Abstract
The production of high-quality seedlings and their use in commercial planting reduce pressure on natural areas. Eugenia dysenterica DC is a native fruit tree from the Brazilian Cerrado, whose nutritional requirements are still unclear. This study aimed to evaluate the effects of nitrogen (N) and phosphorus (P) supplementation on the physiology, growth and nutrient uptake, and use efficiencies of E. dysenterica seedlings grown in glasshouse conditions. The following rates were used in separate experiments: 0, 50, 100, 200, and 400 mg dm−3 N and 0, 100, 200, 400, and 600 mg dm−3 P. The experiment was conducted in a randomized block with four replications. The lowest N rate (50 mg dm−3) increased the stomatal conductance (gS) and, consequently, resulted in the highest transpiration (E), electron transport (ETR), and photosynthetic (A) rates. Also, rates of 50 mg dm−3 and 100 mg dm−3 N increased the Root Uptake Efficiency (RUE) and plant Nutrient Use Efficiency (NUE) for macronutrients and the RUE for micronutrients, stimulating plant growth. Phosphorous fertilization resulted in the maximum values for photosynthesis, electron transport rate, total dry mass, and NUE at the 200 mg dm−3 rate. The results of this study suggest that fertilization with 50 mg dm−3 N and 200 mg dm−3 P is suitable for the development of E. dysenterica seedlings.
1. Introduction
The Cerrado is a Brazilian savanna ecosystem that has a high diversity of flora and fauna and originally covered 200 million hectares; in recent years, more than 50% of its area has been converted to grain crops and pastures [1]. Eugenia dysenterica DC, popularly known as cagaita, is a native fruit of the Brazilian Cerradoin the Myrtaceae family. It is an economically important species because its fruits can be used as food, having high levels of vitamin A and C [2], and in the pharmaceutical and cosmetics industries [3]; the trees are used to supply cork and timber for the construction industry as well as for firewood [4].
According to the regulations in the Brazilian Forestry Code (Law No. 12651 of 25 May 2012), the collection of non-timber forest products in legal reserve areas is permitted only for the purpose of subsistence farming. The law reduces the commercial exploitation of native plants because of threats to biodiversity and the number of conserved areas in the Cerrado biome. Thus, as an alternative, seedling production in orchards and their planting for commercial purposes will reduce native area exploitation. To accomplish this, more information is needed, such as the nutritional requirements of the Cerrado´s native species.
Several arboreal species of the Brazilian savanna, including cagaita, are well adapted to low-fertility soils, particularly those with high acidity, high aluminum content [5], and water deficit conditions in some months of the year; these are all typical characteristics of the Cerrado region [6]. However, studies have shown variations in the productivity of E. dysenterica [7], such as reduced fruit yield after the first five years under natural conditions [8]. Among the primary factors affecting fruit development, inconsistent production was highly correlated with low nutritional levels in the leaves. This suggests that fertilization is an important factor in the production of E. dysenterica fruits.
Appropriate nutrition allows adequate growth and increases the resistance of plants to attacks from pathogens [9]; it also contributes to the allocation of photoassimilatesto fruit development and the standardization of fruit production. Therefore, studies on the nutritional requirements of this species are important to stimulate faster and more uniform plant growth. Considering this need, it is important to highlight that nitrogen (N) and phosphorus (P) are macronutrients required in large quantities by plants during their initial development [10]. These nutrients are often present in limiting amounts in Cerrado soils. Nitrogen availability is often correlated with soil organic matter concentrations [11]. Cerrado soils tend to have minimal organic matter and, thus, low natural plant available N [6]. Plant P availability is strongly correlated with soil pH and mineralogy (including P quality) [12]. The natural P availability for plants tends to be low in these soils due to the presence of iron and aluminum oxides in the clay fraction and the low activity of clay minerals such as kaolinite [6].
Based on this background, the objective of the present study was to determine adequate rates of N and P for E. dysenterica DC seedling production, based on the physiological and morphological traits and nutrient uptake and use efficiencies of seedlings produced in Cerrado soil in glasshouse conditions.
2. Results
2.1. Physiological Traits
The N (0, 50, 100, 200 and 400 mg dm−3) and P (0, 100, 200, 400 and 600 mg dm−3) rates used in this study differentially influenced gas exchange and chlorophyll a fluorescence traits in E. dysenterica seedlings. The photosynthetic rate (A), stomatal conductance (gS) and transpiration (E) were lowest in plants grown in the absence of N or at the highest N rate (400 mg dm−3) (Figure 1). Each of these measured parameters showed a dramatic initial increase with the first increment of N fertilizer (50 mg dm−3) and then dropped at the highest rate (400 mg dm−3). Similarly, the P response increased with the first increment of fertilizer (100 mg dm−3 P); however, it differed as the response plateaued with increasing P rates (Figure 1).
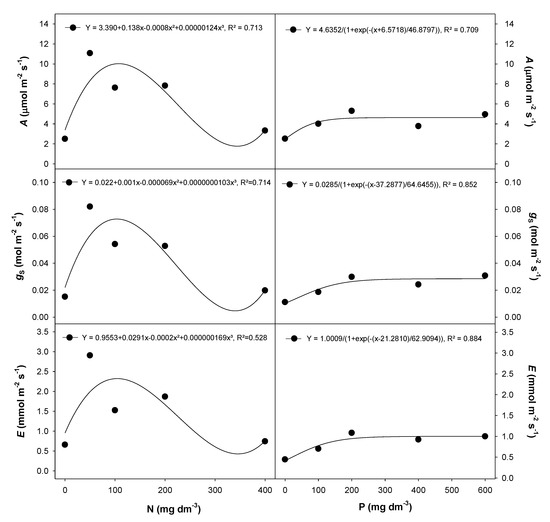
Figure 1.
Gas exchange traits in E. dysenterica seedlings grown at varying rates of nitrogen and phosphorus. Photosynthetic rate (A), stomatal conductance (gS) and transpiration rate (E), in Eugenia dysenterica DC seedlings grown at varying rates of nitrogen (N) and phosphorus (P) for 278 d. Dots represent mean (n = 4). Means and SEM are shown as Supplementary Material (Table S1).
E. dysenterica cultivation at various N and P rates did not change the initial chlorophyll fluorescence (F0, ~306.2) or the potential quantum yield of PSII (Fv/Fm, ~0.844) (data shown as supplementary material; Table S2). The effective quantum yield of PSII (ΦPSII) and the electron transport rate (ETR) were higher in plants grown with between 50 to 200 mg dm−3 N and 200 mg dm−3 P (Figure 2). The ΦNPQ showed a dramatic increase in E. dysenterica plants exposed to 400 mg dm−3 N and was reduced in plants exposed to 600 mg dm−3 P (Figure 2).
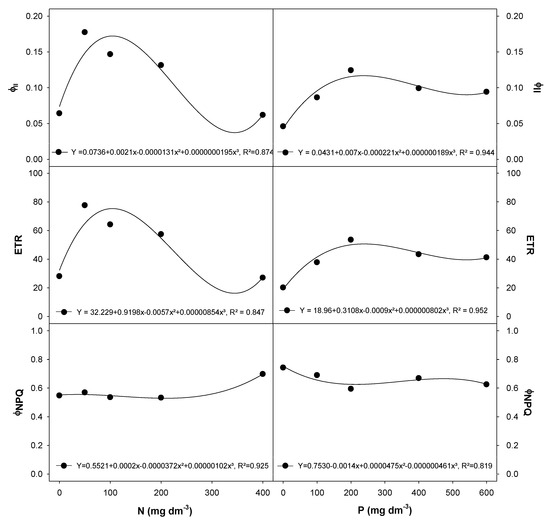
Figure 2.
Chlorophyll a fluorescence traits in E. dysenterica seedlings grown at varying rates of nitrogen and phosphorus. Effective quantum yield of photosystem II (ΦPSII), electron transport rate (ETR) and regulated quantum yield of nonphotochemical energy dissipation (ΦNPQ) in Eugenia dysenterica DC seedlings grown at varying rates of nitrogen (N) and phosphorus (P) for 278 d. Dots represent mean (n = 4). Means and SEM are shown as Supplementary Material (Table S2).
2.2. Morphological Traits
The highest values for height (H) and stem diameter (SD) (Figure 3), and leaf number and root length (data shown as Supplementary Material; Table S3) were observed at 50 to 100 mg dm−3 N. The peak values were 6.6 LN and 70.8 cm RL. However, N application at higher rates (>200 mg dm−3) negatively affected the H and SD in E. dysenterica (Figure 3). The total dry matter (TDM) in plants grown at 200 and 400 mg dm−3 was 7% and 52% lower, respectively, than that in the 50 mg dm−3 N treatment (data shown as Supplementary Material; Table S3). The ratio between shoot dry matter (SDM) and root dry matter (RDM) (SDM/RDM) increased only at the highest N rate (Figure 3).
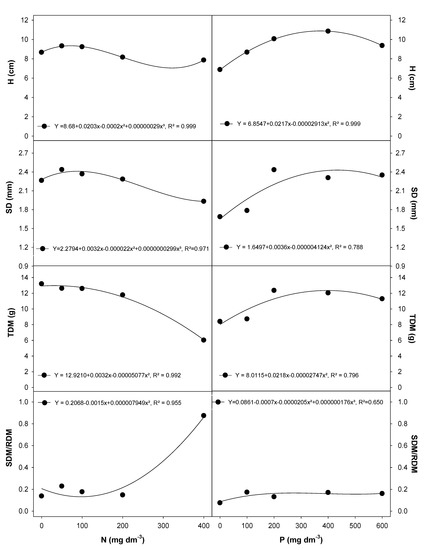
Figure 3.
Morphological traits in E. dysenterica seedlings grown at varying rates of nitrogen and phosphorus. Height (H), stem diameter (SD), total dry matter (TDM), and the ratio between shoot dry matter and root dry matter (SDM/RDM) in Eugenia dysenterica DC seedlings grown at varying rates of nitrogen (N) and phosphorus (P) for 278 d. Dots represent mean (n = 4). Means and SEM are shown as Supplementary Material (Table S3).
2.3. Contribution of Nitrogen and Phosphorous Rates to Physiological and Morphological Changes in Eugenia dysenterica Seedlings
The contributions of N and P doses to physiological and morphological traits were determined by principal component analysis (PCA). The first two main components (PC1 and PC2) explained 67.0% and 64.4% of the total data variation for the N and P rates, respectively (Figure 4). It was observed that the variables A, gS, E, ΦPSII, ETR, RDM, TDM and SDM madea higher contribution toPC1 (Figure 4A). At the rate of 50 mg dm−3 N, E. dysenterica seedlings presented higher values of A, gS, E, ΦPSII, ETR and SDM than in other treatments (Figure 4A), and the physiological variables showed higher values than the morphological variables.
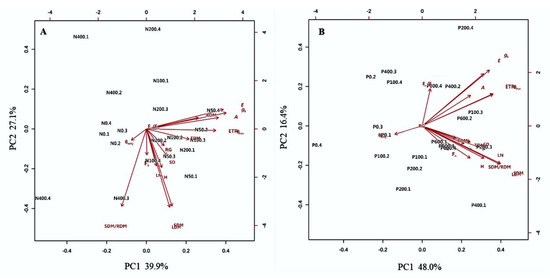
Figure 4.
Principal component analysis (PCA) of the physiological and morphological parameters of E. dysenterica at varying nitrogen and phosphorus rates. Principal component analysis (PCA) of the mean values for the physiological and morphological characteristics of Eugenia dysenterica DC grown at varying (A) nitrogen (N; 0, 50, 100, 200, and 400 mg dm−3) and (B) phosphorus (P; 0, 100, 200, 400, and 600 mg dm−3) rates for 278 d. Images A and B show score variations in these traits along the first two main component axes, with the percentage of variation explained.
Phosphorus positively influenced all physiological and morphological traits of E. dysenterica (Figure 4B). However, it was observed that the 200 mg dm−3 rate of P resulted in the highest values of A, gS, E, ΦPSII and ETR. The PC1 also indicated that LN, SDM/RDM, and SDM were highly correlated with 200 mg dm−3 P.
2.4. Nitrogen and Phosphorous Uptake and Use Efficiency
The Root Uptake Efficiency (RUE) and plant Nutrient Use Efficiency (NUE) for macro- and micronutrients in E. dysenterica seedlings were differentially affected by the availability of N and P in the soil (Figure 5, Figure 6 and Figure 7). The increase in the N rate positively affected the RUE-N and RUE-Mg at the highest rate. RUE-P and RUE-S were stimulated from 200 mg dm−3 N, and RUE-Ca, RUE-Cu, and RUE-Fe were stimulated from the lowest rate (50 mg dm−3 N). Even less responsive, NUE-Ca and NUE-Mg decreased from 200 mg dm−3 N, and NUE-Cu and NUE-Zn were reduced at all doses. The NUE of the other macronutrients was not affected by the increasing N rate.
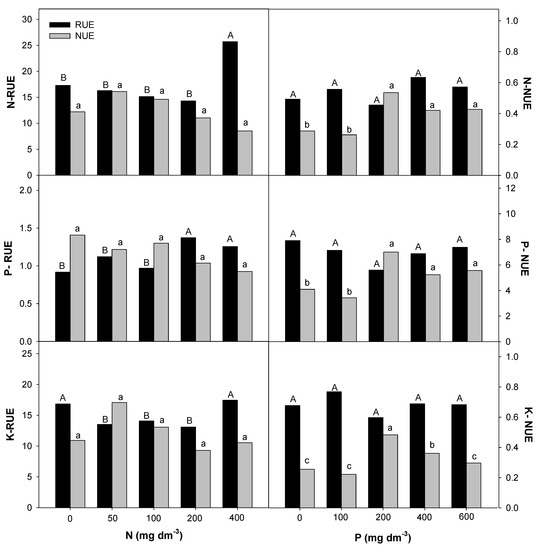
Figure 5.
Root uptake efficiency (RUE) and nutrient use efficiency (NUE) for nitrogen (N), phosphorus (P), and potassium (K) in E. dysenterica seedlings. Root uptake efficiency (RUE, mg g−1) and plant nutrient use efficiency (NUE, g2 mg−1) for the macronutrients N, P, and K in Eugenia dysenterica DC seedlings grown at varying rates of nitrogen and phosphorus for 278 d. Bars represent means (n = 4). Means followed by the same letters, uppercase for RUE and lowercase for NUE, do not differ by the Scott-Knott clustering test (p > 0.05).
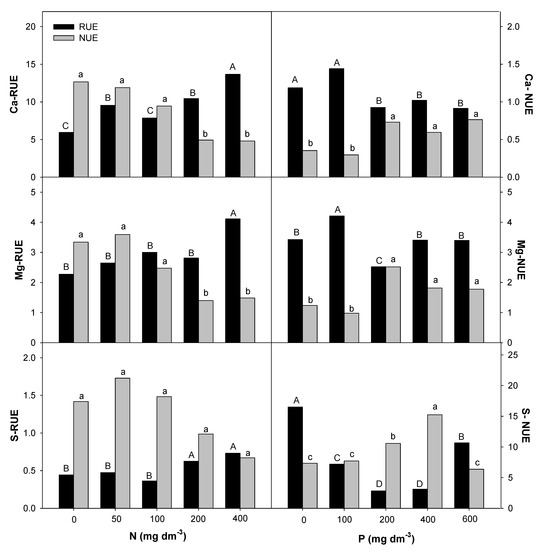
Figure 6.
Root uptake efficiency (RUE) and nutrient use efficiency (NUE) for calcium (Ca), magnesium (Mg), and sulfur (S) in E. dysenterica seedlings. Root uptake efficiency (RUE, mg g−1) and plant nutrient use efficiency (NUE, g2 mg−1) for the macronutrients Ca, Mg, and S in Eugenia dysenterica DC seedlings grown at varying rates of nitrogen and phosphorus for 278 d. Bars represent means (n = 4). Means followed by the same letters, uppercase for RUE and lowercase for NUE, do not differ by the Scott-Knott clustering test (p > 0.05).
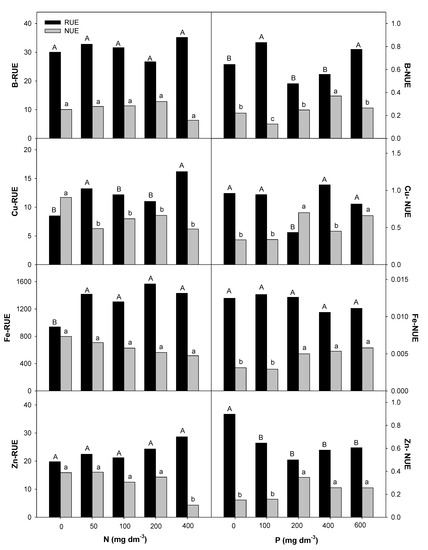
Figure 7.
Root uptake efficiency (RUE) and nutrient use efficiency (NUE) for micronutrients in E. dysenterica seedlings. Root uptake efficiency (RUE, mg g−1) and plant nutrient use efficiency (NUE, g2 g−1) for the micronutrients boron (B), copper (Cu), iron (Fe), and zinc (Zn) in Eugenia dysenterica DC seedlings grown at varying rates of nitrogen and phosphorus for 278 d. Bars represent means (n = 4). Means followed by the same letters, uppercase for RUE and lowercase for NUE, do not differ by the Scott-Knott clustering test (p > 0.05).
P availability positively affected RUE-Mg and RUE-B at the lowest rate (100 mg dm−3) in E. dysenterica plants. RUE-Ca was reduced starting from 100 mg dm−3 P, and RUE-S and RUE-Zn were reduced at all P rates. The uptake efficiencies of the other nutrients evaluated in E. dysenterica plants were not affected by the varying doses of P in the soil (Figure 6). The NUE values for most nutrients (N, P, K, Ca, Mg, S, Cu, Fe, Zn) were higher at 200 mg dm−3 P, that at other rates but did not differ significantly between the higher rates (400 and 600 mg dm−3 P) (Figure 5, Figure 6 and Figure 7).
3. Discussion
3.1. Nitrogen Fertilization Increased Photosynthetic Rates of Eugenia dysenterica
The N and P play key roles in the photochemical and biochemical stages of photosynthesis because they are constituents of photosynthetic process-related proteins, including photosynthetic pigments [13] and the enzyme RuBisCO [14]. Changes in RuBisCO activity can occur either because of changes in the levels of P in the plant that affect the allocation of N to RuBisCO [15] or because N deficiency can hinder the uptake of P and induce changes in RuBisCO activity [14]. The availability of N in the soil contributed to the higher photosynthetic activity in the E. dysenterica seedlings. However, we observed that the highest values for the photosynthetic rate and photochemical processes occurred in plants that were cultivated at the lowest rate of N (50 mg dm−3), demonstrating the low requirement for this nutrient in the early stages of E. dysenterica development.
This trend was also observed in Passiflora alata seedlings that obtained maximum photosynthetic values at low N doses (up to 146 mg kg−1) 176 d after staking [16]. It has been reported that tree plants exhibit increased N demand with increasing age and during fruit production [17], and two main sources of N that fruit trees use for their vegetative growth and reproduction are root N uptake and internal N cycling. The N available for root uptake can be derived from mineral fertilizers or from the mineralization of natural soil N. Plant roots are able to take up the available N forms, i.e., nitrate (NO3−) and ammonium (NH4+) [11,18].
The rates of P used in the present study significantly altered the photosynthetic activity of E. dysenterica seedlings. The photosynthetic rate (A), stomatal conductance (gS), transpiration rate (E), electron transport rate (ETR), and PSII quantum yield (ΦPSII) were responsive to 200 mg dm−3 P. The P availability can stimulate stoma production by epidermal cells, and consequently increase stomatal conductance and photosynthesis [19]. A higher stomatal density was observed by Seika and Yano [20] in Vigna sinensis subjected to 450 mg of P fertilizer. On the other hand, Salter et al. [21] found that native plants grown under P-limited conditions showed impaired photosynthesis due to reduced stomatal conductance. Moreover, the increase in photosynthetic rates and photochemical processes observed in E. dysenterica in both N and P treatments was accompanied by lower nonphotochemical energy dissipation (ΦNPQ), suggesting increased thermal dissipation related to the xanthophyll cycle [22] as an alternative sink for excess energy under stress conditions.
The contribution of phosphorous to the photosynthetic process is related to its function as a substrate for ATP synthesis in the chloroplast stroma, whereas inorganic P deficiency caused by low P availability reduces ATP production in the stroma and CO2 fixation [23]. In addition to maintaining plant development, high photosynthetic rates are expected to result in higher fruit yield and uniformity. This occurs because the yield per plant is directly related to the production of soluble solids [24] and fruits are the main sinks of photoassimilates during the reproductive stage.
3.2. EugeniaDysenterica Growth Was Stimulated by Phosphate Fertilization
Adequate N and P are important nutrients for the initial development of tree plants [25]. The N is required throughout plant growth, and N deficiency is considered one of the most limiting factors for plant growth [11,26]. Besides a lack of fertilization, this nutrient deficiency can also be related to the form of N absorbed by plants (mostly nitrate), which is readily leachable. To favor N acquisition and mobilization, the stimulation of root development and plant biomass production should be promoted [12,27].
P is recognized as an important component of plants as an energy carrier and in nucleic acids and signaling pathway proteins (such as protein kinases) [28]. P is the main limiting element for Eucalyptus grandis seedling growth [29] and is critical for the development of slow-growing tree species [30,31].
An increase in the height and stem diameter of Cedrela fissilis seedlings, which also occur in the Cerrado biome, was promoted by increased rates of N (40 to 160 mg dm−3) [32]. Nevertheless, in the present study, the increase in N fertilization up to 200 mg dm−3 did not affect the growth of the young E. dysenterica plants under glasshouse conditions. As observed for E. dysenterica, mature watermelon plants provided with a high N supply also showed a lower performance in root growth than plants grown under low N conditions [33]. Similarly, previous studies reported that the morphological traits of Talisia esculenta [25] and E. dysenterica [34], particularly height and stem diameter, were not responsive to N doses under controlled conditions. This suggests that this species has low nutritional requirements for N in the earlier stages of development.
On the other hand, the cultivation of plants at the highest N rate (400 mg dm−3) had negative effects on the growth of young E. dysenterica after 278 d of cultivation. Similar behavior in growth and dry mass production was observed at doses higher than 504 mg dm−3 N in Talisia esculenta seedlings [25] and at doses of 200 mg dm−3 N in Annona crassiflora [35].
Unlike N fertilization, P fertilization from the rate of 100 mg dm−3 significantly influenced the height of E. dysenterica, while rates from 200 mg dm−3 P increased the stem diameter. This demonstrates the role of P in shoot biomass production in E. dysenterica. Similarly, Araucaria angustifolia was shown to be responsive to increased P availability in terms of height, stem diameter, and shoot dry matter, with higher values at 472 mg dm−3 P fertilization [36]. Bessa et al. [37] also observed a high correlation between plant growth traits and the availability of N, P, K, and S for E. dysenterica seedlings grown in hydroponic solution for 190 d.
The SDM/RDM of E. dysenterica was not influenced by the N rates (0 to 200 mg dm−3) and showed higher values in P-fertilized plants, than in the absence of added P. The higher translocation of photoassimilates correlated with auxin accumulation [38] is a characteristic behavior of Cerrado species that are well adapted to nutrient-limiting conditions [39]. An important aspect to be considered when evaluating the response of E. dysenterica seedlings to nutrient rates is related to the morphology and initial establishment of the species. The interleaved periods of root and shoot development observed in the growth of native plants [39] may influence the biometric and nutritional responses of the seedlings.
3.3. Effect of Nitrogen and Phosphorous Rates on Root Uptake and Plant Nutrient Use Efficiencies
Studies on the uptake and use efficiencies of native species are rare; however, attention has been paid to the importance of N and P in the initial development of these plants [40,41]. The interaction of N with other nutrients varies with environmental conditions (soil and climate), species, and genotype, and is affected by the forms of N absorbed by plants (N-NO3− or N-NH4+) [42]. The N-RUE in E. dysenterica seedlings was responsive up to the highest evaluated rate (400 mg dm−3 N); however, the seedlings showed lower NUE under this condition, possibly suggesting a negative effect on the dry matter production of the species. Nevertheless, it has been reported that N redistribution in trees occurs seasonally [26], and a higher N requirement can begin after two years of cultivation [43].
The uptake and use efficiencies for P were affected by the gradual increase in the supply of N. This is important because soils with a high level of iron and aluminum oxides, such as Cerrado soils, preferentially present positive charges that are capable of adsorbing various anions, such as phosphate ions, making P less available to plants [12,44]. Even if it has not been observed in E. dysenterica seedlings, the positive interaction between N and K has been described for crops [42], because K plays an important role as a counterion to nitrate (NO3−) transport in xylem [45].
The N availability favored the uptake efficiency of Ca, Mg and S. The transport of nitrate ions in plasma membranes requires the activity of proton pumps (H+-ATP-ase) that depend on Mg as an enzyme cofactor [46]. The Ca is taken up by the root apical zone; however, it is immobile in the phloem [47]. This nutrient is required during cell division and cell wall synthesis and is therefore important for plant development [48]. Like N, S is highly mobile in soil and is taken up mainly in the form of sulfate (SO42−) [47]. In plant chloroplasts, S is a component of nitrite reductase, an enzyme responsible for reducing NO2− to NH3 [49], where nitrite is an intermediate product that is toxic to plants.
Unlike for N, the availability of various P rates had little effect on the nutrient uptake efficiency of E. dysenterica; however, it increased the plant nutrient use efficiency, especially at 200 mg dm−3 P. Trees have been reported to store P in biomass as a long-term adaptive mechanism [31]. According to these authors, in slow-growing species, there is also a tendency to accumulate more P than N to partially offset the gradual decrease in P in the soil.
The nutrient K plays an important role in stomatal control and cell expansion [50]. It has also been observed that K use efficiency is directly related to sugar accumulation and, consequently, fruit development and quality [51,52]. The higher K-NUE in E. dysenterica was observed at rates of 50 mg dm−3 N and 200 mg dm−3 P. These results were similar to those observed in Stryphnodendron polyphyllum, a native Cerrado species that showed higher NUE-K with a supply of 62 mg dm−3 N and 125 mg dm−3 P [40].
The interrelationship between P and Mg occurs in various metabolic processes in plants because Mg is required by the phosphorylative enzymes that incorporate and transfer inorganic P, in addition to being responsible for forming cross-bridges between these enzymes and ATP and ADP [9]. As observed in E. dysenterica, the highest NUE-Mg was found under 200 mg dm−3 P. A similar interaction was observed for the Cerrado species Jacaranda decurrens subsp. symmetrifoliolata, in which P addition increased the Mg concentration in the roots of these plants [53]. P is the most limiting nutrient with respect to growth and biomass production in E. dysenterica seedlings [34]; in the present work, 200 mg dm−3 P caused increases in macro- and micronutrient use efficiency.
Like macronutrients, micronutrients, despite being required in smaller quantities, had their uptake and use efficiency affected by the availability of N and P in the soil. Cerrado soils have low availability of boron (B), copper (Cu), molybdenum (Mo), and zinc (Zn) [6], possibly compromising the development of native trees [54] and affecting fruit production. In these soils, pH correction by liming may decrease the availability of micronutrients such as Mn, Cu, and Zn [3,55], highlighting the importance of understanding micronutrient uptake and use efficiencies for E. dysenterica seedling production.
The availability of N favored Cu, Fe, and Zn uptake in E. dysenterica seedlings. The interactions of N with micronutrients occur because of changes in rhizosphere pH values and the N forms used. N available as nitrate (NO3−) causes increased rhizosphere pH, with a reduction in micronutrient uptake, while ammonium N (NH4+) results in decreased rhizosphere pH values and consequently higher micronutrient uptake [42]. Therefore, the use of ammoniacal sources may favor the uptake of micronutrients in E. dysenterica. Similarly, Carlisle et al. [56] found that the supply of ammoniacal N resulted in increased Fe, Cu, and Zn concentrations in wheat plants. Even at higher uptake efficiencies, the use efficiency of these nutrients was lower in E. dysenterica seedlings, under a high soil N rate (600 mg dm−3).
The RUE-B was not affected by the varying rates of N and increased at 100 mg dm−3 in soil. Liming generally decreases B because the formation of Al(OH)3 by Al3+ precipitation in solution allows the uptake of large amounts of B [6]. The NUE-B for E. dysenterica seedlings was lower with N availability and greater with the supply of 400 mg dm−3 P. B has low mobility in plants but is essential because it influences Ca2+ utilization and cell membrane integrity [57]; this is because it is associated with the pectin content in the cell wall [58].
The highest RUE-Cu was observed at the lowest N rate (50 mg dm−3) but higher NUE-Cu was observed when the plants were exposed to increasing P rates. This emphasizes the importance of phosphate fertilization for micronutrient use efficiency in E. dysenterica seedlings. Exchangeable Cu is adsorbed by organic matter. In the Cerrado scenario, with low organic matter content and low pH, exchangeable Cu is expected to be prevalent and available to plants, as observed in the appropriate Cu levels in various tree species [59]. Cu is a constituent of important oxidase enzymes, including cytochrome oxidase, ascorbic acid oxidase, and lactase, and is involved in the photosynthetic process [42].
The highest NUE-Zn was observed at 200 mg P dm−3 in E. dysenterica seedlings. Increasing P rates also provided higher NUE-Zn in Stryphnodendron polyphyllum seedlings [40]. These data suggest that micronutrient retranslocation depends on an adequate supply of P, which promotes uniform growth and favors fruit production.
We observed favorable responses in the physiological and morphological traits and nutrient use efficiency of E. dysenterica seedlings grown under 200 mg dm−3 P and 50 mg dm−3 N, with P being more limiting to the initial development of this species. Multivariate analysis allowed us to verify that both physiological and morphological assessments were important in the investigation. Our data also demonstrate the importance of fertilization in seedling production programs for commercial plantations, the consolidation of seed orchards and even the use of these species in restoration programs in degraded areas. It is important to favor more uniform fruit production, as non-uniform fruit is a common problem under natural conditions in the Cerrado [7].
4. Materials and Methods
4.1. Plant Material and Growth Conditions
Ripe E. dysenterica fruits were collected from plants located in the municipality of Montes Claros de Goiás, GO, Brazil (19°53′ S latitude, 44°25′ W longitude and 592 m altitude). The specimens were deposited in the herbarium of the Goiano Federal Institute (IF Goiano), Campus Rio Verde (n. 630/2017). After collection, the fruits were homogenized and processed to obtain the seeds.
For both studies, a clayey red Latosol was collected from the 20–40 cm soil layer in a native area of the IF Goiano. The soil had the following chemical attributes, determined according to Silva et al. [60]: pH (CaCl2) = 4.3; organic matter = 27 g kg−1; P (Mehlich-1) = 1.0 mg dm−3; K = 0.11 cmolc dm−3; Ca = 0.4 cmolc dm−3; Mg = 0.1 cmolc dm−3; Al = 0.3 cmolc dm−3; H + Al = 4.5 cmolc dm−3; aluminum saturation = 33% and base saturation (V) = 12%.Based on these values, the soil was incubated (for 20 d) with dolomitic limestone (PRNT 70%), adopting the basis saturation criterion (V%) to 60%.
The experiments were carried out in pots containing 2.8 dm3 of the soil, which was previously subjected to basic fertilization [61] with 100 mg dm−3 potassium (K), 40 mg dm−3 sulfur (S), 1.33 mg dm−3 copper (Cu), 0.81 mg dm−3 boron (B), 4 mg dm−3 zinc (Zn), 3 mg dm−3 manganese (Mn), and 0.15 mg dm−3 molybdenum (Mo). Calcium (Ca) and magnesium (Mg) were supplied by liming. The salts used in the fertilizer solutions were KCl, MnSO4, ZnSO4, CuSO4, H3BO3, and MoO3. After fertilization, four cagaita seeds were added per pot. At 30 d after germination, thinning was performed, retaining two plants per pot. The criteria for plant selection involved uniformity of morphological features, i.e., two expanded pairs of leaves, the absence of contamination (fungi), and adequate spacing in the pot. During the experimental period, the soil moisture was checked every two days and maintained well irrigated with deionized water when necessary.
The experiments were carried out in a glasshouse of the Plant Tissue Culture Laboratory of IF Goiano, Rio Verde, Goiás, Brazil. Temperature and humidity conditions during the experimental period were recorded using a data logger (Novus, model LogBox-RHT-LCD, Porto Alegre, RS, Brazil) and are presented in Figure 8.
Figure 8.
Glasshouse temperature (°C) and relative humidity (%) conditions during the experimental period. Temperature (°C) maximum, medium, and minimum (A); and relative humidity (%) maximum, medium, and minimum (B) in the glasshouse during the experimental period.
4.2. Treatments and Experimental Design
Two independent experiments were conducted, one for each nutrient evaluated. For the treatments corresponding to rates of N and P, the nutrients were not added to the basic fertilizer previously mentioned.
The experimental design was completely randomized, with five dosages for each nutrient (N and P) and four replicates, with 20 pots per experiment. The N and P rates in the treatments were added separately, using the salts NH4SO4, NH4Cl, NH4(H2PO4), and CaHPO4, corresponding to: (i) N: 0, 50, 100, 200, and 400 mg dm−3; and (ii) P: 0, 100, 200, 400, and 600 mg dm−3.
4.3. Evaluations
The experiments lasted 278 d. Then, the physiological and morphological traits were measured, and samples were collected to analyze their nutritional content.
4.3.1. Physiological Traits
Gas exchangeparameters were evaluated in the last completely expanded leaves to measure the photosynthetic rate (A, µmol m−2 s−1), stomatal conductance (gS, mol m−2 s−1) and transpiration rate (E, mmol m−2 s−1). The evaluations were carried out between 9:00 and 11:00 am using an Infrared Gas Analyzer (IRGA; LI-6400XT, Li-Cor, Lincoln, NE, USA), under constant photosynthetically active radiation (PAR, 1000 μmol of photons m−2 s−1) and the atmospheric concentration of CO2 (~405 μmol mol−1), temperature (~31 °C) and relative humidity (~36%).
The characteristics of chlorophyll a fluorescence were measured using a fluorometer (6400-40, Li-Cor, Lincoln, NE, USA) coupled with an IRGA on the same leaf for which the gas exchange data were obtained. Initially, the leaves were adapted to the dark (when the PSII reaction centers are open) to obtain the minimum (F0) and maximum (Fm) fluorescence, with the application of the measuring light (~0.03 µmol m−2 s−1) and a saturation pulse (>3000 µmol m−2 s−1), respectively. The potential quantum yield of photosystem II (PSII) was determined as Fv/Fm = (Fm − F0)/Fm. After illumination with continuous actinic light (~1000 µmol m−2 s−1) for 40 s, a saturation pulse was applied to determine the maximum fluorescence (Fm') and steady-state (Fs) in the leaves adapted to light. The effective quantum yield of photochemical energy conversion in PSII (ΦPSII = [Fm' − Fs]/Fm') and the quantum yields of regulated nonphotochemical energy dissipation (ΦNPQ = [Fs/Fm'] − [Fs/Fm]) were calculated.
The ΦPSII was used to estimate the apparent rate of electron transport (ETR = ΦPSII × PAR × ABSleaf × 0.5; μmol electrons m−2 s−1), in which PAR is the photosynthetically active radiation (µmol photons m−2 s−1); ABSleaf corresponds to the fraction of incident light that is absorbed by the leaves, and 0.5 is the value corresponding to the fraction of excitation energy distributed to PSII.
4.3.2. Morphological Traits
The plants were measured to determine their height (H, cm), stem diameter (SD, mm), leaf number (LN), and root length (RL, cm). The shoot and root system were collected and dried individually in a forced-air oven at 65 °C until reaching a constant weight to obtain the shoot dry matter (SDM, g) and root dry matter (RDM, g). From these values, the ratio of SDM/RDM was calculated.
4.3.3. Root Uptake Efficiency and Plant Nutrient Use Efficiency
For the determination of the nutritional content of the leaves and roots, the dry plant material was ground in a Wiley mill, and ~500 mg of the dry ash was extracted with nitric-percholoric (3:1) digestion according to Silva et al. [60]. Nitrogen (N) was measured via titration by the Kjeldahl method using a nitrogen distiller (TE-0364, Tecnal, Piracicaba, SP, Brazil). Phosphorous (P) and sulfur (S) were determined via the molybdenum blue and turbidity with barium chloride methodologies, respectively, using molecular absorption spectrophotometry (SP1105, Tecnal, Piracicaba, SP, Brazil). Potassium (K) was analyzed using flame photometry (B462, Tecnal, Piracicaba, SP, Brazil), and Ca, Mg, Cu, Fe, and Zn were analyzed using atomic absorption spectrophotometry (Savant AA, GBC Scientific Equipment, Braeside, VIC, Australia). B was analyzed using the azomethine-H method [57]. The nutrient contents were expressed as g kg−1.
Based on these findings, the nutrients were determined on a dry matter basis (in g plant−1) and multiplied by the nutrient content (in g and mg for macronutrients and micronutrients, respectively). Using the results of the dry matter and nutrient contents in the plant, the root uptake efficiency (RUE) and plant nutrient use efficiency (NUE) indexes were calculated according to Swiader et al. [62] and Siddiqi and Glass [63], respectively, using the following equations:
4.4. Statistical Analysis
Physiological and morphological data were submitted to analysis of variance (ANOVA) by the F test (p ≤ 0.05) and adjusted to linear or nonlinear regression models. Standardized data were also assessed by principal component analysis (PCA). The root uptake efficiency and plant nutrient use efficiency data were analyzed by ANOVA and the means were grouped using the clustering Scott-Knott method (p ≤ 0.05). All statistical analyses were performed using the R program (v. 3.6.1, R Foundation, Vienna, Austria), and the graphs were plotted using SigmaPlot software (v. 11.0, Systat Software GmbH, Erkrath, Germany).
5. Conclusions
Nitrogen fertilization increased the photosynthetic activity and growth of E. dysenterica seedlings at rates between 50 and 200 mg dm−3. The highest evaluated N rate (400 mg dm−3) resulted in the lowest nutrient use efficiency.
Phosphate fertilization from 100 mg dm−3 increased the growth of E. dysenterica. The highest photosynthetic activity and macro- and micronutrient use efficiencies were observed at the P rate of 200 mg dm−3. Taken together, the data suggest that fertilization with 50 mg dm−3 N and 200 mg dm−3 P is suitable for the development of E. dysenterica seedlings.
Supplementary Materials
The following are available online at https://www.mdpi.com/2223-7747/9/6/722/s1, Table S1: Photosynthetic rate (A, µmol m−2 s−1), stomatal conductance (gS, mol m−2 s−1) and transpiration rate (E, mmol m−2 s−1) in Eugenia dysenterica DC seedlings grown at varying rates of nitrogen (N, mg dm−3) and phosphorus (P, mg dm−3) for 278 d. Table S2: Initial fluorescence (F0), potential quantum yield of photosystem II (Fv/Fm), electron transport rate (ETR), effective quantum yield of photosystem II (ΦPSII) and regulated quantum yield of nonphotochemical energy dissipation (ΦNPQ) in Eugenia dysenterica DC seedlings grown at varying rates of nitrogen (N, mg dm−3) and phosphorus (P, mg dm−3) for 278 d. Table S3: Height (H, cm), stem diameter (SD, mm), leaf number (LN), root length (RL, cm), total dry mass (TDM, g) and the ratio between shoot dry matter and root dry matter (SDM/RDM) in Eugenia dysenterica DC seedlings grown at varying rates of nitrogen (N, mg dm−3) and phosphorus (P, mg dm−3) for 278 d.
Author Contributions
D.N.d.R. and F.G.S. designed the research. R.d.C.S., D.N.R. and T.C.d.O. conducted the experiments. C.M. performed the physiological measurements. R.d.C.S. and D.N.d.R. performed the morphological and nutritional analyses. E.M.J. designed the figures. All authors discussed the data. D.N.d.R. and C.M. wrote the manuscript with contributions from M.B.F., F.B.d.S., and E.M.J. All authors have read and approved the manuscript to be published.
Funding
This work was supported by the Higher Education Personnel Improvement Coordination (CAPES), the Goiás Research Foundation (FAPEG.), and the Goiano Federal Institute of Education, Science and Technology, Campus Rio Verde (IFGoiano-RV).
Acknowledgments
D.N.d.R., C.M., and M.B.F. are grateful to CAPES and R.d.C.S. and F.G.S. are grateful to the Brazilian National Council for Scientific and Technological Development (CNPq) for fellowships granted.
Conflicts of Interest
All authors declare no conflicts of interest.
References
- Rezende-Silva, S.L.; Costa, A.C.; Dyszyb, F.H.; Batista, P.F.; Crispim-Filho, A.J.; Nascimento, K.J.T. Pouteria torta is a remarkable native plant for biomonitoring the glyphosate effects on Cerrado vegetation. Ecol. Indic. 2019, 102, 497–506. [Google Scholar] [CrossRef]
- Arruda, H.S.; Botrel, D.A.; Fernandes, R.V.B.; Almeida, M.E.F. Development and sensory evaluation of products containing the Brazilian Savannah fruits araticum (Annona crassiflora Mart.) and cagaita (Eugenia dysenterica Mart.). Braz. J. Food Technol. 2016, 19, e2015105. [Google Scholar] [CrossRef]
- Moreira, L.C.; Ávila, R.I.; Veloso, D.F.M.C.; Pedrosa, T.N.; Lima, E.S.; Couto, R.O.; Lima, E.M.; Batista, A.C.; Paula, J.R.; Valadares, M.C. In vitro safety and efficacy evaluations of a complex botanical mixture of Eugenia dysenterica DC. (Myrtaceae): Prospects for developing a new dermocosmetic product. Toxicol. In Vitro 2017, 1, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tunholi, V.P.; Ramos, M.A.; Scariot, A. Availability and use of woody plants in a agrarian reform settlement in the cerrado of the state of Goiás, Brazil. Acta Bot. Bras. 2013, 27, 604–612. [Google Scholar] [CrossRef]
- Rodrigues, A.A.; Vasconcelos Filho, S.C.V.; Müller, C.; Rodrigues, D.A.; Sales, J.F.; Zuchi, J.; Costa, A.C.; Rodrigues, C.L.; Silva, A.A.; Barbosa, D.P. Tolerance of Eugenia dysenterica to aluminum: Germination and plant growth. Plants 2019, 8, 317. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.S.; Guilherme, L.R.G. A career perspective on soil management in the Cerrado region of Brazil. Adv. Agron. 2016, 137, 1–72. [Google Scholar] [CrossRef]
- Souza, E.R.B.; Naves, R.V.; Oliveira, M.F. Initial fruiting of the cagaita tree (Eugenia dysenterica DC) cultivated in Goiânia, Goiás, Brazil. Rev. Bras. Frutic. 2013, 35, 906–909. [Google Scholar] [CrossRef]
- Camilo, Y.M.V.; Souza, E.R.B.; Vera, R.; Naves, R.V. Phenology, production and precocity of Eugenia dysenterica plants aiming breeding. Rev. Ciênc. Agrár. 2013, 36, 192–198. [Google Scholar]
- Huber, D.; Romheld, V.; Weinmann, M. Relationship between Nutrition, Plant Diseases and Pests. In Marschener’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier/Academic Press: Amsterdam, The Netherlands, 2012; pp. 283–298. [Google Scholar]
- Pallardy, S.G. Nitrogen Metabolism. In Physiology of Woody Plants, 3rd ed.; Pallardy, S.G., Ed.; Academic Press: Missouri, MO, USA, 2008; pp. 233–254. [Google Scholar]
- Pilbeam, D.J. Nitrogen. In Plant Nutrition Handbook, 2nd ed.; Pilbeam, D.J., Barker, A.V., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2015; pp. 20–64. [Google Scholar] [CrossRef]
- Hopkins, B.G. Phosphorus. In Plant Nutrition Handbook, 2nd ed.; Pilbeam, D.J., Barker, A.V., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2015; pp. 65–126. [Google Scholar] [CrossRef]
- Singh, S.K.; Reddy, V.R.; Fleisher, D.H.; Timlin, D.J. Relationship between photosynthetic pigments and chlorophyll fluorescence in soybean under varying phosphorus nutrition at ambient and elevated CO2. Photosynthetica 2017, 55, 421–433. [Google Scholar] [CrossRef]
- Cheng, L.; Fuchigami, L.H. Rubisco activation state decreases with increasing nitrogen content in apple leaves. J. Exp. Bot. 2000, 51, 1687–1694. [Google Scholar] [CrossRef] [PubMed]
- Warren, C.R.; Adams, M.A. Phosphorus affects growth and partioning of nitrogen to Rubisco in Pinus pinaster. Tree Physiol. 2002, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Freitas, J.C.O.; Almeida, A.A.F.; Lago, M.F.; Souza, M.M.; Junior, J.O.S. Morphophysiological characteristics of clonal plants Passiflora alata grown in different doses of nitrogen and shading levels. Rev. Bras. Frutic. 2012, 34, 859–872. [Google Scholar] [CrossRef]
- Barreto, P.A.B.; Gama-Rodrigues, A.C.; Gama-Rodrigues, E.F.; Barros, N.F. Nitrogen balance in soil under eucalyptus plantations. Rev. Bras. Cien Solo 2012, 36, 1239–1248. [Google Scholar] [CrossRef]
- Carranca, C.; Brunetto, G.; Tagliavini, M. Nitrogen nutrition of fruit trees to reconcile productivity and environmental concerns. Plants 2018, 7, 4. [Google Scholar] [CrossRef]
- Jacob, J.; Lawlor, D.W. Stomatal and mesophyll limitations of photosynthesis in phosphate deficient sunflower, maize and wheat plants. J. Exp. Bot. 1991, 42, 1003–1011. [Google Scholar] [CrossRef]
- Seika, N.; Yano, K. Stomatal density of cowpea correlates with carbon isotope discrimination in different phosphorus, water and CO2 environments. New Phytol. 2008, 179, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Salter, W.T.; Turnbull, T.L.; Okazaki, Y.; Saito, K.; Kreuzwieser, J.; Rennenberg, H.; Adams, M.A. Plant and soil P determine functional attributes of subalpine Australian plants. Arct Antarct. Alp. Res. 2018, 50, 1. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Role of the xanthophyll cycle in photoprotection elucidated by measurements of light-induced absorbance changes, fluorescence and photosynthesis in leaves of Hedera canariensis. Photosynth. Res. 1990, 5, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Carstensen, A.; Herdean, A.; Schimidt, S.B.; Sharma, A.; Spetea, C.; Pribil, M.; Husted, S. The impacts of phosphorus deficiency on the photosynthetic electron transport chain. Plant. Physiol. 2018, 177, 271–284. [Google Scholar] [CrossRef]
- Duarte, T.F.; Bron, I.U.; Ribeiro, R.V.; Machado, E.C.; Mazzafera, P.; Shimizu, M.M. Effect of crop loading on quality of ‘Valencia’ orange fruit. Rev. Bras. Frutic. 2011, 33, 823–829. [Google Scholar] [CrossRef]
- Tosta, M.S.; Almeida, J.P.N.; Góes, G.B.; Freire, P.A.; Mendonça, V. Nitrogen fertilization in the production of seedlings of Talisia suculenta (A. ST. Hil) Radlk. R. Bras. Eng. Agríc. Ambiental. 2017, 21, 443–447. [Google Scholar] [CrossRef]
- Millard, P.; Grelet, G. Nitrogen storage and remobilization by trees: Ecophysiological relevance in a changing world. Tree Physiol. 2010, 30, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- van Oosten, M.J.; Dell’Aversana, E.; Ruggiero, A.; Cirillo, V.; Gibon, Y.; Woodrow, P.; Maggio, A.; Carillo, P. Omeprazole treatment enhances nitrogen use efficiency through increased nitrogen uptake and assimilation in corn. Front. Plant. Sci. 2019, 10, 1507. [Google Scholar] [CrossRef] [PubMed]
- van de Wiel, C.C.M.; van der Linden, C.G.; Scholten, O.E. Improving phosphorus use efficiency in agriculture: Opportunities for breeding. Euphytica 2016, 207, 1–22. [Google Scholar] [CrossRef]
- Tng, D.Y.; Janos, D.P.; Jordan, G.J.; Weber, E.; Bowman, D.M. Phosphorus limits Eucalyptus grandis seedling growth in an unburnt rain forest soil. Front. Plant. Sci. 2014, 5, 527. [Google Scholar] [CrossRef] [PubMed]
- Razaq, M.; Zhang, P.; Shen, H.; Salahuddin. Influence of nitrogen and phosphorus on the growth and root morphology of Acer mono. PLoS ONE 2017, 12, e0171321. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Trees increase their P : N ratio with size. Glob. Ecol. Biogeogr. 2015, 24, 147–156. [Google Scholar] [CrossRef]
- Freibeiger, M.B.; Guerrini, I.A.; Galetti, G.; Fernandes, D.M.; Corrêa, J.C. Early growth and nutrition of cedar (Cedrela fissilis Vell.) as affected by nitrogen rates. Rev. Árvore. 2013, 37, 385–392. [Google Scholar] [CrossRef]
- Ulas, A.; Doganci, E.; Ulas, F.; Yetisir, H. Root-growth characteristics contributing to genotypic variation in nitrogen efficiency of bottle gourd and rootstock potential for watermelon. Plants 2019, 8, 77. [Google Scholar] [CrossRef]
- Melo, J.T.; Haridasan, M. Response of cagaita (Eugenia dysenterica DC) seedlings to N, P, K, Ca and Mg doses, 1st ed.; EMBRAPA—Boletim de Pesquisa e Desenvolvimento: Planaltina, DF, Brazil, 2009; p. 27. (In Portuguese) [Google Scholar]
- Costa, A.M.; Carlos, L.; Silva, P.O.; Barbosa, K.P.; Rodrigues, C.R. Nitrogen and potassium fertilization in the initial growth of Annona crassiflora Mart. Florest. Ambiente. 2019, 26, 1–9. [Google Scholar] [CrossRef]
- Constantino, V.; Barbosa, J.Z.; Motta, A.V.; Dolinski, M.A.; Prior, S.A.; Zanette, F. Initial growth of Araucaria angustifolia (Bertol.) Kuntze in response to fertilization with nitrogen, phosphorus and potassium. Floresta 2019, 49, 99–108. [Google Scholar] [CrossRef]
- Bessa, L.A.; Moreira, M.A.; Silva, F.G.; Mota, C.S.; Vitorino, L. Growth, nutrient concentration and principal component analysis of Cagaita (Eugenia dysenterica DC.) seedlings grown in nutrient solution. Aust. J. Crop Sci. 2016, 10, 425–433. [Google Scholar] [CrossRef]
- Shen, H.; Chen, J.; Wang, Z.; Yang, C.; Sasaki, T.; Yamamoto, Y.; Matsumoto, H.; Yan, X. Root plasma membrane H+-ATPase is involved in the adaptation of soybean to phosphorus starvation. J. Exp. Bot. 2006, 57, 1353–1362. [Google Scholar] [CrossRef] [PubMed]
- Silveira, C.E.S.; Palhares, D.; Pereira, L.A.R.; Pereira, K.B.D.; Silva, F.A.B. Strategies of plant establishment of two Cerrado species: Byrsonima basiloba Juss. (Malpighiaceae) and Eugenia dysenterica Mart. Ex DC. (Myrtaceae). Plant. Spec. Biol. 2013, 28, 130–137. [Google Scholar] [CrossRef]
- Carnevali, N.H.S.; Marchetti, M.E.; Vieira, M.C.; Carnevali, T.O.; Ramos, D.D. Nutritional efficiency of Stryphnodendron polyphyllum seedlings in function of nitrogen and phosphorus. Ciên. Florest. 2016, 26, 449–461. [Google Scholar] [CrossRef]
- Pinto, J.V.; Vieira, M.C.; Zárate, N.A.H.; Formagio, A.S.N.; Cardoso, C.A.L.; Carnevali, T.O.; Souza, P.H.N. Effect of soil nitrogen and phosphorus on early development and essential oil composition of Schinus terebinthifolius Raddi. J. Essent. Oil Bear. Pl. 2016, 19, 247–257. [Google Scholar] [CrossRef]
- Fageria, N.K. Nitrogen interaction with other nutrients. In Nitrogen Management in Crop Production; Fageria, N.K., Ed.; CRC Press, Taylor and Francis Group: Boca Raton, FL, USA, 2014; pp. 245–274. [Google Scholar]
- Pimentel, L.D.; Bruckner, C.H.; Martinez, H.E.P.; Motoike, S.Y.; Manfio, C.E.; Santos, R.C. Effect of nitrogen and potassium rates on early development of macaw palm. Rev. Bras. Ciênc. Solo 2015, 39, 1671–1680. [Google Scholar] [CrossRef]
- Pavinato, P.S.; Merlin, A.; Rosolem, C.A. Phosphorus fractions in Brazilian Cerrado soils as affected by tillage. Soil Till. Res. 2009, 105, 149–155. [Google Scholar] [CrossRef]
- White, P.J. Long-distance transport in the xylem and phloem. In Marschener’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier/Academic Press: Amsterdam, The Netherlands, 2012; pp. 42–70. [Google Scholar]
- Grzebisz, W. Potassium fertilization of arable crops—The crop rotation oriented concept. Fertiliz 2005, 3, 328–341. [Google Scholar] [CrossRef]
- Marschner, P.; Rengel, Z. Nutrient availability in soils. In Marschener’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier/Academic Press: Amsterdam, The Netherlands, 2012; pp. 315–330. [Google Scholar]
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition, 5th ed.; Kluwer Academic Publisher: Dodrecht, The Netherlands, 2001; pp. 210–212. [Google Scholar]
- Andrés, Z.; Pérez-Hormaeche, J.; Leidi, E.O.; Schulucking, K.; Steinhorst, L.; Mclachlan, D.H.; Pardo, J.M. Control of vacuolar dynamics and regulation of stomatal aperture by tonoplast potassium uptake. Proc. Natl. Acad. Sci. USA 2014, 111, 17. [Google Scholar] [CrossRef] [PubMed]
- Alva, A.K.; Mattos-Junior, D.; Paramasivam, S.; Patil, B.; Dou, H.; Sajwan, K. Potassium management for optimizing citrus production and quality. Int. J. Fruit Sci. 2006, 6, 3–43. [Google Scholar] [CrossRef]
- Quaggio, J.A.; Mattos-Junior, D.; Boaretto, R.M. Sources and rates of potassium for sweet orange production. Sci. Agricola 2011, 68, 369–375. [Google Scholar] [CrossRef]
- Gonçalves, W.V.; Vieira, M.C.D.; Carnevali, T.O.; Zárate, N.A.H.; Aran, H.D.V.R.; Mineli, K.C.S. Nitrogen and phosphorus fertilization promotes aerial part development and affect nutrient uptake by carobinha of the Brazilian Cerrado. Am. J. Plant Sci. 2017, 8, 3377–3398. [Google Scholar] [CrossRef][Green Version]
- Bessa, L.A.; Silva, F.G.; Moreira, M.A.; Teodoro, J.P.R.; Soares, F.A.L. Characterization of nutrient deficiency in Hancornia speciosa Gomes seedlings by omitting micronutrients from the nutrient solution. Rev. Bras. Frutic. 2013, 35, 616–624. [Google Scholar] [CrossRef]
- Teixeira, I.R.; Borém, A.; Araújo, G.A.A.; Fontes, R.L.F. Manganese and zinc leaf application on common bean grown on a “Cerrado”. Soil Sci. Agric. 2004, 61, 77–81. [Google Scholar] [CrossRef]
- Carlisle, E.; Myers, S.; Raboy, V.; Bloom, A. The effects of inorganic nitrogen form and CO2 concentration on wheat yield and nutrient accumulation and distribution. Front. Plant Sci. 2012, 3, 1–13. [Google Scholar] [CrossRef]
- Kobayashi, M.; Nakagawa, H.; Asaka, T.; Matoh, T. Boraterhamnogalacturonan II bonding reinforced by Ca2+ retains pectic polysaccharides in higher-plant cell walls. Plant Physiol. 1999, 119, 199–204. [Google Scholar] [CrossRef]
- Hu, H.; Brown, P.H. Localization of boron in cell walls of squash and tobacco and its association with pectin. Plant Physiol. 1994, 105, 681–689. [Google Scholar] [CrossRef]
- Brun, F.G.K.; Brin, E.J.; Gerber, D.; Szymczak, D.A.; Londero, A.K.; Meyer, E.; Navrosky, M.C. Nutrition facts and limits for micronutrients in tree species used in urban forestry. An. Acad. Bras. Ciênc. 2017, 89, 1881–1893. [Google Scholar] [CrossRef]
- Silva, F.C.; Abreu, M.F.; Perez, D.V.; Eira, P.A.; Abreu, C.A.; Raij, B.; Gianello, C.; Coelho, A.M.; Quaggio, J.A.; Tedesco, M.J.; et al. Chemical Analysis Methods for Soil Fertility Evaluation, 2nd ed.; Embrapa Solos: Rio de Janeiro, Brazil, 2009; p. 627. [Google Scholar]
- Malavolta, E.; Vitti, G.C.; Oliveira, A.S. Evaluation of Plant Nutritional Status: Principles and Applications, 2nd ed.; Potafos: Piracicaba, Brazil, 1997; pp. 215–241. [Google Scholar]
- Swiader, J.M.; Chyan, Y.; Freiji, F.G. Genotypic differences in nitrate uptake and utilization efficiency in pumpkin hybrids. J. Plant Nutr. 1994, 17, 1687–1699. [Google Scholar] [CrossRef]
- Siddiqi, M.Y.; Glass, A.D.M. Utilization index: A modified approach to the estimation and comparison of nutrient utilization efficiency in plants. J. Plant Nutr. 1981, 4, 289–302. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).