Comparative Germination Ecology of Two Endemic Rhaponticum Species (Asteraceae) in Different Climatic Zones of the Ligurian and Maritime Alps (Piedmont, Italy)


Abstract
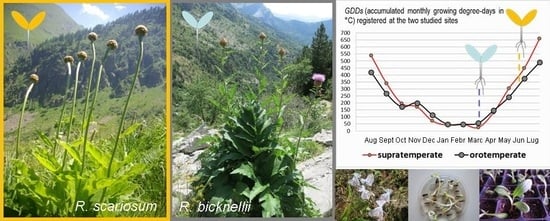
1. Introduction
2. Results
2.1. Study 1—Microclimatic Conditions at the Growing Sites and in the Nursery
2.2. Study 2—Conversion of Temperature Data into Different Climatic Parameters
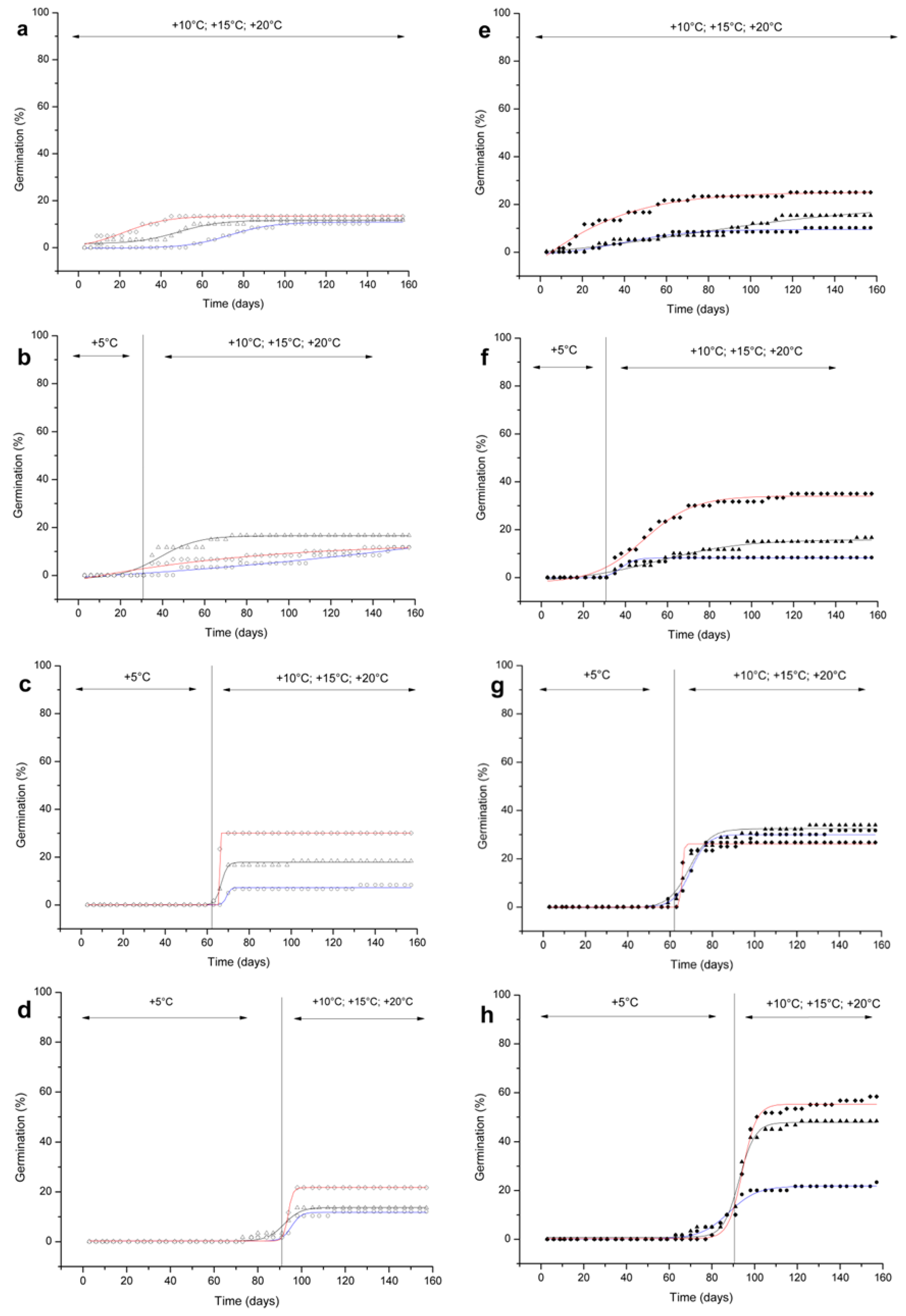
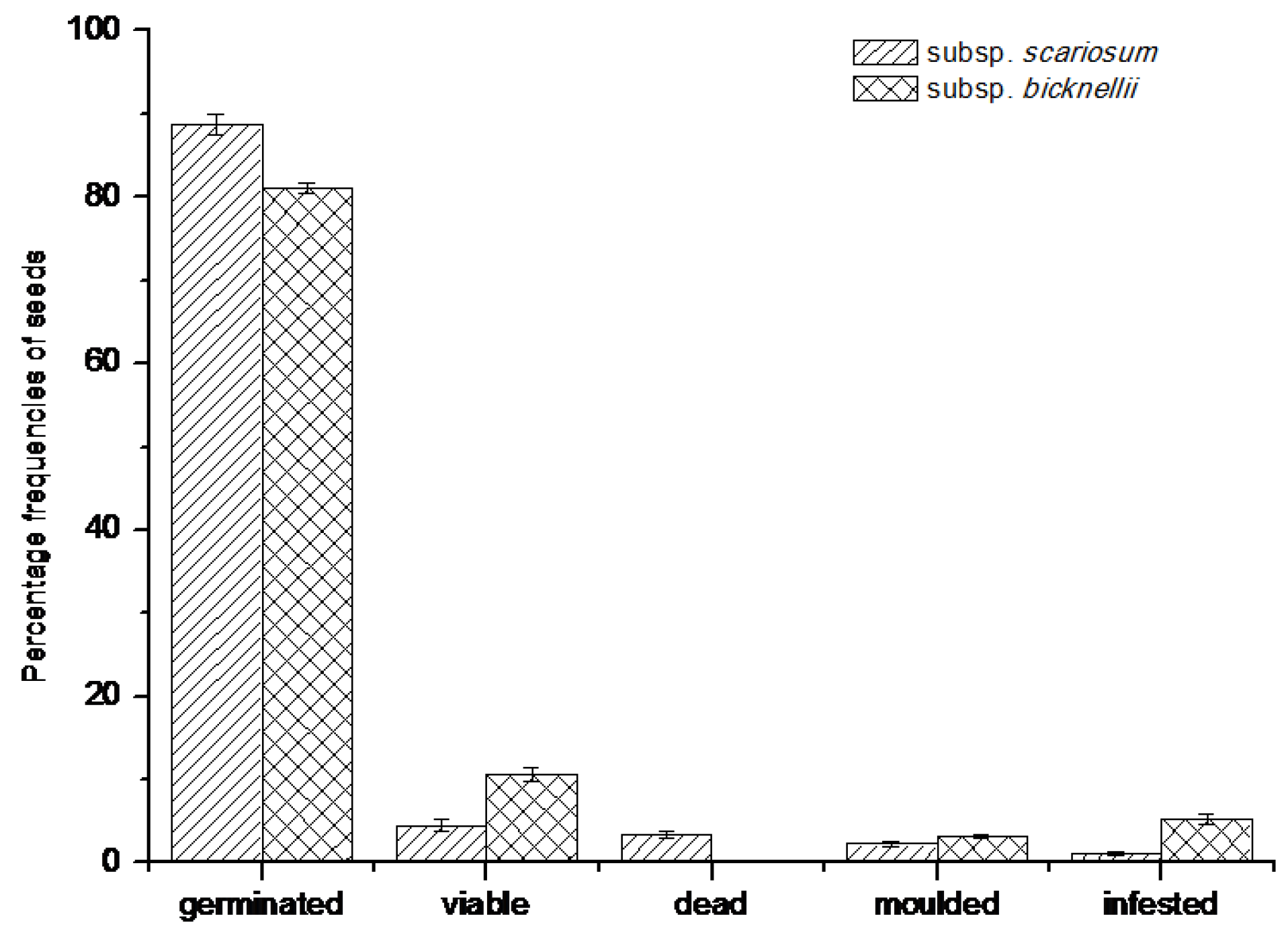
2.3. Study 3—Germination Experiments under Controlled Conditions in the Lab
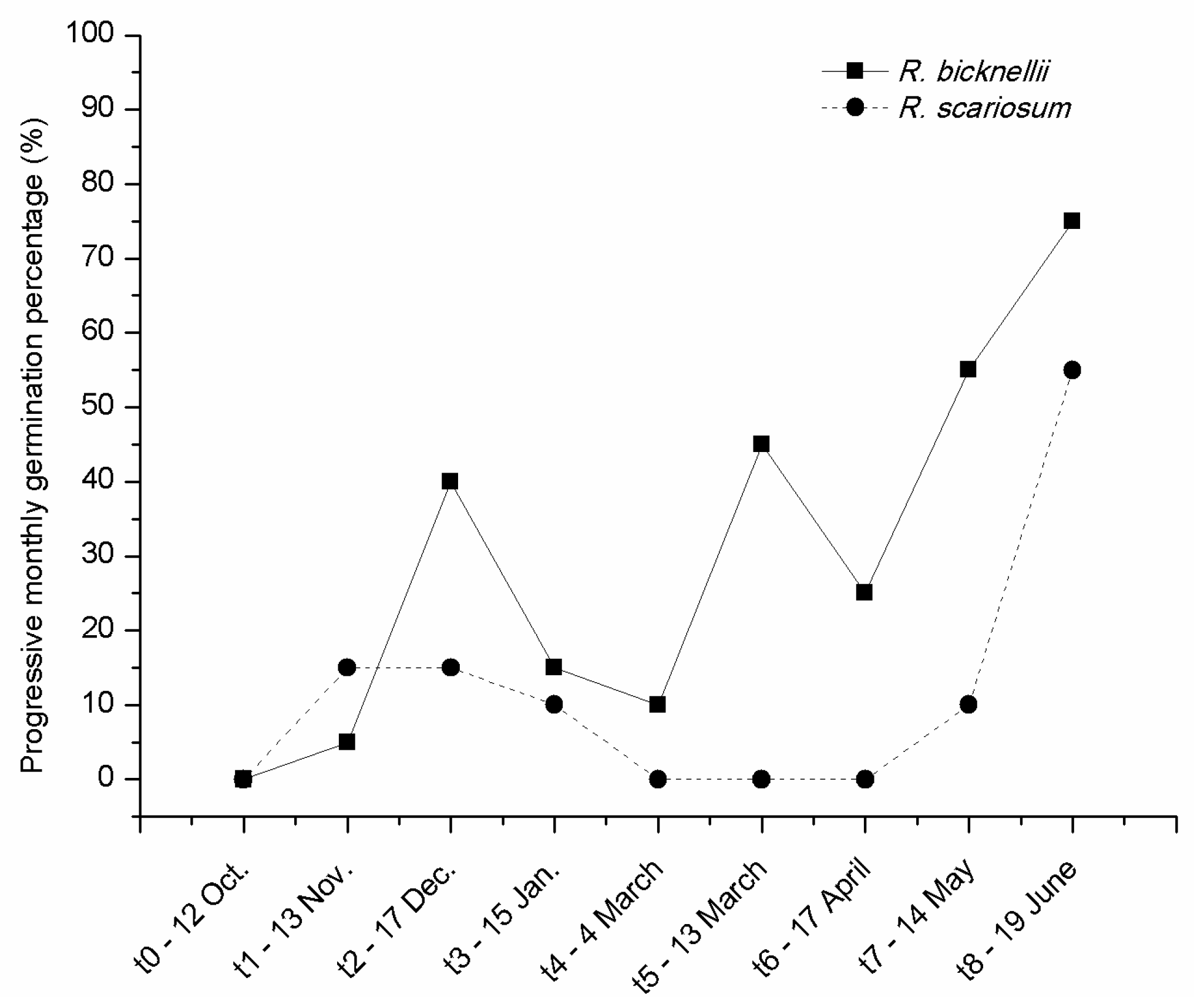
2.4. Study 4—Seed Burial Experiments in the Field and in the Plant Nursery
3. Discussion
3.1. Germination Experiments under Controlled Conditions in the Lab
3.1.1. Seed Responses to Cold Stratification
3.1.2. Physiological Dormancy
3.1.3. R. scariosum and R. bicknellii do not Share Common Germination Strategies
3.2. Seed Burial Experiments in the Field and in the Plant Nursery
3.2.1. R. scariosum and R. bicknellii Form Transient Soil Seed Banks
3.2.2. Seed in the Soil in Relation to Different Microclimatic Conditions
3.3. Microclimatic Conditions at the Growing Sites and in the Nursery
4. Materials and Methods
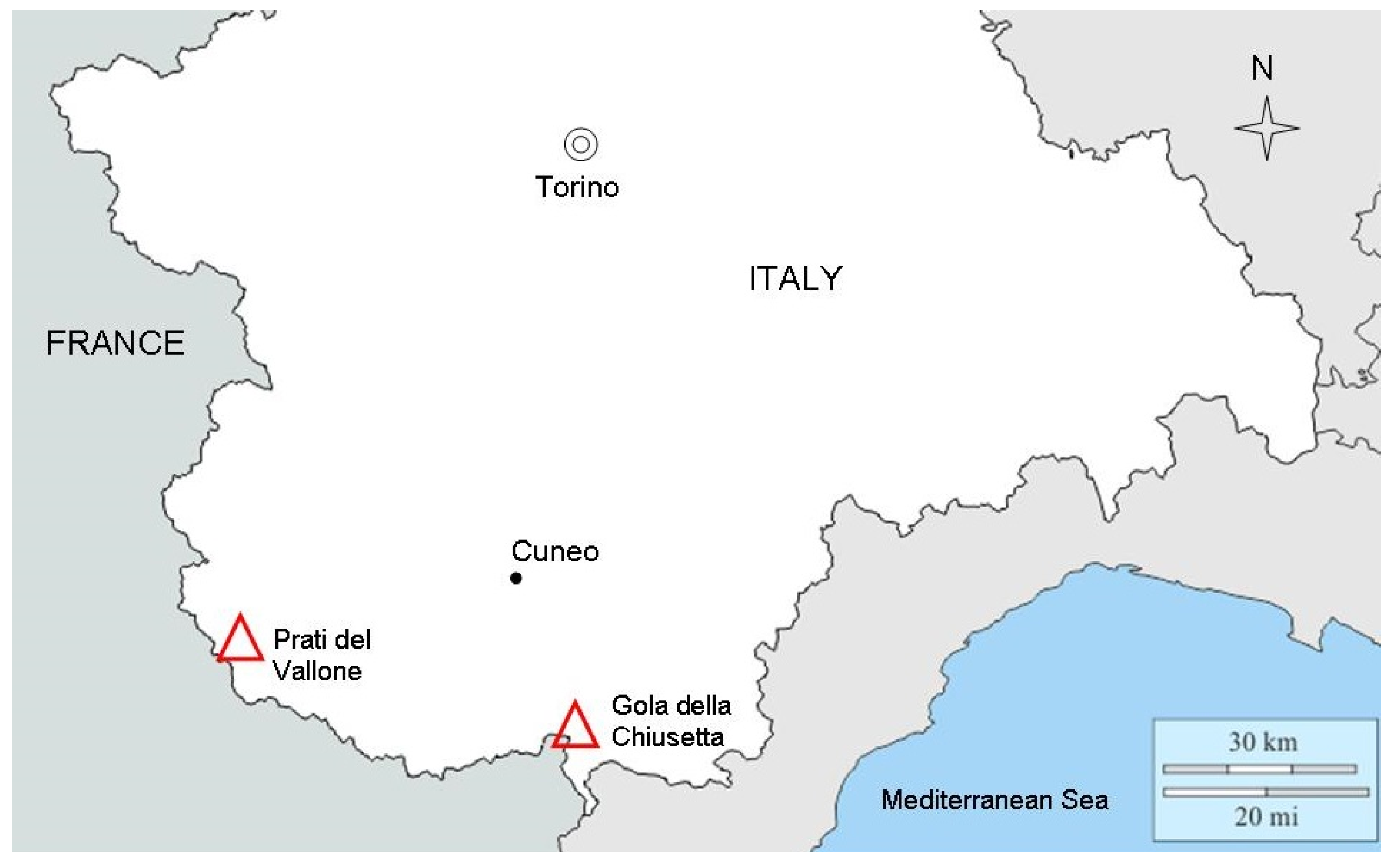
4.1. Species Taxonomy, Distribution and Study Sites
4.2. Study 1—Microclimatic Conditions at the Growing Sites and in the Plant Nursery
4.3. Study 2—Conversion of Temperature Data into Different Climatic Parameters
4.4. Study 3—Germination Experiments under Controlled Conditions in the Lab
4.4.1. Seed Collection and Preparation
4.4.2. Germination Tests
4.5. Study 4—Seed Burial Experiments at the Study Sites and in the Plant Nursery
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Donohue, K. Seeds and seasons: Interpreting germination timing in the field. Seed Sci. Res. 2005, 15, 175–187. [Google Scholar] [CrossRef]
- Penfield, S.; King, J. Towards a systems biology approach to understanding seed dormancy and germination. Proc. R. Soc. B Biol. Sci. 2009, 276, 3561–3569. [Google Scholar] [CrossRef] [PubMed]
- Forbis, T.A. Seedling demography in an alpine ecosystem. Am. J. Bot. 2003, 90, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.E.N.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Bernareggi, G.; Carbognani, M.; Mondoni, A.; Petraglia, A. Seed dormancy and germination changes of snowbed species under climate warming: The role of pre- and post-dispersal temperatures. Ann. Bot. 2016, 118, 529–539. [Google Scholar] [CrossRef]
- Hoyle, G.L.; Cordiner, H.; Good, R.B.; Nicotra, A.B. Effects of reduced winter duration on seed dormancy and germination in six populations of the alpine herb Aciphyllya glacialis (Apiaceae). Conserv. Physiol. 2014, 2, 1–11. [Google Scholar] [CrossRef]
- Cochrane, A.; Yates, C.J.; Hoyle, G.L.; Nicotra, A.B. Will among-population variation in seed traits improve the chance of species persistence under climate change? Glob. Ecol. Biogeogr. 2014, 24, 12–24. [Google Scholar] [CrossRef]
- Donohue, K.; de Casas, R.R.; Burghardt, L.; Kovach, K.; Willis, C.G. Germination, postgermination adaptation, and species ecological ranges. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds, Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Baskin, C.C.; Baskin, J.M.; Auken, O.W. Role of temperature in dormancy break and/or germination of autumn-maturing achenes of eight perennial Asteraceae from Texas, U.S.A. Plant Species Biol. 1998, 13, 13–20. [Google Scholar] [CrossRef]
- Nikolaeva, M.G. Physiology of Deep Dormancy in Seeds; Izdatel’stvo ‘Nauka’: Leningrad, Russia, 1969. [Google Scholar]
- Nikolaeva, M.G. Factors controlling the seed dormancy pattern. In The Physiology and Biochemistry of Seed Dormancy and Germination; Khan, A.A., Ed.; North-Holland Publ.: Amsterdam, The Netherlands, 1977; pp. 51–74. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sc. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Schwienbacher, E.; Navarro-Cano, J.A.; Neuner, G.; Erschbamer, B. Seed dormancy in alpine species. Flora Morphol. Distrib. Funct. Ecol. Plants 2011, 206, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Mondoni, A.; Rossi, G.; Orsenigo, S.; Probert, R.J. Climate warming could shift the time of seed germination alpine plants. Ann. Bot. 2012, 110, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Cavieres, L.A.; Arroyo, M.T.K. Seed germination protocol to cold stratification period and thermal regime in Phacelia secunda (Hydrophyllaceae). Plant Ecol. 2000, 149, 1–8. [Google Scholar] [CrossRef]
- Mondoni, A.; Pedrini, S.; Bernareggi, G.; Rossi, G.; Abeli, T.; Probert, R.J.; Ghitti, M.; Bonomi, C.; Orsenigo, S. Climate warming could increase recruitment success in glacier foreland plants. Ann. Bot. 2015, 116, 907–916. [Google Scholar] [CrossRef] [PubMed]
- Giménez-Benavides, L.; Escudero, A.; Pérez-García, F. Seed germination of high mountain Mediterranean species: Altitudinal, interpopulation and interannual variability. Ecol. Res. 2005, 20, 433–444. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Sierra-Almeida, A. Assessing the importance of cold-stratification for seed germination in alpine plant species of the High-Andes of central Chile. Perspect. Plant Ecol. Evol. Syst. 2018, 30, 125–131. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar] [CrossRef]
- Fernández-Pascual, E.; Seal, C.E.; Pritchard, H.W. Simulating the germination response to diurnally alternating temperatures under climate change scenarios: Comparative studies on Carex diandra seeds. Ann. Bot. 2015, 115, 201–209. [Google Scholar] [CrossRef]
- Kozlowski, G.; Bürcher, S.; Fleury, M.; Huber, F. The Atlantic elements in the Swiss flora: Distribution, diversity, and conservation status. Biodiv. Cons. 2009, 18, 649–662. [Google Scholar] [CrossRef][Green Version]
- Diadema, K.; Noble, V. La flore des Alpes Maritimes et de la Principauté de Monaco. Originalité et Diversité; Naturalia Publications: Turriers, France, 2011. [Google Scholar]
- Gallino, B.; Pallavicini, G. La Vegetazione Delle Alpi Liguri e Marittime; Blu Edizioni: Peveragno (Cuneo), Italy, 2000. [Google Scholar]
- Rivas-Martínez, S.; Penas, A.; Díaz, T.E. Bioclimatic map of Europe–Thermotypes. Cartographic Service 2004; University of Leon. Available online: http//www.globalbioclimatics.org/form/tb_med.htm (accessed on 23 March 2020).
- Rivas-Martínez, S.; Sáenz, S.R.; Penas, A. Worldwide bioclimatic classification system. Glob. Geobot. 2011, 1, 1–634. [Google Scholar]
- Banfi, E.; Galasso, G.; Soldano, A. Notes on systematics and taxonomy for the Italian vascular flora. 2. Nat. Hist. Sci. 2011, 152, 85. [Google Scholar] [CrossRef][Green Version]
- Pignatti, S. Flora d’Italia; Edagricole: Bologna, Italia, 2017. [Google Scholar]
- Tison, J.M.; de Foucault, B. Flora Gallica: Flore de France; Biotope Èditions: Mèze, France, 2014. [Google Scholar]
- GCC: Global Compositae Checklist in the Catalogue of Life. Available online: https://www.gbif.org/dataset/d849eefa-ef3b-4f2c-a023-500fa5521a91 (accessed on 18 March 2020).
- Rivas-Martínez, S. Global bioclimatics (Clasificaciòn Bioclimática de la Tierra). Phytosociological Research Center. 2004. Available online: http//www.globalbioclimatics.org/book/bioc/global_bioclimatics_0.htm (accessed on 18 March 2020).
- Worldwide Bioclimatic Classification System. Available online: http://www.globalbioclimatics.org/form/bi_med.htm (accessed on 18 March 2020).
- McMaster, G.S.; Wilhelm, W.W. Growing degree-days: One equation, two interpretations. Agric. For. Meteorol. 1997, 87, 291–300. [Google Scholar] [CrossRef]
- Amen, R.D. The Extent and Role of Seed Dormancy in Alpine Plants. Q. Rev. Biol. 1966, 41, 271–281. [Google Scholar] [CrossRef]
- Sommerville, K.D.; Martyn, A.J.; Offord, C.A. Can seed characteristics or species distribution be used to predict the stratification requirements of herbs in the Australian Alps? Bot. J. Linn. Soc. 2013, 172, 187–204. [Google Scholar] [CrossRef]
- Hoyle, G.L.; Steadman, K.J.; Good, R.B.; McIntosh, E.J.; Galea, L.M.E.; Nicotra, A.B. Seed germination strategies: An evolutionary trajectory independent of vegetative functional traits. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef]
- Schütz, W.; Milberg, P.; Schutz, W. Seed Dormancy in Carex canescens: Regional differences and ecological consequences. Oikos 1997, 78, 420. [Google Scholar] [CrossRef]
- Shimono, Y.; Kudo, G. Comparisons of germination traits of alpine plants between fellfield and snowbed habitats. Ecol. Res. 2005, 20, 189–197. [Google Scholar] [CrossRef]
- García-Fernández, A.; Escudero, A.; Lara-Romero, C.; Iriondo, J.M. Effects of the duration of cold stratification on early life stages of the Mediterranean alpine plant Silene ciliata. Plant Biol. 2015, 17, 344–350. [Google Scholar] [CrossRef]
- Mondoni, A.; Orsenigo, S.; Müller, J.V.; Carlsson-Graner, U.; Jiménez-Alfaro, B.; Abeli, T. Seed dormancy and longevity in subarctic and alpine populations of Silene suecica. Alp. Bot. 2018, 128, 71–81. [Google Scholar] [CrossRef]
- Fernández-Pascual, E.; Jiménez-Alfaro, B.; Bueno, Á. Comparative seed germination traits in alpine and subalpine grasslands: Higher elevations are associated with warmer germination temperatures. Plant Biol. 2016, 19, 32–40. [Google Scholar] [CrossRef]
- Schwienbacher, E.; Marcante, S.; Erschbamer, B. Alpine species seed longevity in the soil in relation to seed size and shape. A 5-year burial experiment in the Central Alps. Flora Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 19–25. [Google Scholar] [CrossRef]
- Schütz, W. Dormancy characteristics and germination timing in two alpine Carex species. Basic Appl. Ecol. 2002, 3, 125–134. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar] [CrossRef]
- Thompson, K.; Bakker, J.P.; Bekker, R.M.; Hodgson, J.G. Ecological correlates of seed persistence in soil in the north-west European flora. J. Ecol. 1998, 86, 163–169. [Google Scholar] [CrossRef]
- Venn, S.E.; Morgan, J.W. Germination characteristics of mountain celery Aciphylla glacialis (F. Muell.) Benth. (Apiaceae). Vic. Nat. 2009, 126, 4–12. [Google Scholar]
- Thompson, K. The functional ecology of seed banks. In Seeds: The Ecology of Regeneration in Plant Communities; Fenner, M., Ed.; CAB International: Wallingford, UK, 1992; pp. 231–258. [Google Scholar]
- Thompson, K.; Bakker, J.P.; Bakker, R.M. The Soil Seed Banks of North West Europe: Methodology, Density and Longevity; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Thompson, K.; Band, S.R.; Hodgson, J.G. Seed size and shape predict persistence in soil. Funct. Ecol. 1993, 7, 236–241. [Google Scholar] [CrossRef]
- Arroyo, M.T.K.; Cavieres, L.A.; Castor, C.; Humaña, A.M. Soil seed pool; persistent seed bank and standing vegetation in a high alpine site in the central Chilean Andes. Oecologia 1999, 119, 126–132. [Google Scholar] [CrossRef]
- Funes, G.; Basconcelo, S.; Díaz, S.; Cabido, M. Seed bank dynamics in tall-tussock grasslands along an altitudinal gradient. J. Veg. Sci. 2003, 14, 253–258. [Google Scholar] [CrossRef]
- Chambers, J.C.; MacMahon, J.A. A Day in the Life of a Seed: Movements and fates of seeds and their implications for natural and managed systems. Annu. Rev. Ecol. Syst. 1994, 25, 263–292. [Google Scholar] [CrossRef]
- Nogués-Bravo, D.; Araújo, M.B.; Errea, M.P.; Martínez-Rica, J.P. Exposure of global mountain systems to climate warming during the 21st Century. Glob. Environ. Chang. 2007, 17, 420–428. [Google Scholar] [CrossRef]
- Venn, S.E.; Morgan, J.W. Patterns in alpine seedling emergence and establishment across a stress gradient of mountain summits in south-eastern Australia. Plant Ecol. Divers. 2009, 2, 5–16. [Google Scholar] [CrossRef]
- Scherrer, D.; Körner, C. Infra-red thermometry of alpine landscapes challenges climatic warming projections. Glob. Chang. Biol. 2009, 16, 2602–2613. [Google Scholar] [CrossRef]
- Greuter, W.; Aghababian, M.; Wagenitz, G. Vaillant on Compositae -systematic concepts and nomenclatural impact. Taxon 2005, 54, 149–174. [Google Scholar] [CrossRef]
- Hidalgo, O.; Garcia-Jacas, N.; Garnatje, T.; Susanna, A. Phylogeny of Rhaponticum (Asteraceae; Cardueae-Centaureinae) and related genera inferred from nuclear and chloroplast DNA sequence data: Taxonomic and biogeographic implications. Ann. Bot. 2006, 97, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Khela, S. The IUCN Red List of Threatened Species. Rhaponticum scariosum. 2013. Available online: https://www.iucnredlist.org/species/202988/44866836 (accessed on 23 March 2020).
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.P.; Raus, T.; Čarni, A.; Ŝumberova, K.; Willner, W.; Dengler, J.; Gavilán García, R.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sc. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Pesaresi, S.; Galdenzi, D.; Biondi, E.; Casavecchia, S. Bioclimate of Italy: Application of the worldwide bioclimatic classification system. J. Maps 2014, 10, 538–553. [Google Scholar] [CrossRef]
- Stokew, P. Temperature and seed dormancy. In Encyclopedia of Plant Physiology; Ruhland, W., Ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1965; Volume 15/2, pp. 746–803. [Google Scholar]
- Crawley, M.J. The R Book; John Wiley & Sons: Chichester, UK, 2013. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Estimate | SE | z-Value | p |
|---|---|---|---|---|
| Intercept | −3.228926 | 0.325025 | −9.934 | <2 × 10−16 *** |
| Cold stratification (Str) | 0.004309 | 0.003156 | 1.365 | 0.17218 |
| Incubation temperature (T) | 0.083810 | 0.016694 | 5.020 | 5.16 × 10−7 *** |
| R. bicknellii | 0.201804 | 0.247832 | 0.814 | 0.41549 |
| Str x R. bicknellii | 0.011759 | 0.004139 | 2.841 | 0.00449 ** |
| Factor | Estimate | SE | z-Value | p |
|---|---|---|---|---|
| Intercept | −2.9257 | 0.4524 | −6.467 | 1.00 × 10−10 *** |
| Cold stratification (Str) | 0.0043 | 0.0032 | 1.350 | 0.177 |
| Incubation temperature (T) | 0.0649 | 0.0262 | 2.484 | 0.013 * |
| Factor | Estimate | SE | t-Value | p |
|---|---|---|---|---|
| Intercept | −3.2281 | 0.3883 | −8.314 | 4.65 × 10−16 *** |
| Cold stratification (Str) | 0.0162 | 0.0027 | 6.004 | 3.06 × 10−9 *** |
| Incubation temperature (T) | 0.0963 | 0.0212 | 4.432 | 1.08 × 10−5 *** |
| Species | Study Areas (Acronyms) | Habitat Code | SIC Code b | Geology | Altitude (m a.s.l.) | Aspect | Thermoclimatic Belts c |
|---|---|---|---|---|---|---|---|
| R. scariosum | PDV a | Alpine pastures: 6150 | IT 1160021 | Siliceous rocks | 1915 | South-East | Orotemperate |
| R. bicknellii | GDC a | Alpine pastures: 6170 * | IT 1160057 | Calcareous rocks | 1840 | South | Supratemperate |
| Treatments in the Dark | Cold Stratification Period (in Days) | Incubation Temperatures (°C) | |||||
|---|---|---|---|---|---|---|---|
| 0 | 30 | 60 | 90 | 10 | 15 | 20 | |
| 1 | X | X | |||||
| 2 | X | X | |||||
| 3 | X | X | |||||
| 4 | X | X | |||||
| 5 | X | X | |||||
| 6 | X | X | |||||
| 7 | X | X | |||||
| 8 | X | X | |||||
| 9 | X | X | |||||
| 10 | X | X | |||||
| 11 | X | X | |||||
| 12 | X | X | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carasso, V.; Mucciarelli, M.; Dovana, F.; Müller, J.V. Comparative Germination Ecology of Two Endemic Rhaponticum Species (Asteraceae) in Different Climatic Zones of the Ligurian and Maritime Alps (Piedmont, Italy). Plants 2020, 9, 708. https://doi.org/10.3390/plants9060708
Carasso V, Mucciarelli M, Dovana F, Müller JV. Comparative Germination Ecology of Two Endemic Rhaponticum Species (Asteraceae) in Different Climatic Zones of the Ligurian and Maritime Alps (Piedmont, Italy). Plants. 2020; 9(6):708. https://doi.org/10.3390/plants9060708
Chicago/Turabian StyleCarasso, Valentina, Marco Mucciarelli, Francesco Dovana, and Jonas V Müller. 2020. "Comparative Germination Ecology of Two Endemic Rhaponticum Species (Asteraceae) in Different Climatic Zones of the Ligurian and Maritime Alps (Piedmont, Italy)" Plants 9, no. 6: 708. https://doi.org/10.3390/plants9060708
APA StyleCarasso, V., Mucciarelli, M., Dovana, F., & Müller, J. V. (2020). Comparative Germination Ecology of Two Endemic Rhaponticum Species (Asteraceae) in Different Climatic Zones of the Ligurian and Maritime Alps (Piedmont, Italy). Plants, 9(6), 708. https://doi.org/10.3390/plants9060708