Phytosociological Study, Diversity and Conservation Status of the Cloud Forest in the Dominican Republic


 ,
,  , and
, and 
Abstract
1. Introduction
2. Results
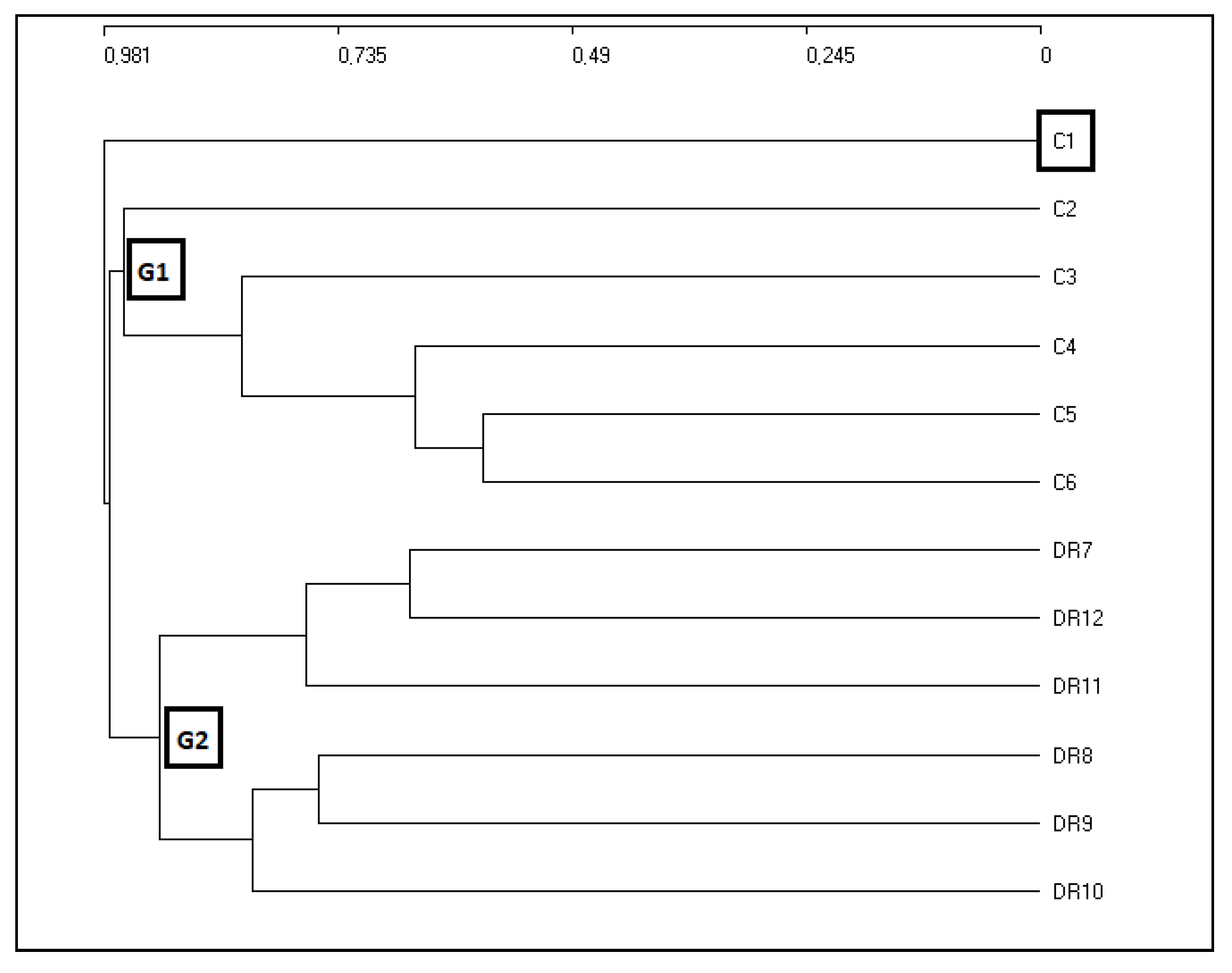
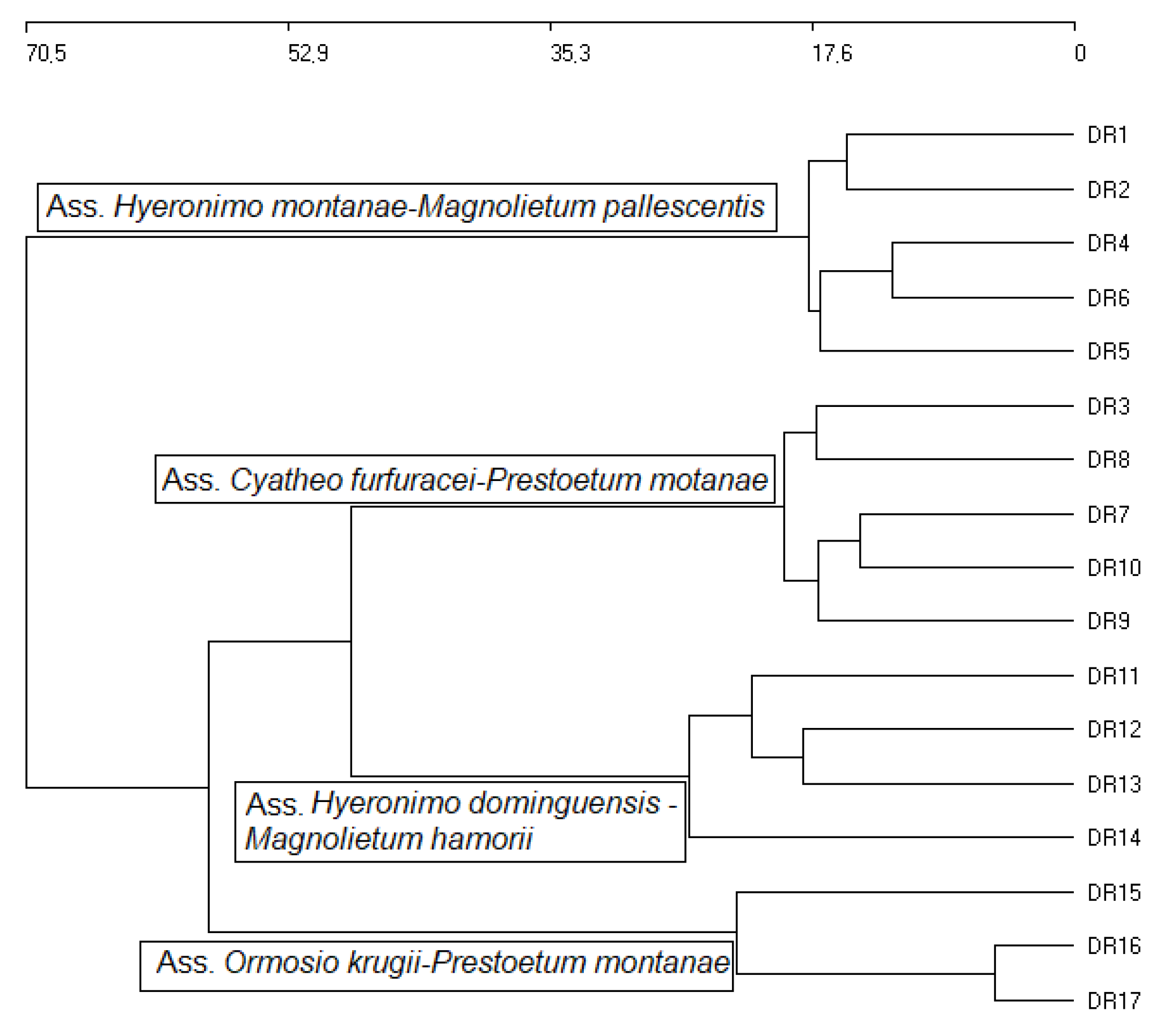
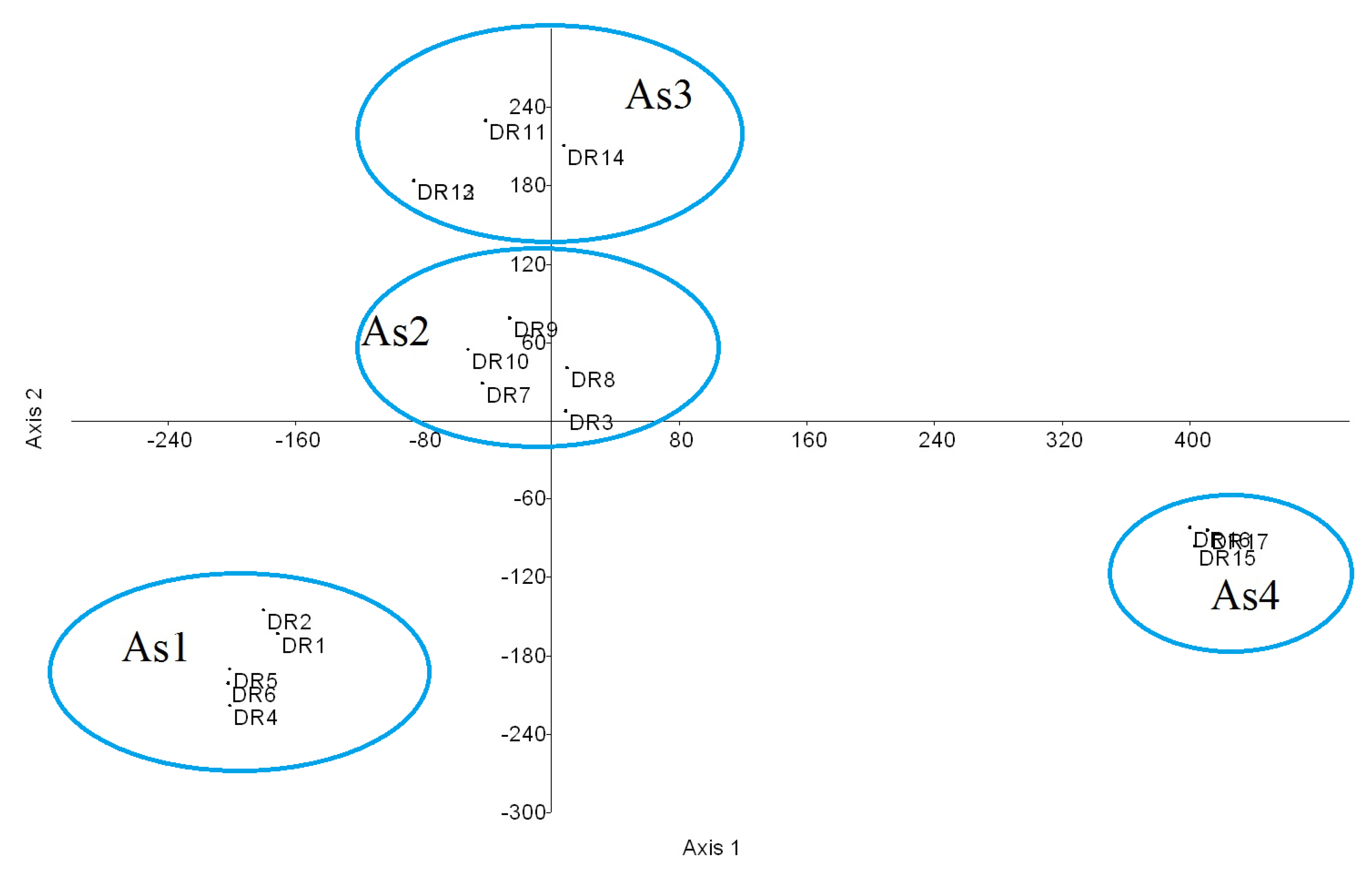
2.1. Phytosociological Study
2.2. Conservation Status of the Associations
3. Discussion
Syntaxonomical Checklist for the Cloud Forest of Hispaniola
- Cyrillo-Weinmannietea pinntae Borhidi 1996
- Cyrillo-Weinmannietalia pinnatae Borhdi 1996
- Rondeletio ochraceae-Clusion roseae Cano, Cano–Ortiz & Veloz all. nova hoc loco
- Cyatheo furfuracei-Prestoetum motanae Cano, Cano–Ortiz & Veloz ass. nova hoc loco
- Ormosio krugii-Prestoetum montanae Cano, Cano–Ortiz & Veloz ass. nova hoc loco
- Ocoteo-Cyrilletea racemiflorae Borhidi 1996
- Ocoteo cuneatae-Magnolietalia cubensis Borhidi & Muñiz in Borhidi 1996
- Rondeletio ochraceae-Didymopanion tremuli Cano, Cano–Ortiz & Veloz all. nova hoc loco
- Hyeronimo montanae-Magnolietum pallescentis Cano, Cano–Ortiz & Veloz ass. nova hoc loco
- Hyeronimo dominguensis-Magnolietum hamorii Cano, Cano–Ortiz & Veloz ass. nova hoc loco
4. Materials and Methods
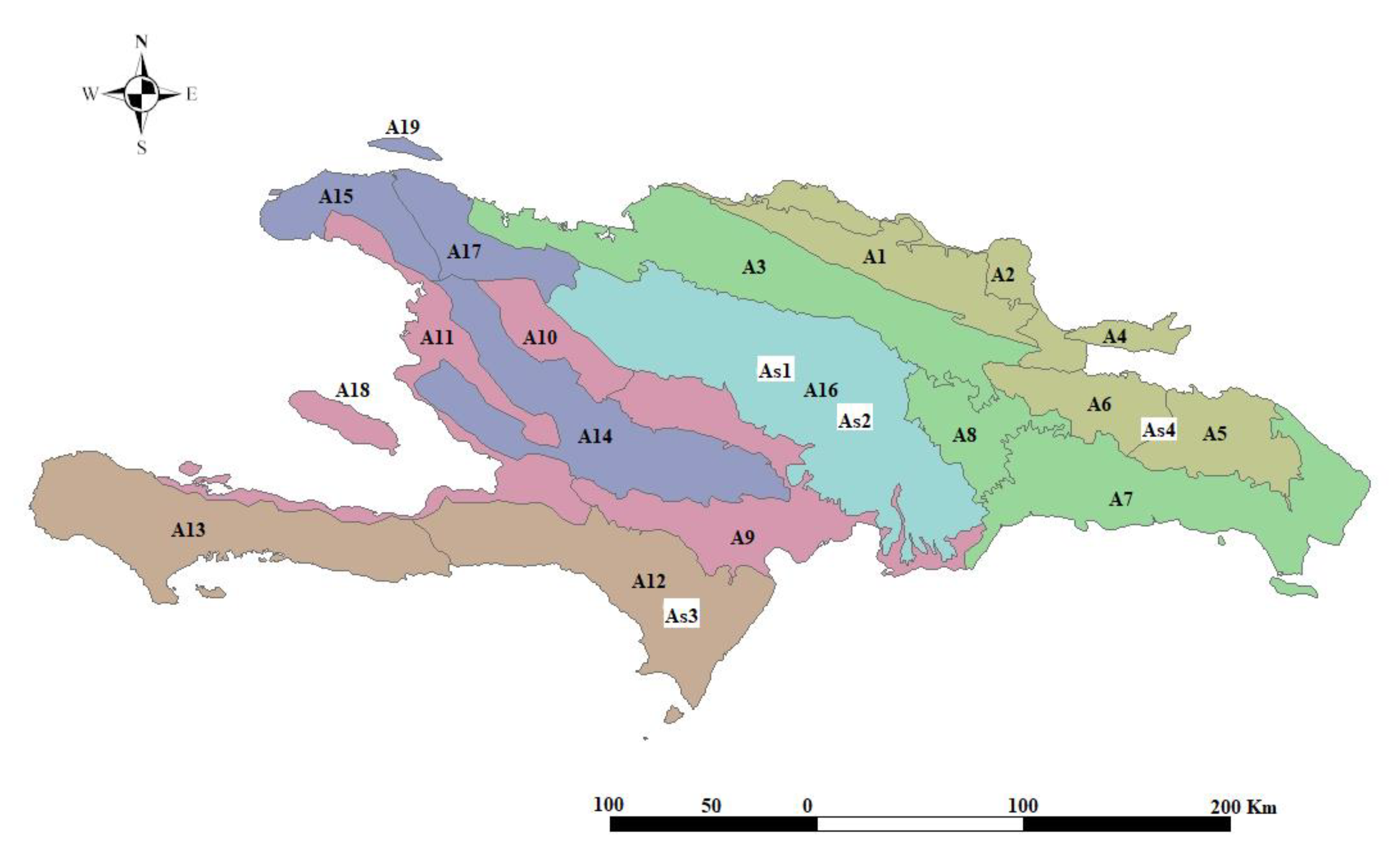
Study Area
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° rel. | 4 | 5 | 10 | 11 | 12 | |||
|---|---|---|---|---|---|---|---|---|
| N° order | DR1 | DR2 | DR4 | DR5 | DR6 | |||
| Altitude | 1481 | 1474 | 1473 | 1441 | 1465 | |||
| Area in m2 × 10 | 200 | 100 | 200 | 50 | 200 | |||
| Cover ratio In % | 100 | 90 | 100 | 100 | 100 | |||
| Xn in m. | 15 | 15 | 10 | 4 | 20 | |||
| Characteristics of the association and higher units | Family | Biotype | Status | |||||
| Magnolia pallescens Urb. & Ekm. | Magnoliaceae | A | E | 3 | 3 | 5 | 1 | 4 |
| Cyathea furfuracea Baker | Cyatheaceae | A | N | 2 | 3 | 2 | 4 | 2 |
| Chionanthus domingensis Lam. | Oleaceae | A | N | 2 | 3 | 3 | 1 | 2 |
| Gonocalyx tetrapterus A. Liogier | Ericaceae | Tr | E | 1 | 2 | 3 | 1 | 2 |
| Hyeronima montana A. Liogier | Euphorbiaceae | A | E | + | 3 | 2 | 4 | 4 |
| Didymopanax tremulus Krug. & Urb. | Araliaceae | A | E | 5 | 2 | 3 | - | 5 |
| Persea oblongifolia Kopp. | Lauraceae | A | E | 2 | 2 | 3 | 1 | 3 |
| Arthrostylidium multispicatum Pilger | Poaceae | Tr | E | 2 | 3 | 2 | 1 | 2 |
| Rondeletia ochracea Urb. | Rubiaceae | A | E | 1 | 1 | 2 | 3 | 3 |
| Alsophila minor (D.C. Eaton) R.M. Tryon | Cyatheaceae | A | N | 2 | 2 | 2 | 2 | 2 |
| Tabebuia vinosa A. Gentry | Bignoniaceae | A | E | 1 | + | 1 | 1 | + |
| Ditta maestrensis Borhidi | Euphorbiaceae | A | N | 1 | 2 | 3 | 2 | 2 |
| Smilax populnea Kunt var. horrida O.E. Schulz | Smilacaceae | Tr | N | 1 | 3 | 1 | 1 | + |
| Ilex macfadyenii (Walp.) Rehder | Aquifoliaceae | A | N | 1 | 3 | + | + | + |
| Clusia clusioides (Griseb.) D’Arcy | Clusiaceae | A | N | + | 1 | 1 | 1 | 2 |
| Cyrilla racemiflora L. | Cyrillaceae | A | N | 2 | 2 | 3 | - | 2 |
| Vaccinium racemosum (Vahl) Wilbur & Luteyn | Ericaceae | Tr | N | 2 | 3 | - | 1 | 1 |
| Cinnamomum alainii (C.K. Allen) A. Liogier | Lauraceae | A | E | - | + | 2 | 1 | 2 |
| Marcgravia rubra A. Liogier | Marcgraviaceae | Tr | E | 1 | 1 | 2 | - | 2 |
| Myrsine coriacea (Sw.) R. Br. | Myrsinaceae | A | N | 1 | 2 | + | + | - |
| Pinguicula casabitoana J. Jiménez | Lentibulariaceae | Ep | E | + | + | 1 | - | - |
| Vriesea sintenisii (Baker) L.B. Smith & Pitt. | Bromeliaceae | Ep | N | - | - | 2 | 1 | 2 |
| Ocotea nemodaphne Mez | Lauraceae | A | N | + | 1 | - | - | 2 |
| Brunellia comocladifolia H. & B. | Brunelliaceae | A | N | 1 | + | - | - | - |
| Ocotea leucoxylon (Sw.) Mez | Lauraceae | A | N | 1 | - | - | - | + |
| Schradera subsessilis Steyermark | Rubiaceae | Tr | N | 1 | - | 2 | - | - |
| Mikania venosa A. Liogier | Asteraceae | Tr | E | - | 2 | - | - | + |
| Chaetocarpus domingensis Proctor | Euphorbiaceae | A | E | - | - | 1 | - | + |
| Odontadenia polyneura (urb.) Wood. | Apocynaceae | Tr | E | - | - | - | + | + |
| Myrsine nubicola A. Liogier | Myrsinaceae | A | E | + | - | - | - | - |
| Prestoea montana (Grah.) Nichol | Arecaceae | A | N | - | 2 | - | - | - |
| Weinmannia pinnata L. | Cunoniaceae | A | N | - | + | - | - | - |
| Odontosoria uncinella (Kunze) Fée | Polypodiaceae | Tr | N | - | + | - | - | - |
| Persea krugii Mez | Lauraceae | A | N | - | - | - | 1 | - |
| Epidendrum carpophorum Barb. Rodr. | Orchidaceae | Ep | N | - | - | - | + | - |
| Pleurothallis domingensis Cogn. | Orchidaceae | Ep | E | - | - | - | + | - |
| Byrsonima lucida (Mill.) L.c. rich. | Malpighiaceae | A | N | - | - | - | + | - |
| Dilomilis montana (Sw.) Summerh. | Orchidaceae | Ep | N | - | - | - | + | - |
| Companions species | ||||||||
| Styrax ochraceus Urb. | Styracaceae | Ar | E | 1 | 1 | 1 | 1 | 1 |
| Palicourea alpina (Sw.) DC. | Rubiaceae | Ar | N | 1 | 3 | 1 | 1 | + |
| Torralbasia cuneifolia (C. Wright) Krug. & Urb. | Celastraceae | Ar | N | - | + | 4 | 2 | 3 |
| Macrocarpaea domingensis Urb. | Gentianaceae | Ar | E | 1 | - | + | 1 | 2 |
| Psychotria domingensis Jacq. | Rubiaceae | Ar | N | 3 | - | 1 | 1 | + |
| Polygala fuertesii (Urb.) Blake | Polygalaceae | Ar | E | 1 | - | 2 | 5 | - |
| Psychotria guadalupensis (DC.) Howard | Rubiaceae | Ar | N | + | 3 | - | - | + |
| Baccharis myrsinites (Lam.) Pers. | Asteraceae | Ar | N | - | - | 1 | 1 | + |
| Bocconia frutescens L. | Papaveraceae | Ar | N | + | - | - | - | - |
| Clidemia umbellata (Miller) L.O. Wms. | Melastomataceae | Ar | N | + | - | - | - | - |
| Vernonia buxifolia (Cass.) Less. | Asteraceae | Ar | N | + | - | - | - | - |
| Cestrum coelophlebium O. E. Schulz | Solanaceae | Ar | E | - | + | - | - | - |
| Lyonia alainii W. Judd. | Ericaceae | Ar | E | - | - | 1 | - | - |
| Clidemia hirta (L.) D. don | Melastomataceae | Ar | N | - | - | + | - | - |
| Renealmia jamaicensis (Gaertn.) Horan var. puberula (Gagn.) Maas | Zingiberaceae | H | N | + | 3 | 1 | 1 | + |
| Lobelia rotundifolia Juss. | Campanulaceae | H | E | 1 | - | 1 | - | + |
| Gleychenia bifida (Willd.) Spreng. | Gleycheniaceae | H | N | 2 | - | 1 | - | |
| Blechnum occidentale L. | Blechnaceae | H | N | + | - | 1 | - | + |
| Lycopodium clavatum L. | Lycopodiaceae | H | N | - | - | 1 | - | + |
| Peperomia hernandifolia (Vahl) A. Dietr. | Piperaceae | H | N | - | - | - | + | - |
| Lycopodium cernuum L. | Lycopodiaceae | H | N | 2 | - | - | - | - |
| Odontosoria aculeata (L.) J. Sm. | Polypodiaceae | H | N | - | - | - | + | - |
| Isachne rigidifolia (Poir.) Urb. | Poaceae | H | N | - | - | - | - | 1 |
| Machaerina cubensis (Kük.) T. Koyama | Cyperaceae | H | N | - | - | - | - | + |
| N° rel. | 6 | 13 | 14 | 15 | 17 | ||||
|---|---|---|---|---|---|---|---|---|---|
| N° order | DR3 | DR7 | DR8 | DR9 | DR10 | ||||
| Altitude | 1097 | 1373 | 1377 | 1251 | 1200 | ||||
| Area in m2× 10 | 200 | 50 | 100 | 100 | 50 | ||||
| Cover ratio In % | 100 | 100 | 100 | 100 | 100 | ||||
| Xn in m. | 20 | 9 | 9 | 15 | 7 | ||||
| Characteristics of the association and higher units | Family | Biotype | Status | ||||||
| Prestoea montana (Grah.) Nichol | Arecaceae | A | N | 5 | 4 | 5 | 5 | 4 | |
| Arthrostylidium multispicatum Pilger | Poaceae | Tr | E | 2 | 3 | 2 | 1 | 2 | |
| Cyathea furfuracea Baker | Cyatheaceae | A | N | 2 | 1 | 2 | 2 | + | |
| Dendropanax arboreus (L.) Dcne & Planch. | Araliaceae | A | N | 2 | - | + | + | + | |
| 3Alsophila minor (D.C. Eaton) R.M. Tryon | Cyatheaceae | A | N | 1 | 1 | 2 | 1 | - | |
| Ocotea leucoxylon (Sw.) Mez | Lauraceae | A | N | - | + | + | + | + | |
| Coccoloba wrightii Lindau | Polygnonaceae | A | N | - | 1 | + | 2 | + | |
| Alchornea latifolia Sw. | Euphorbiaceae | A | N | 2 | + | - | 1 | - | |
| Turpinia occidentalis (Sw.) G. Don | Staphyleaceae | A | N | - | - | + | 2 | 1 | |
| Brunellia comocladifolia H. & B. | Brunelliaceae | A | N | 2 | - | - | - | + | |
| Byrsonima lucida (Mill.) L.c. Rich. | Malpighiaceae | A | N | - | 1 | - | - | + | |
| Calyptrantes selleanus Urb. & Ekm. | Myrtaceae | A | E | - | - | - | - | + | |
| Cecropia screberiana Miq. | Moraceae | A | N | 2 | - | 2 | - | - | |
| Dichaea glauca (Sw.) Lindley | Orchidaceae | Ep | N | - | + | - | + | 1 | |
| Epidendrum anceps Jacq. | Orchidaceae | Ep | N | - | 1 | - | - | - | |
| Epidendrum jamaicense Lindl | Orchidaceae | Ep | N | - | - | - | + | - | |
| Epidendrum ramosum Jacq. | Orchidaceae | Ep | N | - | - | + | + | - | |
| Epidendrum ramosum Jacq. | Orchidaceae | Ep | N | - | - | - | - | - | |
| Grammitis asplenifolia (L.) Proctor | Grammitidaceae | Ep | N | - | + | - | - | - | |
| Guatteria blainii (Griseb.) Urb. | Annonaceae | A | N | - | + | - | - | + | |
| Guzmania monostrachya (Sw.) Rusby | Bromeliaceae | Ep | N | - | + | + | - | - | |
| Malpighia macracantha Ekm. & Nied. | Malpighiaceae | A | E | - | - | - | 2 | - | |
| Jacquiniella globosa (Jacq.) Schlechter | Orchidaceae | Ep | N | - | + | - | - | - | |
| Didymopanax tremulus Krug. & Urb. | Araliaceae | A | E | 1 | - | - | - | - | |
| Miconia mirabilis (Aubl.) L.O. Willians | Melastomataceae | A | N | - | + | - | - | - | |
| Exostema elliptica Griseb. | Rubiaceae | A | N | - | - | + | - | - | |
| Microgramma piloselloides L. | Polypodiaceae | Ep | N | - | - | + | - | - | |
| Camparettia falcata Poepp. & Endl. | Orchidaceae | Ep | N | - | - | - | + | - | |
| Antrophyum lanceolatum (L.) Kaulf. | Adiantaceae | Ep | N | - | - | + | - | - | |
| Myrsine coriacea (Sw.) R. Br. | Myrsinaceae | A | N | - | + | - | - | + | |
| Niphidium crassifolium (L.) Lell. | Polypodiaceae | Ep | N | - | - | + | - | - | |
| Oncidium variegatum (Sw.) Sw. | Orchidaceae | Ep | N | - | - | - | + | - | |
| Ophioglossum palmatum L. | Ophioglossaceae | Ep | N | - | - | - | - | - | |
| Phlebodium aureum (L.) J. Smith | Polypodiaceae | Ep | N | - | - | - | + | - | |
| Pleurothallis domingensis Cogn. | Orchidaceae | Ep | E | - | + | - | - | + | |
| Pothuya nudicaulis (L.) Regel | Bromeliaceae | Ep | N | - | - | - | + | - | |
| Rondeletia ochracea Urb. | Rubiaceae | A | E | - | + | - | 3 | - | |
| Companions species | |||||||||
| Myrcia splendens (Sw.) DC. | Myrtaceae | Ar | N | - | 5 | 2 | 2 | 5 | |
| Psychotria domingensis Jacq. | Rubiaceae | Ar | N | - | 3 | 3 | 1 | 1 | |
| Tabebuia bullata A. Gentry | Bignoniaceae | Ar | E | 1 | - | + | + | + | |
| Blechnum tuerckheimii A. Brause | Blechnaceae | H | E | - | 1 | 2 | 3 | - | |
| Psychotria guadalupensis (DC.) Howard | Rubiaceae | Ar | N | - | 3 | - | 1 | 1 | |
| Renealmia jamaicensis (Gaertn.) Horan var. puberula (Gagn.) Maas | Zingiberaceae | H | N | - | 2 | 2 | - | + | |
| Mikania venosa A. Liogier | Asteraceae | Tr | E | - | - | + | + | 2 | |
| Sagraea fuertesii (Cogn.in Urb.) Alain | Melastomataceae | Ar | E | - | 1 | - | - | 1 | |
| Senecio lucens (Poir) Urb. | Asteraceae | Tr | E | - | - | + | 2 | 1 | |
| Smilax havanensis Jacq. | Smilacaceae | Tr | N | - | + | - | - | - | |
| Solanum crotonoides Lam. | Solanaceae | Ar | N | - | 1 | - | - | + | |
| Solanum virgatum Lam. | Solanaceae | Ar | N | - | - | + | - | - | |
| Stigmaphyllon emarginatum (L.) A. Juss. | Malpighiaceae | Tr | N | - | - | - | - | + | |
| Uncinia hamata (L.) Urb. | Cyperaceae | H | N | - | + | + | + | - | |
| Vaccinium racemosum (Vahl) Wilbur & Luteyn | Ericaceae | Tr | N | - | + | - | - | - | |
| Vitis tiliifolia H. & B. ex Willd. | Vitaceae | Tr | N | - | - | - | - | + | |
| Vittaria lineata (L.) Smith | Pteridaceae | Ep | N | - | - | + | - | - | |
| Blechnum occidentale L. | Blechnaceae | H | N | - | 1 | 2 | - | + | |
| Cestrum coelophlebium O. E. Schulz | Solanaceae | Ar | E | - | - | - | 1 | + | |
| Cestrum inclusum Urb. | Solanaceae | Ar | E | - | - | 5 | - | - | |
| Cissampelos pareira L. | Menispermiaceae | Tr | N | 1 | - | + | - | - | |
| Commelina elegans Kunth | Commelinaceae | H | N | - | - | + | - | - | |
| Daphnosis crassifolia (Poir.) Meiss. | Thymelaeaceae | Ar | N | - | + | - | - | - | |
| Diplazium hastile (Christ.) C. Chr. | Athyriaceae | H | N | - | - | 2 | - | - | |
| Diplazium hians Kuntze | Athyriaceae | H | N | - | - | - | 2 | - | |
| Gleychenia bifida (Willd.) Spreng. | Gleycheniaceae | H | N | 1 | - | - | - | + | |
| Gomedesia lindeniana Berg. | Myrtaceae | Ar | N | - | - | - | - | 1 | |
| Gyrotaenia myriocarpa Griseb. | Urticaceae | Ar | N | - | - | + | - | - | |
| Hyptis americana (Poir.) Briq. | Lamiaceae | Ar | N | - | + | - | - | - | |
| Ichnanthus pallens (Sw.) Munro | Poaceae | H | N | - | 1 | + | + | - | |
| Ipomoea furcyensis Urb. | Convolvulaceae | Tr | E | - | - | - | + | - | |
| Lasianthus lanceolatus (Griseb.) Gómez Maza | Rubiaceae | Ar | N | - | 1 | - | - | - | |
| Lobelia robusta Graham | Campanulaceae | Ar | E | - | - | + | - | - | |
| Lobelia rotundifolia Juss. | Campanulaceae | H | E | - | + | - | - | - | |
| Odontadenia polyneura (urb.) Wood. | Apocynaceae | Tr | E | - | - | - | - | 1 | |
| Odontosoria uncinella (Kunze) Fée | Polypodiaceae | Tr | N | - | + | - | - | - | |
| Olyra latifolia L. | Poaceae | H | N | - | - | - | + | - | |
| Palicourea alpina (Sw.) DC. | Rubiaceae | Ar | N | - | + | - | - | + | |
| Peperomia hernandifolia (Vahl) A. Dietr. | Piperaceae | H | N | - | + | - | - | - | |
| Pilea geminata Urb. | Urticaceae | H | E | - | - | 2 | - | - | |
| Polypodium loriceum L. | Polypodiaceae | Ep | N | - | - | - | + | - | |
| Pothomorphe peltata (L.) Miquel | Piperaceae | Ar | N | - | - | + | - | - | |
| Mucuna urens (L.) Fawc. & Rendle | Fabaceae | Tr | N | - | + | - | - | - | |
| Myrcia deflexa (Poir) DC. | Myrtaceae | Ar | N | - | - | - | 1 | + | |
| N° rel. | 23 | 24 | 25 | 26 | |||
|---|---|---|---|---|---|---|---|
| N° order | DR11 | DR12 | DR13 | DR14 | |||
| Altitude | 1207 | 1239 | 1233 | 1140 | |||
| Area in m2× 10 | 200 | 200 | 200 | 200 | |||
| Cover ratio In % | 100 | 100 | 100 | 100 | |||
| Xn in m. | 25 | 15 | 20 | 15 | |||
| Characteristics of the association and higher units | Family | Biotype | Status | ||||
| Magnolia hamorii Howard | Magnoliaceae | A | E | 5 | 2 | 2 | 5 |
| Hyeronima domingensis Urb. | Euphorbiaceae | A | E | 5 | 2 | 5 | + |
| Cyathea fulgens C. Chr. | Cyatheaceae | A | N | 2 | 2 | 2 | 1 |
| Myrsine coriacea (Sw.) R. Br. | Myrsinaceae | A | N | 1 | 1 | 1 | 2 |
| Didymopanax tremulus Krug. & Urb. | Araliaceae | A | E | + | 5 | 2 | 3 |
| Brunellia comocladifolia H. & B. | Brunelliaceae | A | N | 2 | 1 | - | - |
| Prestoea montana (Grah.) Nichol | Arecaceae | A | N | + | 2 | 2 | 3 |
| Beilschmiedia pendula (Sw.) Hemsl. | Lauraceae | A | N | 2 | - | 1 | - |
| Ocotea leucoxylon (Sw.) Mez | Lauraceae | A | N | - | 1 | 1 | + |
| Calyptrantes selleanus Urb. & Ekm. | Myrtaceae | A | E | + | 1 | 1 | - |
| Weinmannia pinnata L. | Cunoniaceae | A | N | 2 | 2 | 2 | - |
| Pleurothalis ruscifolia (Jaq.) R. Br. | Orchidaceae | Ep | N | 1 | 2 | 2 | - |
| Elleanthus cephalotus Garay & Sweet | Orchidaceae | Ep | N | 2 | 2 | 1 | - |
| Elaphoglossum crinitum (L.) C. Chr. | Lomariopsidaceae | Ep | N | 1 | 1 | + | - |
| Columnea sanguinea Urb. | Gesneriaceae | ArEp | N | 1 | 2 | 1 | - |
| Elaphoglossum latifolium (Sw.) J. Sm. | Lomariopsidaceae | Ep | N | 2 | 2 | 2 | - |
| Miconia prasina (Sw.) DC. | Melastomataceae | A | N | 1 | - | 1 | - |
| Rondeletia ochracea Urb. | Rubiaceae | A | E | 1 | 1 | 1 | - |
| Alchornea latifolia Sw. | Euphorbiaceae | A | N | 1 | - | - | + |
| Dendropanax arboreus (L.) Dcne & Planch. | Araliaceae | A | N | 1 | - | - | - |
| Miconia mirabilis (Aubl.) L.O. Willians | Melastomataceae | A | N | - | 1 | - | + |
| Epidendrum ramosum Jacq. | Orchidaceae | Ep | N | - | - | 2 | + |
| Ophioglossum palmatum L. | Ophioglossaceae | Ep | N | + | 1 | - | - |
| Ocotea acarina C.K. Allen | Lauraceae | A | E | - | - | 2 | 1 |
| Chionanthus domingensis Lam. | Oleaceae | A | N | - | - | 2 | - |
| Ocotea nemodaphne Mez | Lauraceae | A | N | - | 1 | - | - |
| Ilex macfadyenii (Walp.) Rehder | Aquifoliaceae | A | N | - | 1 | - | - |
| Niphidium crassifolium (L.) Lell. | Polypodiaceae | Ep | N | 2 | - | - | - |
| Polypodium loriceum L. | Polypodiaceae | Ep | N | 1 | - | - | - |
| Epidendrum jamaicense Lindl | Orchidaceae | Ep | N | - | 2 | - | - |
| Phlebodium aureum (L.) J. Smith | Polypodiaceae | Ep | N | 1 | - | - | - |
| Dichaea glauca (Sw.) Lindley | Orchidaceae | Ep | N | - | 2 | - | - |
| Epidendrum carpophorum Barb. Rodr. | Orchidaceae | Ep. | N | - | - | 1 | - |
| Ocotea floribunda (Sw.) Mez | Lauraceae | A | N | - | - | - | 1 |
| Anacheilium cochleatum (L.) Hoffm. | Orchidaceae | Ep | N | - | - | - | + |
| Ocotea patens (Sw.) Nees | Lauraceae | A | N | - | - | - | + |
| Guarea guidonea Sleumer | Meliaceae | A | N | 1 | - | - | - |
| Maxillaria coccinea (Jacq.) L.O. Wms. | Orchidaceae | Ep | N | - | 2 | - | - |
| Ocotea foeniculacea Mez | Lauraceae | A | N | - | 1 | - | - |
| Cecropia screberiana Miq. | Moraceae | A | N | - | - | - | 1 |
| Beilschmiedia pendula (Sw.) Hemsl. | Lauraceae | A | N | - | - | - | 1 |
| Companions species | |||||||
| Psychotria domingensis Jacq. | Rubiaceae | Ar | N | 2 | 2 | 2 | 1 |
| Mikania venosa A. Liogier | Asteraceae | Tr | E | 1 | 2 | 1 | 2 |
| Gomedesia lindeniana Berg. | Myrtaceae | Ar | N | 1 | 1 | 1 | 2 |
| Lasianthus bahorucanus Zanoni | Rubiaceae | H | E | 2 | 2 | 1 | 1 |
| Columnea domingensis (Urb.) Wiehler | Gesneriaceae | Ar | E | 2 | 1 | + | 1 |
| Odontosoria uncinella (Kunze) Fée | Polypodiaceae | Tr | N | 3 | 2 | 2 | 2 |
| Mecranium ovatum Cog. | Melastomataceae | Ar | E | 2 | 1 | 1 | 1 |
| Vriesea tuercheimii (Mez.) L.B. Smith | Bromeliaceae | H | E | 2 | 2 | 2 | 1 |
| Nephrolepis biserrata (Sw.) Schott | Lomariopsidaceae | H | N | 2 | 2 | 2 | 2 |
| Peperomia hernandifolia (Vahl) A. Dietr. | Piperaceae | H | N | + | 1 | 1 | 1 |
| Psychotria guadalupensis (DC.) Howard | Rubiaceae | Ar | N | 2 | 2 | 1 | - |
| Myrcia deflexa (Poir) DC. | Myrtaceae | Ar | N | 2 | 1 | 1 | 2 |
| Lomariposis sorbifolia (L.) Feé | Lomariopsidaceae | H | N | 1 | - | 1 | 1 |
| Hedyosmum domingense Urb. | Chloranthaceae | Ar | E | - | 1 | 1 | + |
| Lomariposis sorbifolia (L.) Feé | Lomariopsidaceae | H | N | 2 | 2 | - | 1 |
| Renealmia jamaicensis (Gaertn.) Horan var. puberula (Gagn.) Maas | Zingiberaceae | H | N | 2 | 1 | 2 | - |
| Vaccinium racemosum (Vahl) Wilbur & Luteyn | Ericaceae | Tr | N | - | 1 | 1 | - |
| Macrocarpaea domingensis Urb. | Gentianaceae | Ar | E | - | 2 | 1 | - |
| Polygala fuertesii (Urb.) Blake | Polygalaceae | Ar | E | - | 1 | 1 | - |
| Arthrostylidium multispicatum Pilger | Poaceae | Tr | E | 3 | 2 | - | - |
| Torralbasia cuneifolia (C. Wright) Krug. & Urb. | Celastraceae | Ar | N | - | 1 | 1 | |
| Mucuna urens (L.) Fawc. & Rendle | Fabaceae | Tr | N | 1 | - | - | 2 |
| Schlegelia brachyantha Griseb. | Schlegeliaceae | Tr | N | 1 | - | - | + |
| Meriania involucrata (Desv.) Naud. | Melastomataceae | Ar | E | - | 1 | 1 | - |
| Hypolepis hispaniolica Mason | Polypodiaceae | Tr | E | - | 2 | - | 1 |
| Arthrostylidium sarmentosum Pilger | Poaceae | Tr | N | - | 2 | 2 | - |
| Blechneum fragile (Liebm.) Morton & Lellinger | Blechnaceae | H | N | - | 2 | 2 | - |
| Ilex tuerckheimii Loes. | Aquifoliaceae | Ar | E | - | - | + | - |
| Cordia dependens Urb. & Ekm. | Boraginaceae | Ar | E | - | - | - | + |
| Passiflora rubra L. | Passifloraceae | Tr | N | - | - | - | + |
| Eupatorium odoratum L. | Asteraceae | Ar | N | - | - | - | + |
| Mikania cordifolia (L.) Willd. | Asteraceae | Tr | N | - | - | - | 1 |
| Psychotria liogieri Sateyerm | Rubiaceae | Ar | N | - | - | - | + |
| Marattia kaulfussii J. Smith | Marattiaceae | H | N | 1 | - | - | - |
| Asplenium radicans L. | Aspleniaceae | H | N | 1 | - | - | - |
| Smilax domingensis Willd. | Smilacaceae | Tr | N | + | - | - | - |
| Leandra limoides (Urb.) W. Judd & Skean | Melastomataceae | Ar | E | - | 1 | - | - |
| Hillia parasitica Jacq. | Rubiaceae | Tr | N | - | 2 | - | - |
| Cestrum daphnoides Griseb. | Solanaceae | Ar | E | - | 1 | - | - |
| Tibouchina longifolia (Vahl) Baill. | Melastomataceae | Ar | N | - | 1 | - | - |
| Clidemia umbellata (Miller) L.O. Wms. | Melastomataceae | Ar | N | - | - | - | + |
| Schradera subsessilis Steyermark | Rubiaceae | Tr | E | 1 | - | - | - |
| Marcgravia rubra A. Liogier | Marcgraviaceae | Tr | E | - | - | 1 | - |
| Lobelia rotundifolia Juss. | Campanulaceae | H | E | - | 1 | - | - |
| Blechnum occidentale L. | Blechnaceae | H | N | - | - | - | + |
| Cissampelos pareira L. | Menispermiaceae | Tr | N | - | - | - | + |
| Myrcia splendens (Sw.) DC. | Myrtaceae | Ar | N | - | - | - | 3 |
| Ichnanthus pallens (Sw.) Munro | Poaceae | H | N | - | - | - | 1 |
| Sagraea fuertesii (Cogn.in Urb.) Alain | Melastomataceae | Ar | E | - | 1 | - | - |
| N° rel. | 13 | 15a | 15b | |||
|---|---|---|---|---|---|---|
| N° order | DR15 | DR16 | DR17 | |||
| Altitude | 519 | 541 | 530 | |||
| Area in m2× 10 | 200 | 200 | 200 | |||
| Cover ratio In % | 75 | 100 | 100 | |||
| Xn inm. | 15 | 12 | 15 | |||
| Characteristics of the association and higher units | Family | Biotype | Status | |||
| Prestoea montana (Grah.) Nichol | Arecaceae | A | N | 3 | 4 | 4 |
| Cecropia screberiana Miq. | Moraceae | A | N | 3 | 2 | 3 |
| Alchornea latifolia Sw. | Euphorbiaceae | A | N | 2 | 5 | 4 |
| Miconia mirabilis (Aubl.) L.O. Willians | Melastomataceae | A | N | 3 | 2 | 2 |
| Miconia prasina (Sw.) DC. | Melastomataceae | A | N | 1 | 1 | 1 |
| Guarea guidonea Sleumer | Meliaceae | A | N | + | 4 | 4 |
| Cyathea arborea (L.) J.E. Smith | Cyatheaceae | A | N | 3 | 4 | 4 |
| Turpinia occidentalis (Sw.) G. Don | Staphyleaceae | A | N | 1 | + | 1 |
| Clusia rosea Jacq. | Clusiaceae | A | N | 1 | + | + |
| Ocotea globosa (Aubl.) Schlecht. & Cham. | Lauraceae | A | N | 2 | 1 | 1 |
| Casearea arborea (L.C. Rich.) Urb. | Flacourtiaceae | A | N | 1 | 1 | + |
| Oreopanax capitatus (Jacq.) Decne. & Planch. | Araliaceae | A | N | 2 | 3 | 3 |
| Didymopanax morototoni (Aubl.) Decne. & Planch | Araliaceae | A | N | 2 | 3 | 3 |
| Byrsonima spicata (Cav.) Kunth | Malpighiaceae | A | N | + | 1 | 1 |
| Buchenavia tetraphylla (Aubl.) R. A. Howard | Combretaceae | A | N | 1 | 1 | 1 |
| Sloanea berteriana Choisy | Elaeocarpaceae | A | N | 1 | 1 | 2 |
| Ormosia krugii Urb. | Fabaceae | A | N | 2 | 2 | 2 |
| Miconia serrulata (DC.) Naud. | Melastomataceae | A | N | + | + | 1 |
| Bactris plumeriana Mart. | Arecaceae | A | E | - | 1 | 1 |
| Myrsine coriacea (Sw.) R. Br. | Myrsinaceae | A | N | 1 | 1 | - |
| Ocotea leucoxylon (Sw.) Mez | Lauraceae | A | N | - | 2 | 2 |
| Inga fagifolia (L.) Willd. ex Benth. | Mimosaceae | A | N | - | + | + |
| Inga vera Willd. | Mimosaceae | A | N | - | + | + |
| Cupania americana L. | Sapindaceae | A | N | 2 | - | - |
| Hirtella triandra Sw. | Chrysobalanaceae | A | N | + | - | - |
| Miconia racemosa (Aubl.) DC. | Melastomataceae | A | N | 1 | - | - |
| Zantoxylum martinicensis (Lam.) DC. | Rutaceae | A | N | 1 | - | - |
| Guzmania monostrachya (Sw.) Rusby | Bromeliaceae | Ep | N | + | - | - |
| Microgramma piloselloides L. | Polypodiaceae | Ep | N | + | - | - |
| Companions species | ||||||
| Cnemidaria horrida (L.) K. Presl | Cyatheaceae | Ar | N | 2 | 2 | 2 |
| Pytirogramma calomelanos (L.) Link | Polypodiaceae | H | N | 1 | + | + |
| Ipomoea tiliacea (Willd.) Choisy | Convolvulaceae | Tr | N | + | 2 | 2 |
| Mucuna urens (L.) Fawc. & Rendle | Fabaceae | Tr | N | 1 | 2 | 2 |
| Solanum torvum Sw. | Solanaceae | Ar | N | 1 | 1 | 1 |
| Mikania cordifolia (L.) Willd. | Asteraceae | Tr | N | 2 | 1 | 1 |
| Psychotria domingensis Jacq. | Rubiaceae | Ar | N | - | 2 | 1 |
| Pothomorphe peltata (L.) Miquel | Piperaceae | Ar | N | - | 2 | 2 |
| Tibouchina longifolia (Vahl) Baill. | Melastomataceae | Ar | N | 1 | - | + |
| Nepsera aquatica (Aubl.) Naud. | Melastomataceae | Ar | N | 1 | - | + |
| Syngonium podophyllum Schott | Araceae | Tr | N | 2 | + | - |
| Urera baccifera (L.) Gaud. | Urticaceae | Ar | N | - | 2 | 2 |
| Psychotria uliginosa Sw. | Rubiaceae | Ar | N | - | 2 | 2 |
| Coccocypselum herbaceum Aubl. | Rubiaceae | H | N | + | - | - |
| Piper adunculum L. | Piperaceae | Ar | N | 1 | - | - |
| Cissus verticillata (L.) Nicholson & Farris | Vitaceae | Tr | N | 1 | - | - |
| Neurolaena lobata (L.) Cass. | Asteraceae | H | N | + | - | - |
| Triunfetta semitriloba Jacq. | Tiliaceae | H | N | 1 | - | - |
| Clidemia umbellata (Miller) L.O. Wms. | Melastomataceae | Ar | E | 1 | - | - |
| Gleychenia bifida (Willd.) Spreng. | Gleycheniaceae | H | N | 1 | - | - |
| Lycopodium clavatum L. | Lycopodiaceae | H | N | 1 | - | - |
| Ichnanthus pallens (Sw.) Munro | Poaceae | H | N | 1 | - | - |
| Nephrolepis multiflora (Roxb.) Jarret | Lomariopsidaceae | H | N | 1 | - | - |
| Smilax domingensis Willd. | Smilacaceae | Tr | N | + | - | - |
| Mimosa pudica L. | Mimosaceae | H | N | 1 | - | - |
| Palicourea crocea (Sw.) Schultes | Rubiaceae | Ar | N | 1 | - | - |
| Urena lobata L. | Malvaceae | Ar | N | 1 | - | - |
| Hedychium coronarium Koen. | Zingiberaceae | H | I | 1 | - | - |
| Solanum jamaicense Mill. | Solanaceae | Ar | N | 1 | - | - |
| Entada gigas (L.) Fawc. & Rendle | Fabaceae | Tr | N | + | - | - |
References
- García, R.; Mejía, M.; Peguero, B.; Jiménez, F. Flora endémica de la sierra de Bahoruco, República Dominicana. Moscosoa 2001, 12, 9–44. [Google Scholar]
- De Los Ángeles, I.; Clase, T.; Peguero, B. Flora y vegetación del Parque Nacional El Choco, Sosúa, provincia Puerto Plata, República Dominicana. Moscosoa 2005, 14, 10–55. [Google Scholar]
- Guerrero, A.; Jiménez, F.; Höner, D.; Zanoni, T. La flora y la vegetación de la loma Barbacoa, Cordillera Central, República Dominicana. Moscosoa 1997, 9, 84–116. [Google Scholar]
- Höner, D.; Jiménez, F. Flora vascular y vegetación de la loma la Herradura (Cordillera Oriental), República Dominicana. Moscosoa 1994, 8, 65–85. [Google Scholar]
- May, T. Regeneración de la Vegetación arbórea y arbustiva en un terreno de cultivos abandonado durante 12 años en la zona de bosques húmedos montanos (Reserva Científica Ébano Verde, Cordillera Central, República Dominicana). Moscosoa 1994, 8, 131–149. [Google Scholar]
- May, T. Fases tempranas de la sucesión en un bosque nublado de Magnolia pallescens después de un incendio (Loma Casabito, Reserva Científica de Ebano Verde, Cordillera Central, República Dominicana). Moscosoa 1997, 9, 117–144. [Google Scholar]
- May, T. Tres años de obervaciones fenológicas en el bosque nublado de Casabito (Reserva científica Ebano Verde, Cordillera Central, República Dominicana). Moscosoa 1998, 10, 164–178. [Google Scholar]
- May, T. Respuesta de la vegeteción en un calimetal de Dicranopteris pectinata después de un fuego, en la parte oriental de la Cordillera Central, República Dominicana. Moscosoa 2000, 11, 113–132. [Google Scholar]
- May, T. Composition, structure and diversity in broadleaved cloud forests in the Ebano Verde scientific reserve (Cordillera Central range, Dominican Republic). Moscosoa 2007, 15, 156–176. [Google Scholar]
- May, T.; Peguero, B. Vegetación y flora de la loma el Mogote, Jarabacoa, Cordillera Central, República Dominicana. Moscosoa 2000, 11, 11–37. [Google Scholar]
- Mejía, M.; García, R.; Jiménez, F. Sub-región fitogeográfica Barbacoa-Casabito: Riqueza florística y su importancia en la conservación de la flora de la isla Española. Moscosoa 2000, 11, 57–106. [Google Scholar]
- Mejía, M.; Jiménez, F. Flora y vegetación de la loma la Humeadora, Cordillera Central, República Dominicana. Moscosoa 1998, 10, 10–46. [Google Scholar]
- Mejía, M.; Pimentel, J.; García, R. Árboles y Arbustos de la región Cársica de los Haitises, República Dominicana. Moscosoa 2011, 17, 90–114. [Google Scholar]
- Veloz, A. Flora y vegetación del Monte Jota, Sierra de Bahoruco, Provincia Independencia, República Dominicana. Moscosoa 2007, 15, 206–217. [Google Scholar]
- Hager, J.; Zanoni, T. La vegetación natural de la República Dominicana: Una nueva clasificación. Moscosoa 1993, 7, 39–81. [Google Scholar]
- Rivas-Martínez, S.; Navarro, G.; Penas, A.; Costa, M. Biogeographic map. of Sourh America. A preliminary survey. Int. J. Geobot. Res. 2011, 1, 21–40. [Google Scholar] [CrossRef]
- Zanoni, T. La flora y la vegetación de loma Diego de Ocampo, Cordillera Septentrional, República Dominicana. Moscosoa 1990, 6, 19–45. [Google Scholar]
- Zanoni, T.; Mejía, M.M.; Pimentel, J.D.; García, R.G. La flora y vegetación de los Haitises, República Dominicana. Moscosoa 1990, 6, 46–97. [Google Scholar]
- Cano, E.; Cano Ortiz, A. Establishment of biogeographic areas by distributing endemic flora and habitats (Dominican Republic, Haiti, R.). In Global Advances in Biogeography; Stevens., L., Ed.; InTechOpen: Rijeka, Croatia, 2012; pp. 99–118. [Google Scholar] [CrossRef]
- Cano, E.; Veloz Ramirez, A.; Cano Ortiz, A.; Esteban Ruiz, F.J. Distribution of Central American melastomataceae: Biogeographical análisis of the Caribbean Islands. Acta Bot. Gall. 2009, 156, 527–557. [Google Scholar] [CrossRef][Green Version]
- Cano, E.; Veloz Ramirez, A.; Cano Ortiz, A. Contribution to the biogeography of the Hispaniola (Dominican Republic, Haiti). Acta Bot. Gall. 2010, 157, 581–598. [Google Scholar] [CrossRef]
- Cano, E.; Veloz Ramirez, A.; Cano Ortiz, A. Phytosociological study of the Pinus occidentalis forests in the Dominican Republic. Plant Biosyst. 2011, 145, 286–297. [Google Scholar] [CrossRef]
- Cano, E.; Cano Ortiz, A.; Del Río González, S.; Alatorre Cobos, J.; Veloz, A. Bioclimatic map of the Dominican Republic. Plant Soc. 2012, 49, 81–90. [Google Scholar] [CrossRef]
- Cano, E.; Cano Ortiz, A.; Veloz, A. Contribution to the knowledge of the edaphoxerophilous communities of the Samana Peninsula (Dominican Republic). Plant Soc. 2015, 52, 3–8. [Google Scholar] [CrossRef]
- Cano, E.; Veloz, A. Contribution to the knowledge of the plant communities of the Caribbean-Cibensean sector in the Dominican Republic. Acta Bot. Gall. 2012, 159, 201–210. [Google Scholar] [CrossRef]
- Cano Ortiz, A.; Musarella, C.M.; Piñar Fuentes, J.C.; Spampinato, G.; Veloz, A.; Cano, E. Vegetation of the dry bioclimatic areas in the Dominican Republic. Plant Biosyst. 2015, 149, 451–472. [Google Scholar] [CrossRef]
- Cano Ortiz, A.; Musarella, C.M.; Piñar Fuentes, J.C.; Pinto Gomes, C.J.; Cano, E. Distribution patterns of endemic flora to define hotspots on Hispaniola. Syst. Biodivers. 2016, 14, 261–275. [Google Scholar] [CrossRef]
- Cano Ortiz, A.; Musarella, C.M.; Cano, E. Biogeographical Areas of Hispaniola (Dominican Republic, Republic of Haiti). In Plant Ecology—Traditional Approaches to Recent Trends; Yousaf, Z., Ed.; InthechOpen: London, UK, 2017; pp. 165–189. [Google Scholar] [CrossRef]
- Cano Ortiz, A.; Musarella, C.M.; Piñar Fuentes, J.C.; Bartolomé Esteban, C.; Quinto-Canas, R.; Pinto Gomes, C.J.; Del Río González, S.; Cano, E. Advances in the knowledge of the vegetation of Hispaniola (Caribbean Central America). In Vegetation; Sebata, A., Ed.; InthechOpen: London, UK, 2018; pp. 83–99. [Google Scholar] [CrossRef]
- Cano Ortiz, A.; Musarella, C.M.; Piñar Fuentes, J.C.; Pinto Gomes, C.J.; Del Río González, S.; Cano, E. Diversity and conservation status of mangrove communities in two areas of Mesocaribea biogeographic region. Curr. Sci. India 2018, 115, 534–540. [Google Scholar] [CrossRef]
- Borhidi, A. Phytogeography and Vegetation Ecology of Cuba; Academiai Kiado: Budapest, Hungary, 1991. [Google Scholar]
- Borhidi, A. Phytogeography and Vegetation Ecology of Cuba; Academiai Kiado: Budapest, Hungary, 1996. [Google Scholar]
- Samek, V. Regiones Fitogeográficas de Cuba; Serie No 15; Academia de Ciencias de Cuba: Havana, Cuba, 1973. [Google Scholar]
- Reyes, O.J. Estudio sinecológico de las pluvisilvas submontanas sobre rocas del complejo metamórfico. Foresta Veracruzana 2005, 7, 15–22. [Google Scholar]
- Reyes, O.J.; Acosta Cantillo, F. Fitocenosis en los bosques siempre verdes de Cuba Oriental. I. Ocoteo-Phoebietum elongatae en los mogotes de la grán meseta de Guantánamo. Foresta Veracruzana 2010, 12, 9–16. [Google Scholar]
- Reyes, O.J.; Acosta Cantillo, F. Fitocenosis en los bosques siempre verdes de Cuba Oriental. II. Guareo guidoniae-Zanthoxyletum martinicensis en Sagua Baracoa. Foresta Veracruzana 2010, 12, 1–6. [Google Scholar]
- Reyes, O.J.; Acosta Cantillo, F. Fitocenosis en los bosques siempre verdes de Cuba Oriental. III. Pruno-Guareetum guidoniae en la sierra Maestra. Foresta Veracruzana 2011, 13, 1–6. [Google Scholar]
- Rivas-Martínez, S. Sinopsis biogeográfica, bioclimática y vegetacional de América del Norte. Fitosociologia 2004, 41, 19–52. [Google Scholar]
- Rivas-Martínez, S.; Loidi, J. Bioclimatology of the Iberian Peninsula. Itinera Geobot. 1999, 13, 41–47. [Google Scholar]
- Lugo, A.E.; Francis, J.K.; Frangi, J.L. Prestoea Montana (R. Graham) Nichols. Sierra Palm; US Department of Agriculte, Forest Service, Southern Forest Experimental Station: New Orleans, LA, USA, 1998; pp. 420–450. [Google Scholar]
- Rangel-Ch, J.O. La biodiversidad de Colombia: Significado y distribución regional. Rev. Acad. Colomb. Cienc. Ex. Fis. Nat. 2015, 39, 176–200. [Google Scholar] [CrossRef]
- Meier, W. Flora and Vegetation des Avila-National Parks Venezuela (Kuestenkordillere) unter Besonderer Beruecksichtigung der Nebelwaldstufe; Dissertationes Botanicae: Stuttgard, Germany, 1998.
- Rangel-Churio, J.O. La riqueza de las plantas con flores de Colombia. Caldasia 2015, 37, 279–307. [Google Scholar] [CrossRef]
- Cuatrecasas, J. Aspectos de la vegetación natural de Colombia. Rev. Acad. Col. Cs. Ex. Fis. Nat. 1958, 7, 306–312. [Google Scholar]
- Bussmann, R.W. Bosques andinos del sur de Ecuador, clasificación, regeneración y uso. Rev. Peru. Biol. 2005, 12, 203–216. [Google Scholar] [CrossRef]
- Cleef, A.M. The Vegetation of the Páramos of the Colombian Cordillera Oriental; Dissertationes Botanicae: Stuttgard, Germany, 1981. [Google Scholar]
- Cleef, A.M.; Rangel-Churio, J.O.; Van der Hammen, T.; Jaramillo, M.R. La Vegetación de las Selvas del Transecto Butirica; Van der Hammen, T., Ruiz, P.M., Eds.; Studies on Tropical Andean Ecosystems: Vaduz, Liechtenstein, 1984; Volume 2, pp. 267–406. [Google Scholar]
- Musarella, C.M. Solanum torvum Sw. (Solanaceae): A new alien species for Europe. Genet. Resour. Crop. Evol. 2020, 67, 515–522. [Google Scholar] [CrossRef]
- García, R.; Mejía, M.; Zanoni, T. Composición florística y principales asociaciones vegetales en la Reserva Científica de Evano Verde, Cordillera Central, República Dominicana. Moscosoa 1994, 8, 86–130. [Google Scholar]
- Cano Ortiz, A.; Musarella, C.M.; Quinto Canas, R.; Piñar Fuentes, J.C.; Pinto Gomes, C.J.; Cano, E. The cloud forest in the Dominican Republic: Diversity and conservation status. bioRxiv 2019. [Google Scholar] [CrossRef]
- Mollat, H.; Wagner, B.M.; Cepek, P.; Weiss, W. Mapa Geológico de la República Dominicana 1:250,000; Die Bundesanstalt für Geowissenschaften und Rohstoffe: Hannover, Germany, 2004. [Google Scholar]
- Rivas-Martínez, S.; Sánchez Mata, D.; Costa, M. North American boreal and western temperate forest vegetation. Syntaxonomical synopsis ot the potential natural plant communities of North America, II. Itinera Geobot. 1999, 12, 5–326. [Google Scholar]
- Rivas-Martínez, S. Notions on dynamic-catenal phytosocilogy as a basis of landscape science. Plant Biosyst. 2005, 139, 135–144. [Google Scholar] [CrossRef]
- Van der Maarel, E. Transformation of cover-abundance values in phytosociology and its effects on community similarity. Vegetatio 1979, 39, 97–114. [Google Scholar]
- Braun-Blanquet, J. Fitosociología: Bases Para el Estudio de las Comunidades Vegetales; Blume: Madrid, Spain, 1979. [Google Scholar]
- Gehu, J.M.; Rivas Martínez, S. Nociones Fondamentales de Phytosocilogie; Dierschke, H., Ed.; Syntaxonomie: Cramer, Vaduz, 1981; pp. 1–33. [Google Scholar]
- Weber, H.E.; Moravecm, H.; Theurillat, J.P. International Code of Phytosociological Nomenclature. J. Veg. Sc. 2000, 11, 739–768. [Google Scholar] [CrossRef]
- Liogier, A.H. La Flora de la Española; Jardín Botánico Nacional Dr. Rafael Ma. Moscoso: Santo Dominogo, República Dominicana, 1996–2000; Volume I-IX. [Google Scholar]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria; New York Botanical Garden’s Virtual Herbarium. Available online: http://sweetgum.nybg.org/ih/ (accessed on 5 May 2020).
- Hammer, Ø.; Harper, D.A.T.; Paul, D.R. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]





| Ageratina paucibracteata (Alain) King et Robins. | Micropholis polita (Griseb.) Pierre |
| Alsophila aquilina C. Chr. | Mozartia gundlachii (Kr. & Urb.) Urb. |
| Allophyllus cristalensis Lippold | Myrica cacuminis Britt and Wils. |
| Ascyrum hypericoides L. | Myrica punctata Griseb. |
| Bactris cubensis Burret | Ocotea cuneata (Griseb.) Urb. |
| Buchenavia capitata (Vahl.) Eichl. | Oplismenus hirtellus (L.) Beauv. |
| Byrsonima biflora Griseb. | Ossaea muricata (Griseb.) Wr. ex Sam. |
| Byrsonima coriacea R. Br. | Ouratea revoluta (Wr ex Griseb.) Engl. |
| Byrsonima orientensis Bisse | Panicum boliviense Hack. |
| Calophyllum utile Bisse | Panicum glutinosum Sw. |
| Calycogonium grisebachii Triana | Panicum scoparium L. |
| Calycogonium lindenianum Naud. | Pardiaea maestrensis Borhidi and Catassus |
| Calyptrantes punctata Griseb. | Pera ekmanii Urb. |
| Callicarpa ferruginea Sw. | Phaius tankervilliae (Banks) Blume |
| Cestrum laurifolium L’Hérit | Pharus latifolius L. |
| Clethra cubensis A. Rich. | Philodendron lacerum (Jacq.) Schott |
| Clidemia strigillosa (Sw.) DC. | Phyllanthus pachystylus Urb. |
| Clusia minor L. | Phyllanthus subcarnosus Wr ex Muell. Arg. |
| Clusia tetrastigma Vesque | Pilocarpus racemosus Vahl. |
| Coccocypselum x lanceolatum (Urb.) Borhidi | Pinus maestrensis Bisse |
| Coccoloba costata Wr. Inn Sauv. | Piper holguinianum Trel. |
| Coccoloba ekmanii O. C. Schmidt | Pithecellobium arboreum (L.) Urb. |
| Coccoloba retusa Griseb. | Platygine triandra Borhidi |
| Coussarea urbaniana Standl. | Pleurothalis tribuloides (Sw.) Lindl. |
| Critonia dalea (L.) DC. | Pleurothalis velaticaulis Rchb. |
| Cyathea araneosa (Sw.) Domin | Podocarpus ekmanii Urb. |
| Cyrilla nipensis Urb. | Polygala oblongata (Britt.) Blake |
| Chrysophyllum argenteum Jacq. | Polypodium phyllitidis L. |
| Dalbergaria cubensis (Urb.) Borhidi | Polystachya cubensis Schltr. |
| Dendropanax nervosus (Urb et Ekm.) A. C. Sm. | Protium cubense (Rose) Urb. |
| Dennstaedtia adiantoides (H. & B) Moore | Psidium balium Urb. |
| Desmodium herbaceum (L.) Benth. & Oerst. | Psychotria shaferi Urb. |
| Dicranopteris flexuosa (Schrad.) Mett. | Pteridium caudatum (L.) Max. |
| Dichaea hystricina Rchb. | Pteris rigida Sw. |
| Dilomilis oligophylla (Schltr.) Summerh. | Pytirogramma sulphurea (Sw.) Max. |
| Dinema cubincola (Borhidi) H. Dietr. | Rajania nipensis Howard |
| Dipholis cubensis (Griseb.) Pierre | Raudolfia salicifolia Griseb. |
| Ditta myricoides Griseb. | Rhamnidium nipense Urb. |
| Elaphoglossum firmum (Mett.) Urb. | Rondeletia calophylla Standl ex Britt. |
| Eugenia rigida Berg. | Rondeletia naguensis Britt and Wils |
| Garrya fadyenii Hook. | Rubus turquinensis Rydb. |
| Gesneria pachyclada Urb. | Salacia nipensis Britt. |
| Gesneria viridiflora (Dcne) Kuntze | Sapium cubense Britt ex Wils. |
| Graffenrieda refescens Britt. & Wils. | Sapium erythrospermum (Griseb.) Muell. Arg. |
| Guatteria moralesii Urb. | Sapium jamiaicense Sw. |
| Guettarda lindeniana A. Rich. | Savia cuneifolia Urb. |
| Habenaria monorrhiza (Sw.) Rchb. | Scleria pilosissima Britt. |
| Hedyosmum grissebachii Solms | Schradera cubensis Steyerm. |
| Heterotrichum umbellatum (Mill.) Urb. | Simaruba laevis Griseb. |
| Hyeronima nipensis Urb. | Sloanea curatellifolia Griseb. |
| Ichnanthus mayarensis (Wr.) Hitchc. | Solanum cristalense Amsh. |
| Illicium cubense A.C.Sm. | Solanum pachyneurum O.E. Schulz |
| Jacquiniella globosa (Jacq.) Schlechter | Solanum virgatum Lam. |
| Lasiacis sloanei (L.) Hitchc. | Solonia reflexa Urb. |
| Leucocroton wrightii Griseb. | Stelis ophioglossoides (Jacq.) Sw. |
| Litachne pauciflora (Swartz) P. Beauverd | Tabebuia hypoleuca Griseb. |
| Lobelia assurgens L. | Talauna minor Urb. subsp. oblongifolia (León) Borhidi |
| Lophosoria quadripinnata (Gmel.) C. Chr. | Terminalia nipensis Alain |
| Lycopodium dichotomum Jacq. | Trema cubensis Urb. |
| Lyonia calycosa (Samll) Urb. | Trichomanes crispum L. |
| Lyonia maestrensis Acuña and Roig | Vaccinium leonis Acuña & Roig |
| Magnolia cubensis Urb. | Vandenboschia scandens (L.) Copel |
| Marcgravia evenia Kr et Urb. | Vanilla phaeantha Rchb. |
| Mataiba domingensis (DC.) Radlk. | Vanilla wrightii Rchb. |
| Mecodium polyanthos (Sw.) Copel | Vernonia parvuliceps Ekm. |
| Meriania leucantha Sw. subsp. nana (Naud.) Borhidi | Viburnum villosum Sw. |
| Meringium fucoides (Sw.) Copel | Wallenia laurifolia (A. Rich.) Wr in Sauv subsp. pinetorum (Britt.) Borhidi |
| Miconia acunae Borhidi | Wigandia reflexa Brand |
| Miconia alternifolia Griseb. | Zanthoxyllum cubense P. Wils. |
| Miconia dodecandra (Desv.) Cogn. |
| Alsophila minor (D.C.Eaton) R.M.Tryon | Magnolia hamorii Howard |
| Anacheilium cochleatum (L.) Hoffm. | Magnolia pallescens Urb. & Ekm. |
| Antrophyum lanceolatum (L.) Kaulf. | Malpighia macracantha Ekm. & Nied. |
| Arthrostylidium multispicatum Pilger | Marattia kaulfussii J. Smith |
| Arthrostylidium sarmentosum Pilger | Marcgravia rubra A. Liogier |
| Asplenium radicans L. | Maxillaria coccinea (Jacq.) L.O. Wms. |
| Baccharis myrsinites (Lam.) Pers. | Mecranium ovatum Cog. |
| Bactris plumeriana Mart. | Meriania involucrata (Desv.) Naud. |
| Beilschmiedia pendula (Sw.) Hemsl. | Miconia mirabilis (Aubl.) L.O. Willians |
| Blechneum fragile (Liebm.) Morton & Lellinger | Miconia prasina (Sw.) DC. |
| Blechnum tuerckheimii A. Brause | Miconia racemosa (Aubl.) DC. |
| Bocconia frutescens L. | Microgramma piloselloides L. |
| Buchenavia tetraphylla (Aubl.) R.A. Howard | Mikania cordifolia (L.) Willd. |
| Byrsonima lucida (Mill.) L.C. Rich. | Mikania venosa A. Liogier |
| Byrsonima spicata (Cav.) Kunth | Mimosa pudica L. |
| Calyptrantes selleanus Urb. & Ekm. | Mucuna urens (L.) Fawc. & Rendle |
| Camparettia falcata Poepp. & Endl. | Myrcia deflexa (Poir) DC. |
| Casearea arborea (L.C. Rich.) Urb. | Myrsine nubicola A. Liogier |
| Cecropia screberiana Miq. | Nephrolepis multiflora (Roxb.) Jarret |
| Cestrum coelophlebium O. E. Schulz | Nepsera aquatica (Aubl.) Naud. |
| Cestrum daphnoides Griseb. | Neurolaena lobata (L.) Cass. |
| Cestrum inclusum Urb. | Niphidium crassifolium (L.) Lell. |
| Cinnamomum alainii (C.K. Allen) A. Liogier | Ocotea acarina C. K. Allen |
| Cissampelos pareira L. | Ocotea floribunda (Sw.) Mez |
| Cissus verticillata (L.) Nicholson & Farris | Ocotea foeniculacea Mez |
| Clidemia umbellata (Miller) L.O. Wms. | Ocotea globosa (Aubl.) Schlecht. & Cham. |
| Clusia clusioides (Griseb.) D’Arcy | Ocotea nemodaphne Mez |
| Cnemidaria horrida (L.) K. Presl | Ocotea patens (Sw.) Nees |
| Coccoloba wrightii Lindau | Odontadenia polyneura (Urb.) Wood. |
| Columnea domingensis (Urb.) Wiehler | Olyra latifolia L. |
| Columnea sanguinea Urb. | Oncidium variegatum (Sw.) Sw. |
| Commelina elegans Kunth | Ophioglossum palmatum L. |
| Cordia dependens Urb. & Ekm. | Oreopanax capitatus (Jacq.) Decne. & Planch. |
| Cupania americana L. | Ormosia krugii Urb. |
| Cyathea fulgens C. Chr. | Palicourea crocea (Sw.) Schultes |
| Cyathea furfuracea Baker | Passiflora rubra L. |
| Chaetocarpus domingensis Proctor | Peperomia hernandifolia (Vahl) A. Dietr. |
| Daphnosis crassifolia (Poir.) Meiss. | Persea krugii Mez |
| Dendropanax arboreus (L.) Dcne & Planch. | Persea oblongifolia Kopp. |
| Dichaea glauca (Sw.) Lindley | Phlebodium aureum (L.) J. Smith |
| Didymopanax tremulus Krug. & Urb. | Pilea geminata Urb. |
| Dilomilis montana (Sw.) Summerh. | Pinguicula casabitoana J. Jiménez |
| Diplazium hastile (Christ.) C. Chr. | Piper adunculum L. |
| Diplazium hians Kuntze | Pleurothalis ruscifolia (Jaq.) R. Br. |
| Ditta maestrensis Borhidi | Pleurothallis domingensis Cogn. |
| Elaphoglossum crinitum (L.) C. Chr. | Polygala fuertesii (Urb.) Blake |
| Elaphoglossum latifolium (Sw.) J. Sm. | Polypodium angustifolium Sw. |
| Elleanthus cephalotus Garay & Sweet | Polypodium loriceum L. |
| Entada gigas (L.) Fawc. & Rendle | Pothomorphe peltata (L.) Miquel |
| Epidendrum anceps Jacq. | Pothuya nudicaulis (L.) Regel |
| Epidendrum carpophorum Barb. Rodr. | Prestoea montana (Grah.) Nichol |
| Epidendrum jamaicense Lindl | Psychotria domingensis Jacq. |
| Epidendrum ramosum Jacq. | Psychotria liogieri Sateyerm |
| Eupatorium odoratum L. | Psychotria uliginosa Sw. |
| Exostema elliptica Griseb. | Pytirogramma calomelanos (L.) Link |
| Gleychenia bifida (Willd.) Spreng. | Renealmia jamaicensis (Gaertn.) Horan var. puberula (Gagn.) Maas |
| Gomedesia lindeniana Berg. | Rondeletia ochracea Urb. |
| Gonocalyx tetrapterus A. Liogier | Sagraea fuertesii (Cogn.in Urb.) Alain |
| Grammitis asplenifolia (L.) Proctor | Schlegelia brachyantha Griseb. |
| Guarea guidonea Sleumer | Schradera subsessilis Steyermark |
| Guatteria blainii (Griseb.) Urb. | Senecio lucens (Poir.) Urb. |
| Guzmania monostrachya (Sw.) Rusby | Sloanea berteriana Choisy |
| Gyrotaenia myriocarpa Griseb. | Smilax havanensis Jacq. |
| Hedychium coronarium Koen. | Smilax populnea Kunt var. horrida O.E. Schulz |
| Hedyosmum domingense Urb. | Solanum crotonoides Lam. |
| Hirtella triandra Sw. | Solanum jamaicense Mill. |
| Hyeronima domingensis Urb. | Solanum torvum Sw. |
| Hyeronima montana A. Liogier | Solanum virgatum Lam. |
| Hypolepis hispaniolica Mason | Stigmaphyllon emarginatum (L.) A. Juss. |
| Hyptis americana (Poir.) Briq. | Styrax ochraceus Urb. |
| Ichnanthus pallens (Sw.) Munro | Syngonium podophyllum Schott |
| Ilex tuerckheimii Loes. | Tabebuia bullata A. Gentry |
| Inga fagifolia (L.) Willd. ex Benth. | Tabebuia vinosa A. Gentry |
| Inga vera Willd. | Torralbasia cuneifolia (C. Wright) Krug. & Urb. |
| Ipomoea furcyensis Urb. | Triunfetta semitriloba Jacq. |
| Ipomoea tiliacea (Willd.) Choisy | Turpinia occidentalis (Sw.) G. Don |
| Isachne rigidifolia (Poir.) Urb. | Uncinia hamata (L.) Urb. |
| Lasianthus bahorucanus Zanoni | Urena lobata L. |
| Leandra limoides (Urb.) W. Judd & Skean | Urera baccifera (L.) Gaud. |
| Lobelia robusta Graham | Vaccinium racemosum (Vahl) Wilbur & Luteyn |
| Lobelia rotundifolia Juss. | Vernonia buxifolia (Cass.) Less. |
| Lomariopsis sorbifolia (L.) Fée | Vitis tiliifolia H. & B. ex Willd. |
| Lycopodium cernuum L. | Vittaria lineata (L.) Smith |
| Lycopodium clavatum L. | Vriesea sintenisii (Baker) L.B. Smith & Pitt. |
| Lyonia alainii W. Judd. | Vriesea tuercheimii (Mez.) L.B. Smith |
| Macrocarpaea domingensis Urb. | Zanthoxyllum martinicensis (DC.) Lam. |
| Machaerina cubensis (Kük.) T. Koyama |
| Species | As1 | As2 | As3 | As4 | P |
|---|---|---|---|---|---|
| Myrsine coriacea (Sw.) R. Br. | IV | III | V | III | 4 |
| Ocotea leucoxylon (Sw.) Mez | I | IV | III | III | 4 |
| Prestoea montana (Grah.) Nichol | I | V | V | V | 4 |
| Psychotria domingensis Jacq. | IV | IV | V | III | 4 |
| Gleychenia bifida (Willd.) Spreng. | II | I | - | I | 3 |
| Clidemia umbellata (Miller) L.O. Wms. | I | I | - | I | 3 |
| Renealmia jamaicensis (Gaertn.) Horan var. puberula (Gagn.) Maas | V | III | V | - | 3 |
| Arthrostylidium multispicatum Pilger | V | IV | III | - | 3 |
| Rondeletia ochracea Urb. | V | II | V | - | 3 |
| Didymopanax tremulus Krug. & Urb. | IV | I | V | - | 3 |
| Psychotria guadalupensis (DC.) Howard | III | III | V | - | 3 |
| Mikania venosa A. Liogier | II | IV | V | - | 3 |
| Odontosoria uncinella (Kunze) Fée | II | II | V | - | 3 |
| Brunellia comocladifolia H. & B. | II | III | III | - | 3 |
| Lobelia rotundifolia Juss. | III | I | I | - | 3 |
| Alchornea latifolia Sw. | - | III | I | V | 3 |
| Miconia mirabilis (Aubl.) L.O. Willians | - | II | I | V | 3 |
| Mucuna urens (L.) Fawc. & Rendle | - | II | I | V | 3 |
| Nephrolepis multiflora (Roxb.) Jarret | - | I | III | I | 3 |
| Ilex macfadyenii (Walp.) Rehder | V | - | I | - | 2 |
| Chionanthus domingensis Lam. | V | - | I | - | 2 |
| Macrocarpaea domingensis Urb. | IV | - | III | - | 2 |
| Polygala fuertesii (Urb.) Blake | IV | - | III | - | 2 |
| Marcgravia rubra A. Liogier | IV | - | I | - | 2 |
| Alsophila minor (D. C. Eaton) R. M. Tryon | V | III | - | - | 2 |
| Palicourea alpina (Sw.) DC. | V | II | - | - | 2 |
| Blechnum occidentale L. | III | III | - | - | 2 |
| Cyrilla racemiflora L. | IV | IV | - | - | 2 |
| Ocotea nemodaphne Mez | III | - | I | - | 2 |
| Schradera subsessilis Steyermark | II | - | I | - | 2 |
| Lycopodium clavatum L. | II | - | - | I | 2 |
| Odontadenia polyneura (urb.) Wood. | II | I | - | - | 2 |
| Byrsonima lucida (Mill.) DC. | I | II | - | - | 2 |
| Weinmannia pinnata L. | I | - | V | - | 2 |
| Epidendrum carpophorum Barb. Rodr. | I | - | I | - | 2 |
| Epidendrum carpophorum Barb. Rodr. | I | - | I | - | 2 |
| Pleurothallis domingensis Cogn. | I | II | - | - | 2 |
| Cestrum coelophlebium O. E. Schulz | I | II | - | - | 2 |
| Olyra latifolia L. | I | I | - | - | 2 |
| Cecropia screberiana Miq. | - | III | - | V | 2 |
| Turpinia occidentalis (Sw.) G. Don | - | III | - | V | 2 |
| Mikania cordifolia (L.) Willd. | - | I | - | V | 2 |
| Pothomorphe peltata (L.) Miquel | - | I | - | III | 2 |
| Ichnanthus pallens (Sw.) Munro | - | III | - | I | 2 |
| Guzmania monostrachya (Sw.) Rusby | - | II | - | I | 2 |
| Dendropanax arboreus (L.) Dcne & Planch. | - | IV | I | - | 2 |
| Dichaea glauca (Sw.) Lindley | - | III | III | - | 2 |
| Epidendrum ramosum Jacq. | - | III | I | - | 2 |
| Gomedesia lindeniana Berg. | - | II | V | - | 2 |
| Myrcia deflexa (Poir) DC. | - | II | V | - | 2 |
| Peperomia hernandifolia (Vahl) A. Dietr. | - | II | V | - | 2 |
| Vriesea tuercheimii (Mez.) L.B. Smith | - | I | V | - | 2 |
| Cyathea fulgens C. Chr. | - | I | V | - | 2 |
| Magnolia hamorii Howard | - | I | V | - | 2 |
| Mecranium ovatum Cog. | - | I | V | - | 2 |
| Lasianthus bahorucanus Zanoni | - | I | V | - | 2 |
| Nephrolepis biserrata (Sw.) Schott | - | I | V | - | 2 |
| Columnea domingensis (Urb.) Wiehler | - | I | V | - | 2 |
| Hedyosmum domingense Urb. | - | I | III | - | 2 |
| Lomariposis sorbifolia (L.) Feé | - | I | III | - | 2 |
| Beilschmiedia pendula (Sw.) Hemsl. | - | I | III | - | 2 |
| Vaccinium racemosum (Vahl) Wilbur & Luteyn | IV | I | III | - | 2 |
| Ocotea acarina C. | - | I | I | - | 2 |
| Hypolepis hispaniolica Mason | - | I | I | - | 2 |
| Schlegelia brachyantha Griseb. | - | I | I | - | 2 |
| Sagraea fuertesii (Cogn.in Urb.) Alain | - | II | I | - | 2 |
| Niphidium crassifolium (L.) Lell. | - | I | I | - | 2 |
| Phlebodium aureum (L.) J. Smith | - | I | I | - | 2 |
| Polypodium loriceum L. | - | I | I | - | 2 |
| Epidendrum jamaicense Lindl | - | I | I | - | 2 |
| Microgramma piloselloides L. | - | I | - | I | 2 |
| Miconia prasina (Sw.) DC. | - | - | III | V | 2 |
| Guarea guidonea Sleumer | - | - | I | V | 2 |
| Tibouchina longifolia (Vahl) Baill. | - | - | I | III | 2 |
| Smilax domingensis Willd. | - | - | I | I | 2 |
| Magnolia pallescens Urb. & Ekm. | V | - | - | - | 1 |
| Styrax ochraceus Urb. | V | - | - | - | 1 |
| Hyeronima montana A. Liogier | V | - | - | - | 1 |
| Cyathea furfuracea Baker | V | - | - | - | 1 |
| Clusia clusioides (Griseb.) D’Arcy | V | - | - | - | 1 |
| Ditta maestrensis Borhidi | V | - | - | - | 1 |
| Persea oblongifolia Kopp. | V | - | - | - | 1 |
| Smilax populnea Kunt var. horrida O.E. Schulz | V | - | - | - | 1 |
| Tabebuia vinosa A. Gentry | V | - | - | - | 1 |
| Gonocalyx tetrapterus A. Liogier | V | - | - | - | 1 |
| Cinnamomum alainii (C.K. Allen) A. Liogier | IV | - | - | - | 1 |
| Vriesea sintenisii (Baker) L.B. Smith & Pitt. | III | - | - | - | 1 |
| Baccharis myrsinites (Lam.) Pers. | III | - | - | - | 1 |
| Pinguicula casabitoana J. Jiménez | III | - | - | - | 1 |
| Chaetocarpus domingensis Proctor | II | - | - | - | 1 |
| Odontosoria aculeata (L.) J. Sm. | I | - | - | - | 1 |
| Myrsine nubicola A. Liogier | I | - | - | - | 1 |
| Persea krugii Mez | I | - | - | - | 1 |
| Lycopodium cernuum L. | I | - | - | - | 1 |
| Isachne rigidifolia (Poir.) Urb. | I | - | - | - | 1 |
| Machaerina cubensis (Kük.) T. Koyama | I | - | - | - | 1 |
| Vernonia buxifolia (Cass.) Less. | I | - | - | - | 1 |
| Lyonia alainii W. Judd. | I | - | - | - | 1 |
| Clidemia hirta (L.) D. don | I | - | - | - | 1 |
| Bocconia frutescens L. | I | - | - | - | 1 |
| Dilomilis montana (Sw.) Summerh. | I | - | - | - | 1 |
| Myrcia splendens (Sw.) DC. | - | IV | - | - | 1 |
| Cissampelos pareira L. | - | III | - | - | 1 |
| Uncinia hamata (L.) Urb. | - | III | - | - | 1 |
| Tabebuia bullata A. Gentry | - | III | - | - | 1 |
| Blechnum tuerckheimii A. Brause | - | III | - | - | 1 |
| Senecio lucens (Poir) Urb. | - | III | - | - | 1 |
| Coccoloba wrightii Lindau | - | III | - | - | 1 |
| Guatteria blainii (Griseb.) Urb. | - | II | - | - | 1 |
| Solanum crotonoides Lam. | - | II | - | - | 1 |
| Vitis tiliifolia H. & B. ex Willd. | - | I | - | - | 1 |
| Anacheilium cochleatum (L.) Hoffm. | - | I | - | - | 1 |
| Antrophyum lanceolatum (L.) Kaulf. | - | I | - | - | 1 |
| Camparettia falcata Poepp. & Endl. | - | I | - | - | 1 |
| Passiflora rubra L. | - | I | - | - | 1 |
| Smilax havanensis Jacq. | - | I | - | - | 1 |
| Stigmaphyllon emarginatum (L.) A. Juss. | - | I | - | - | 1 |
| Commelina elegans Kunth | - | I | - | - | 1 |
| Diplazium hastile (Christ.) C. Chr. | - | I | - | - | 1 |
| Diplazium hians Kuntze | - | I | - | - | 1 |
| Epidendrum anceps Jacq. | - | I | - | - | 1 |
| Grammitis asplenifolia (L.) Proctor | - | I | - | - | 1 |
| Jacquiniella globosa (Jacq.) Schlechter | - | I | - | - | 1 |
| Oncidium variegatum (Sw.) Sw. | - | I | - | - | 1 |
| Pothuya nudicaulis (L.) Regel | - | I | - | - | 1 |
| Vittaria lineata (L.) Smith | - | I | - | - | 1 |
| Cestrum inclusum Urb. | - | I | - | - | 1 |
| Cordia dependens Urb. & Ekm. | - | I | - | - | 1 |
| Daphnosis crassifolia (Poir.) Meiss. | - | I | - | - | 1 |
| Eupatorium odoratum L. | - | I | - | - | 1 |
| Gyrotaenia myriocarpa Griseb. | - | I | - | - | 1 |
| Hyptis americana (Poir.) Briq. | - | I | - | - | 1 |
| Lasianthus lanceolatus (Griseb.) Gómez Maza | - | I | - | - | 1 |
| Lobelia robusta Graham | - | I | - | - | 1 |
| Psychotria liogieri Sateyerm | - | I | - | - | 1 |
| Solanum virgatum Lam. | - | I | - | - | 1 |
| Pilea geminata Urb. | - | I | - | - | 1 |
| Exostema elliptica Griseb. | - | I | - | - | 1 |
| Malpighia macracantha Ekm. & Nied. | - | I | - | - | 1 |
| Ocotea floribunda (Sw.) Mez | - | I | - | - | 1 |
| Ocotea patens (Sw.) Nees | - | I | - | - | 1 |
| Ipomoea furcyensis Urb. | - | I | - | - | 1 |
| Columnea sanguinea Urb. | - | - | V | - | 1 |
| Elaphoglossum crinitum (L.) C. Chr. | - | - | V | - | 1 |
| Elaphoglossum latifolium (Sw.) J. Sm. | - | - | V | - | 1 |
| Elleanthus cephalotus Garay & Sweet | - | - | V | - | 1 |
| Pleurothalis ruscifolia (Jaq.) R. Br. | - | - | V | - | 1 |
| Hyeronima domingensis Urb. | - | - | V | - | 1 |
| Calyptrantes selleanus Urb. & Ekm. | - | - | V | - | 1 |
| Torralbasia cuneifolia (C. Wright) Krug. & Urb. | IV | - | III | - | 1 |
| Meriania involucrata (Desv.) Naud. | - | - | III | - | 1 |
| Miconia punctata (Desr.) D. Don | - | - | III | - | 1 |
| Ophioglossum palmatum L. | - | - | III | - | 1 |
| Blechneum fragile (Liebm.) Morton & Lellinger | - | - | III | - | 1 |
| Arthrostylidium sarmentosum Pilger | - | - | III | - | 1 |
| Ilex tuerckheimii Loes. | - | - | I | - | 1 |
| Leandra limoides (Urb.) W. Judd & Skean | - | - | I | - | 1 |
| Maxillaria coccinea (Jacq.) L.O. Wms. | - | - | I | - | 1 |
| Asplenium radicans L. | - | - | I | - | 1 |
| Cestrum daphnoides Griseb. | - | - | I | - | 1 |
| Polypodium angustifolium Sw. | - | - | I | - | 1 |
| Ocotea foeniculacea Mez | - | - | I | - | 1 |
| Hillia parasitica Jacq. | - | - | I | - | 1 |
| Marattia kaulfussii J. Smith | - | - | I | - | 1 |
| Buchenavia tetraphylla (Aubl.) R. A. Howard | - | - | - | V | 1 |
| Byrsonima spicata (Cav.) Kunth | - | - | - | V | 1 |
| Casearea arborea (L. C. Rich.) Urb. | - | - | - | V | 1 |
| Clusia rosea Jacq. | - | - | - | V | 1 |
| Cyathea arborea (L.) J.E. Smith | - | - | - | V | 1 |
| Didymopanax morototoni (Aubl.) Decne. & Planch | - | - | - | V | 1 |
| Pytirogramma calomelanos (L.) Link | - | - | - | V | 1 |
| Miconia serrulata (DC.) Naud. | - | - | - | V | 1 |
| Ocotea globosa (Aubl.) Schlecht. & Cham. | - | - | - | V | 1 |
| Oreopanax capitatus (Jacq.) Decne. & Planch. | - | - | - | V | 1 |
| Ormosia krugii Urb. | - | - | - | V | 1 |
| Sloanea berteriana Choisy | - | - | - | V | 1 |
| Cnemidaria horrida (L.) K. Presl | - | - | - | V | 1 |
| Solanum torvum Sw. | - | - | - | V | 1 |
| Ipomoea tiliacea (Willd.) Choisy | - | - | - | V | 1 |
| Inga fagifolia (L.) Willd. ex Benth. | - | - | - | III | 1 |
| Inga vera Willd. | - | - | - | III | 1 |
| Bactris plumeriana Mart. | - | - | - | III | 1 |
| Nepsera aquatica (Aubl.) Naud. | - | - | - | III | 1 |
| Syngonium podophyllum Schott | - | - | - | III | 1 |
| Psychotria uliginosa Sw. | - | - | - | III | 1 |
| Urera baccifera (L.) Gaud. | - | - | - | III | 1 |
| Mimosa pudica L. | - | - | - | I | 1 |
| Neurolaena lobata (L.) Cass. | - | - | - | I | 1 |
| Triunfetta semitriloba Jacq. | - | - | - | I | 1 |
| Cupania americana L. | - | - | - | I | 1 |
| Hirtella triandra Sw. | - | - | - | I | 1 |
| Miconia racemosa (Aubl.) DC. | - | - | - | I | 1 |
| Zantoxylum martinicensis (Lam.) DC. | - | - | - | I | 1 |
| Cissus verticillata (L.) Nicholson & Farris | - | - | - | I | 1 |
| Entada gigas (L.) Fawc. & Rendle | - | - | - | I | 1 |
| Palicourea crocea (Sw.) Schultes | - | - | - | I | 1 |
| Piper adunculum L. | - | - | - | I | 1 |
| Coccocypselum herbaceum Aubl. | - | - | - | I | 1 |
| Hedychium coronarium Koen. | - | - | - | I | 1 |
| Solanum jamaicense Mill. | - | - | - | I | 1 |
| Urena lobata L. | - | - | - | I | 1 |
| DR1 | DR2 | DR3 | DR4 | DR5 | DR6 | DR7 | DR8 | DR9 | DR10 | DR11 | DR12 | DR13 | DR14 | DR15 | DR16 | DR17 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shannon_T | 3.612 | 3.443 | 2.451 | 3.566 | 3.464 | 3.557 | 3.557 | 3.458 | 3.424 | 3.389 | 3.786 | 3.972 | 3.781 | 3.702 | 3.917 | 3.496 | 3.499 |
| Shannon_Ca | 3.170 | 3.247 | 2.165 | 3.097 | 3.119 | 3.152 | 2.819 | 2.606 | 2.803 | 2.563 | 3.119 | 3.154 | 3.013 | 2.683 | 3.173 | 3.061 | 3.027 |
| Shannon_Co | 2.591 | 1.718 | 1.099 | 2602 | 2.232 | 2.507 | 2.910 | 2901 | 2.655 | 2.814 | 3.066 | 3.391 | 3.162 | 3.256 | 3.277 | 2.458 | 2.532 |
| Shannon_Ne | 3.178 | 2.947 | 2.160 | 3.000 | 2.997 | 3.119 | 3.347 | 3.244 | 3.104 | 2.998 | 3.561 | 3.601 | 3.388 | 3.458 | 3.897 | 3.435 | 3.438 |
| Shannon_E | 2.574 | 2.508 | 1.089 | 2.732 | 2.490 | 2.557 | 1.891 | 1.842 | 2.137 | 2.158 | 2.410 | 2.803 | 2.658 | 2.074 | 0.000 | 0.000 | 0.000 |
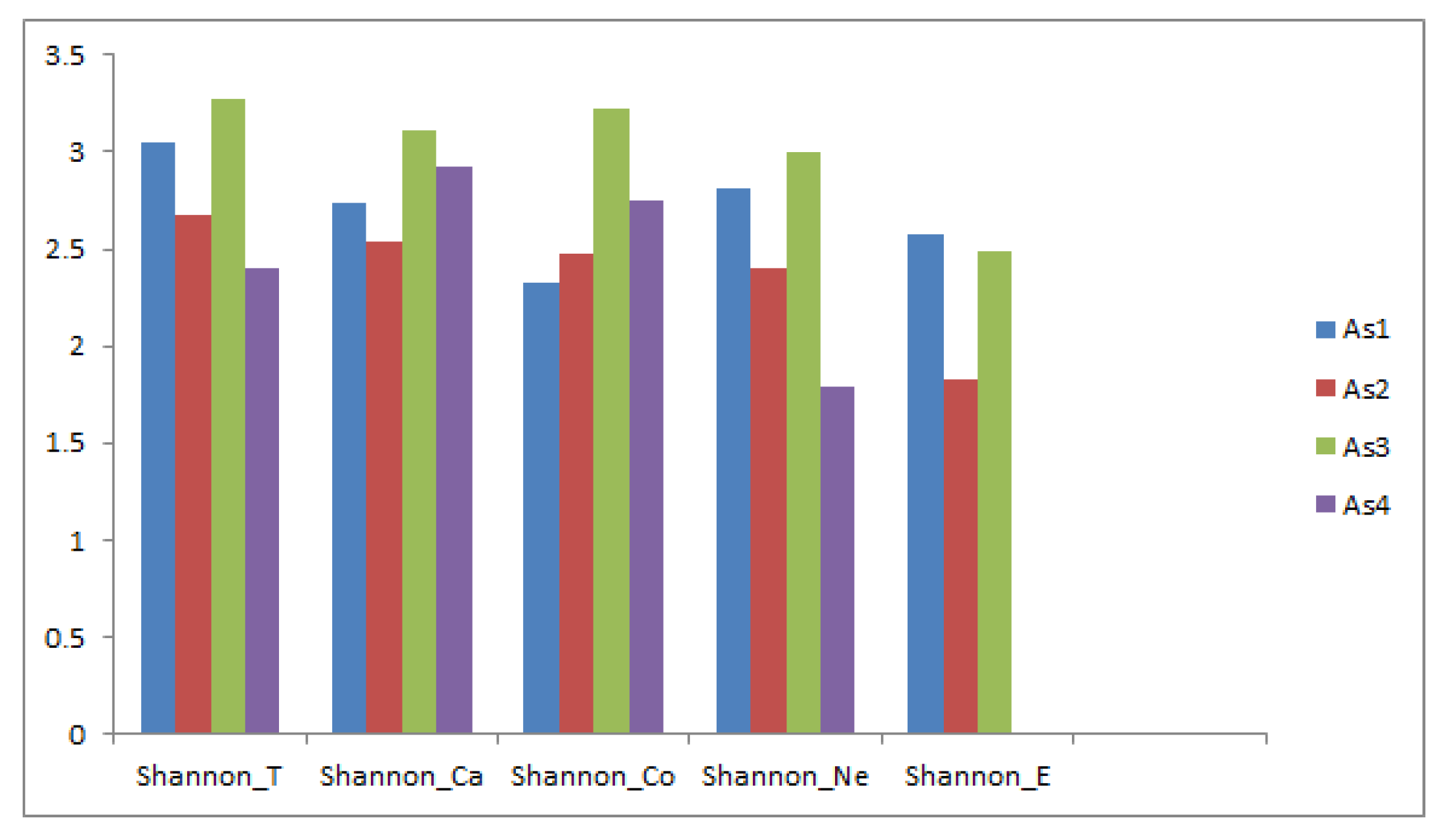
| As1 | As2 | As3 | As4 | |
|---|---|---|---|---|
| Shannon_T | 3.049 | 2.681 | 3.268 | 2.400 |
| Shannon_Ca | 2.743 | 2.533 | 3.105 | 2.921 |
| Shannon_Co | 2.330 | 2.475 | 3.218 | 2.755 |
| Shannon_Ne | 2.810 | 2.397 | 2.994 | 1.795 |
| Shannon_E | 2.572 | 1.823 | 2.486 | 0.000 |
| Plant Communities | Authors | N. of Relevés | |
|---|---|---|---|
| C1 | Clethro-Pinetum maestrensis Borhidi 1991 (Cuba) | Borhidi [31]. Table 139, page 624 | 5 |
| C2 | Hyeronimo-Sloanetum curatellifoliae Borhidi 1991 (Cuba) | Borhidi [31]. Table 140, page 627 | 5 |
| C3 | Alchorneo-Calophylletum rivularis Reyes 2005 (Cuba) | Reyes [32]. Table 1 | 6 |
| C4 | Pruno-Guareetum guidoniae Reyes & Acosta 2011 (Cuba) | Reyes & Acosta [35]. Table 2 | 4 |
| C5 | Ocoteo-Phoebietum elongatae Reyes & Acosta 2010 (Cuba) | Reyes & Acosta [33]. Table 1 | 7 |
| C6 | Guareo guidoniae-Zantoxyletum martinicensis Reyes & Acosta 2010 (Cuba) | Reyes & Acosta [34]. Table 1 | 8 |
| Total 35 relevés | |||
| DR7 | Hyeronimo montanae-Magnolietum pallescentis nova (DR) | Own relevés | 5 |
| DR8 | Cyatheo furfuracei-Prestoetum montanae nova (DR) | Own relevés | 5 |
| DR9 | Hyeronimo dominguensis-Magnolietum hamorii nova (DR) | Own relevés | 4 |
| DR10 | Ormosio krugii-Prestoetum montanae nova (DR) | Own relevés | 3 |
| DR11 | Vegetation relevés (DR) | May & Peguero [10] Table 1 page 23 | 3 |
| DR12 | Vegetation relevés (DR) | May [9] Table 1 page 171 | 12 |
| Total 32 relevés |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cano Ortiz, A.; Musarella, C.M.; Pinto Gomes, C.J.; Quinto Canas, R.; Piñar Fuentes, J.C.; Cano, E. Phytosociological Study, Diversity and Conservation Status of the Cloud Forest in the Dominican Republic. Plants 2020, 9, 741. https://doi.org/10.3390/plants9060741
Cano Ortiz A, Musarella CM, Pinto Gomes CJ, Quinto Canas R, Piñar Fuentes JC, Cano E. Phytosociological Study, Diversity and Conservation Status of the Cloud Forest in the Dominican Republic. Plants. 2020; 9(6):741. https://doi.org/10.3390/plants9060741
Chicago/Turabian StyleCano Ortiz, Ana, Carmelo Maria Musarella, Carlos José Pinto Gomes, Ricardo Quinto Canas, José Carlos Piñar Fuentes, and Eusebio Cano. 2020. "Phytosociological Study, Diversity and Conservation Status of the Cloud Forest in the Dominican Republic" Plants 9, no. 6: 741. https://doi.org/10.3390/plants9060741
APA StyleCano Ortiz, A., Musarella, C. M., Pinto Gomes, C. J., Quinto Canas, R., Piñar Fuentes, J. C., & Cano, E. (2020). Phytosociological Study, Diversity and Conservation Status of the Cloud Forest in the Dominican Republic. Plants, 9(6), 741. https://doi.org/10.3390/plants9060741