1. Introduction
Single-species is a unit of the whole ecosphere. Although most biological populations are multi-species, there is no single population in the strict sense, but because of artificial breeding, there are a lot of single population resources in many human-created environments, which provide indispensable roles for human production and life. Due to the economic interests and production needs, people need to develop the single population resources for a long time and continuously, and to give full play to the best use value on the basis of the lowest cost consumption as possible, so the control and prediction of the single population is particularly important.
Single-species models are widely used in the field of mathematical biology, such as pest management, optimal management of biological resources, epidemic prevention and control, cell growth regulation and so on. Many scholars (see [
1,
2,
3]) have obtained some good properties of these models by quantitative analysis, and the results help us forecast and control the actual production. Wang et al. [
4] studied a single-species model and found an optimal harvesting strategy which allows the output to reach a maximum and remains constant. At the same time, the population quantity can attain the maximum level in a precise time interval when the population has been harvested. Dou et al. [
5] addressed a non-autonomous Logistic single-species model. The analytical expressions about the best harvesting strategy have been obtained by Pontryagin control and optimization principles of impulsive systems. Recently, many articles have also focused on the study of single-species population model (see [
6,
7,
8,
9]).
We know that both discrete and distributed delays tend to bring more rich and complex dynamics to a population dynamic system. For researchers, although models with delay are more complex than without delay, the research results of these models are closer to actual life. In 1948, Hutchinson [
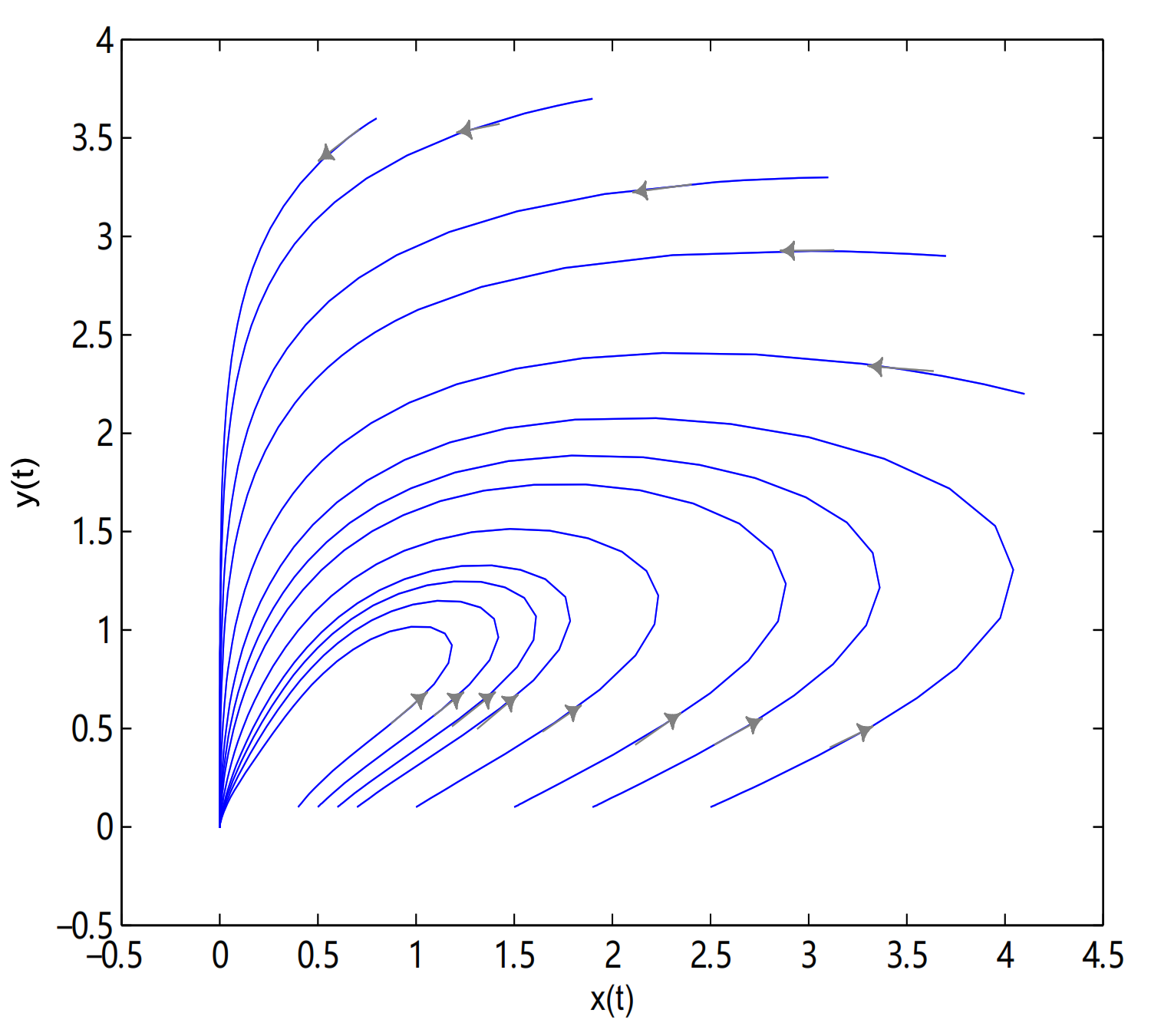
10] analyzed a Logistic delay equation characterizing animal populations, and the form is
which could vividly simulate the population size. In (
1), the food supply at time
t depends on the population number at time
. More generally, many scholars studied some models in which the current population density continuously depends on the population density in the past period. For example, Cushing [
11] proposed the following single population model with distributed delay
and detailed qualitative results were obtained. Pang et al. [
12] considered the following single population model with distributed delay and impulsive state feedback control
and sufficient conditions for the existence and stability of the order-1 periodic solution and limit cycles were obtained, where
. Ruan and Wolkowicz [
13] built a chemostat model about single population with distributed delay, and they obtained that the system existed Hopf bifurcations by taking the average time delay as the bifurcation parameter. Lian et al. [
14] considered a predator-prey system with Holling type IV functional response and time delay. They studied the effects of time delay on this system and obtained the conditions of local asymptotic stability of the positive equilibrium and the existence of local Hopf bifurcations by applying the delay as a bifurcation parameter. Yao et al. [
15] investigated the global asymptotic stability of fractional-order complex-valued differential equations with distributed delays. Based on the Laplace transform method, a novel necessary and sufficient condition for the stability is established by embedding the characteristic equation into a two-dimensional complex system. The algebraic criterion is expressed by the fractional exponent, coefficients, and the delay. Many single-species population models with time delay were studied and rich results were obtained (see [
16,
17,
18,
19,
20,
21,
22]).
In addition, many scholars have studied the single population model with different forms of harvest. Clark [
23] introduced the following single population model with linear harvest,
Clark introduced the single population model (
4) with linear harvest, analytical expressions for optimal harvest of a renewable resource stock, which is subject to a stochastic process are found. These expressions give the optimal harvest as an explicit feedback control law. All relations in system (
4), including the stochastic process, may be arbitrary functions of the state variable (stock). In 1978, Ludwig et al. [
24] put forward the following single population model with nonlinear harvest,
Nonlinear harvesting has been proposed and some novel conclusions have been obtained. Tan et al. [
25] considered the following single population system with implusive disturbance and nonlinear harvest:
They obtained good dynamic properties. Li et al. [
26] established and analyzed a single-species model with delay weak kernel and constant rate harvesting:
The existence of the equilibrium point of the model (
7) is obtained, and the properties of the positive equilibrium state of the population are studied.
Population model with distributed delay and harvest plays an important role in ecosystem, which can reflect the growth rule of invertebrate population better and help us get the best policy of harvesting. This paper is mainly aimed at qualitative analysis of a single-species population model with distributed delay and nonlinear harvest. By the establishment of a single-species population model of differential equations, and using stability analysis methods to discuss the existence and stability of equilibria of the system, we can forecast the development of the population.
The rest of this paper is organized as follows. In
Section 2, we establish a single-species model with distributed delay and nonlinear harvest and make a statement about the model. Then, we will discuss the existence and stability of the equilibria of the system under three different cases from
Section 3,
Section 4 and
Section 5. In
Section 6, we verify our results by numerical simulations. The conclusions will be given in
Section 7.
2. The Model
We combine distributed delay and nonlinear harvest to consider a new single-species model. From biological and economic points of view, Michaelis-Menten type harvesting is more realistic among the several types of harvesting. Thus, a single-species model with distributed delay and Michaelis-Menten type harvesting is established as follows:
where
denotes the density of the population at time
t,
r denotes the intrinsic rate of increase of the population,
and
c are positive constants.
indicates the impact of distributed time delay. The term
represents the effect of distributed delay from the following aspects: The integral for
s from
to
t represents the continuous influence on current state
from the past time, which shows the influence interval for distributed delay. That is, the entire interval from the past to current time
t has impact on current state
. The kernel function
represents the influential degree to current state
from time
s to
t.
represents the density of population at the past time
s. Therefore, the term
represents the distributed delay in the model.
is simplified Michaelis-Menten type harvesting (the details of this kind of harvesting can be seen in the Refs. [
27,
28,
29]).
Take the transform
which yields the following equivalent system
Considering the practical significance of system (
9), we discuss system (
9) in
in this paper.
We consider the existence of equilibria of system (
9). Let
the solutions of the equations are
and
( if
), where
Denoting
we can also write
We denote
,
and
throughout this paper. In the following discussion, we always assume
. Next, we analyze the existence and stability of the equilibria of system (
9) under three cases.
3. The Existence and Stability of the Equilibria for
Let us consider the existence and stability of the equilibria of system (
9) for the case
. Obviously,
and
in this case.
(1)
is clearly an equilibrium of system (
9).
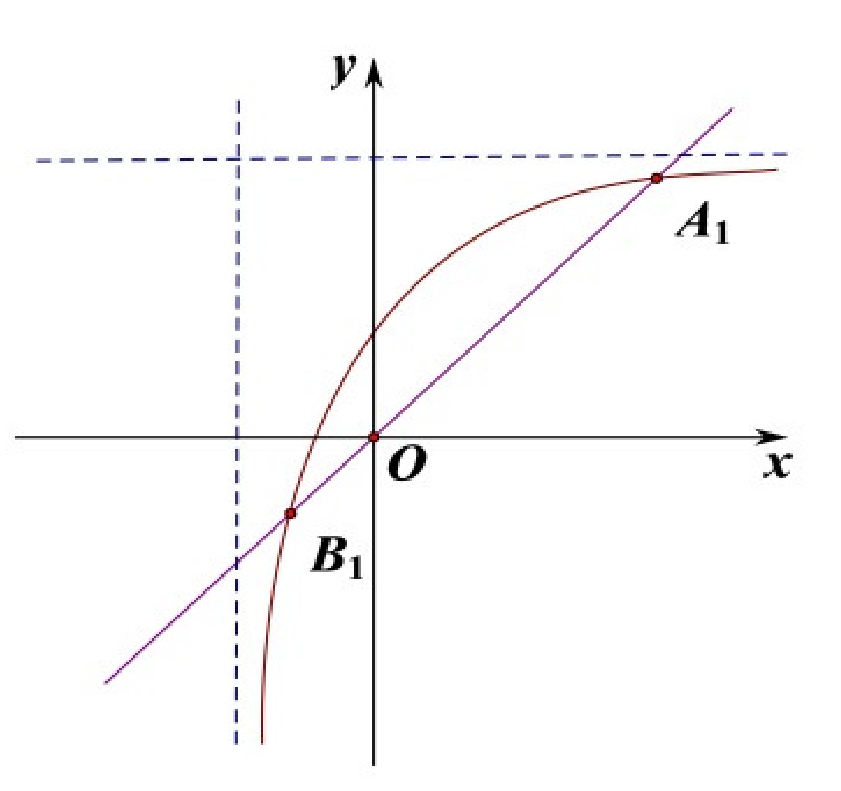
(2) When
,
is a positive equilibrium of system (
9),
is a negative equilibrium of system (
9) (see
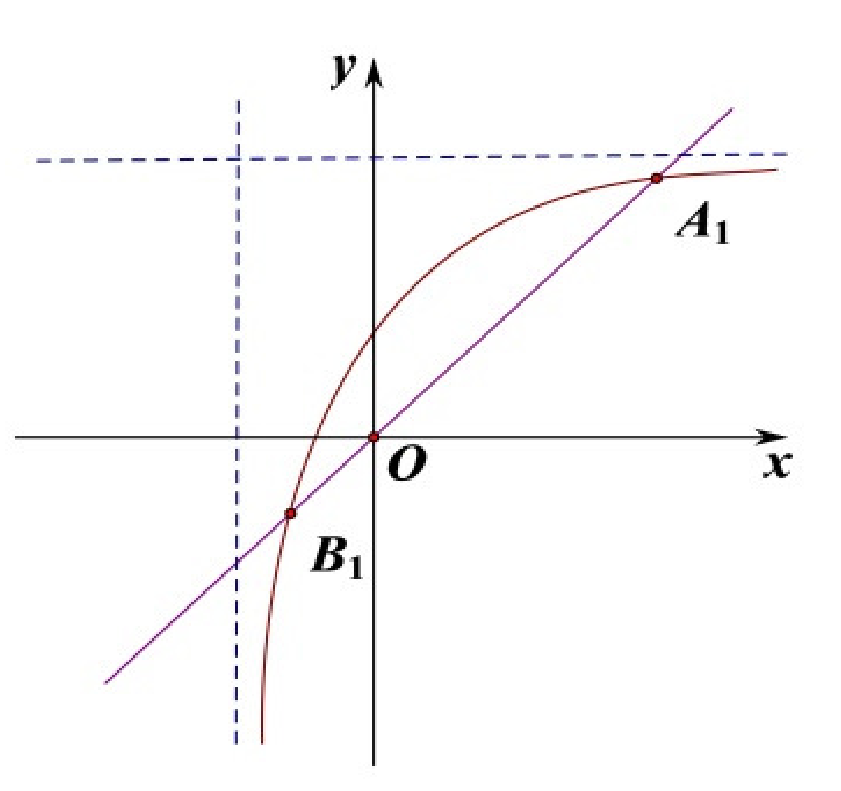
Figure 1).
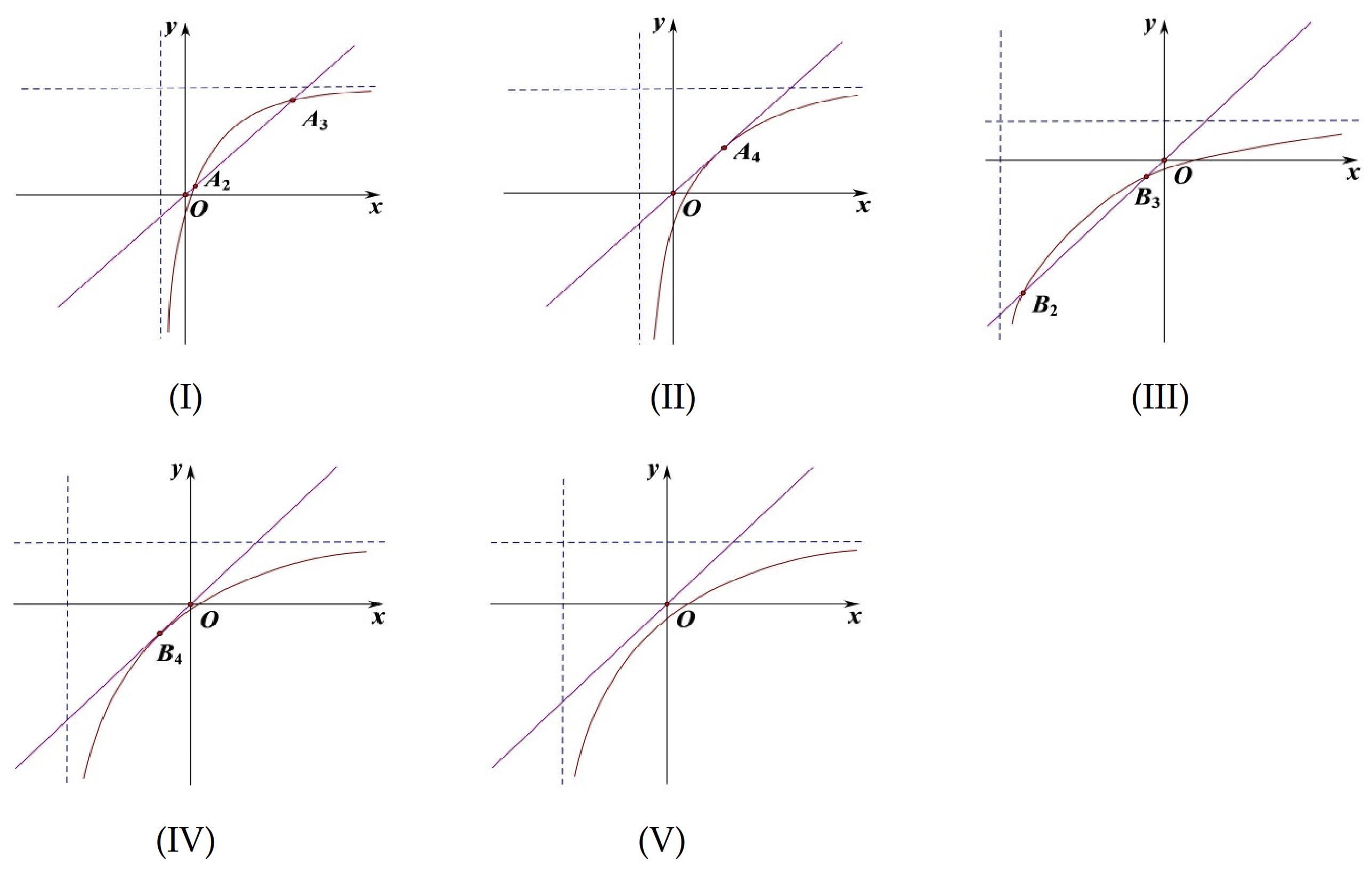
x denotes the density of the population, y indicates the impact of distributed time delay. There are two solid lines on every sub-chart. The straight line is the equation: , the curve is the equation: . The dotted lines in the Figures are asymptotes to the equation: . The intersections of two solid lines are shown in the following figure.
Theorem 1. If , the equilibrium of system (9) is a saddle point. If and the equilibrium of system (9) is globally asymptotically stable, where . Proof. When
, the Jacobi matrix of system (
9) is
At the equilibrium
O, the Jacobi matrix of system (
9) becomes
which allows us to obtain
Since
, then
. Obviously,
is a saddle point.
At the equilibrium
, the Jacobi matrix of system (
9) becomes
Then we get
Since we can also write
and
, it is easy for us to simplify
as follows
Since , we know that and , then .
Next, we simplify
as follows
Let
We have
for
,
as
,
when
.
We discuss the situation of i.e., .
If , then , the positive equilibrium is locally stable in this case. Next, we are going to prove that there is no closed cycle in . There is a unique positive equilibrium in , for convenience, we substitute for . Suppose that there is a closed orbit around , which is, , so there must exist a periodic T such that .
We calculate the index
of the closed orbit
where
We rewrite system (
9) as follows
Integrating the two sides along the closed orbit
, we get
So we know that
and
also satisfy the following algebraic equation
Hence
where
is the coordinate of the equilibrium
. We obtain the index
of the closed orbit
As
, we have
The closed orbit
is stable as
, which is contradicted by the fact that the equilibrium
is asymptotically stable. Therefore, there is no closed orbit.
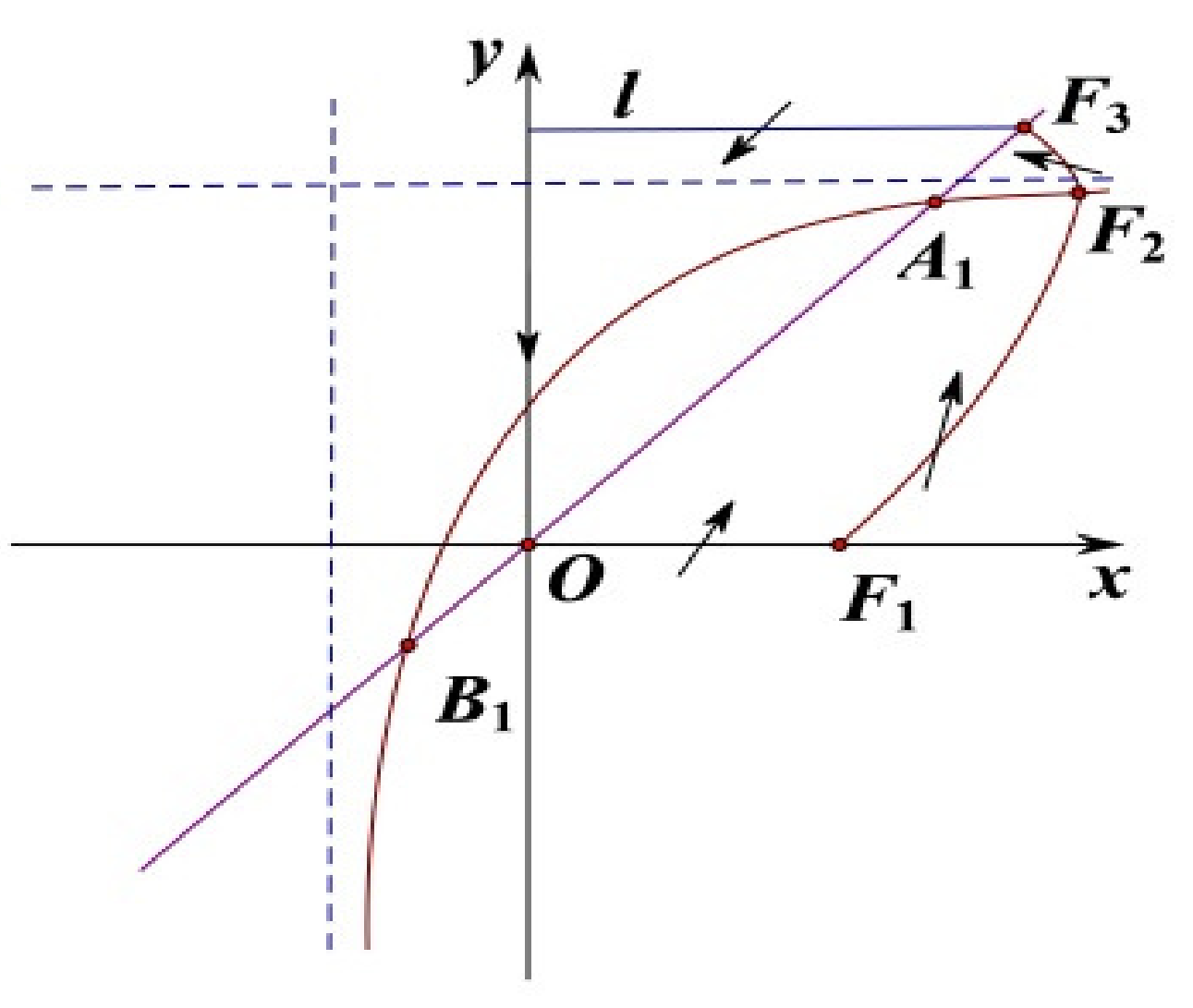
Now, we are going to prove that the solution of system (
9) is eventually uniformly bounded (see
Figure 2). Take a point
on
x-axis, draw the trajectory passing through the point
of the following equation
and the trajectory intersects isoclinic line
at
. Comparing the direction of trajectory of system (
9) with the tangent direction of trajectory
of the above system on
yields that
So the trajectory on
of system (
9) goes through into the interior of
from the outer of
when the trajectory of system (
9) intersects
. Then draw the trajectory of the following equation
passing through the point
, and the trajectory will intersect isoclinic line
at
. Comparing the direction of trajectory of system (
9) with the tangent direction of trajectory
of above system on
, we have
So the trajectory on
of system (
9) goes through into the interior of
from the outer of
when the trajectory of system (
9) intersects
. Then draw the line
l from the point
, which is perpendicular to
y-axis. The trajectory of system (
9) goes through into the interior of
l from the outer of
l when the trajectory of system (
9) intersects line
l. Furthermore,
y-axis is an isoclinic line, and the trajectory of system (
9) goes through into the interior of
x-axis from the outer of
x-axis when intersects
x-axis. According to the above analysis, the solution of system (
9) is eventually uniformly bounded in
. In addition, there is no closed orbit in
. So the unique positive equilibrium
is globally asymptotically stable. □
Next, we discuss the case of i.e., .
We shift point
of system (
9) to the origin and system (
9) can be rewritten as
where
is a
function of at least fourth order about
x. In this case,
, that is
, then
becomes
We can obtain
by
. So we can know that the eigenvalues of
are
and
(
i is the complex unit, similarly hereinafter) by simple calculation. Taking non-singular transformations
and
, and also denoting
by
, respectively, system (
11) becomes
where
where
is a
function of at least fourth order about
x and
y.
and
are
functions and satisfy
.
Using the formula for the Lyapunov number of Andronov et al. [
30], we have the first Lyapunov number
of the equilibrium
of system (
9) as follows
If
holds, it follows that
, where
So we obtain the following theorem:
Theorem 2. If and condition (12) holds, a supercritical Hopf bifurcation occurs near when η passes through 0, where . Then, we discuss the case of i.e., .
When
and
, the positive equilibrium
is an unstable focus or node. Given what has been proved before, it is not difficult to obtain that the solution of system (
9) is also uniformly bounded in
. Since the unique positive equilibrium
is unstable, we can obtain that there exists at least one limit cycle which is stable in region
by Poincaré-Bendixson Theorem.
Particularly, we have known that a Hopf bifurcation occurs when
and condition (
12) holds. That is to say, the equilibrium
is not a true center. The derivative of system (
9) at the point
is denoted as
. We can get the eigenvalues of
are
, i.e.,
We also define the matrix
which satisfies
We obtain that
. Using the Friedrich Theorem of Hopf bifurcation, we get the truth that there exists a unique stable limit cycle near
when
and condition (
12) holds. So we get the following Theorem:
Theorem 3. If and , there exists at least one limit cycle which is stable in Ω
. Particularly, there exists a unique stable limit cycle near when and condition (12) holds, where . 5. The Existence and Stability of the Equilibria for
Let us consider the existence and stability of the equilibria for the case .
(1)
is clearly an equilibrium of system (
9).
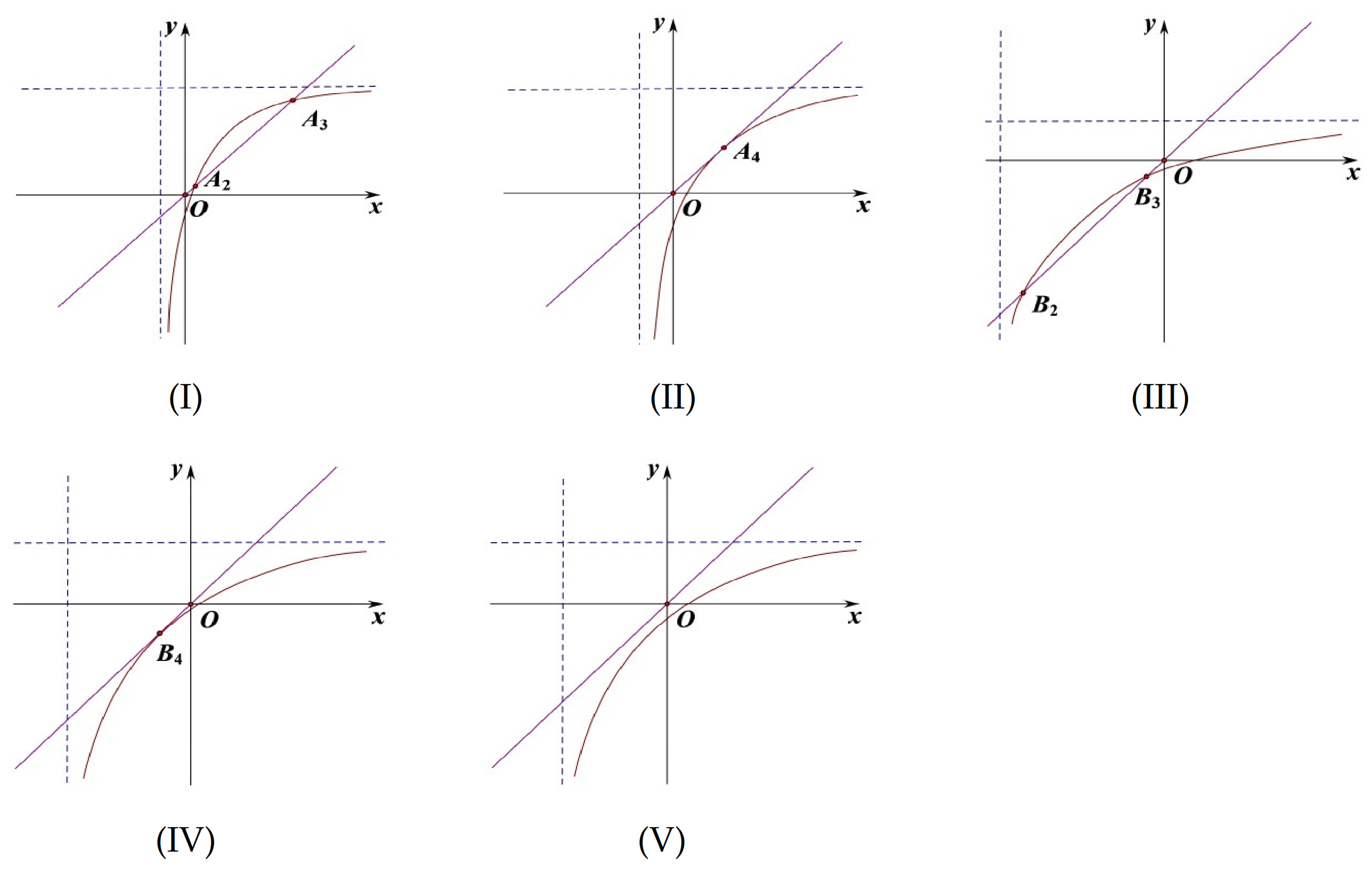
(2) When
and
,
is a positive equilibrium of system (
9), where
(see
Figure 4I).
(3) When
and
,
is a negative equilibrium of system (
9). This case will not be discussed (see
Figure 4II).
(4) When
and
, system (
9) has no more equilibria except
(see
Figure 4III).
Theorem 6. If and the equilibrium of system (9) is an unstable node, and system (9) has a singular line. If and the equilibrium is a saddle node. Proof. Firstly, we analyze the stability of the equilibrium
of system (
9). We take
into (10) and get
and
When
, we get
. It can easily be shown that the equilibrium
O is a higher order singular point. We change (
9) into (
19).
Performing the nonsingular transformations
,
and
, we still denote
and
by
and
t, and (
19) becomes
When
, i.e.,
, the positive equilibrium
does not exist under this condition, that is, the equilibrium
O is a triple singularity (
20) becomes
Let
, then
by Taylor expansion at
, we have
After some calculations, we have
By Theorem 7.1 in Chapter 2 of Zhang et al. [
31], we know that
O is an unstable node.
According to what we have proved before, the solution of system (
9) is eventually uniformly bounded in
under this case. Since
is the isoclinic line, it is not possible in the whole plane to have a closed orbit which surrounds the equilibrium
O. In addition, if the closed orbit exists in some quadrant, then there must be an equilibrium in the closed orbit. So we have the conclusion that there is no closed orbit in the whole plane when
and
. Meanwhile, the linear system of system (
19) is as follows
and
. It follows that system (
19) has a singular line
, i.e.,
when
and
. Then all the trajectories tend to the singular line
.
When
, i.e.,
, the positive equilibrium
exists if
, and the negative equilibrium
exists if
, that is, the equilibrium
O is a double singularity. We rewrite (
20) into the following equation
Let
, then
By some calculations, we derive
By Theorem 7.1 in Chapter 2 of [
31], we know that
O is a saddle node. □
Next, we are going to analyze the stability of the equilibrium . Obviously, the equilibrium O as a saddle node simultaneously exists with the equilibrium when and . We first give the Theorem 7 and then prove it.
Theorem 7. If and , the unique positive equilibrium point is locally stable. If and , the unique positive equilibrium is unstable and there exists at least one limit cycle in a small neighborhood of .
Proof. We substitute
into (
10) to derive
which yields
It follows from the inequality
that
.
It is difficult to check the sign of , so we set .
When
then
, which means the positive equilibrium
is locally stable. When
then
, which means the positive equilibrium
is unstable. In addition, the solution of system (
9) is eventually uniformly bounded in
, so there exists at least one limit cycle in a small neighborhood of
.
When
then
, which implies a Hopf bifurcation may occur in system (
9). □
Next, we are going to talk about the situation of i.e., .
Theorem 8. If and condition (22) holds, a supercritical Hopf bifurcation occurs near when ξ passes through 0, where . Proof. Let us now verify the existence of a Hopf bifurcation under the condition
. Suppose
we have
. We shift this point
of system (
9) to the origin. Then, system (
9) can be written as
Setting
, the eigenvalues of
are
and
. Using the nonsingular transformations
,
, and also denoting
by
, respectively, system (
21) becomes
where
and
are
functions and satisfy
.
Applying the formula for the Lyapunov number of Andronov et al. [
30], we have the first Lyapunov number
of the equilibrium point
of system (
9) as follows
If
holds, then
. The Theorem 8 is proved. □
Applying the Friedrich Theorem of Hopf bifurcation as before, we can know that there exists a unique stable limit cycle near
when
and condition (
22) holds.
6. Numerical Simulation
In the following, applying dde23 in matlab, we verify our theoretical results by numerical simulations for system (
9). According to the numerical constraints in the theorems, we first determine the values of
b,
r and
c. Combined with numerical simulations, and try several times to find the value of
that meets the conditions. For the value of
, it is easy to calculate with the given conditions. All figures correspond to the cases in the table above.
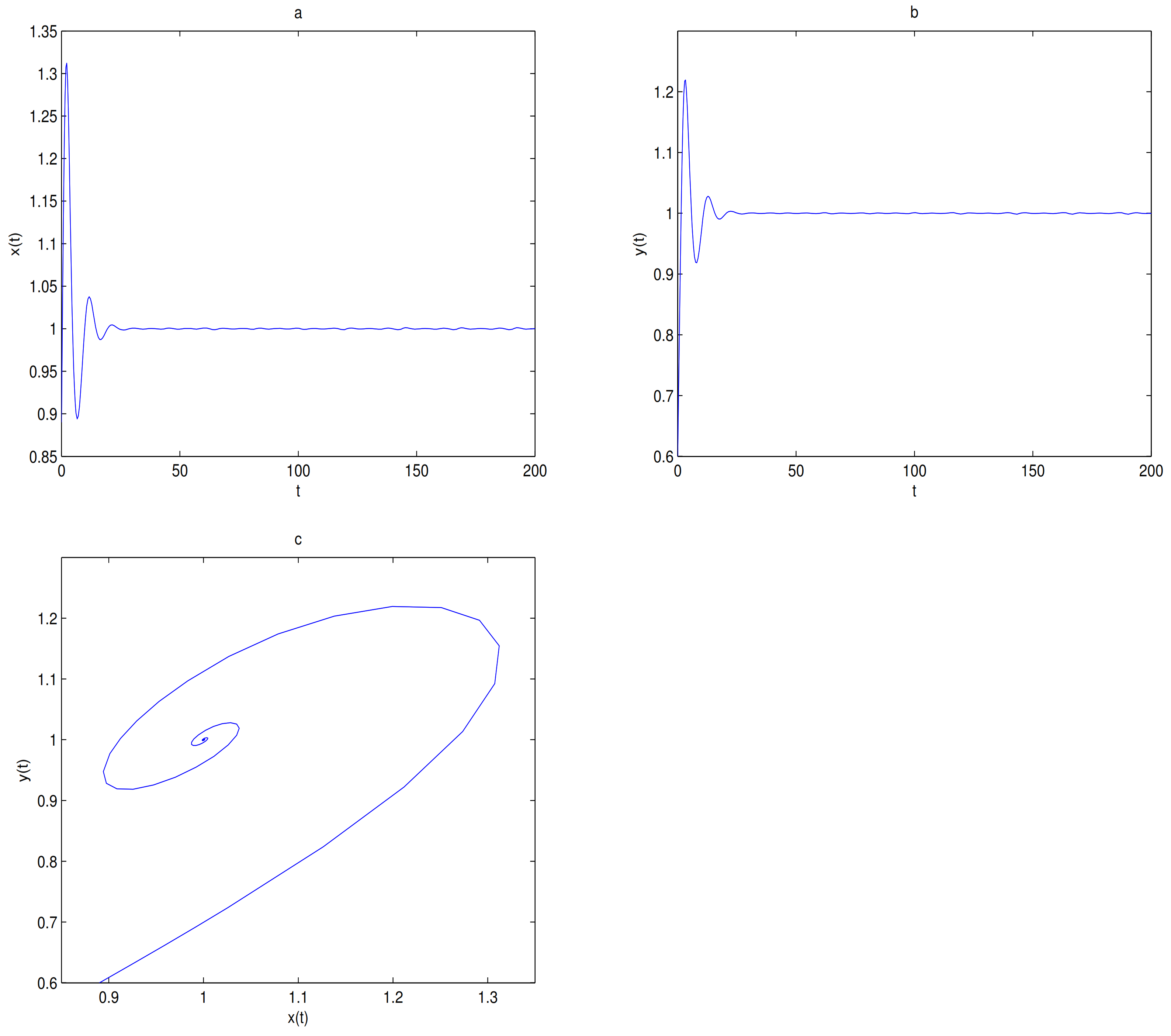
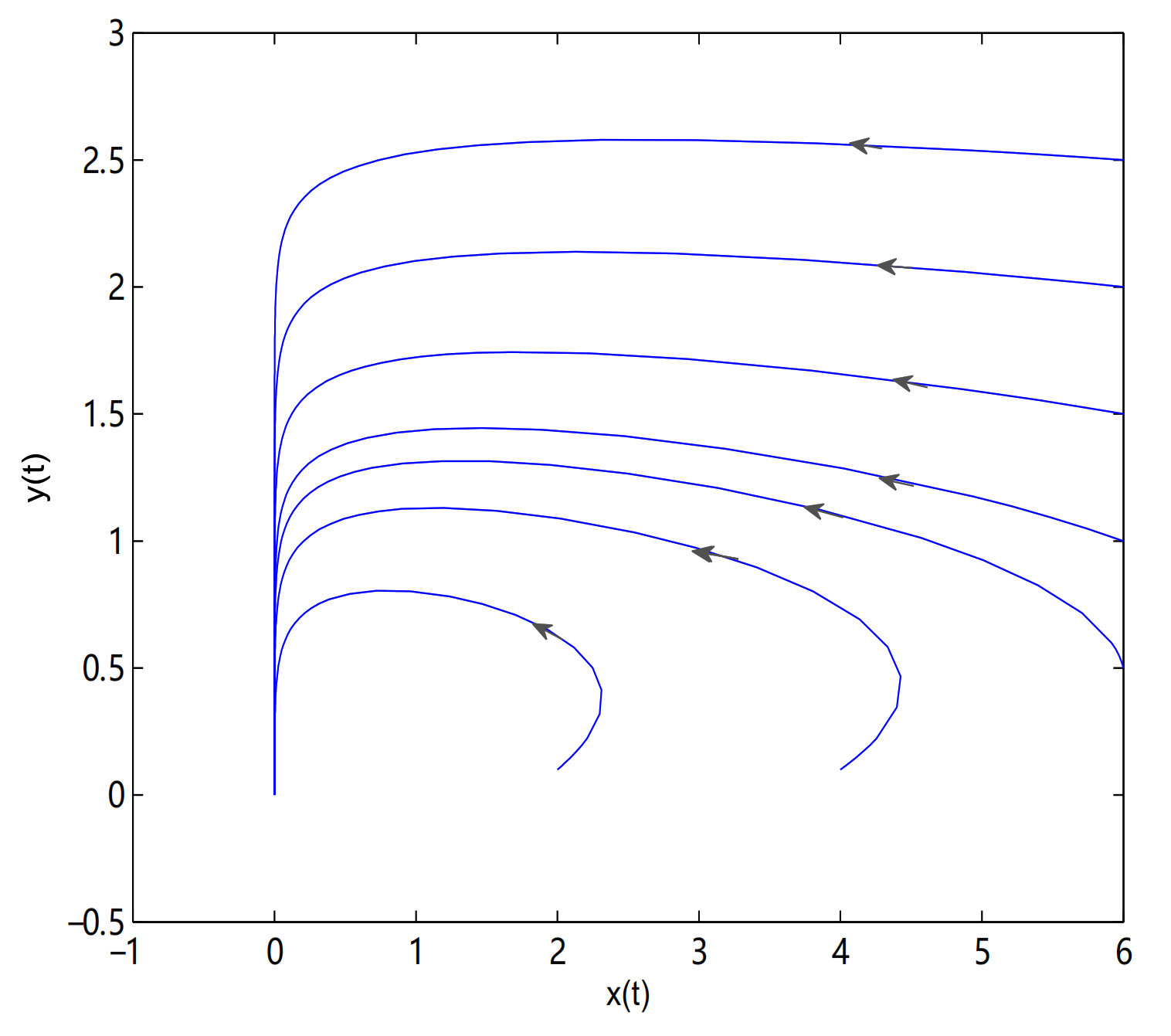
For Theorems 1–3, let
and
,
O is always a saddle.
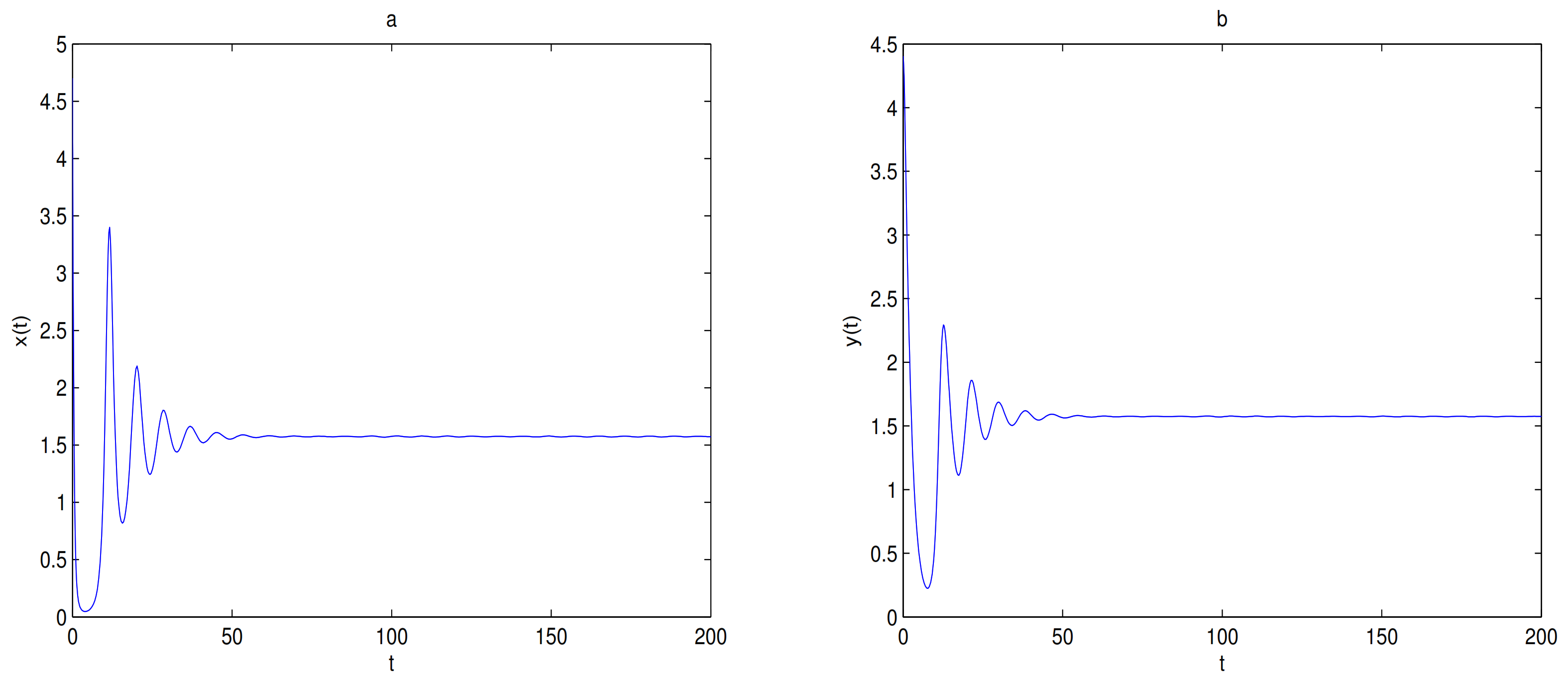
is globally asymptotically stable when
(see
Figure 5) and there exists one limit cycle in
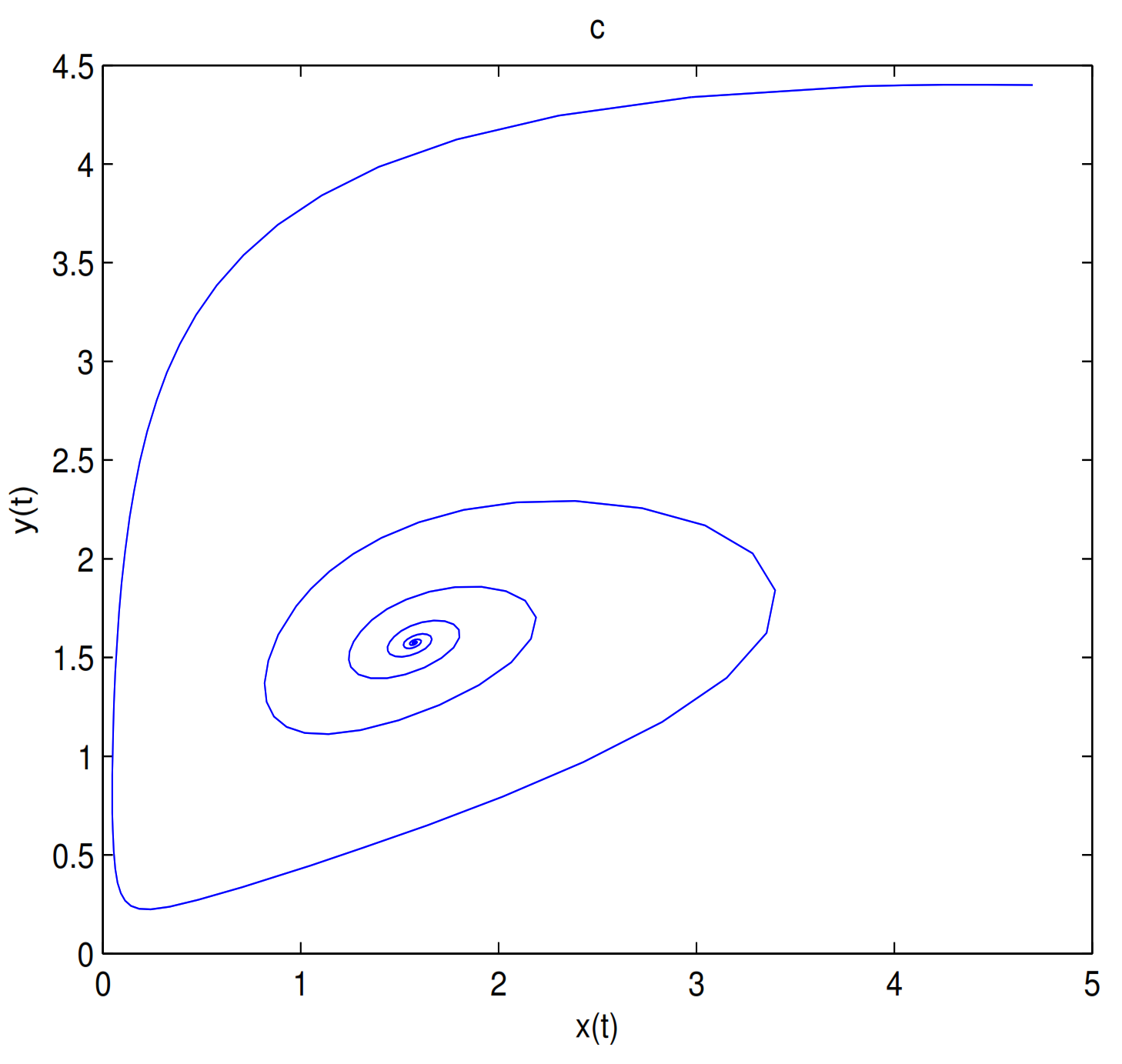
when
, in here
(see
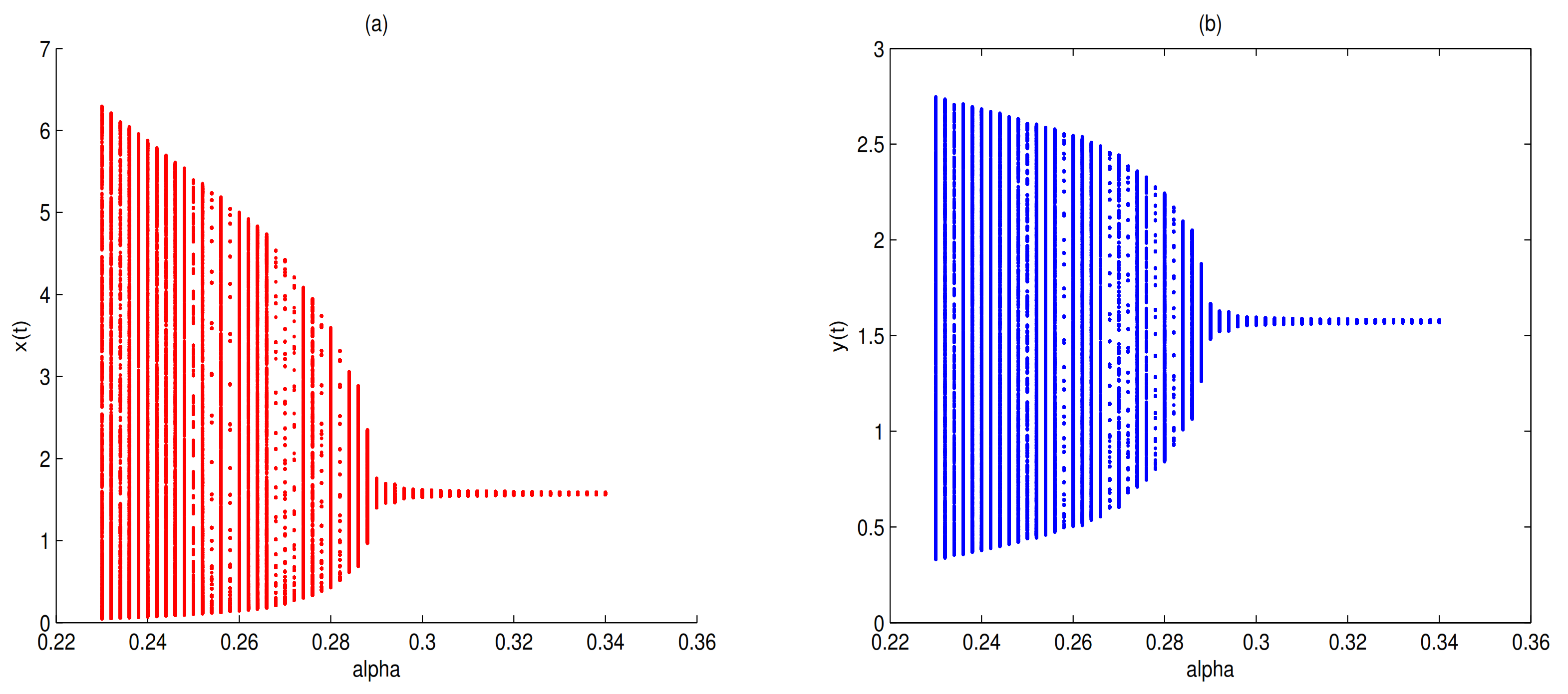
Figure 6). Choosing
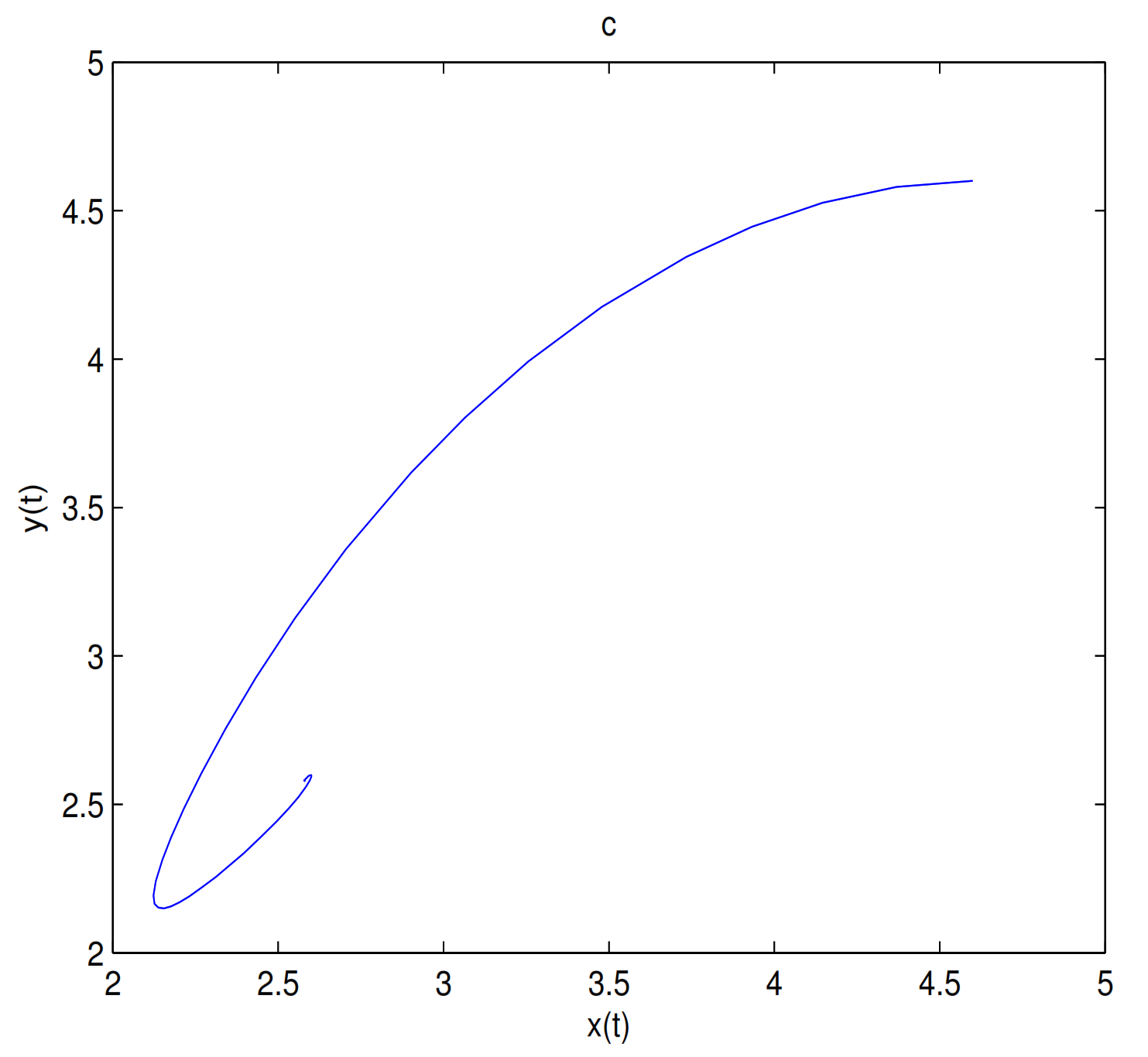
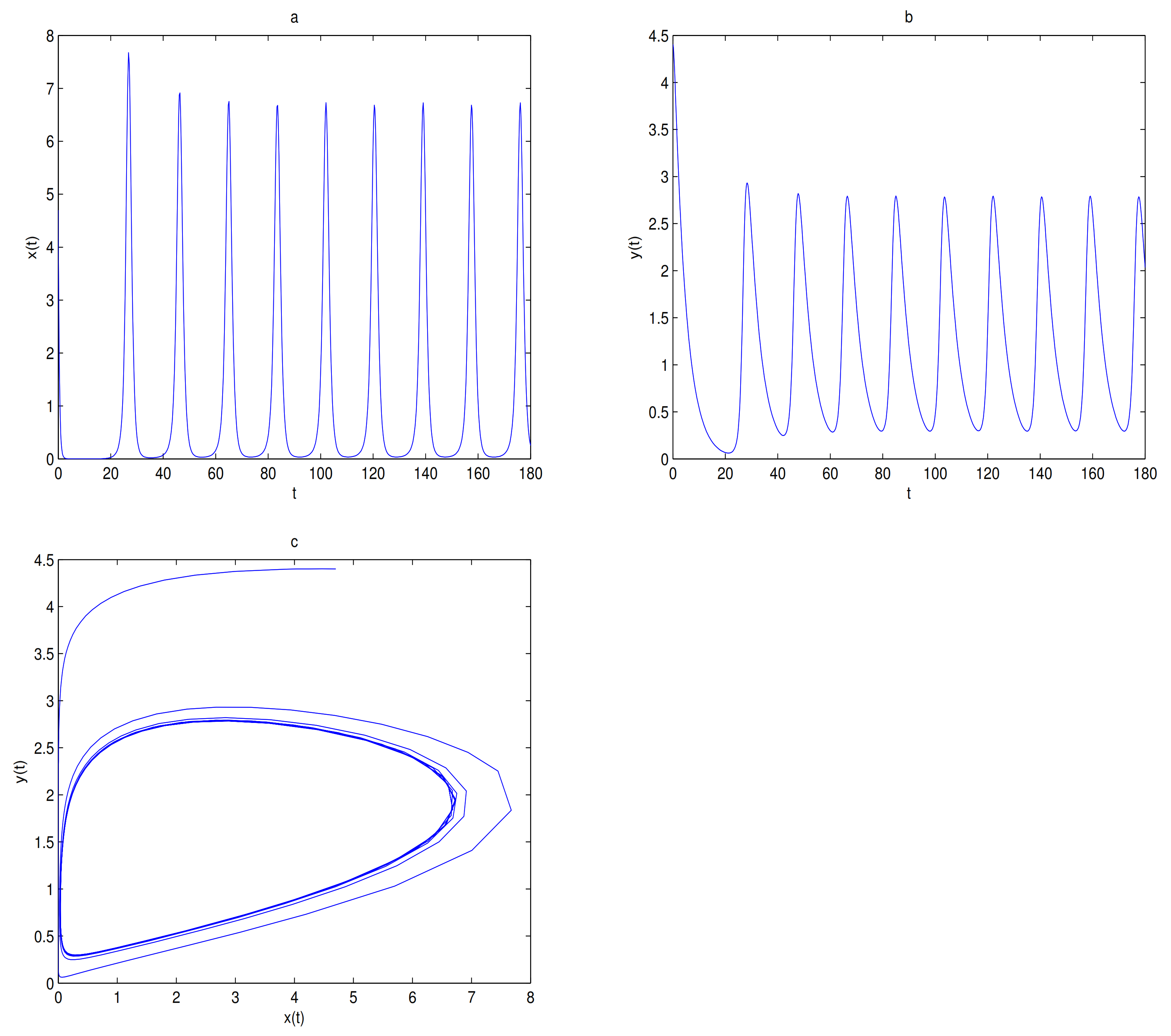
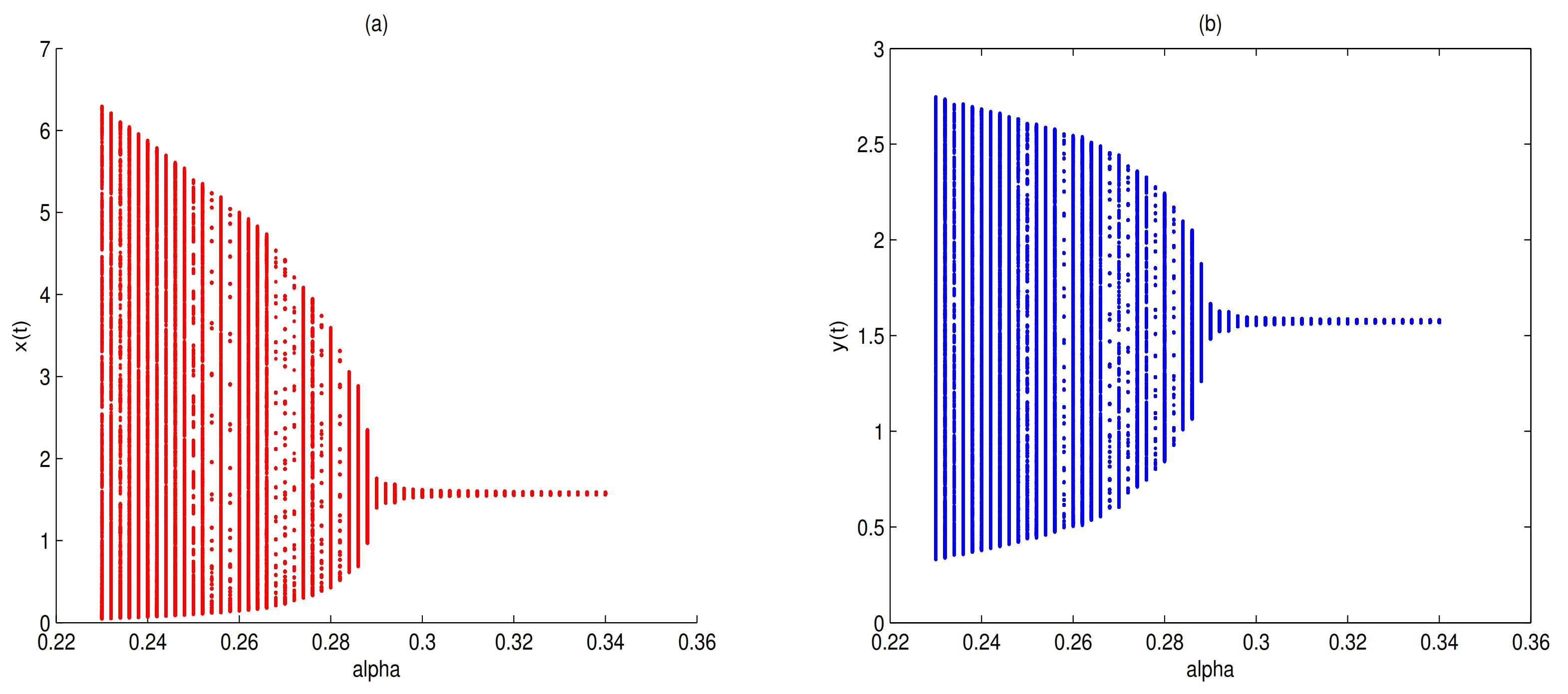
as a bifurcation parameter, a supercritical Hopf bifurcation occurs near
when
passes through 0.288, in here
(see
Figure 7).
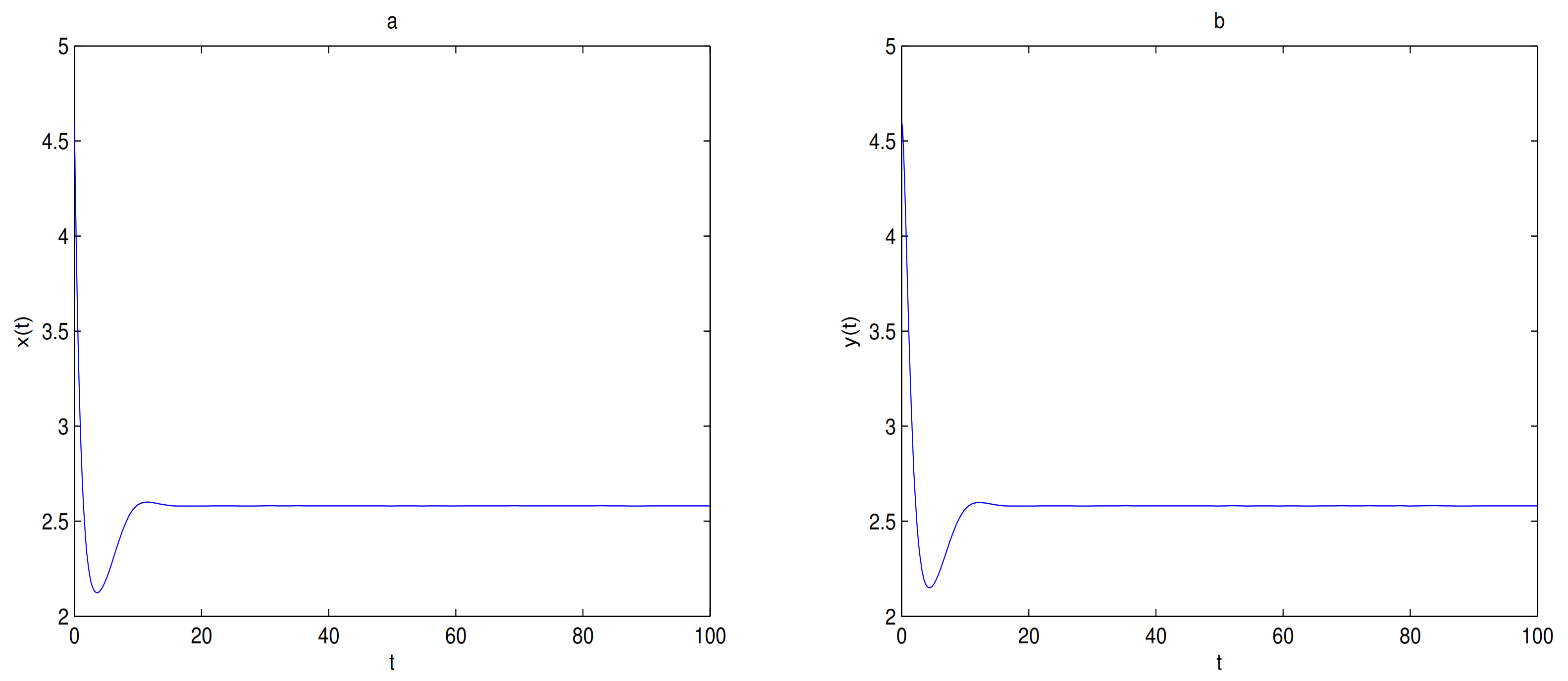
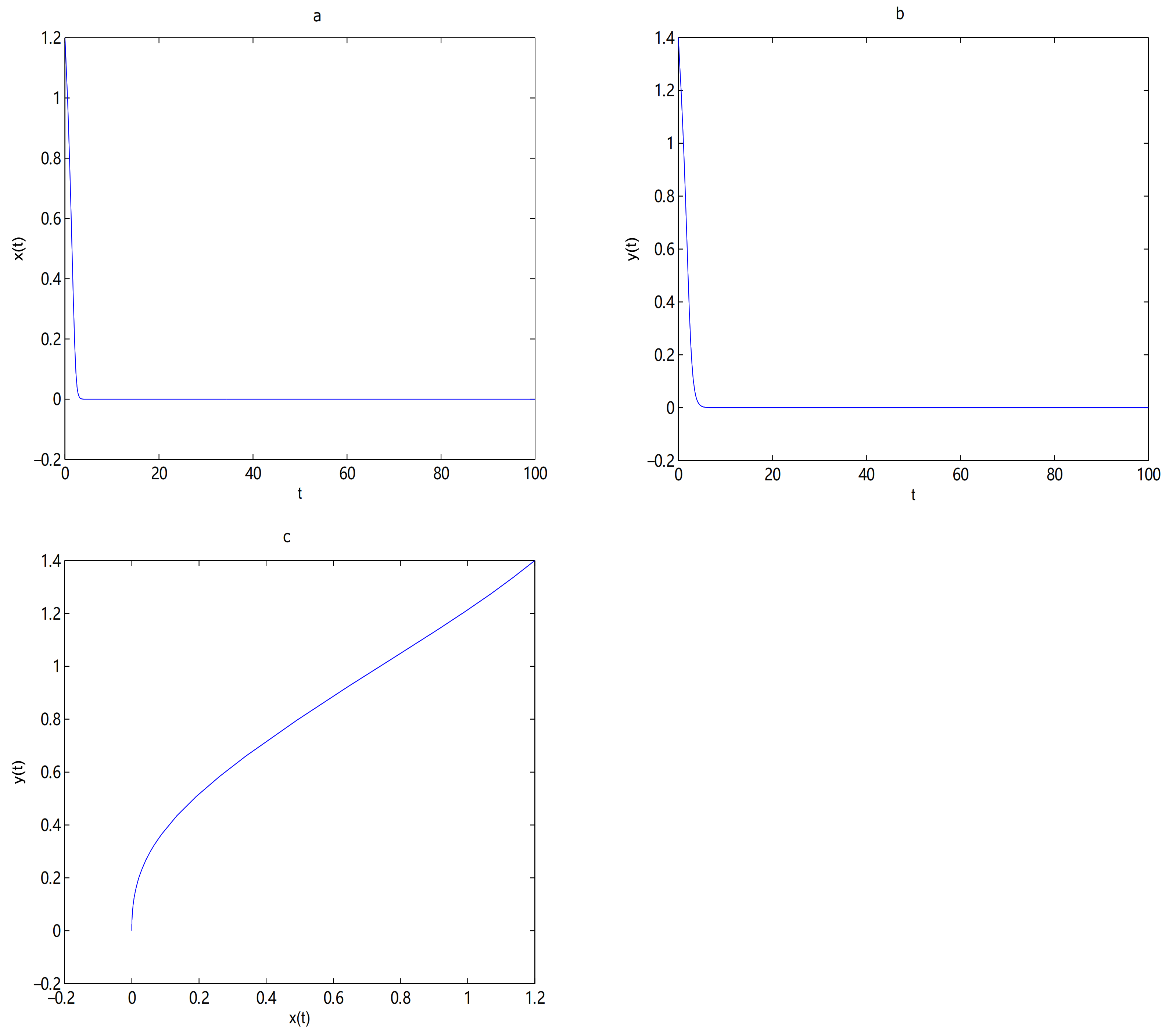
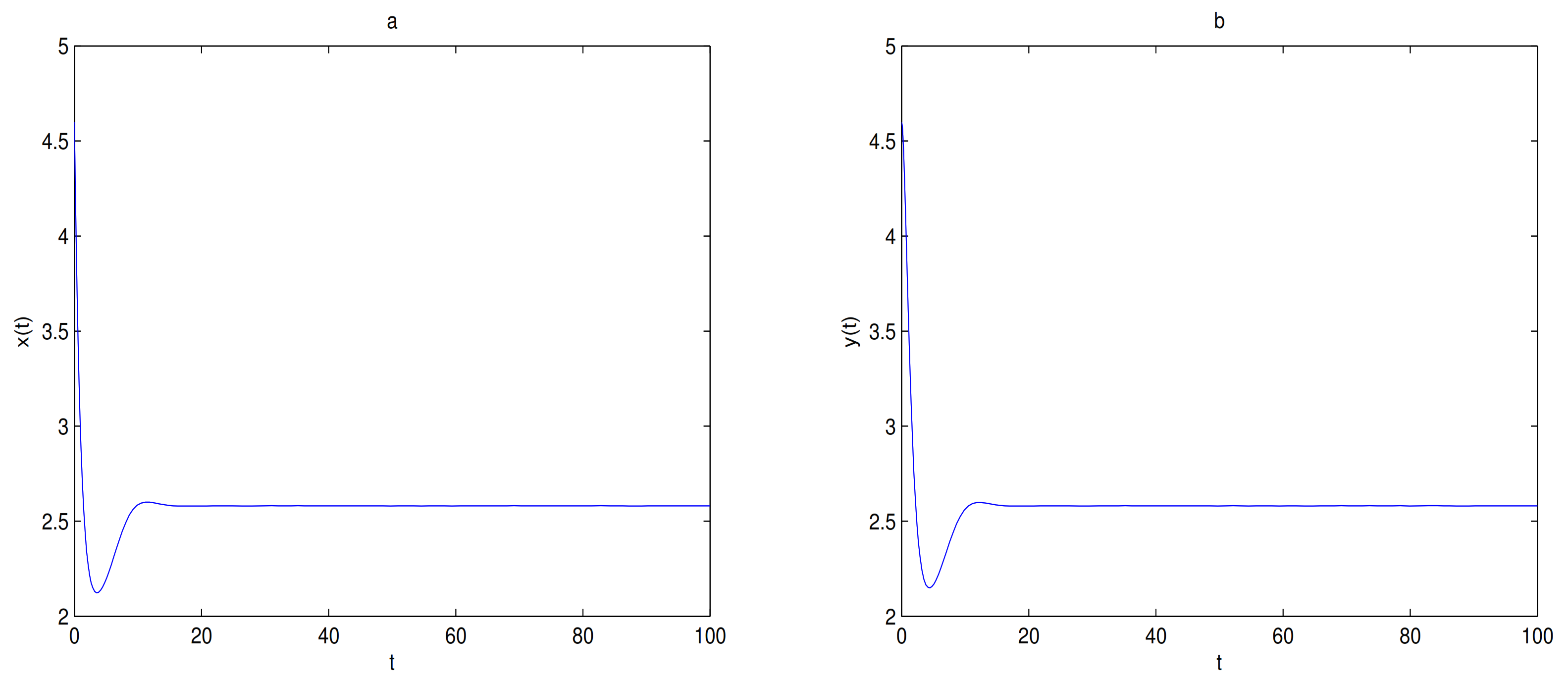
For Theorem 4, let
and
,
O is the only equilibrium point of system (
9) at this time, and
O is globally asymptotically stable. In here initial values are (6, 2.5), (6, 2), (6, 1.5), (6, 1), (4, 0.1) and (2, 0.1), we see that all trajectories tend to
O (see
Figure 8). Let
and
,
O is always locally stable, and
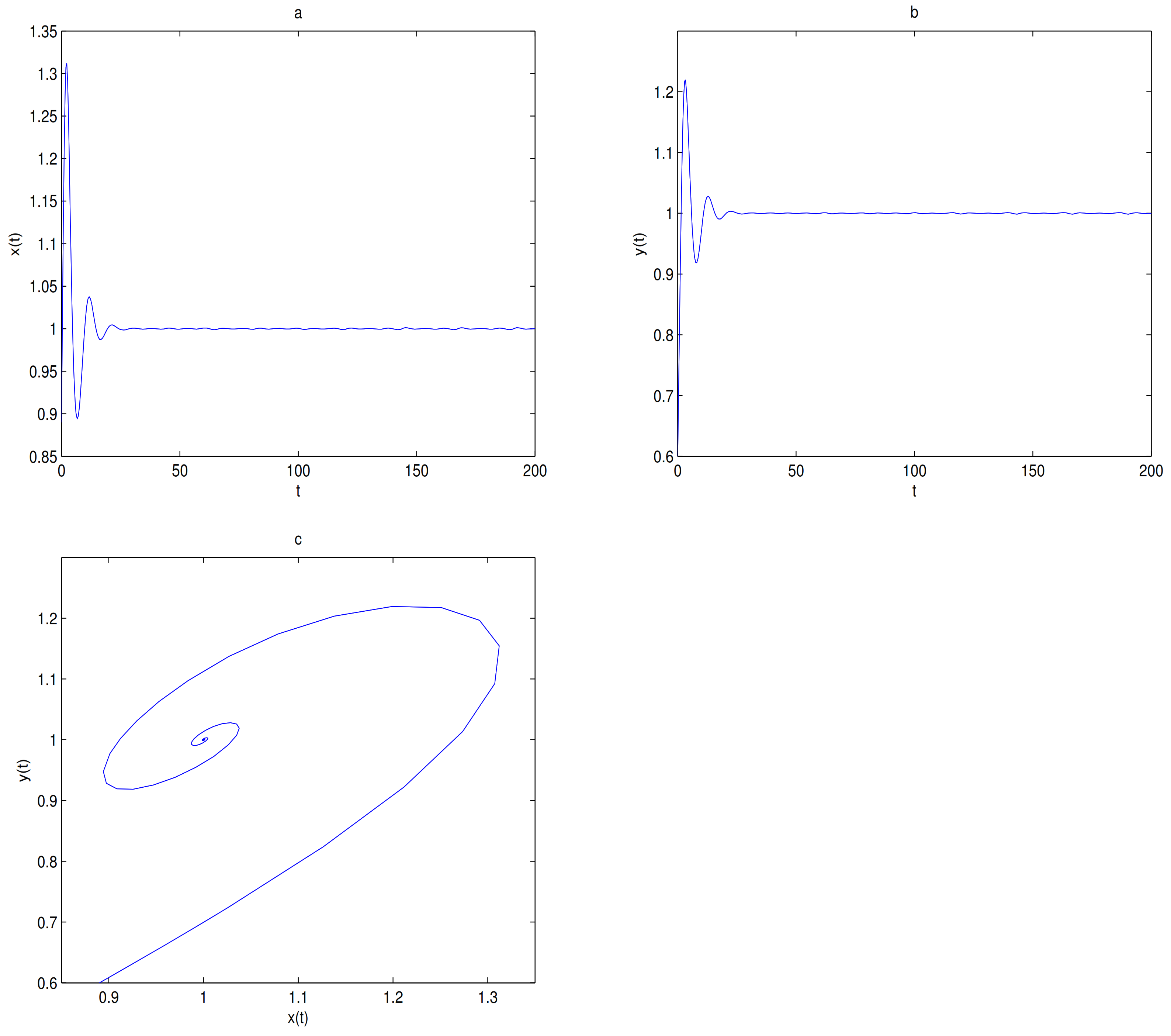
is always a saddle.
is locally stable when
. the trajectory tends to
when the initial value is selected in the attraction domain of
, for example,
(see
Figure 9). The trajectory tends to
O when the initial value is selected in the attraction domain of
O, for example,
(see
Figure 10).
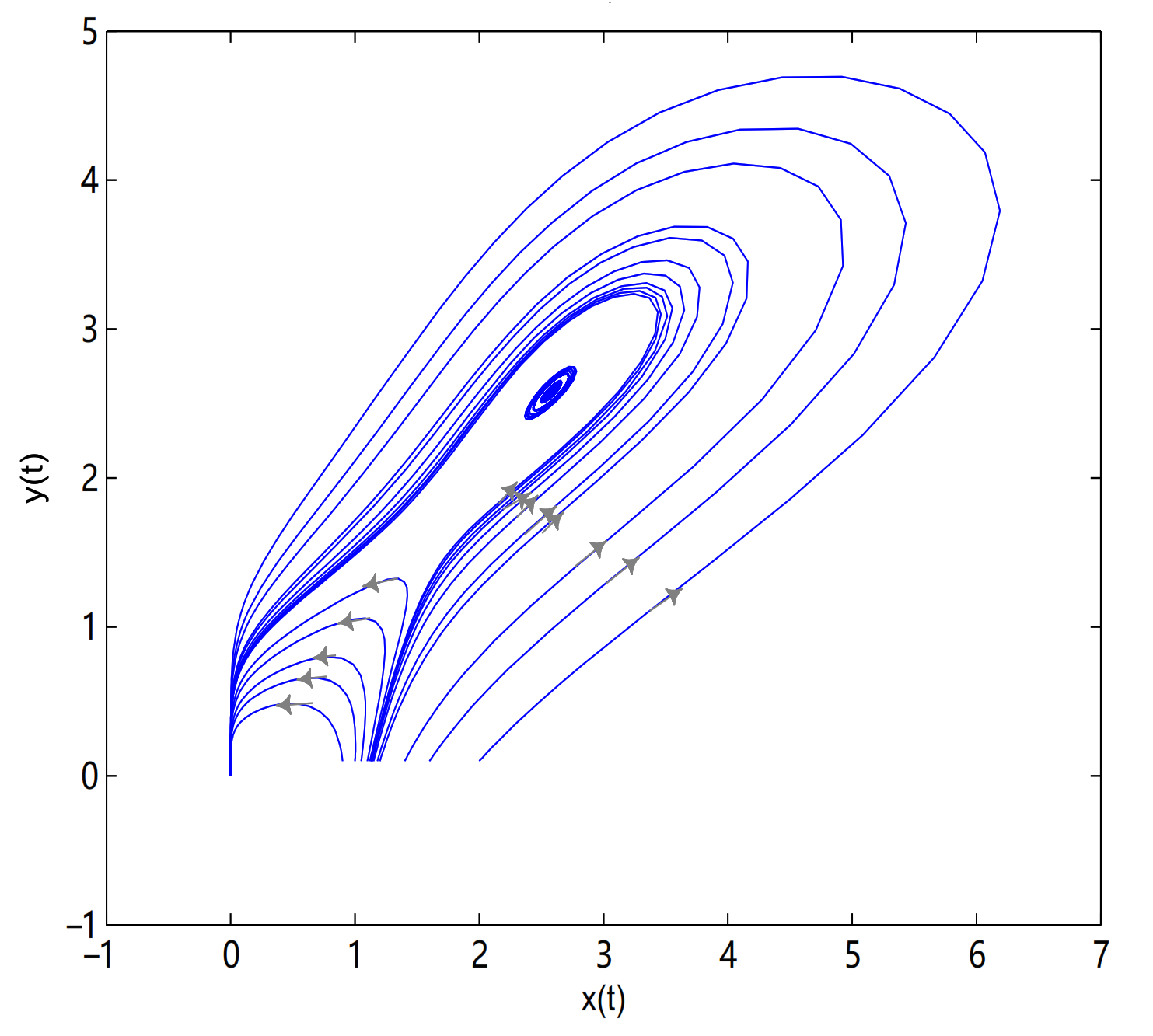
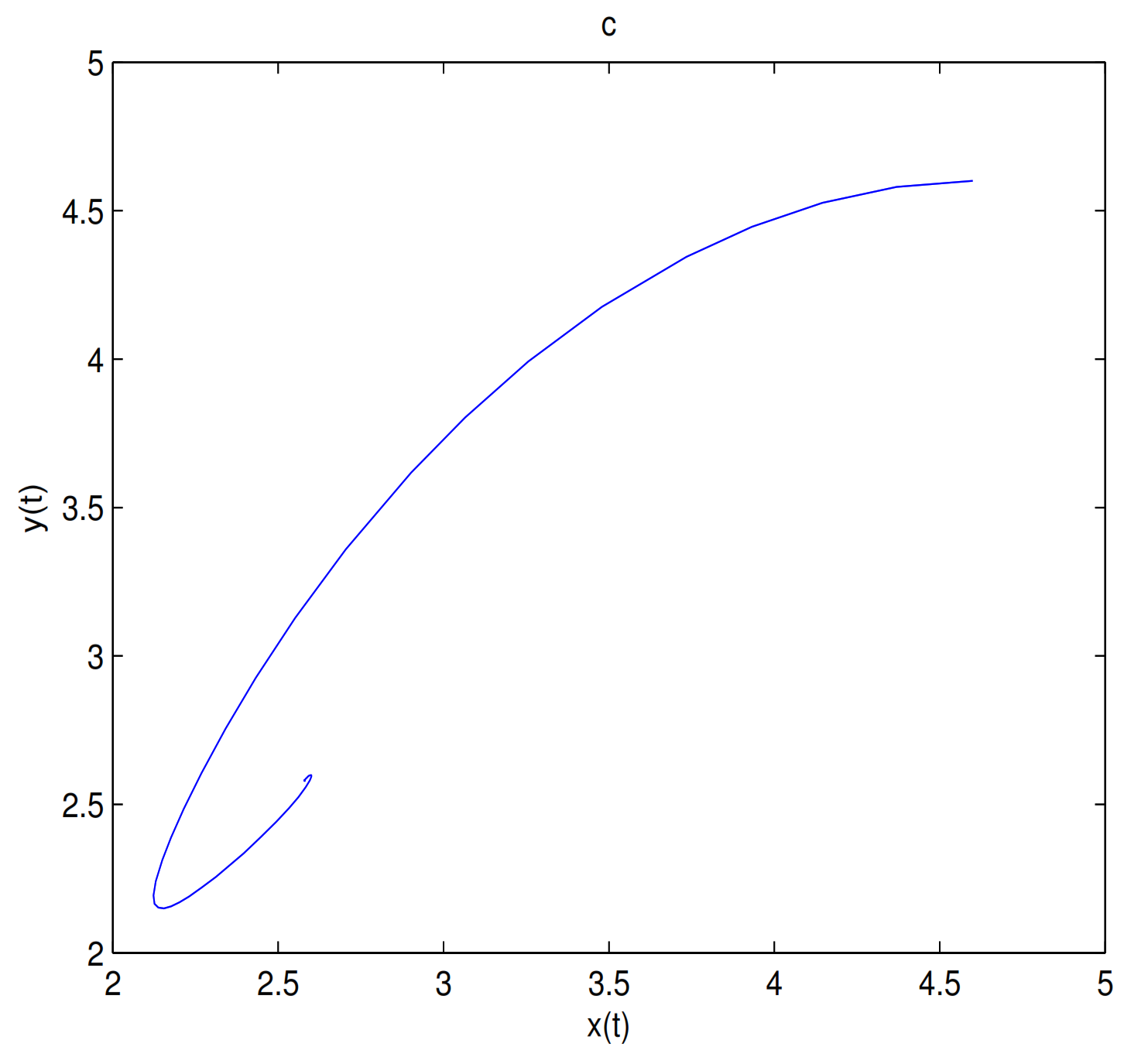
is a center-type equilibrium point when
, in here initial values are (2.57, 2.57), (2.42, 2.42), (2.1, 0.1), (1.6, 0.1), (1.4, 0.1), (1.2, 0.1), (1.18, 0.1), (1.15, 0.1), (1.14, 0.1), (1.135, 0.1), (1.133, 0.1), (1.131, 0.1), (1.13, 0.1), (1.1, 0.1), (1.05, 0.1), (1, 0.1) and (0.9, 0.1) (see
Figure 11).
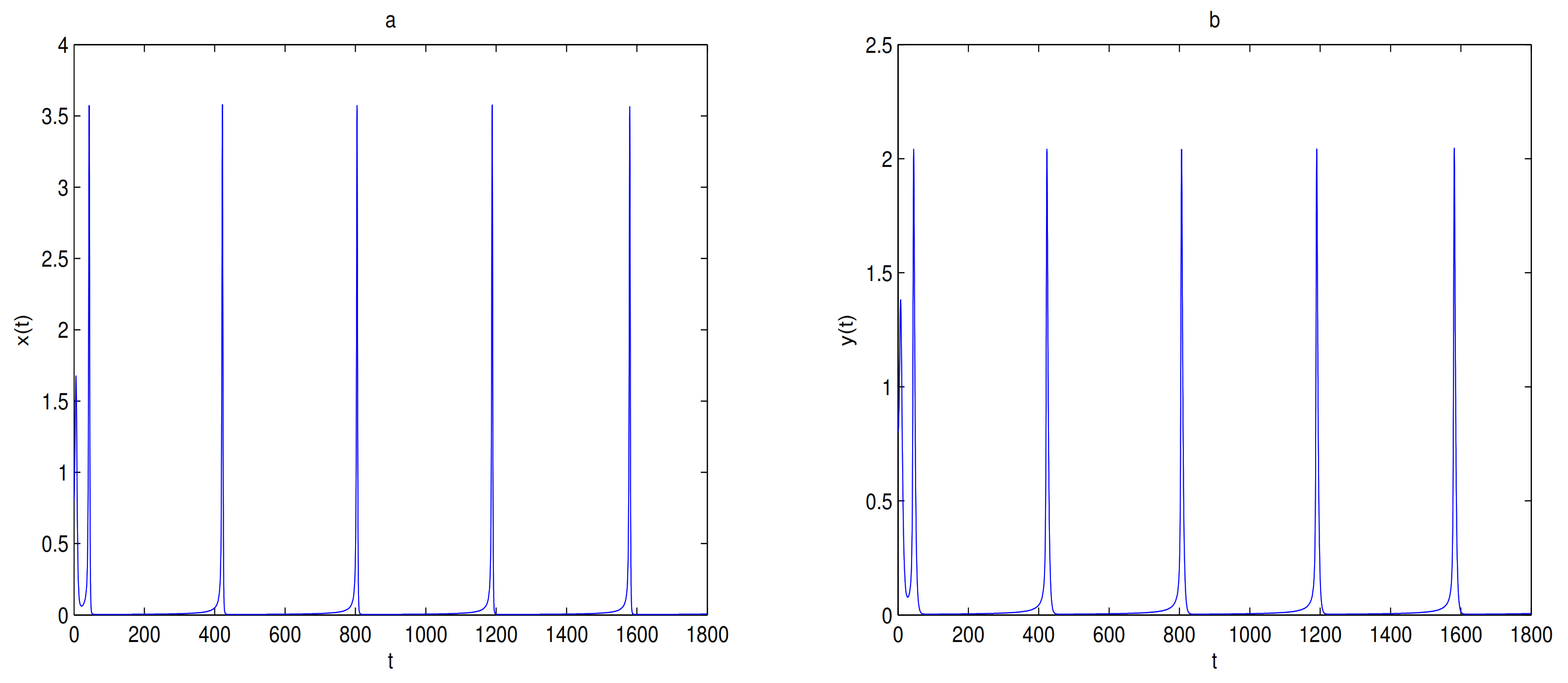
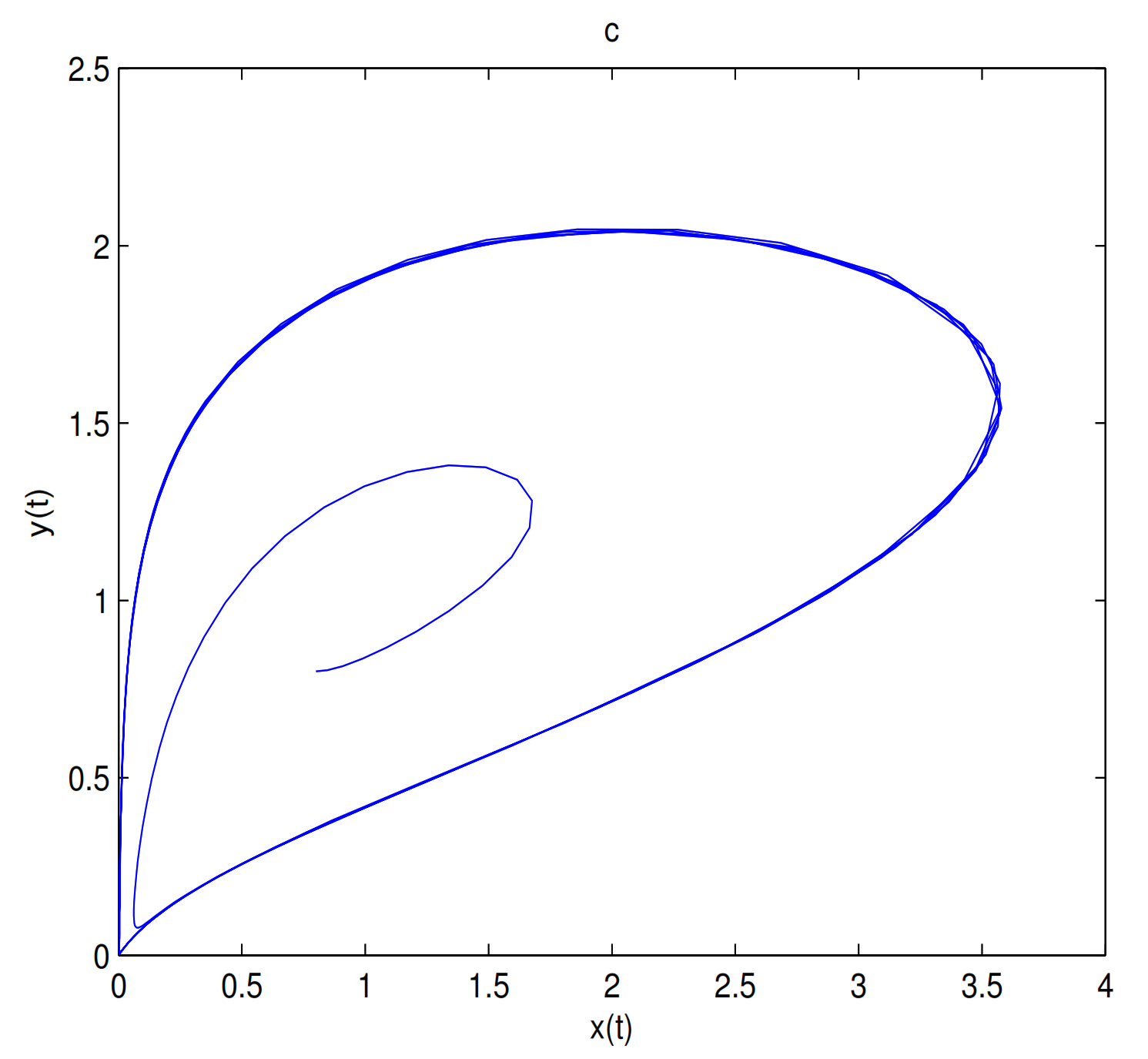
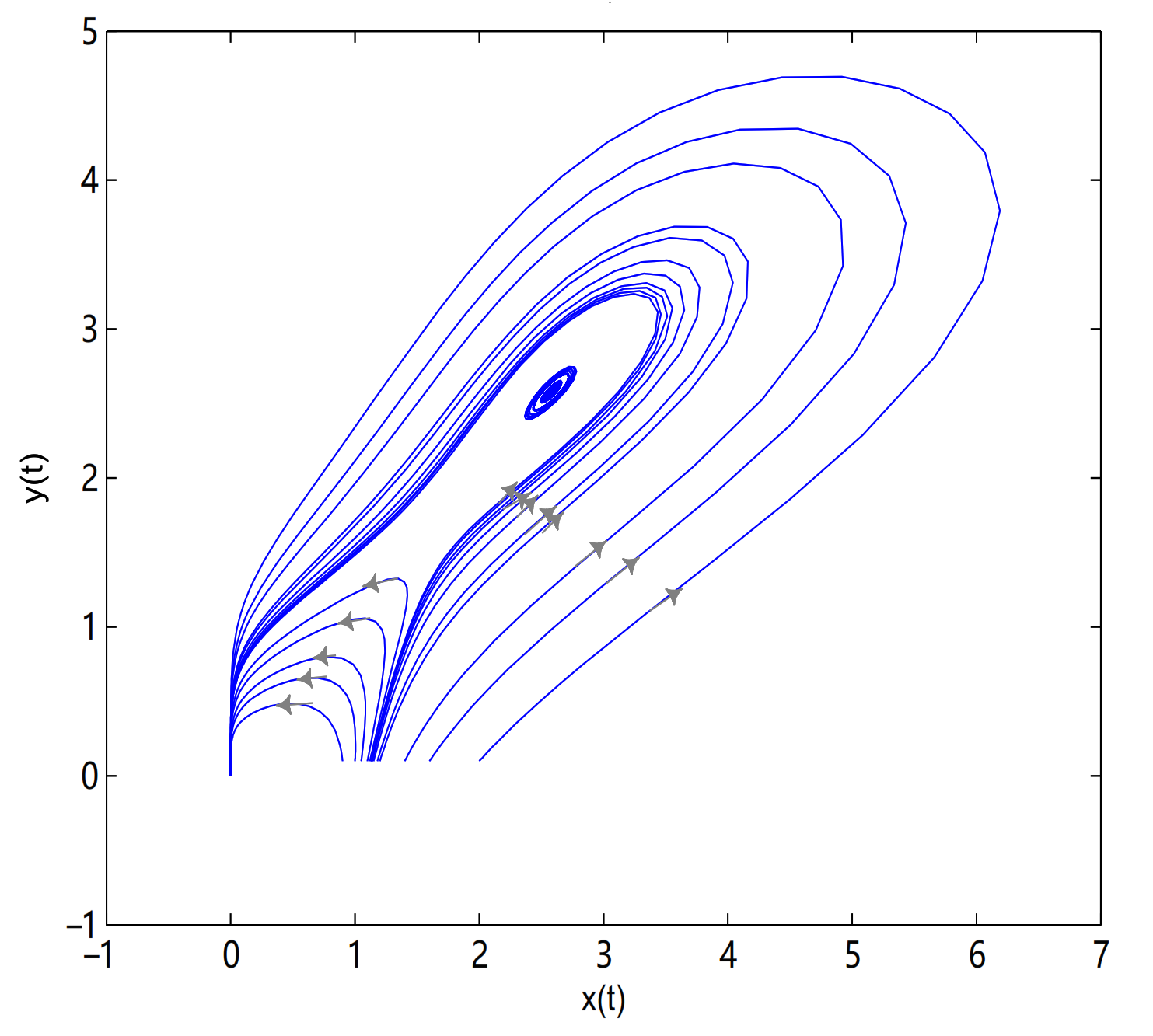
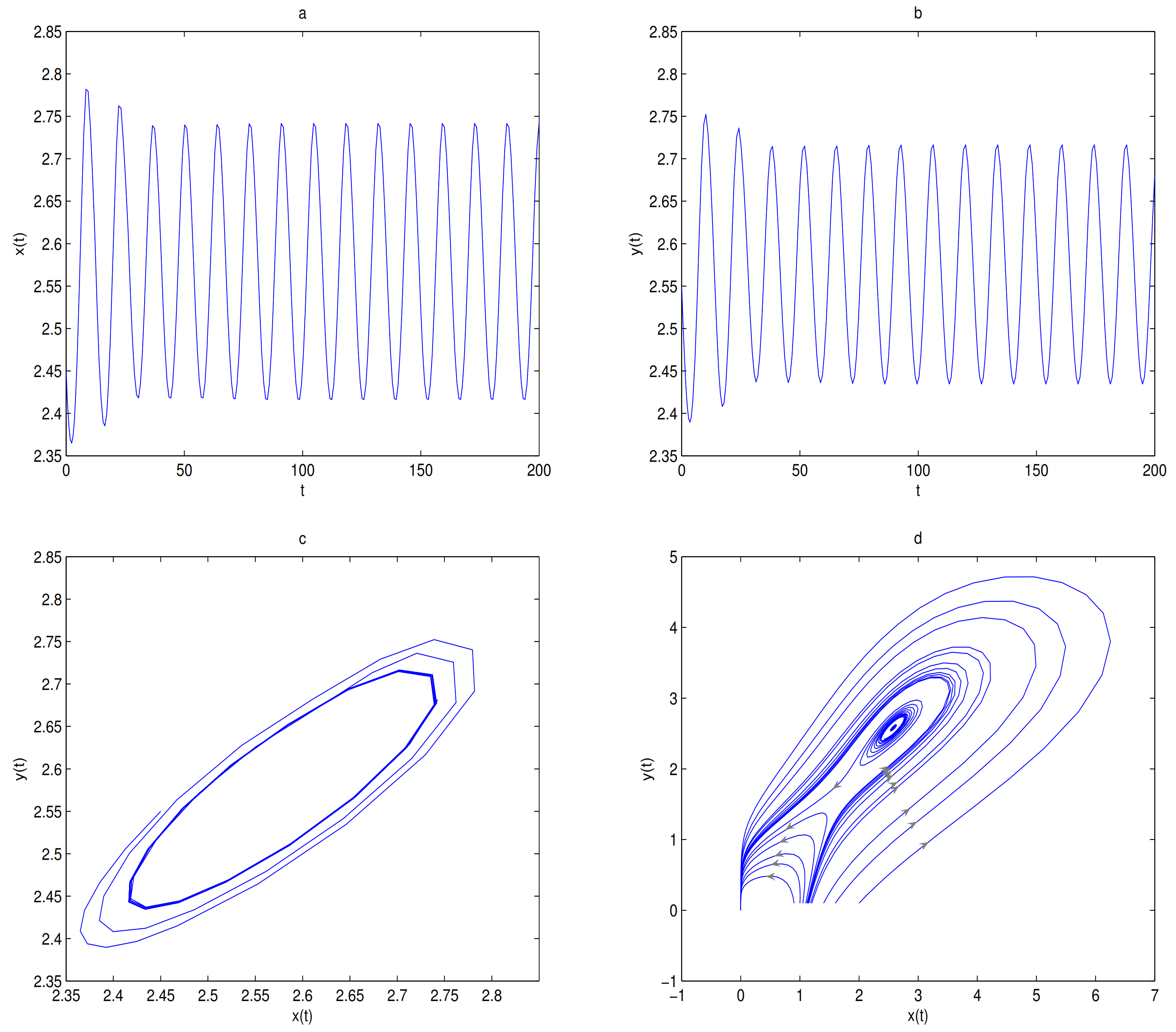
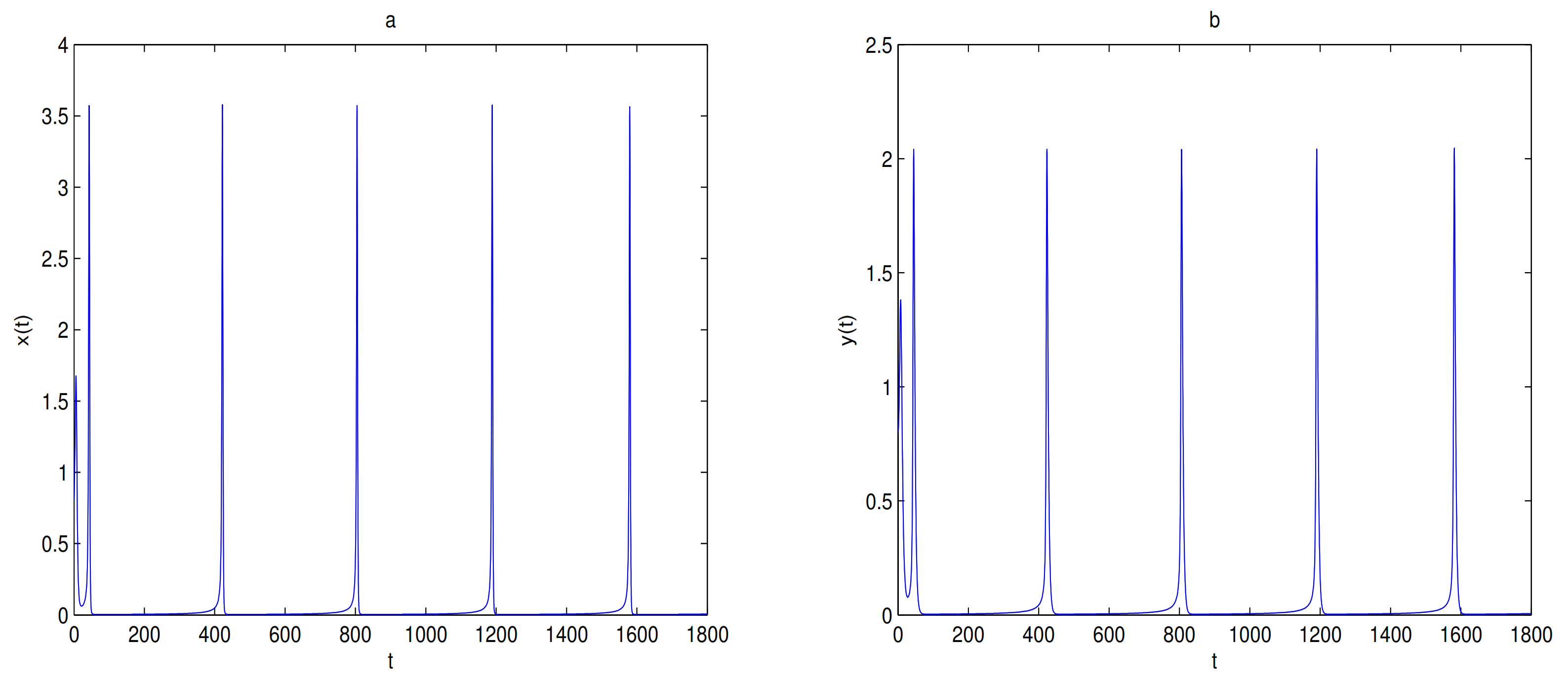
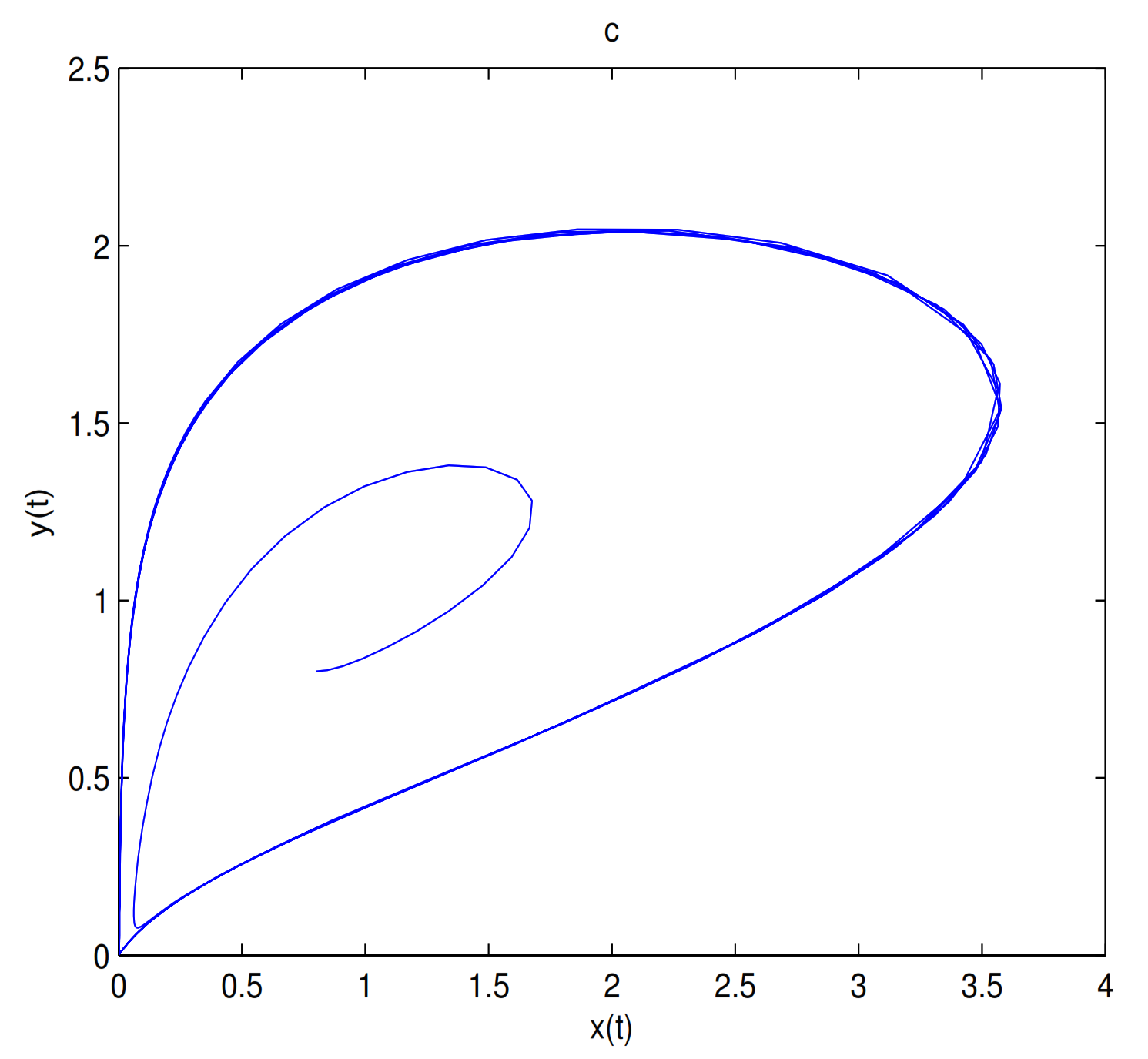
is unstable when
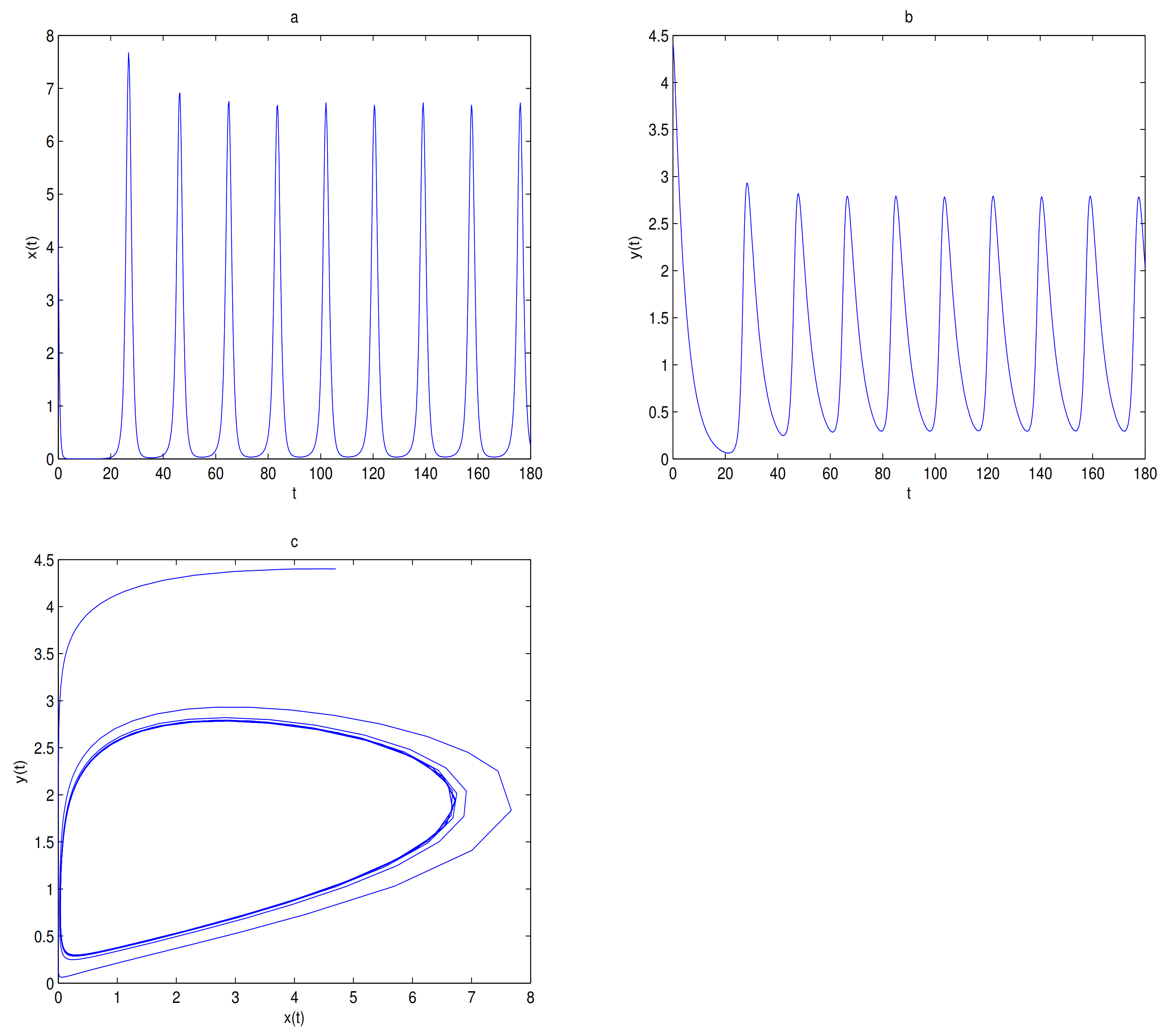
, and a limit cycle is generated by Hopf supercritical bifurcation in the neighborhood of
. We choose
as initial value (see
Figure 12c). We choose the same initial values as in
Figure 11, and we see that the trajectory forms a limit cycle with (2.57,2.57) as the initial value, and everything else tends to
O (
Figure 12d).
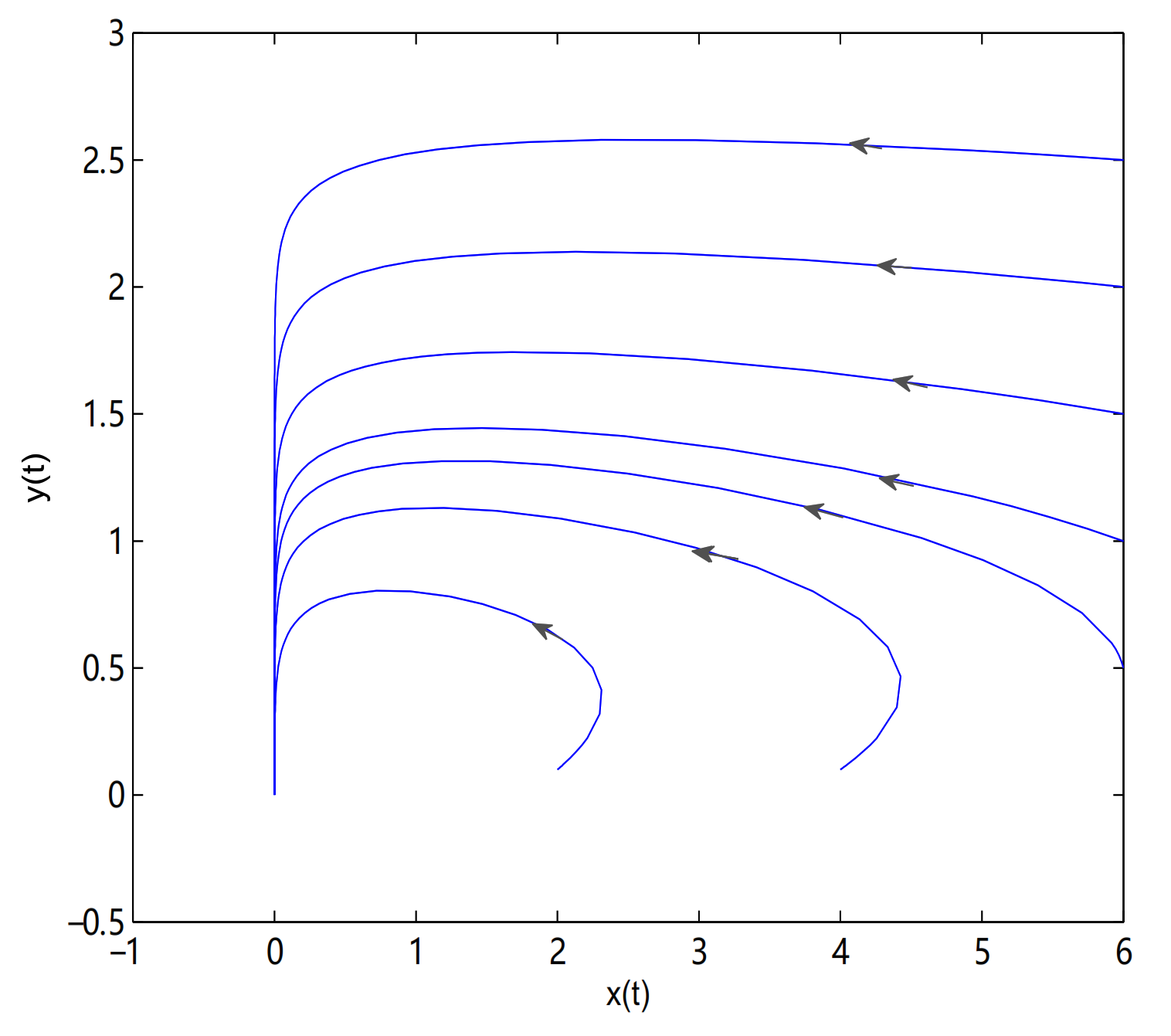
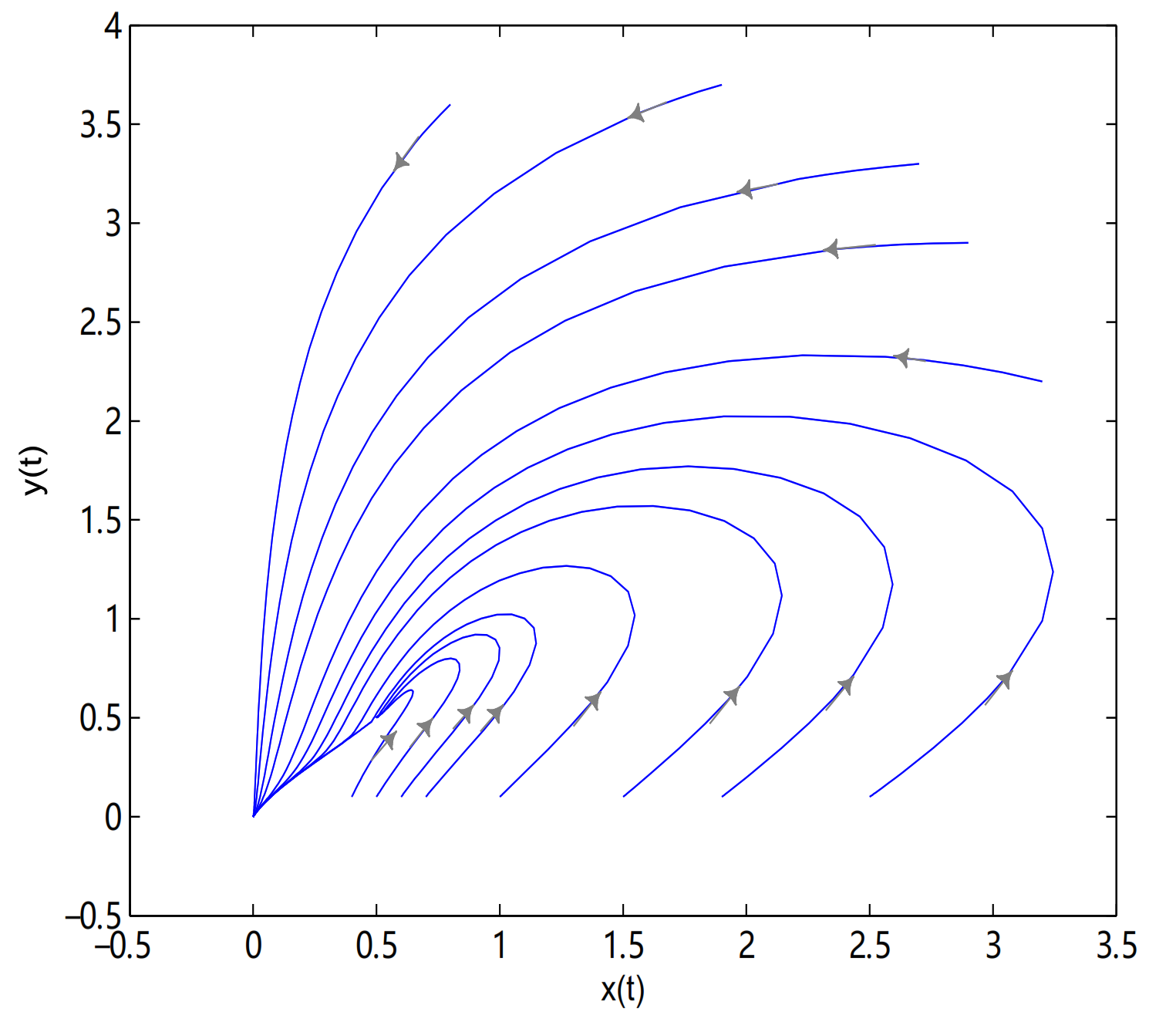
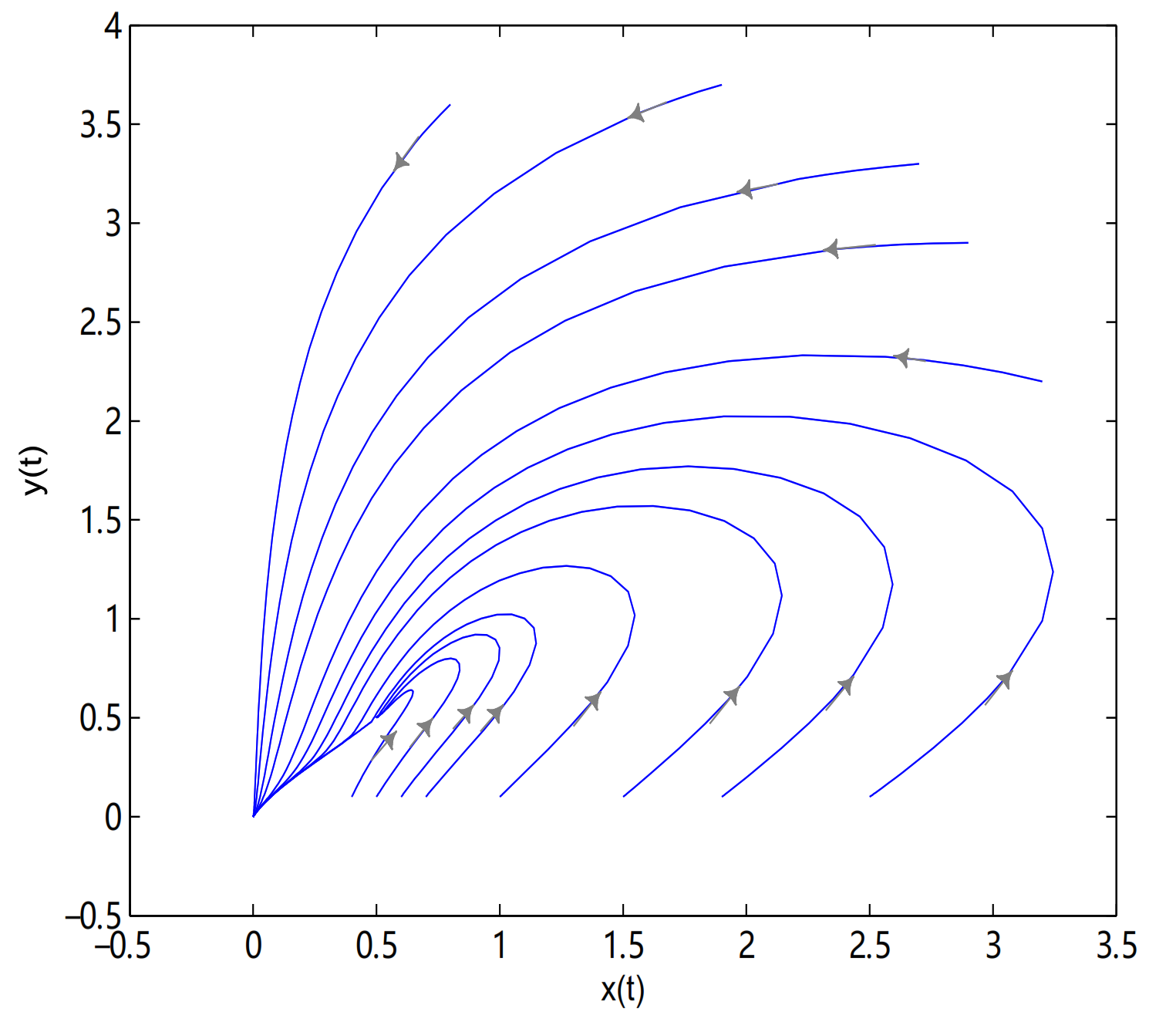
For Theorem 5, let
,
O is always globally asymptotically stable, and
are merged into a single equilibrium
. Meanwhile,
is a saddle node when
, in here initial values are (2.7, 3.3), (2.9, 2.9), (0.8, 3.6), (3.2, 2.2), (1.9, 3.7), (2.5, 0.1), (1.9, 0.1), (1.5, 0.1), (1, 0.1), (0.7, 0.1), (0.6, 0.1), (0.5, 0.1) and (0.4, 0.1) (see
Figure 13).
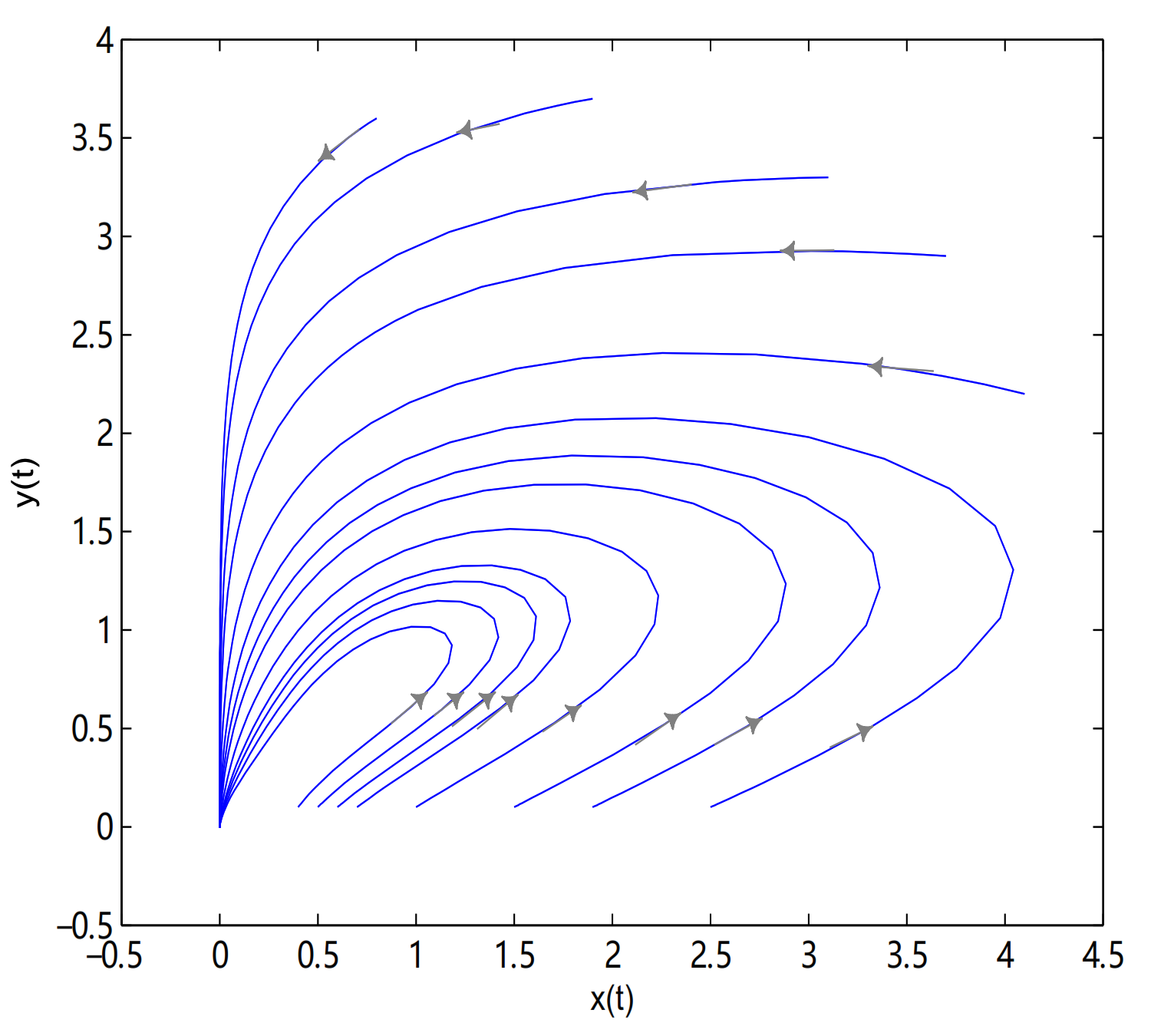
is a degenerate critical point when
, in here initial values are (3.1, 3.3), (3.7, 2.9), (0.8, 3.6), (4.1, 2.2), (1.9, 3.7), (2.5, 0.1), (1.9, 0.1), (1.5, 0.1), (1, 0.1), (0.7, 0.1), (0.6, 0.1), (0.5, 0.1) and (0.4, 0.1) (see
Figure 14).
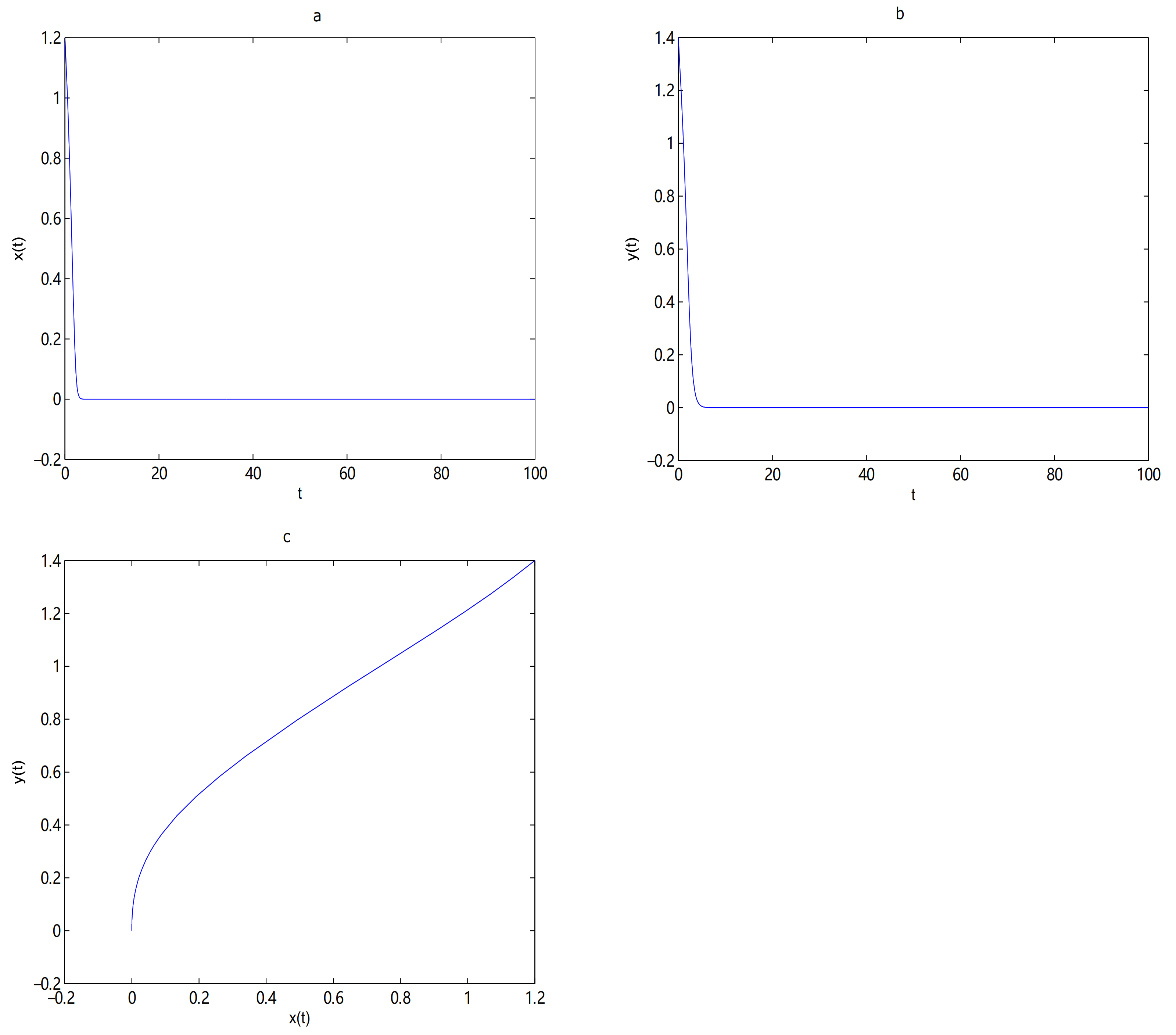
For Theorems 6–8, first, let
and
;
O is always a saddle node.
is unstable and there exists a limit cycle near
when
, the initial value is (0.8, 0.8) (see
Figure 15).
is locally stable when
, and the initial value is (0.89, 0.6) (see
Figure 16). By choosing
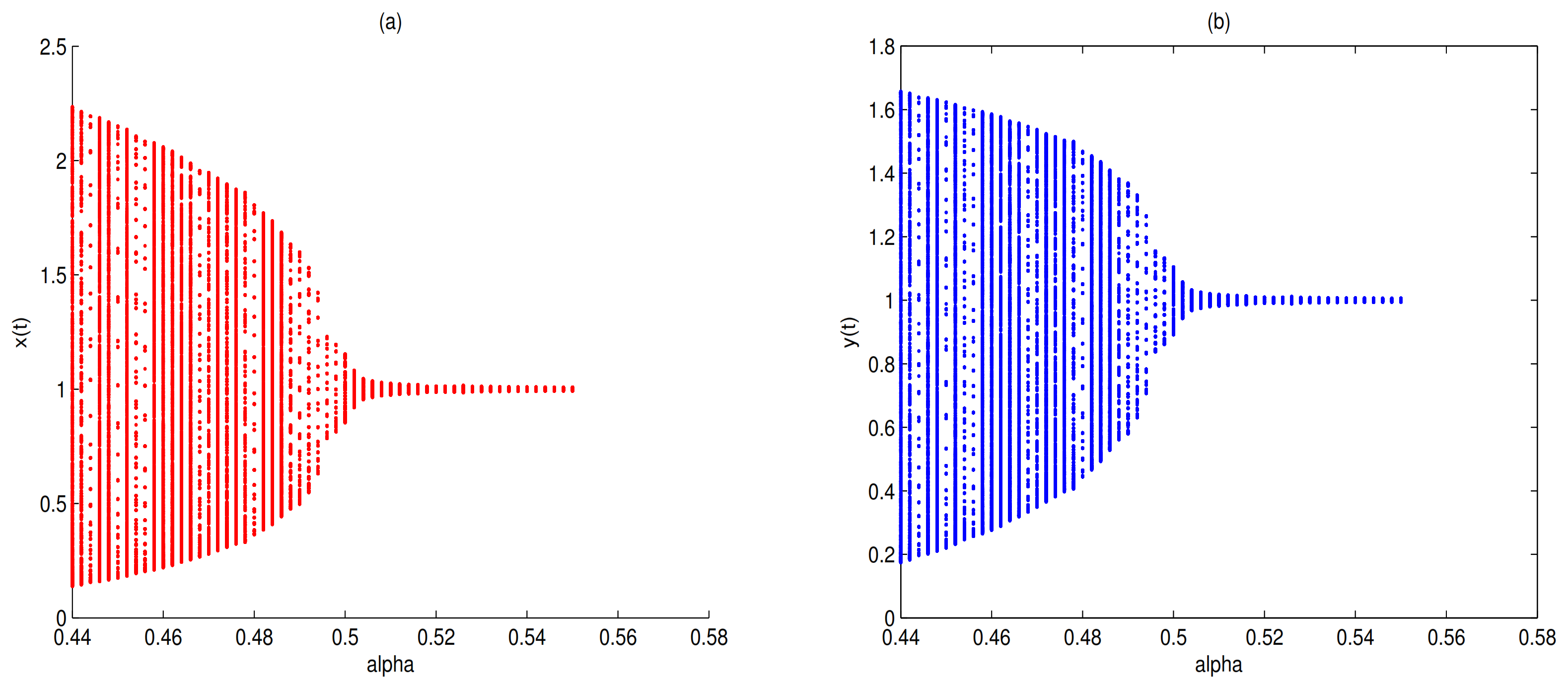
as a bifurcation parameter, a supercritical Hopf bifurcation occurs near
when
passes through 0.5 (see
Figure 17). Let
and
,
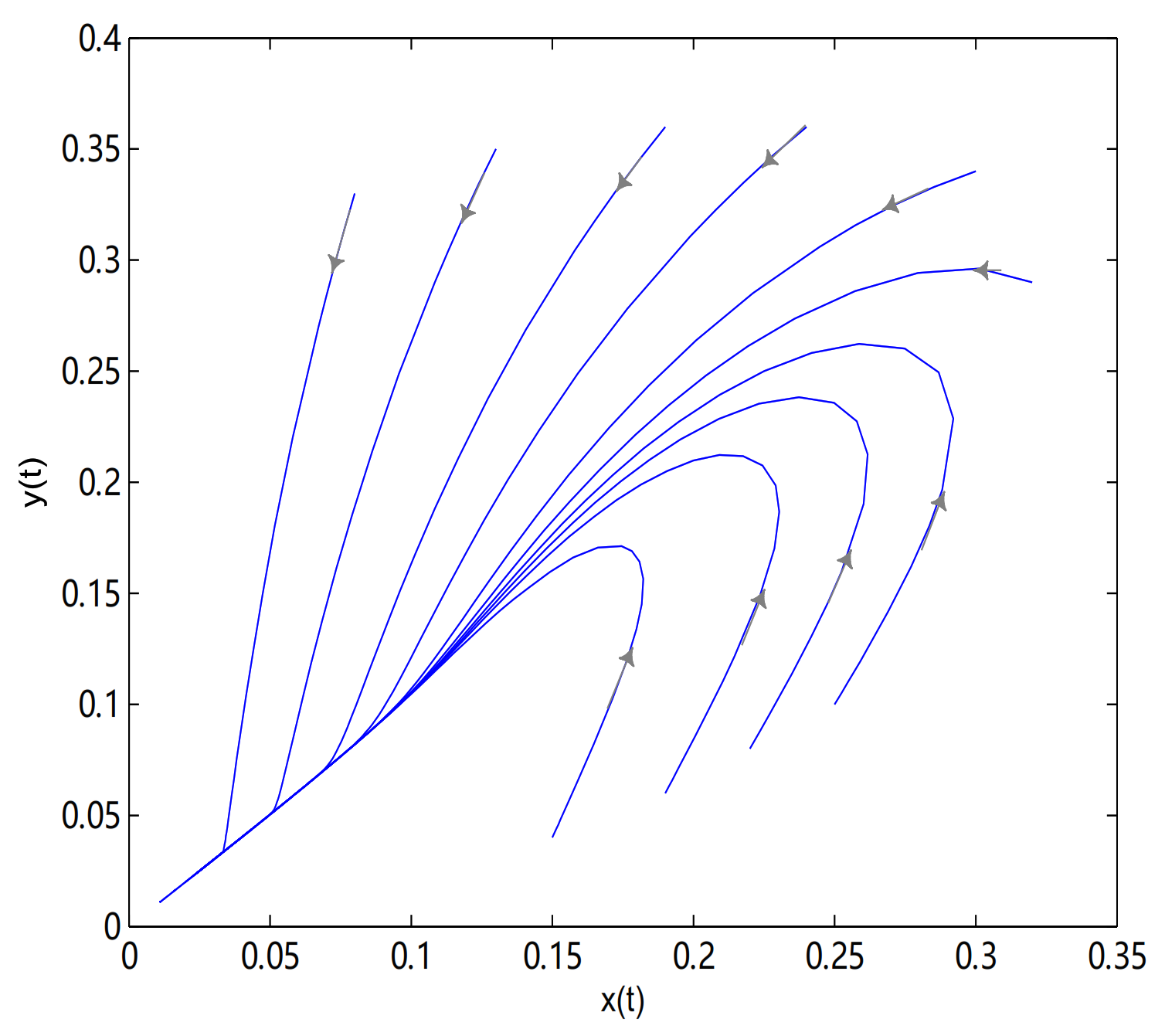
O is an unstable node, in here initial values are (0.28, 0.34), (0.32, 0.29), (0.19, 0.06), (0.25, 0.1), (0.13, 0.35), (0.08, 0.33), (0.24, 0.36), (0.22, 0.08), (0.15, 0.04) and (0.19, 0.36), all trajectories tend to singular line
(see
Figure 18).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}