyleKashiwakura, J.-i., Oritani, K., & Matsuda, T. (2022). The Functional Properties and Physiological Roles of Signal-Transducing Adaptor Protein-2 in the Pathogenesis of Inflammatory and Immune Disorders. Biomedicines, 10(12), 3079. https://doi.org/10.3390/biomedicines10123079