Global Stability of Within-Host Virus Dynamics Models with Multitarget Cells


Abstract
1. Introduction
2. Virus Dynamics Model
2.1. Properties of Solutions
2.2. Steady States
2.3. Global Stability
3. Virus Model with Latency
3.1. Properties of Solutions
3.2. Steady States
3.3. Global Stability
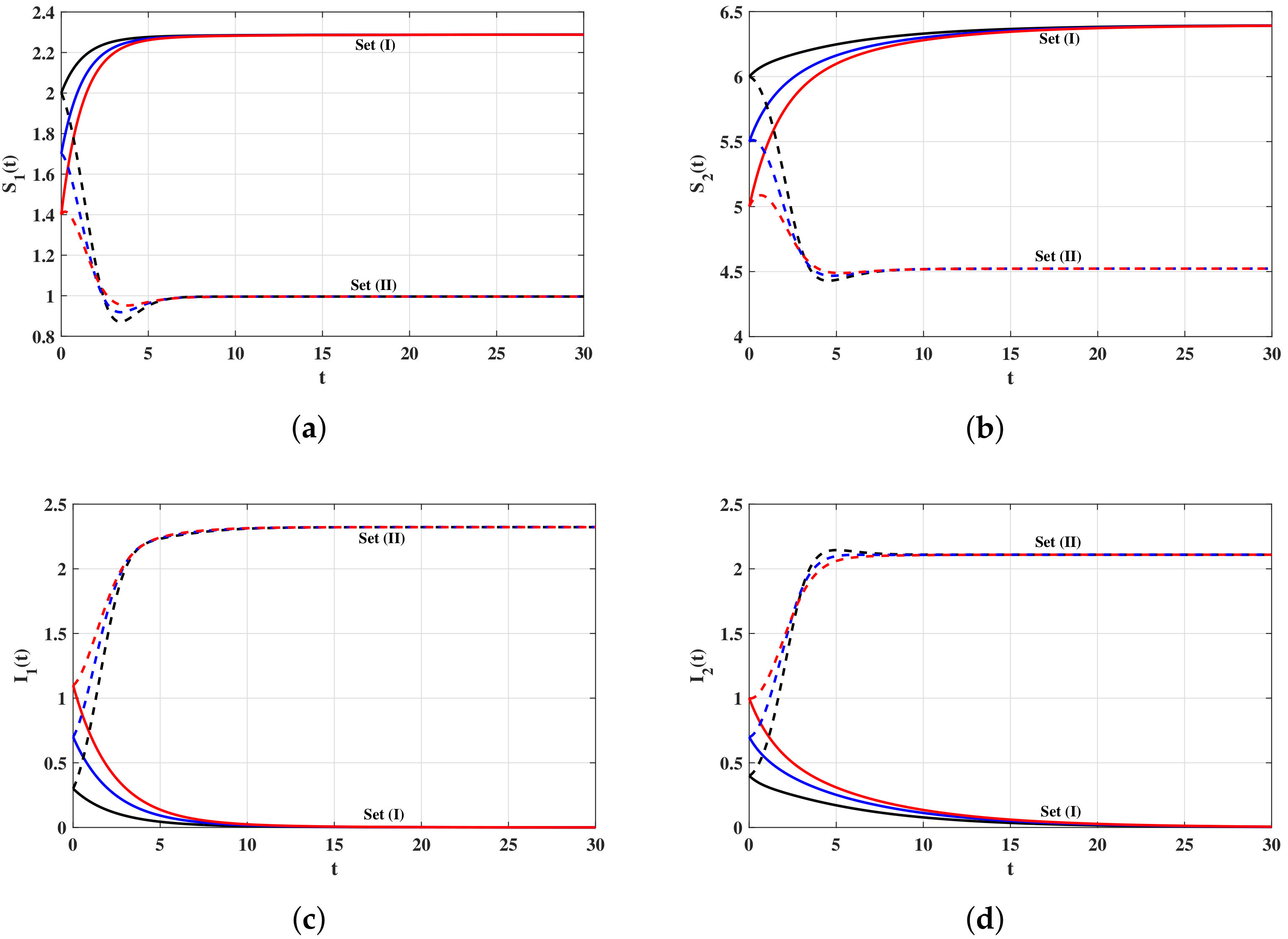
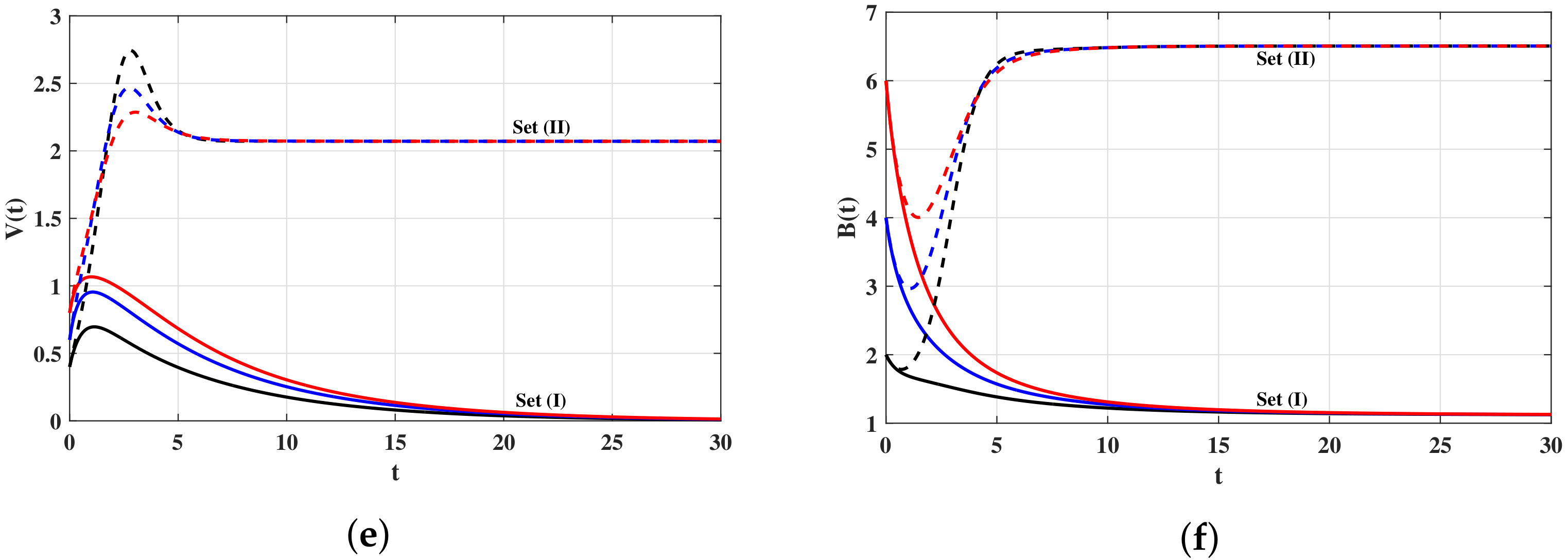
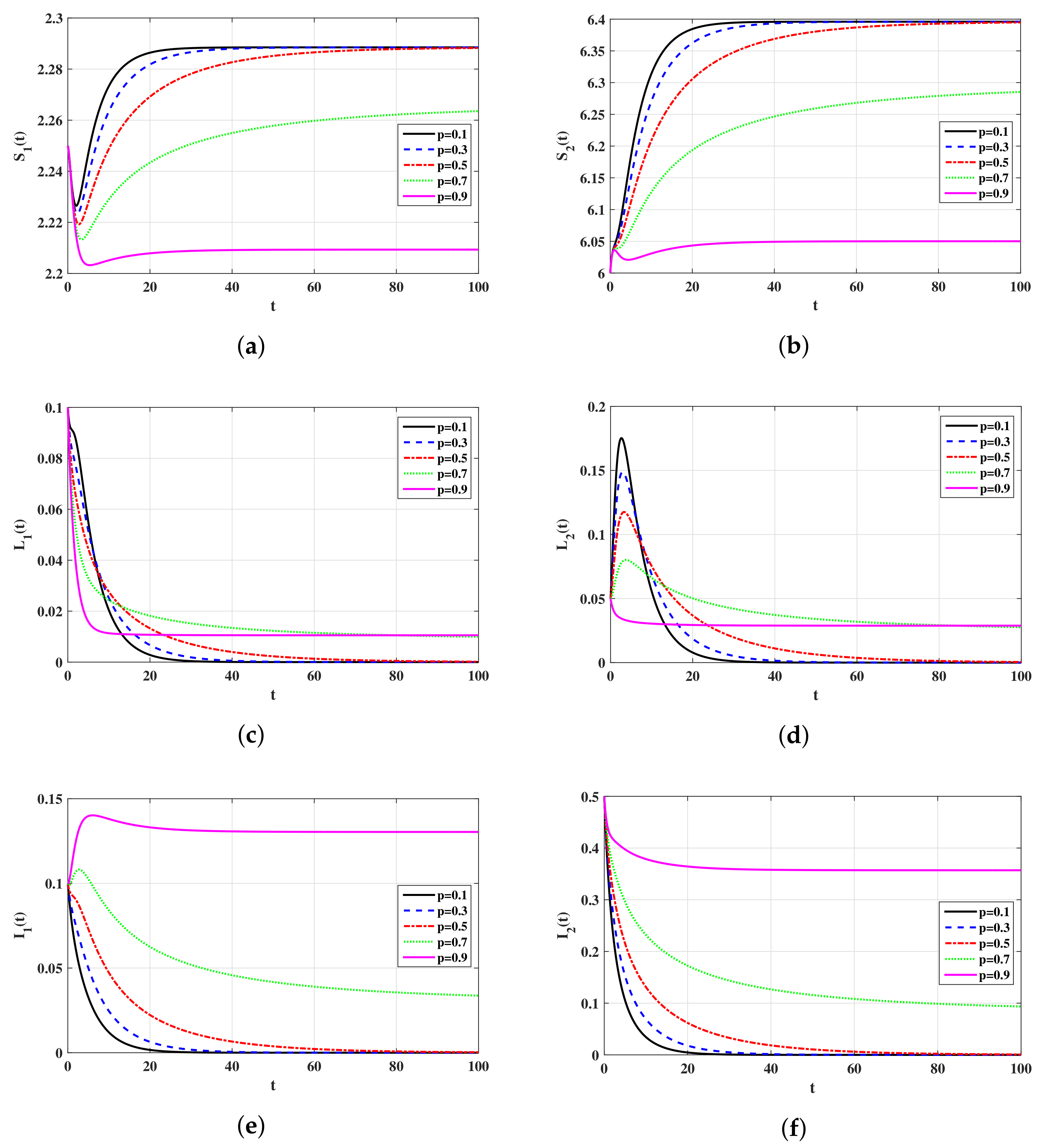
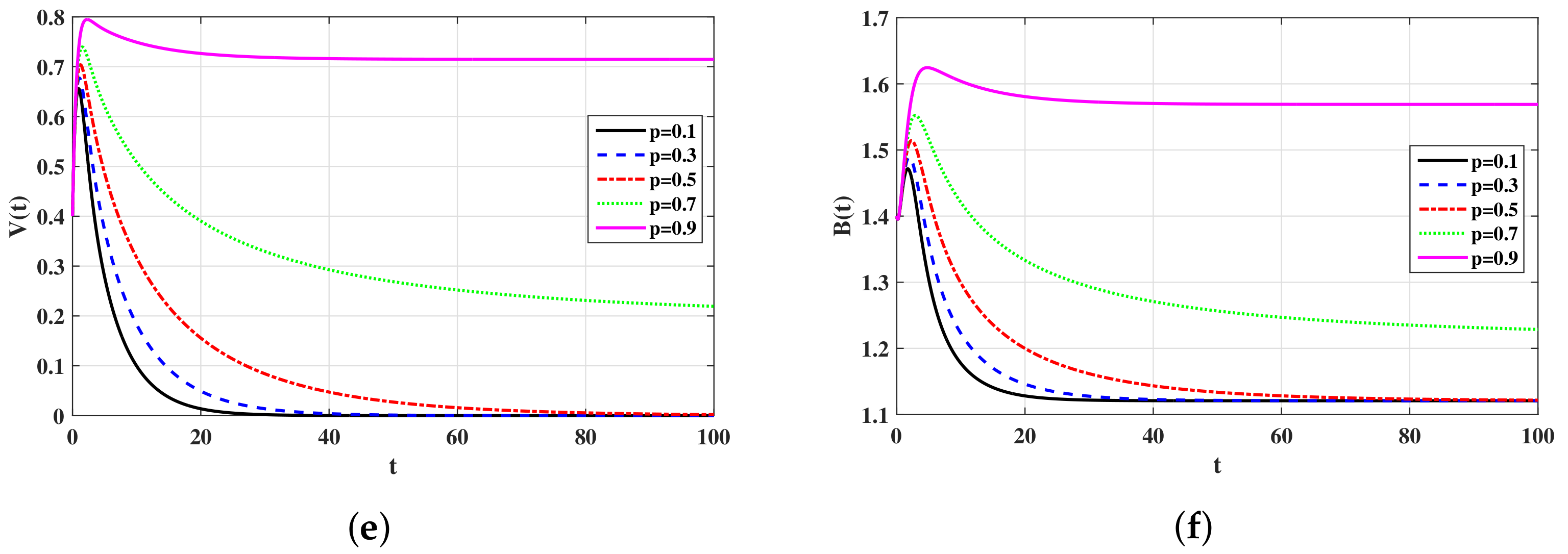
4. Numerical Simulations
4.1. Simulations for Virus Dynamics Model
4.2. Simulations for Virus Model with Latency
5. Conclusions and Discussion
5.1. Effects of Latency on the Virus Dynamics
Author Contributions
Acknowledgments
Conflicts of Interest
Appendix A
References
- Nowak, M.A.; Bangham, C.R.M. Population dynamics of immune responses to persistent viruses. Science 1996, 272, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Bonhoeffer, S.; May, R.M.; Shaw, G.M.; Nowak, M.A. Virus dynamics and drug therapy. Proc. Natl. Acad. Sci. USA 1997, 94, 6971–6976. [Google Scholar] [CrossRef] [PubMed]
- Elaiw, A.M.; Raezah, A.A. Stability of general virus dynamics models with both cellular and viral infections and delays. Math. Meth. Appl. Sci. 2017, 40, 5863–5880. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Elnahary, E.K.; Raezah, A.A. Effect of cellular reservoirs and delays on the global dynamics of HIV. Adv. Differ. Equ. 2018, 2018, 85. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H. Stability of latent pathogen infection model with adaptive immunity and delays. J. Integr. Neurosci. 2018. [Google Scholar] [CrossRef] [PubMed]
- Roy, P.K.; Chatterjee, A.N.; Greenhalgh, D.; Khan, Q.J.A. Long term dynamics in a mathematical model of HIV-1 infection with delay in different variants of the basic drug therapy model. Nonlinear Anal. Real World Appl. 2013, 14, 1621–1633. [Google Scholar] [CrossRef]
- Connell, M.C.; Yang, Y. Global stability of a diffusive virus dynamics model with general incidence function and time delay. Nonlinear Anal. Real World Appl. 2015, 25, 64–78. [Google Scholar] [CrossRef]
- Liu, S.; Wang, L. Global stability of an HIV-1 model with distributed intracellular delays and a combination therapy. Math. Biosci. Eng. 2010, 7, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fu, S. Global stability of a virus dynamics model with intracellular delay and CTL immune response. Math. Meth. Appl. Sci. 2015, 38, 420–430. [Google Scholar] [CrossRef]
- Hattaf, K.; Yousfi, N.; Tridane, A. Mathematical analysis of a virus dynamics model with general incidence rate and cure rate. Nonlinear Anal. Real World Appl. 2012, 13, 1866–1872. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Raezah, A.A.; Hattaf, K. Stability of HIV-1 infection with saturated virus-target and infected-target incidences and CTL immune response. Int. J. Biomath. 2017, 15, 1750070. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Raezah, A.A.; Alofi, B.S. Dynamics of delayed pathogen infection models with pathogenic and cellular infections and immune impairment. AIP Adv. 2018, 8, 025323. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Raezah, A.; Alofi, A.S. Effect of humoral immunity on HIV-1 dynamics with virus-to-target and infected-to-target infections. AIP Adv. 2016, 6, 085204. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Raezah, A.; Alofi, A. Stability of a general delayed virus dynamics model with humoral immunity and cellular infection. AIP Adv. 2017, 7, 065210. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H.; Alofi, A.S. Stability of CTL immunity pathogen dynamics model with capsids and distributed delay. AIP Adv. 2017, 7, 125111. [Google Scholar] [CrossRef]
- Gibelli, L.; Elaiw, A.; Alghamdi, M.A.; Althiabi, A.M. Heterogeneous population dynamics of active particles: Progression, mutations, and selection dynamics. Math. Models Meth. Appl. Sci. 2017, 27, 617–640. [Google Scholar] [CrossRef]
- Huang, G.; Takeuchi, Y.; Ma, W. Lyapunov functionals for delay differential equations model of viral infections. SIAM J. Appl. Math. 2010, 70, 2693–2708. [Google Scholar] [CrossRef]
- Li, B.; Chen, Y.; Lu, X.; Liu, S. A delayed HIV-1 model with virus waning term. Math. Biosci. Eng. 2016, 13, 135–157. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Zhang, X.; Guo, Y.; Wang, H. Analysis of an HIV infection model with treatments and delayed immune response. Appl. Math. Model. 2016, 40, 3081–3089. [Google Scholar] [CrossRef]
- Wang, K.; Fan, A.; Torres, A. Global properties of an improved hepatitis B virus model. Nonlinear Anal. Real World Appl. 2010, 11, 3131–3138. [Google Scholar] [CrossRef]
- Manna, K. Dynamics of a diffusion-driven HBV infection model with capsids and time delay. Int. J. Biomath. 2017, 10, 1750062. [Google Scholar] [CrossRef]
- Peralta, R.; Vargas-De-Leon, C.; Miramontes, P. Global stability results in a SVIR epidemic model with immunity loss rate depending on the vaccine-age. Abstr. Appl. Anal. 2015, 2015, 341854. [Google Scholar] [CrossRef]
- Monica, C.; Pitchaimani, M. Analysis of stability and Hopf bifurcation for HIV-1 dynamics with PI and three intracellular delays. Nonlinear Anal. Real World Appl. 2016, 2, 55–69. [Google Scholar] [CrossRef]
- Neumann, A.U.; Lam, N.P.; Dahari, H.; Gretch, D.R.; Wiley, T.E.; Layden, T.J.; Perelson, A.S. Hepatitis C viral dynamics in vivo and the antiviral efficacy of interferon-alpha therapy. Science 1998, 282, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, M.Y.; Kirschner, D. Mathematical analysis of the global dynamics of a model for HTLV-I infection and ATL progression. Math. Biosci. 2002, 179, 207–217. [Google Scholar] [CrossRef]
- Shi, X.; Zhou, X.; Son, X. Dynamical behavior of a delay virus dynamics model with CTL immune response. Nonlinear Anal. Real World Appl. 2010, 11, 1795–1809. [Google Scholar] [CrossRef]
- Shu, H.; Wang, L.; Watmough, J. Global stability of a nonlinear viral infection model with infinitely distributed intracellular delays and CTL imune responses. SIAM J. Appl. Math. 2013, 73, 1280–1302. [Google Scholar] [CrossRef]
- Wang, J.; Lang, J.; Zou, X. Analysis of an age structured HIV infection model with virus-to-cell infection and cell-to-cell transmission. Nonlinear Anal. Real World Appl. 2017, 34, 75–96. [Google Scholar] [CrossRef]
- Murase, A.; Sasaki, T.; Kajiwara, T. Stability analysis of pathogen-immune interaction dynamics. J. Math. Biol. 2005, 51, 247–267. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zou, D. Global stability of in host viral models with humoral immunity and intracellular delays. J. Appl. Math. Mod. 2012, 36, 1313–1322. [Google Scholar] [CrossRef]
- Wang, T.; Hu, Z.; Liao, F. Stability and Hopf bifurcation for a virus infection model with delayed humoral immunity response. J. Math. Anal. Appl. 2014, 411, 63–74. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H. Global stability of humoral immunity virus dynamics models with nonlinear infection rate and removal. Nonlinear Anal. Real World Appl. 2015, 26, 161–190. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H. Global properties of nonlinear humoral immunity viral infection models. Int. J. Biomath. 2015, 8, 1550058. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H. Stability of a general delay-distributed virus dynamics model with multi-staged infected progression and immune response. Math. Meth. Appl. Sci. 2017, 40, 699–719. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H.; Hattaf, K. Dynamical behaviors of a general humoral immunity viral infection model with distributed invasion and production. Int. J. Biomath. 2017, 10, 1750035. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X. Stability and Hopf bifurcation of a within-host chikungunya virus infection model with two delays. Math. Comput. Simul. 2017, 138, 31–48. [Google Scholar] [CrossRef]
- Deans, J.A.; Cohen, S. Immunology of malaria. Ann. Rev. Microbiol. 1983, 37, 25–49. [Google Scholar] [CrossRef] [PubMed]
- Perelson, A.S.; Essunger, P.; Cao, Y.; Vesanen, M.; Hurley, A.; Saksela, K.; Markowitz, M.; Ho, D.D. Decay characteristics of HIV-1-infected compartments during combination therapy. Nature 1997, 387, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolny, H.M.; Baron, M.J.; Gieschke, R.; Davies, B.E.; Jumbe, N.L.; Beauchemin, C.A.A. Exploring cell tropism as a possible contributor to influenza infection severity. PLoS ONE 2011, 5, e13811. [Google Scholar] [CrossRef] [PubMed]
- Bajaria, S.H.; Webb, G.; Cloyd, M.; Kirschner, D. Dynamics of naive and memory CD4 + T Lymphocytes in HIV-1 disease progression. J. Acquir. Immune Defic. Syndr. 2002, 30, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Payne, R.J.H.; Nowak, M.A.; Blumberg, B.S. Analysis of a cellular model to account for the natural history of infection by the hepatitis B virus and its role in the development of primary hepatocellular carcinoma. J. Theor. Biol. 1992, 159, 215–240. [Google Scholar] [CrossRef]
- Payne, R.J.H.; Nowak, M.A.; Blumberg, B.S. A cellular model to explain the pathogenesis of infection by the hepatitis B virus. Math. Biosci. 1994, 123, 25–58. [Google Scholar] [CrossRef]
- Dahari, H.; Feliu, A.; Garcia-Retortillo, M.; Forns, X.; Neumann, A.U. Second hepatitis C compartment indicated by viral dynamics during liver transplantation. J. Hepatol. 2005, 42, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Elaiw, A.M.; Azoz, S.A. Global properties of a class of HIV infection models with Beddington-DeAngelis functional response. Math. Meth. Appl. Sci. 2013, 36, 383–394. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Hassanien, I.A.; Azoz, S.A. Global stability of HIV infection models with intracellular delays. J. Korean Math. Soc. 2012, 49, 779–794. [Google Scholar] [CrossRef]
- Elaiw, A.M. Global properties of a class of HIV models. Nonlinear Anal. Real World Appl. 2010, 11, 2253–2263. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Almuallem, N.A. Global dynamics of delay-distributed HIV infection models with differential drug efficacy in cocirculating target cells. Math. Meth. Appl. Sci. 2016, 39, 4–31. [Google Scholar] [CrossRef]
- Cunningham, A.L.; Donaghy, H.; Harman, A.N.; Kim, M.; Turville, S.G. Manipulation of dendritic cell function by viruses. Curr. Opin. Microbiol. 2010, 13, 524–529. [Google Scholar] [CrossRef] [PubMed]
- Couderc, T.; Chretien, F.; Schilte, C.; Disson, O.; Brigitte, M.; Guivel-Benhassine, F. A mouse model for Chikungunya: young age and inefficient type-I interferon signaling are risk factors for severe disease. PLoS Pathog. 2008, 4, e29. [Google Scholar] [CrossRef] [PubMed]
- Lum, F.M.; Ng, L.F.P. Cellular and molecular mechanisms of chikungunya pathogenesis. Antivir. Res. 2015, 120, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Ozden, S.; Huerre, M.; Riviere, J.P.; Coffey, L.L.; Afonso, P.V.; Mouly, V. Human muscle satellite cells as targets of Chikungunya virus infection. PLoS ONE 2007, 2, e527. [Google Scholar] [CrossRef] [PubMed]
- Her, Z.; Malleret, B.; Chan, M.; Ong, E.K.; Wong, S.C.; Kwek, D.J. Active infection of human blood monocytes by Chikungunya virus triggers an innate immune response. J. Immunol. 2010, 184, 5903–5913. [Google Scholar] [CrossRef] [PubMed]
- Elaiw, A.M. Global properties of a class of virus infection models with multitarget cells. Nonlinear Dyn. 2012, 69, 423–435. [Google Scholar] [CrossRef]
- Wang, X.; Song, X.; Tang, S.; Rong, L. Analysis of HIV models with multiple target cell populations and general nonlinear rates of viral infection and cell death. Math. Comput. Simul. 2016, 124, 87–103. [Google Scholar] [CrossRef]
- Korobeinikov, A. Global properties of basic virus dynamics models. Bull. Math. Biol. 2004, 66, 879–883. [Google Scholar] [CrossRef] [PubMed]
- LaSalle, J.P. Stability theory of ordinary differential equations. J. Differ. Equ. 1968, 4, 57–65. [Google Scholar] [CrossRef]
- LaSalle, J.P. The stability of dynamical system. In The Stability of Dynamical Systems; SIAM: Philadephia, PE, USA, 1976. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value | Parameter | Value |
|---|---|---|---|
| varied | q | ||
| c | r | ||
| varied | |||
| Parameter | Value | Parameter | Value |
|---|---|---|---|
| le | q | ||
| c | r | ||
| = | |||
| = | |||
| = | varied |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elaiw, A.M.; Alade, T.O.; Alsulami, S.M. Global Stability of Within-Host Virus Dynamics Models with Multitarget Cells. Mathematics 2018, 6, 118. https://doi.org/10.3390/math6070118
Elaiw AM, Alade TO, Alsulami SM. Global Stability of Within-Host Virus Dynamics Models with Multitarget Cells. Mathematics. 2018; 6(7):118. https://doi.org/10.3390/math6070118
Chicago/Turabian StyleElaiw, Ahmed M., Taofeek O. Alade, and Saud M. Alsulami. 2018. "Global Stability of Within-Host Virus Dynamics Models with Multitarget Cells" Mathematics 6, no. 7: 118. https://doi.org/10.3390/math6070118
APA StyleElaiw, A. M., Alade, T. O., & Alsulami, S. M. (2018). Global Stability of Within-Host Virus Dynamics Models with Multitarget Cells. Mathematics, 6(7), 118. https://doi.org/10.3390/math6070118