
Dynamics of Stage-Structured Predator–Prey Model with Beddington–DeAngelis Functional Response and Harvesting

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Local Stability of Origin Equilibrium and Boundary Equilibrium
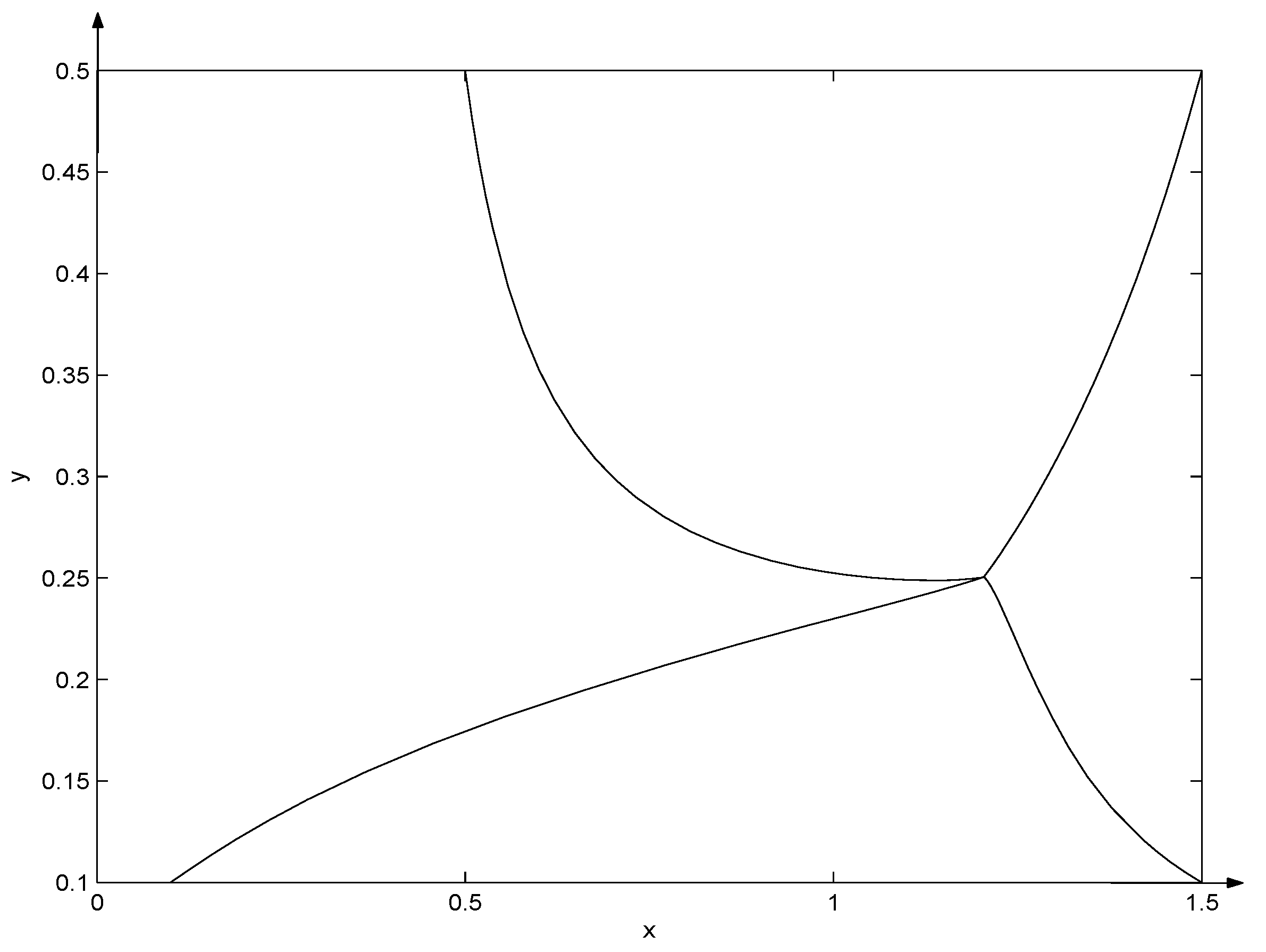
- (i)
- if , then ;
- (ii)
- if , then ;
- (ii)
- if , then .
3. Local Stability of the Positive Equilibrium
3.1. Local Stability of the Positive Equilibrium for
3.2. Local Stability of the Positive Equilibrium for
4. Global Asymptotic Stability of Positive Equilibrium
4.1. Global Asymptotic Stability of Positive Equilibrium for
4.2. Global Asymptotic Stability of Positive Equilibrium for
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fatma, B.; Ali, Y.; Chandan, M. Effects of fear in a fractional-order predator-prey system with predator density-dependent prey mortality. Chaos Soliton. Fract. 2021, 145, 110711. [Google Scholar]
- Jiang, X.; Chen, X.; Huang, T.; Yan, H. Bifurcation and Control for a Predator-Prey System With Two Delays. IEEE Trans. Circuits Syst. II 2021, 68, 376–380. [Google Scholar] [CrossRef]
- Liu, J.; Hu, J.; Yuen, P. Extinction and permanence of the predator-prey system with general functional response and impulsive control. Appl. Math. Model. 2020, 88, 55–67. [Google Scholar] [CrossRef]
- Qiu, H.; Guo, S.; Li, S. Stability and Bifurcation in a Predator–Prey System with Prey-Taxis. Int. J. Bifurcat. Chaos 2020, 30, 2050022. [Google Scholar] [CrossRef]
- Molla, H.; Sarwardi, S.; Sajid, M. Predator-prey dynamics with Allee effect on predator species subject to intra-specific competition and nonlinear prey refuge. J. Math. Comput. Sci. 2021, 25, 150–165. [Google Scholar]
- Roy, J.; Barman, D.; Alam, S. Role of fear in a predator-prey system with ratio-dependent functional response in deterministic and stochastic environment. Biosystems 2020, 197, 104176. [Google Scholar] [CrossRef]
- Song, D.; Li, C.; Song, Y. Stability and cross-diffusion-driven instability in a diffusive predator–prey system with hunting cooperation functional response. Nonlinear Anal. Real 2020, 54, 103106. [Google Scholar] [CrossRef]
- Köhnke, M.C.; Siekmann, I.; Seno, H.; Malchow, H. A type IV functional response with different shapes in a predator–prey model. J. Theor. Biol. 2020, 505, 110419. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Takeuchi, Y. Dynamics of the density dependent predator-prey system with Beddington-DeAngelis functional response. J. Math. Anal. Appl. 2011, 374, 644–654. [Google Scholar] [CrossRef] [Green Version]
- May, R.M. Stability and Complexity in Model Ecosystems (Monographs in Population Biology); Princeton University Press: Princeton, NJ, USA, 1973; Volume 6. [Google Scholar]
- Oaten, A.; Murdoch, W.W. Functional response and stability in predator–prey systems. Am. Nat. 1975, 109, 289–298. [Google Scholar] [CrossRef]
- Vos, M.; Moreno Berrocal, S.; Karamaouna, F.; Hemerik, L.; Vet, L.E.M. Plant-mediated indirect effects and the persistence of parasitoid—Herbivore communities. Ecol. Lett. 2001, 4, 38–45. [Google Scholar] [CrossRef]
- Gross, T.; Ebenhöh, W.; Feudel, U. Enrichment and food chain stability: The impact of different forms of predator–prey interactions. J. Theor. Biol. 2004, 227, 349–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCann, K.; Rasmussen, J.; Umbanhowar, J. The dynamics of spatially coupled food webs. Ecol. Lett. 2005, 8, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Beddington, J.R. Mutual interference between parasites or predators and its effect on searching efficiency. J. Anim. Ecol. 1975, 44, 331–340. [Google Scholar] [CrossRef] [Green Version]
- DeAngelis, D.L.; Goldstein, R.A.; O’Neil, R.V. A model of trophic interaction. Ecology 1975, 56, 881–892. [Google Scholar] [CrossRef]
- Holling, C.S. The components of predation as revealed by a study of small-mammal predation of the European pine sawfly. Canad. Entomol. 1959, 91, 293–320. [Google Scholar] [CrossRef]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 1959, 91, 385–395. [Google Scholar] [CrossRef]
- Solomon, M.E. The natural control of animal populations. J. Anim. Ecol. 1949, 18, 1–35. [Google Scholar] [CrossRef]
- Holling, C.S. The functional response of invertebrate predators to prey density. Mem. Entomol. Soc. Can. 1966, 48, 1–87. [Google Scholar] [CrossRef]
- Kratina, P.; Vos, M.; Bateman, A.; Anholt, B.R. Functional response modified by predator density. Oecologia 2009, 159, 425–433. [Google Scholar] [CrossRef]
- Aiello, W.G.; Freedman, H.I. A time-delay model of single-species growth with stage structure. Math. Biosci. 1990, 101, 139–153. [Google Scholar] [CrossRef]
- Freedman, H.I.; Wu, J.H. Persistence and global asymptotic stability of single species dispersal models with stage structure. Quart. Appl. Math. 1991, 2, 351–371. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zeng, Z. Stationary distribution and extinction of a stochastic ratio-dependent predator–prey system with stage structure for the predator. Physica A 2020, 545, 123310. [Google Scholar] [CrossRef]
- Hu, D.; Li, Y.; Liu, M.; Bai, Y. Stability and Hopf bifurcation for a delayed predator–prey model with stage structure for prey and Ivlev-type functional response. Nonlinear Dyn. 2020, 99, 3323–3350. [Google Scholar] [CrossRef]
- Ghosh, B.; Zhdanova, O.L.; Barman, B.; Frisman, E.Y. Dynamics of stage-structure predator-prey systems under density-dependent effect and mortality. Ecol. Complex. 2020, 41, 100812. [Google Scholar] [CrossRef]
- Liu, S.; Beretta, E. A stage-structured predator-prey model of Beddington-DeAngelis type. SIAM J. Appl. Math. 2006, 66, 1101–1129. [Google Scholar] [CrossRef]
- Song, X.; Chen, L. Optimal harvesting and stability for a two species competitive system with stage structure. Math. Biosci. 2001, 170, 173–186. [Google Scholar] [CrossRef]
- She, Z.; Li, H. Dynamics of a density-dependent stage-structured predator–prey system with Beddington–DeAngelis functional response. J. Math. Anal. Appl. 2013, 406, 188–202. [Google Scholar] [CrossRef]
- Qu, Y.; Wei, J. Bifurcation analysis in a predator-prey system with stage-structure and harvesting. J. Franklin. Inst. 2010, 347, 1097–1113. [Google Scholar] [CrossRef]
- Xiao, D.; Jennings, L.S. Bifurcations of a ratio-dependent predator–prey system with constant rate harvesting. SIAM J. Appl. Math. 2005, 65, 737–753. [Google Scholar] [CrossRef]
- Chen, J.; Huang, J.; Ruan, S.; Wang, J. Bifurcations of invariant tori in predator–prey models with seasonal prey harvesting. SIAM J. Appl. Math. 2013, 73, 1876–1905. [Google Scholar] [CrossRef]
- Yan, X.; Zhang, C. Global stability of a delayed diffusive predator–prey model with prey harvesting of Michaelis–Menten type. Appl. Math. Lett. 2021, 114, 106904. [Google Scholar] [CrossRef]
- Mortuja, M.G.; Chaube, M.K.; Kumar, S. Dynamic analysis of a predator-prey system with nonlinear prey harvesting and square root functional response. Chaos Soliton. Fract. 2021, 148, 111071. [Google Scholar] [CrossRef]
- Brauer, F.; Soudack, A. Stability regions in predator-prey systems with constant rate prey harvesting. J. Math. Biol. 1979, 8, 55–71. [Google Scholar] [CrossRef]
- May, R.M.; Beddington, J.R.; Clark, C.W.; Holt, S.J.; Laws, R.M. Management of multispecies fisheries. Science 1979, 205, 267–277. [Google Scholar] [CrossRef]
- Hale, J.K. Theory of Functial Differential Equations; Springer: New York, NY, USA, 1977. [Google Scholar]
- Kar, T.K.; Pahari, U.K. Modelling and analysis of a prey-predator system with stage-structure and harvesting. Nonlinear Anal. 2007, 8, 601–609. [Google Scholar] [CrossRef]
- Hale, J. Ordinary Differential Equation; Krieger Publishing: Malabar, FL, USA, 1980. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Cheng, X. Dynamics of Stage-Structured Predator–Prey Model with Beddington–DeAngelis Functional Response and Harvesting. Mathematics 2021, 9, 2169. https://doi.org/10.3390/math9172169
Li H, Cheng X. Dynamics of Stage-Structured Predator–Prey Model with Beddington–DeAngelis Functional Response and Harvesting. Mathematics. 2021; 9(17):2169. https://doi.org/10.3390/math9172169
Chicago/Turabian StyleLi, Haiyin, and Xuhua Cheng. 2021. "Dynamics of Stage-Structured Predator–Prey Model with Beddington–DeAngelis Functional Response and Harvesting" Mathematics 9, no. 17: 2169. https://doi.org/10.3390/math9172169
APA StyleLi, H., & Cheng, X. (2021). Dynamics of Stage-Structured Predator–Prey Model with Beddington–DeAngelis Functional Response and Harvesting. Mathematics, 9(17), 2169. https://doi.org/10.3390/math9172169