Abstract
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is a new virus which infects the respiratory system and causes the coronavirus disease 2019 (COVID-19). The coinfection between malaria and COVID-19 has been registered in many countries. This has risen an urgent need to understand the dynamics of coinfection. In this paper, we construct a reaction–diffusion in-host malaria/COVID-19 model. The model includes seven-dimensional partial differential equations that explore the interactions between seven compartments, healthy red blood cells (RBCs), infected RBCs, free merozoites, healthy epithelial cells (ECs), infected ECs, free SARS-CoV-2 particles, and antibodies. The biological validation of the model is confirmed by establishing the nonnegativity and boundedness of the model’s solutions. All equilibrium points with the corresponding existence conditions are calculated. The global stability of all equilibria is proved by picking up appropriate Lyapunov functionals. Numerical simulations are used to enhance and visualize the theoretical results. We found that the equilibrium points show the different cases when malaria and SARS-CoV-2 infections occur as mono-infection or coinfection. The shared antibody immune response decreases the concentrations of SARS-CoV-2 and malaria merozoites. This can have an important role in reducing the severity of SARS-CoV-2 if the immune response works effectively.
MSC:
35B35; 37N25; 92B05
1. Introduction
The coronavirus disease 2019 (COVID-19) is a viral disease that appeared in China at the end of 2019 and spread to most countries of the world. The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is the cause of COVID-19. Malaria-endemic regions face a great challenge due to the possibility of coinfection between malaria and other viral diseases. Indeed, malaria/COVID-19 coinfection has been founded in several countries [1]. This has increased the necessity to understand the dynamics of the coinfection and its effect on the patient.
SARS-CoV-2 is an RNA virus and belongs to the family Coronaviridae [2]. It uses the angiotensin-converting enzyme 2 (ACE2) receptor to step into the ECs [3]. Such receptor is expressed in kidney, heart, gastrointestinal tract, blood vessels, and other organs [4]. The human-to-human transmission of SARS-CoV-2 occurs via respiratory droplets containing viruses [5]. Eleven vaccines for COVID-19 were approved by the World Health Organization (WHO) for emergency use. These include Novavax/Nuvaxovid, Bharat Biotech/Covaxin, CanSino/Convidecia, Pfizer/BioNTech/Comirnaty, Moderna/Spikevax, Serum Institute of India COVOVAX (Novavax formulation), Janssen (Johnson & Johnson)/Jcovden, Oxford/AstraZeneca/Vaxzevria, Serum Institute of India Covishield (Oxford/AstraZeneca formulation), Sinopharm (Beijing)/Covilo, and Sinovac/CoronaVac [6]. There are a number of other effective vaccines that are not yet approved by the WHO. On 22 October 2020, the U.S. Food and Drug Administration (FDA) approved the antiviral drug Veklury (remdesivir) for the treatment of COVID-19 cases who need hospitalization [7]. It is utilized for adults and pediatric patients 12 years of age and older (with weight ≥ 40 kg) [7].
On the other hand, malaria is a parasitic disease attributable to Plasmodium parasites [5,8]. There are five types of Plasmodium parasites: P. malariae, P. knowlesi, P. vivax, P. falciparum, and P. ovale. However, P. falciparum is the deadliest malaria parasite. Infected Anopheles mosquitoes transmit the malaria parasite to humans [8]. There are two stages for malaria infection in the body: the liver stage and the blood stage [8]. The blood stage is responsible for most of the clinical symptoms. At the blood stage, the parasites, in the form of a merozoite attack, infect the red blood cells (RBCs) [8]. After rupturing a cell, 8–32 daughter merozoites are produced [8]. Preventive chemotherapies are utilized to treat malaria infection and their consequences [9]. In this paper, we focus on the blood stage of malaria infection.
Malaria and COVID-19 have common symptoms including fever, headache, fatigue, myalgia and difficulty in breathing [10,11,12]. This can cause difficulty in the clinical diagnosis of malaria and SARS-CoV-2 coinfection [4,5]. Wrong or late diagnosis of coinfection can have a bad effect on the health of the patient [13]. The incubation periods for Plasmodium falciparum malaria and SARS-CoV-2 are 7–14 days and 2–17 days, respectively, and this enhances the possibility of coinfection [5,13]. In fact, malaria/COVID-19 coinfection has been found in several countries [13,14,15]. Some studies indicated that the coinfection could increase the severity of SARS-CoV-2 infection [1,13]. Wilairatana et al. [1] presented a review article and identified studies of malaria/COVID-19 coinfection and compared them from several aspects including: the possible correlations between COVID-19 and malaria, the prevalence of malaria infection among COVID-19 patients, the risk of oxidative stress in the malaria/COVID-19 coinfection, the role of sex in the malaria/COVID-19 coinfection, the effect of malaria coinfection on the clearance of SARS-CoV-2 in COVID-19 patients, the clinical severity of COVID-19, treatment of COVID-19, mean duration of the hospitalized and the underlying comorbidities. Hussein et al. [16] reported that coinfection with malaria and COVID-19 is associated with increased all-cause in-hospital mortality compared to a single-infection with SARS-CoV2. Nevertheless, several studies mentioned that the neutralizing antibodies against Plasmodium falciparum can also be effective against SARS-CoV-2 particles. This can minimize the severity of SARS-CoV-2 infection in coinfected patients [4,17,18,19]. Thus, understanding the dynamics of coinfection is very crucial in order to find better ways to deal with and treat this group of patients.
Mathematical modeling is considered as one of the most substantial tools that is used to support medical studies during epidemics. Malaria models at the blood stage have been explored in many works (see for example [20,21,22,23,24,25,26]). In addition, many COVID-19 models have been formulated and studied. These models can be classified into epidemiological models and in-host models. Epidemiological models study the transmission of COVID-19 between individuals (see for example [27,28,29,30,31,32]). On the other hand, in-host models study the interactions between SARS-CoV-2 and cells inside the body (see for example [33,34,35,36,37,38]). In fact, in-host models have received less attention than between-host models. In a recent work [39], the malaria/SARS-CoV-2 coinfection model has been developed and investigated. All the above-mentioned models assume that parasites, viruses, and cells are distributed homogeneously in the body. However, this assumption is not realistic in biological systems as the diffusion of particles causes spatial variations within the body. Considering spatial diffusion converts the ODE model into a PDE model, which allows the compartments to vary in space and time. This will give a more accurate description of the model’s dynamics. Therefore, some malaria models (see for example [8,40]) and SARS-CoV-2 models (see for example [39,41]) are formulated using partial differential equations (PDEs) to take into account the diffusion of some components in the model. Actually, the coinfection of COVID-19 with malaria is an active area of research. Current studies are trying to deeply understand the dynamics of this coinfection. This will help to effectively treat coinfected patients and save their lives. Mathematical modeling can support these studies and reduce the number of experiments needed to test hypotheses. We noted that a diffusive malaria/COVID-19 coinfection model has not yet been considered. In this paper, we formulate a reaction–diffusion malaria/COVID-19 model. This model considers the interactions between healthy RBCs, infected RBCs, free merozoites, healthy ECs, infected ECs, free SARS-CoV-2 particles, and antibodies. For this model, we (i) validate the boundedness and nonnegativity of solutions, (ii) calculate all model’s equilibria and extract the conditions of their existence, (iii) show the global stability of equilibria, and (iv) enhance the analytical results by executing some numerical simulations.
The paper is written as follows: Section 2 gives a description for the proposed model. Section 3 shows the properties of the model’s solutions. Furthermore, it calculates all models’ equilibria. Section 4 introduces the Lyapunov method to establish the global stability of all model’s equilibria. Section 5 is devoted for numerical simulations. Finally, the results are discussed and some future research points are suggested in Section 6.
2. Reaction–Diffusion Malaria/COVID-19 Model with Immune Response
In this section, we give a detailed description of the proposed model. We construct the malaria/COVID-19 coinfection model as a system of seven PDEs:
for and , where , , , , , , and stand for the concentrations of healthy RBCs, infected RBCs, free merozoites, healthy ECs, infected ECs, free SARS-CoV-2 particles, and antibodies. Healthy RBCs are generated at a constant rate , get infected by merozoites at rate , and die at rate . Infected RBCs die at rate and burst to generate merozoites per infected cell. Free merozoites die at rate and are cleared by antibodies at rate . Healthy ECs are recruited from its source at rate , get infected by SARS-CoV-2 at rate and die at rate . Infected ECs die at rate and release SARS-CoV-2 at rate . SARS-CoV-2 particles are eliminated by antibodies at rate and die at rate . Antibodies die at a natural death rate and are stimulated to target malaria merozoites and SARS-CoV-2 at rates and , respectively. The spatial domain is continuous, bounded and its boundary is smooth. is the Laplacian operator. We assume that each component of the model diffused in the domain with a diffusion coefficient . The initial conditions (ICs) of model (1) are defined as the following:
The boundary conditions are given by the following Neumann boundary conditions (NBCs):
where is the outward normal derivative on . This type of boundary condition simulates a natural barrier that prevents cells and viruses from crossing the boundary.
3. Properties of Solutions
In this section, we verify the basic properties of model (1) including the existence, nonnegativity, and boundedness of the solutions. Furthermore, we evaluate all possible equilibrium points with their conditions of existence.
Let be the set of all bounded and continuous functions from to , and . The positive cone induces a partial order on . Let , where is the Euclidean norm on . This reveals that is a Banach lattice [42,43].
Theorem 1.
Proof.
For any , we define , by
We note that A is locally Lipschitz on . We rewrite system (1)–(3) as the abstract differential equation
where and , . It is possible to show that
According to [42,43,44], systems (1)–(3) have a unique nonnegative mild solution on , which is the maximal existence time interval. Next, we show that the solutions of model (1) are bounded. We define
Since , we obtain that
where . Thus, satisfies the following system:
Let be a solution to the following ODE
Thus, . Comparison principle [45] provides that . Therefore, we have
Accordingly, and are bounded. Let
As , we obtain
where . Comparison principle [45] implies that
This proves the boundedness of and . Finally, we define a function
Since , , and , we have
where . Based on the comparison principle [45], we obtain
Thus, , , and are bounded. Consequently, all solutions are bounded on . Based on the standard theory for semi-linear parabolic systems [46], the solutions are bounded on . □
Proposition 1.
- (1)
- The uninfected equilibrium always exists;
- (2)
- The SARS-CoV-2-free equilibrium without immune response exists if ;
- (3)
- The SARS-CoV-2-free equilibrium exists if ;
- (4)
- The malaria-free equilibrium without immune response exists if ;
- (5)
- The malaria-free equilibrium exists if ;
- (6)
- The malaria/COVID-19 coinfection immune-free equilibrium exists if and ;
- (7)
- The malaria/COVID-19 coinfection equilibrium exists if , , and .
Proof.
Each equilibrium of system (1) satisfies the following algebraic system:
By solving (4), we obtain the following equilibria:
- (1)
- The uninfected equilibrium , whereThus, the equilibrium always exists.
- (2)
- The malaria single-infection without immunity equilibrium is given by , , wherewhere . We note that and are positive, while and are positive for . Thus, exists when . Here, is a threshold parameter, which specifies the establishment of malaria infection.
- (3)
- The malaria single-infection with immunity equilibrium . The components are given bywhere . We see that , , and are always positive, while when . Therefore, exists if . is a threshold parameter which sets the initiation of antibody immune response against malaria merozoites.
- (4)
- The SARS-CoV-2 single-infection without immunity equilibrium is defined as . The components are given bywhere . Notably, and are always positive, while and are positive when . Here, is a threshold parameter which determines the establishment of SARS-CoV-2 infection.
- (5)
- The SARS-CoV-2 single-infection with immunity is given by , , wherewhere . We see that , , and are always positive, while if . Hence, exists if . The threshold parameter marks the establishment of antibody immunity against SARS-CoV-2 infection.
- (6)
- The malaria/SARS-CoV-2 coinfection without immunity equilibrium is given by , , whereThe components and are always positive. and are positive when , while and are positive when . Consequently, exists when and .
- (7)
- The malaria/SARS-CoV-2 coinfection with immunity equilibrium is given by , , whereBy substituting in the fourth equation of model (1), we obtainThus, fulfills the following equationLet us define a function as follows:whereBy computing the value of at , we obtainwhere . We note that ifIn addition, we find thatThus, if
This implies that there exists a root such that . Let and observe that for and ( is naturally satisfied at because coexists with when , but it will not be stable as can be concluded from Theorem 4), we have , , , , and . Similarly, to find the third existence condition of , we form a function of Z and extract the conditions at which there is a positive root. This will give
4. Global Stability of Equilibria
This section confirms the global stability of all equilibrium points by building appropriate Lyapunov functionals. Define a Lyapunov functional
and let be the largest invariant subset of
Theorem 2.
The uninfected equilibrium is globally asymptotically stable (GAS) when and .
Proof.
Define
Then, we have
By calculating the time derivative of , we have
Depending on the Divergence theorem and NBCs, we have
This implies that
We note that when and . In addition, when , , and . The solutions tend to which contains elements with and then and . The third and sixth equations of system (1) imply that . Then, and thus LaSalle’s invariance principle (LIP) [47] assures the global asymptotic stability of when and . □
Theorem 3.
Assume that . Then, the malaria single-infection without immunity equilibrium is GAS if and .
Proof.
Define
Then, we obtain
The equilibrium conditions at are
By computing , we obtain
Thus, we see that if and . In addition, when , , , , and . The solutions tend to , which has and then . From the sixth equation of (1), we obtain . Hence, . Accordingly, LIP proves the global asymptotic stability of if , and . □
Theorem 4.
Suppose that . Then, the malaria single-infection with immunity equilibrium is GAS when .
Proof.
Consider
Then, we obtain
The equilibrium conditions at are computed as
After using the equilibrium conditions to collect terms of Equation (12), we obtain
By using the values of and , we have
Accordingly, is given by
We observe that if . In addition, when , , , , and . We can prove that the elements of satisfy and . Consequently, . Therefore, the global asymptotic stability of is followed by LIP when and . □
Theorem 5.
Assume that . Then, the SARS-CoV-2 single-infection without immunity equilibrium is GAS when and .
Proof.
Define
Then, we obtain
Accordingly, is given by
This implies that if and . In addition, one can show that when , , , , , , and . Thus, . As a result, LIP insures the global asymptotic stability of when , and . □
Theorem 6.
Assume that . Then, the SARS-CoV-2 single-infection with immunity equilibrium is GAS if .
Proof.
Consider
Then, we have
The equilibrium conditions at can be written as
By utilizing Equation (15) to collect terms of Equation (14), we obtain
By using Equation (9), is computed as
Therefore, if . Furthermore, when , , , , and . One can show that . By LIP, the equilibrium is GAS if and . □
Theorem 7.
Assume that and . Then, the malaria/SARS-CoV-2 coinfection without immunity equilibrium is GAS if .
Proof.
Define
Then, we obtain
The equilibrium conditions at can be written as
By using the above conditions, the derivative in (16) becomes
To evaluate the fifth term in (17), we calculate
Accordingly, is provided as
Hence, we have if . In addition, when , , , , , , and . Thus, and, according to LIP, is GAS if , , and . □
Theorem 8.
Suppose that , , and . Then, the malaria/SARS-CoV-2 coinfection with immunity equilibrium is GAS.
Proof.
Consider
Then, we obtain
By using the equilibrium conditions at ,
and the is given by
Therefore, we have . Furthermore, we have when , , , , , , and . Thus, . It follows from LIP that is GAS when , , and . □
5. Numerical Simulations
In this section, we execute some numerical simulations to visualize the analytical results gained previously. The MATLAB PDE solver (pdepe) is used to solve the equations (see the Supplementary File S1 pdex30.m). The spatial domain is selected as with step size and time step size . The initial conditions of model (1) are taken as:
The values are assumed based on previous studies [8,48]. The results are classified into seven cases corresponding to the global stability of each equilibrium point. These cases are obtained by varying five parameters , , , , and , while the rest of the parameters take fixed values as shown in Table 1. We used the values of some parameters which are given in the literature to perform our numerical simulations. We mention that these values are taken from studies for SARS-CoV-2 single-infection and malaria single-infection. To the best of our knowledge, till now, there has been no available data (e.g., the concentrations of SARS-CoV-2 particles, merozoites, antibodies, etc.) from SARS-CoV-2 and malaria coinfection patients. Therefore, estimating the parameters of the coinfection model is sill open for future work.

Table 1.
Model parameters.
Now, we have the following cases:
- (1)
- The varied parameters are . This yields and . This implies that the equilibrium is GAS (see Figure 1), which agrees with Theorem 2. This simulates an individual who has recovered from both malaria and SARS-CoV-2 infections.
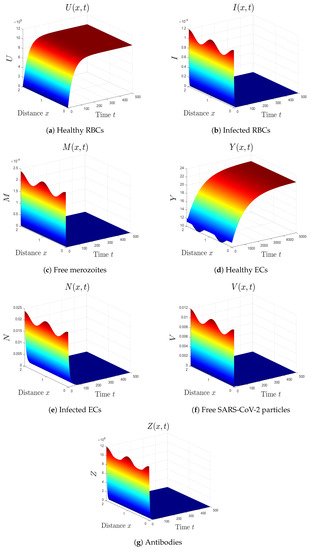 Figure 1. Simulation of system (1) for . The uninfected equilibrium is GAS.
Figure 1. Simulation of system (1) for . The uninfected equilibrium is GAS. - (2)
- The selected parameters are . Then, we obtain , , and . Figure 2 shows that the numerical results agree with the analytical results of Theorem 4. The equilibrium is GAS. This case describes a patient who only has malaria with inactive antibody immune response.
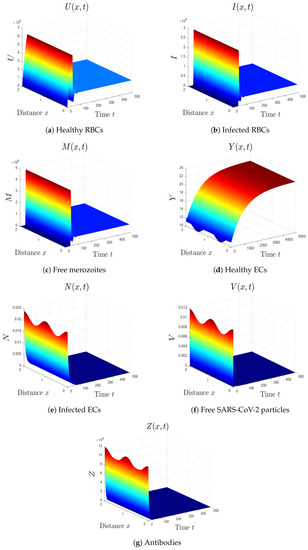 Figure 2. Simulation of system (1) for . The equilibrium is GAS.
Figure 2. Simulation of system (1) for . The equilibrium is GAS. - (3)
- The varied parameters are . This yields and . From Figure 3, we see that the equilibrium is GAS, which illustrates Theorem 4. This case represents a patient who has only malaria with an active antibody immune response.
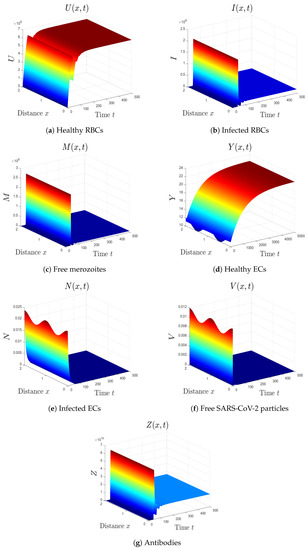 Figure 3. Simulation of system (1) for . The equilibrium is GAS.
Figure 3. Simulation of system (1) for . The equilibrium is GAS. - (4)
- By choosing , we obtain , , and . Figure 4 illustrates the global asymptotic stability of the equilibrium as given by Theorem 5. The patient in this situation suffers from SARS-CoV-2 single-infection with inactive immunity.
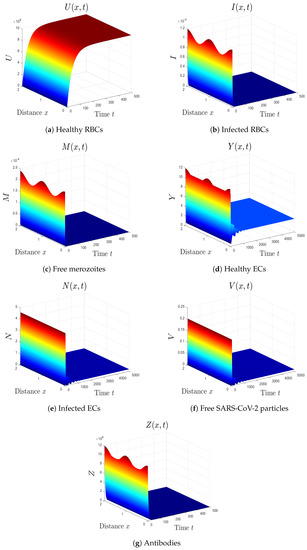 Figure 4. Simulation of system (1) for . The equilibrium is GAS.
Figure 4. Simulation of system (1) for . The equilibrium is GAS. - (5)
- By selecting we obtain , . Accordingly, the equilibrium is GAS (see Figure 5). This result comes in agreement with Theorem 6. The patient in this situation has SARS-CoV-2 single-infection with active immunity. The activation of the antibody immunity causes a reduction in the number of SARS-CoV-2 particles.
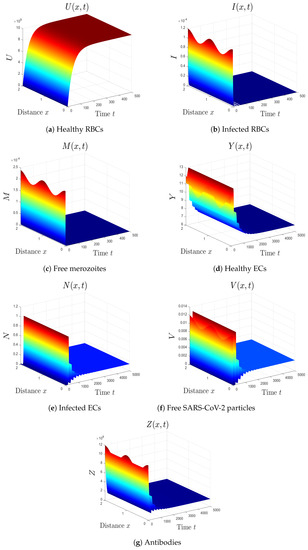 Figure 5. Simulation of system (1) for . The equilibrium is GAS.
Figure 5. Simulation of system (1) for . The equilibrium is GAS. - (6)
- We take . This gives , , and . Thus, the equilibrium is GAS (see Figure 6), which agrees with Theorem 7. Here, the coinfection of malaria and COVID-19 occurs but with inactive antibody immunity. The inactivation of immunity enhances the replication of both SARS-CoV-2 particles and malaria merozoites, which worsens the health state of the patient.
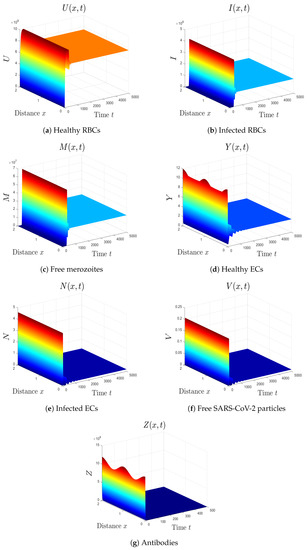 Figure 6. Simulation of system (1) for . The equilibrium is GAS.
Figure 6. Simulation of system (1) for . The equilibrium is GAS. - (7)
- We select . In this case, the threshold parameters are given as , , and . In line with Theorem 8, the equilibrium is GAS (see Figure 7). Under these circumstances, the coinfection of malaria and COVID-19 occurs with active antibody immunity. This action works on reducing the concentrations of both malaria merozoites and SARS-CoV-2 particles.
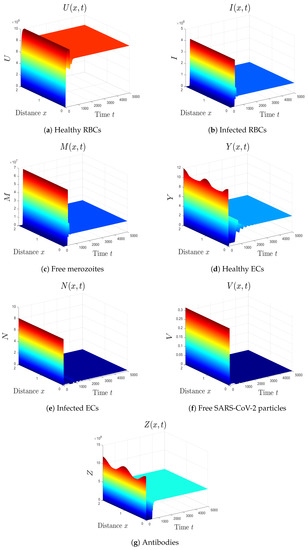 Figure 7. Simulation of system (1) for . The equilibrium is GAS.
Figure 7. Simulation of system (1) for . The equilibrium is GAS.
5.1. Sensitivity Analysis
Sensitivity analysis evaluates a relative change in a variable when a parameter changes. We execute sensitivity analysis for and as they are the main determinants for the stability of the uninfected equilibrium . The normalized forward sensitivity index of a differentiable function with respect to a parameter p is defined as
5.1.1. Sensitivity Analysis of
The normalized forward sensitivity index of is given by
We calculate the sensitivity indices of with respect to each parameter using the values provided in Table 1. The results are listed in Table 2. We note that the sensitivity indices of do not depend on any parameters. For instance, the sensitivity index of with respect to is
Table 2.
Sensitivity indices of .
Therefore, it is useful to justify the sign of the sensitivity indices of . According to Table 2, the number of merozoites produced per infected cell, , the infection rate of RBCs, , and the recruitment rate of healthy RBCs, , are the parameters that increase malaria infection in the body. Conversely, the death rate of uninfected RBCs, , and the death rate of merozoites, , are the parameters that have a crucial role in eliminating malaria infection from the body.
5.1.2. Sensitivity Analysis of
The normalized forward sensitivity index of is given by
As for , we calculate the sensitivity index of each parameter in using the values given in Table 1. The results are presented in Table 3. We see that, when one of the parameters with a positive index (e, , or ) is increased while the other parameters remain constant, this raises the value of . In other words, these parameters lead to the growth of SARS-CoV-2. Conversely, the parameters with negative indices have a role in eliminating SARS-CoV-2 infection from the body.
Table 3.
Sensitivity indices of .
6. Results and Discussion
Malaria/COVID-19 coinfection represents a true concern especially in malaria-endemic regions. Therefore, there is an urgent need to understand the dynamics of this coinfection within a human body. In this paper, we develop a reaction–diffusion in-host malaria/COVID-19 coinfection model. This model considers the interactions between healthy RBCs, infected RBCs, free merozoites, healthy ECs, infected ECs, free SARS-CoV-2 particles and antibodies. We show that the system admits seven equilibrium points and we prove the following:
- (1)
- The uninfected equilibrium always exists. Moreover, is GAS if and . This situation represents an individual who recovered from both malaria and SARS-CoV-2 infections.
- (2)
- The malaria single-infection without immunity equilibrium exists if . In addition, is GAS if and . This simulates the situation of malaria mono-infection patient with inactive immunity.
- (3)
- The malaria single-infection with immunity equilibrium exists if . Moreover, is GAS if . At this point, the antibody immune response is activated to eradicate malaria merozoites.
- (4)
- The SARS-CoV-2 single-infection without immunity equilibrium exists if . In addition, is GAS if and . This point simulates the situation of a patient who is only infected by SARS-CoV-2 and the immune response is inactive.
- (5)
- The SARS-CoV-2 single-infection with immunity equilibrium exists if . It is GAS when . The immune response is activated in the SARS-CoV-2 mono-infection patient.
- (6)
- The malaria/COVID-19 coinfection without immunity equilibrium exists if and . It is GAS when . Here, the coinfection occurs with inactive immune response.
- (7)
- The malaria/COVID-19 coinfection with immunity equilibrium exists, and it is GAS if , , and . This point represents the occurrence of malaria/COVID-19 coinfection with an active antibody immune response.
The numerical results agree with the analytical results. Based on our results, we assume that the malaria/COVID-19 coinfection can be protective as the shared antibody immune response works on clearing SARS-CoV-2. This can decrease the severity of COVID-19. This result comes in agreement with some studies that reported the positive impact of the shared antibody immune response [4,17,18,19]. However, other studies suggested that there is an increased risk of death in malaria patients with SARS-CoV-2 infection [1,13]. Therefore, more studies are required to investigate the impact of coinfection between malaria and COVID-19, to evaluate the effect of the immune system during the coinfection, and to find the suitable ways for treating the coinfected patients. The main limitation of this research work is that we did not estimate the values of the model’s parameters using real data. The reasons are as follows: (1) The data on malaria/COVID-19 coinfection are still very limited; (2) Comparing our results with a small number of real studies may not be very precise; (3) Collecting real data from patient coinfected with malaria and SARS-CoV-2 is not an easy process; (4) Working on experiments to obtain data is beyond the scope of this paper. Thus, the theoretical results obtained in this paper need to be tested against empirical findings when real data become available.
6.1. Conclusions
Malaria/COVID-19 coinfection has been reported in many countries. In this paper, we formulated a reaction–diffusion in-host model to study the coinfection between malaria and COVID-19. We assumed that the shared antibody immune response decreases the concentrations of SARS-CoV-2 and malaria merozoites. This can reduce the severity of SARS-CoV-2 in coinfected patients. The principal limitation of this paper is that we did not use real data to estimate the values of parameters or to compare the results due to the scarcity of data. Therefore, our results need to be validated when real data become available.
6.2. Future Works
The model developed in this work can be improved by (i) using real data to find a good estimation of the parameters’ values, (ii) examining the influence of time delays that may occur during infection or production of SARS-CoV-2 particles and malaria merozoites, (iii) considering viral mutations [41,50,51], (iv) considering the effect of treatments on the progression of both diseases, (v) incorporating the role of CTLs in killing infected RBCs or ECs, and (vi) considering an age-structured model to account for the age structure in the infected cells compartments, which can lead to important observations.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/math10224390/s1, File S1.
Author Contributions
Conceptualization, A.M.E. and A.D.A.A.; Formal analysis, A.M.E. and A.D.A.A.; Investigation, A.D.A.A.; Methodology, A.M.E. and A.D.A.A.; Writing—original draft, A.D.A.A.; Writing—review & editing, A.M.E. and A.D.A.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research work was funded by Institutional Fund Projects under grant no. (IFPIP: 68-130-1443) provided by the Ministry of Education and King Abdulaziz University, DSR, Jeddah, Saudi Arabia.
Data Availability Statement
Not applicable.
Acknowledgments
This research work was funded by Institutional Fund Projects under Grant No. (IFPIP: 68-130-1443). The authors gratefully acknowledge technical and financial support provided by the Ministry of Education and King Abdulaziz University, DSR, Jeddah, Saudi Arabia.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wilairatana, P.; Masangkay, F.; Kotepui, K.; Milanez, G.; Kotepui, M. Prevalence and characteristics of malaria among COVID-19 individuals: A systematic review, meta-analysis, and analysis of case reports. PLoS Neglected Trop. Dis. 2021, 15, e0009766. [Google Scholar] [CrossRef]
- Akula, S.M.; Abrams, S.L.; Steelman, L.S.; Candido, S.; Libra, M.; Lerpiriyapong, K.; Cocco, L.; Ramazzotti, G.; Ratti, S.; Follo, M.Y.; et al. Cancer therapy and treatments during COVID-19 era. Adv. Biol. Regul. 2020, 77, 100739. [Google Scholar] [CrossRef]
- Jyotsana, N.; King, M. The impact of COVID-19 on cancer risk and treatment. Cell. Mol. Bioeng. 2020, 2016, 5230219. [Google Scholar] [CrossRef]
- Hussein, M.; Albashir, A.; Elawad, O.; Homeida, A. Malaria and COVID-19: Unmasking their ties. Malar. J. 2020, 19, 457. [Google Scholar] [CrossRef]
- Di Gennaro, F.; Marotta, C.; Locantore, P.; Pizzol, D.; Putoto, G. Malaria and COVID-19: Common and different findings. Trop. Med. Infect. Dis. 2020, 5, 141. [Google Scholar] [CrossRef]
- Coronavirus Disease (COVID-19), Vaccine Tracker, World Health Organization (WHO). 2021. Available online: https://covid19.trackvaccines.org/agency/who/ (accessed on 1 October 2022).
- The U.S. Food and Drug Administration. Know Your Treatment Options for COVID-19. 2021. Available online: https://www.fda.gov/consumers/consumer-updates/know-your-treatment-options-covid-19 (accessed on 1 October 2022).
- Elaiw, A.M.; Al Agha, A.D. Global analysis of a reaction–diffusion within-host malaria infection model with adaptive immune response. Mathematics 2020, 8, 563. [Google Scholar] [CrossRef]
- Malaria, Fact Sheets, World Health Organization (WHO). 2021. Available online: https://www.who.int/news-room/fact-sheets/detail/malaria (accessed on 1 October 2022).
- Sebastiao, C.; Gaston, C.; Paixao, J.; Sacomboio, E.; Neto, Z.; de Vasconcelos, J.N.; Morais, J. Coinfection between SARS-CoV-2 and vector-borne diseases in Luanda, Angola. J. Med. Virol. 2021, 94, 366–371. [Google Scholar] [CrossRef]
- Pusparani, A.; Henrina, J.; Cahyadi, A. Co-infection of COVID-19 and recurrent malaria. J. Infect. Dev. Ctries. 2021, 15, 625–629. [Google Scholar] [CrossRef]
- Chanda-Kapata, P.; Kapata, N.; Zumla, A. COVID-19 and malaria: A symptom screening challenge for malaria endemic countries. Int. J. Infect. Dis. 2020, 94, 151–153. [Google Scholar] [CrossRef]
- Indari, O.; Baral, B.; Muduli, K.; Mohanty, A.; Swain, N.; Mohakud, N.K.; Jha, H.C. Insights into Plasmodium and SARS-CoV-2 co-infection driven neurological manifestations. Biosaf. Health 2021, 3, 230–234. [Google Scholar] [CrossRef]
- Mahajan, N.N.; Kesarwani, S.N.; Shinde, S.S.; Nayak, A.; Modi, D.N.; Mahale, S.D.; Gajbhiye, R.K. Co-infection of malaria and dengue in pregnant women with SARS-CoV-2. Int. J. Gynecol. Obstet. 2020, 151, 459–462. [Google Scholar] [CrossRef]
- Hussein, R.; Guedes, M.; Ibraheim, N.; Ali, M.M.; El-Tahir, A.; Allam, N.; Abuakar, H.; Pecoits-Filho, R.; Kotanko, P. Co-infection of malaria and early clearance of SARS-CoV-2 in healthcare workers. J. Med. Virol. 2021, 93, 2431–2438. [Google Scholar]
- Hussein, R.; Guedes, M.; Ibraheim, N.; Ali, M.M.; El-Tahir, A.; Allam, N.; Abuakar, H.; Pecoits-Filho, R.; Kotanko, P. Impact of COVID-19 and malaria coinfection on clinical outcomes: A retrospective cohort study. Clin. Microbiol. Infect. 2022, 28, 1152.e1–1152.e6. [Google Scholar] [CrossRef]
- Kalungi, A.; Kinyanda, E.; Akena, D.; Kaleebu, P.; Bisangwa, I. Less Severe Cases of COVID-19 in Sub-Saharan Africa: Could Co-infection or a Recent History of Plasmodium falciparum Infection Be Protective? Front. Immunol. 2021, 12, 565625. [Google Scholar] [CrossRef]
- Parodi, A.; Cozzani, E. Coronavirus disease 2019 (COVID 19) and Malaria. Med. Hypotheses 2020, 143, 110036. [Google Scholar] [CrossRef]
- Iesa, M.; Osman, M.; Hassan, M.; Dirar, A.; Abuzeid, N.; Mancuso, J.J.; Pandey, R.; Mohammed, A.A.; Borad, M.J.; Babiker, H.M. SARS-CoV-2 and Plasmodium falciparum common immunodominant regions may explain low COVID-19 incidence in the malaria-endemic belt. New Microbes New Infect. 2020, 38, 100817. [Google Scholar] [CrossRef]
- Anderson, R.; May, R.; Gupta, S. Nonlinear phenomena in host-parasite interactions. Parasitology 1989, 99, S59–S79. [Google Scholar] [CrossRef]
- Hetzel, C.; Anderson, R. The within-host cellular dynamics of bloodstage malaria: Theoretical and experimental studies. Parasitology 1996, 113, 25–38. [Google Scholar] [CrossRef]
- Saul, A. Models for the in-host dynamics of malaria revisited: Errors in some basic models lead to large over-estimates of growth rates. Parasitology 1998, 117, 405–407. [Google Scholar] [CrossRef]
- Hoshen, M.; Heinrich, R.; Stein, W.; Ginsburg, H. Mathematical modelling of the within-host dynamics of Plasmodium falciparum. Parasitology 2000, 121, 227–235. [Google Scholar] [CrossRef]
- Iggidr, A.; Kamgang, J.; Sallet, G.; Tewa, J. Global analysis of new malaria intrahost models with a competitive exclusion principle. SIAM J. Appl. Math. 2006, 67, 260–278. [Google Scholar] [CrossRef]
- Tumwiine, J.; Mugisha, J.; Luboobi, L. On global stability of the intra-host dynamics of malaria and the immune system. J. Math. Anal. Appl. 2008, 341, 855–869. [Google Scholar] [CrossRef]
- Orwa, T.; Mbogo, R.; Luboobi, L. Mathematical model for the in-host malaria dynamics subject to malaria vaccines. Lett. Biomath. 2018, 5, 222–251. [Google Scholar] [CrossRef]
- Asamoah, J.; Jin, Z.; Sun, G.; Seidu, B. Sensitivity assessment and optimal economic evaluation of a new COVID-19 compartmental epidemic model with control interventions. Chaos Soiltons Fractals 2021, 146, 110885. [Google Scholar] [CrossRef]
- Currie, C.; Fowler, J.; Kotiadis, K.; Monks, T. How simulation modelling can help reduce the impact of COVID-19. J. Simul. 2020, 14, 83–97. [Google Scholar] [CrossRef]
- Krishna, M.V.; Prakash, J. Mathematical modelling on phase based transmissibility of Coronavirus. Infect. Dis. Model. 2020, 5, 375–385. [Google Scholar] [CrossRef]
- Rajagopal, K.; Hasanzadeh, N.; Parastesh, F.; Hamarash, I.; Jafari, S.; Hussain, I. A fractional-order model for the novel coronavirus (COVID-19) outbreak. Nonlinear Dyn. 2020, 101, 711–718. [Google Scholar] [CrossRef]
- Chen, T.; Rui, J.; Wang, Q.; Zhao, Z.; Cui, J.A.; Yin, L. A mathematical model for simulating the phase-based transmissibility of a novel coronavirus. Infect. Dis. Poverty 2020, 9, 24. [Google Scholar] [CrossRef]
- Liu, Z.; Magal, P.; Seydi, O.; Webb, G. Understanding unreported cases in the 2019-nCoV epidemic outbreak in Wuhan, China, and the importance of major public health interventions. SSRN Electronic J. 2020, 1–12. [Google Scholar] [CrossRef]
- Du, S.Q.; Yuan, W. Mathematical modeling of interaction between innate and adaptive immune responses in COVID-19 and implications for viral pathogenesis. J. Med. Virol. 2020, 92, 1615–1628. [Google Scholar] [CrossRef]
- Li, C.; Xu, J.; Liu, J.; Zhou, Y. The within-host viral kinetics of SARS-CoV-2. Math. Biosci. Eng. 2020, 17, 2853–2861. [Google Scholar] [CrossRef]
- Ghosh, I. Within host dynamics of SARS-CoV-2 in humans: Modeling immune responses and antiviral treatments. arXiv 2020, arXiv:2006.02936. [Google Scholar] [CrossRef]
- Pinky, L.; Dobrovolny, H.M. SARS-CoV-2 coinfections: Could influenza and the common cold be beneficial? J. Med. Virol. 2020, 92, 2623–2630. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Al Agha, A.D.; Azoz, S.A.; Ramadan, E. Global analysis of within-host SARS-CoV-2/HIV coinfection model with latency. Eur. Phys. J. Plus 2022, 137, 1–22. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Al Agha, A.D. Global dynamics of SARS-CoV-2/cancer model with immune responses. Appl. Math. Comput. 2021, 408, 126364. [Google Scholar] [CrossRef]
- Al Agha, A.D.; Elaiw, A.M. Global dynamics of SARS-CoV-2/malaria model with antibody immune response. Math. Eng. 2022, 19, 8380–8410. [Google Scholar] [CrossRef]
- Takoutsing, E.; Temgoua, A.; Yemele, D.; Bowong, S. Dynamics of an intra-host model of malaria with periodic antimalarial treatment. Int. J. Nonlinear Sci. 2019, 27, 148–164. [Google Scholar]
- Elaiw, A.M.; Al Agha, A.D.; Alshaikh, M.A. Global stability of a within-host SARS-CoV-2/cancer model with immunity and diffusion. Int. J. Biomath. 2022, 15, 2150093. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, Z. Dynamics of a diffusive HBV model with delayed Beddington-DeAngelis response. Nonlinear Anal. Real World Appl. 2014, 15, 118–139. [Google Scholar] [CrossRef]
- Xu, Z.; Xu, Y. Stability of a CD4+ T cell viral infection model with diffusion. Int. J. Biomath. 2018, 11, 1850071. [Google Scholar] [CrossRef]
- Smith, H.L. Monotone Dynamical Systems: An Introduction to the Theory of Competitive and Cooperative Systems; American Mathematical Society: Providence, RI, USA, 1995. [Google Scholar]
- Protter, M.H.; Weinberger, H.F. Maximum Principles in Differential Equations; Prentic Hall: Englewood Cliffs, NJ, USA, 1967. [Google Scholar]
- Henry, D. Geometric Theory of Semilinear Parabolic Equations; Springer: New York, NY, USA, 1993. [Google Scholar]
- Khalil, H.K. Nonlinear Systems; Prentice-Hall: Englewood Cliffs, NJ, USA, 1996. [Google Scholar]
- Elaiw, A.; Al Agha, A. Global dynamics of reaction–diffusion oncolytic M1 virotherapy with immune response. Appl. Math. Comput. 2020, 367, 124758. [Google Scholar] [CrossRef]
- Almocera, A.S.; Quiroz, G.; Hernandez-Vargas, E.A. Stability analysis in COVID-19 within-host model with immune response. Commun. Nonlinear Sci. Numer. Simul. 2020, 95, 105584. [Google Scholar] [CrossRef]
- Bellomo, N.; Burini, D.; Outada, N. Multiscale models of Covid-19 with mutations and variants. Netw. Heterog. Media 2022, 17, 293–310. [Google Scholar] [CrossRef]
- Bellomo, N.; Burini, D.; Outada, N. Pandemics of mutating virus and society: A multi-scale active particles approach. Philos. Trans. A Math. Phys. Eng. Sci. 2022, 380, 20210161. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).