Physiological and Biochemical Changes in Sugar Beet Seedlings to Confer Stress Adaptability under Drought Condition

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Treatment
2.2. Determination of Plant Height, Fresh Weight, and Dry Weight
2.3. Photosynthetic Pigments Analysis
2.3.1. Chlorophyll (Chl) and Carotenoid (Car) Determination
2.3.2. Determination of Photosystem II Quantum Yield
2.4. Physiological Traits
2.4.1. Leaf Relative Water Content
2.4.2. Drought Tolerance Index
2.4.3. The Membrane Stability Index (MSI)
2.5. Determination of Lipid Peroxidation
2.6. Estimation of Hydrogen Peroxide (H2O2)
2.7. Determination of Osmolytes
2.7.1. Free Proline Content
2.7.2. Glycine Betaine Content
2.7.3. Total Soluble Carbohydrate (TSC) and Sucrose Content
2.8. Determination of Antioxidant Enzyme Activities
2.9. Determination of Total Phenolic Content, Total Flavonoid Content, and Antioxidant Capacity
2.10. Statistical Analysis
3. Results and Discussions
3.1. Response to Drought Stress on Plant Growth Characteristics
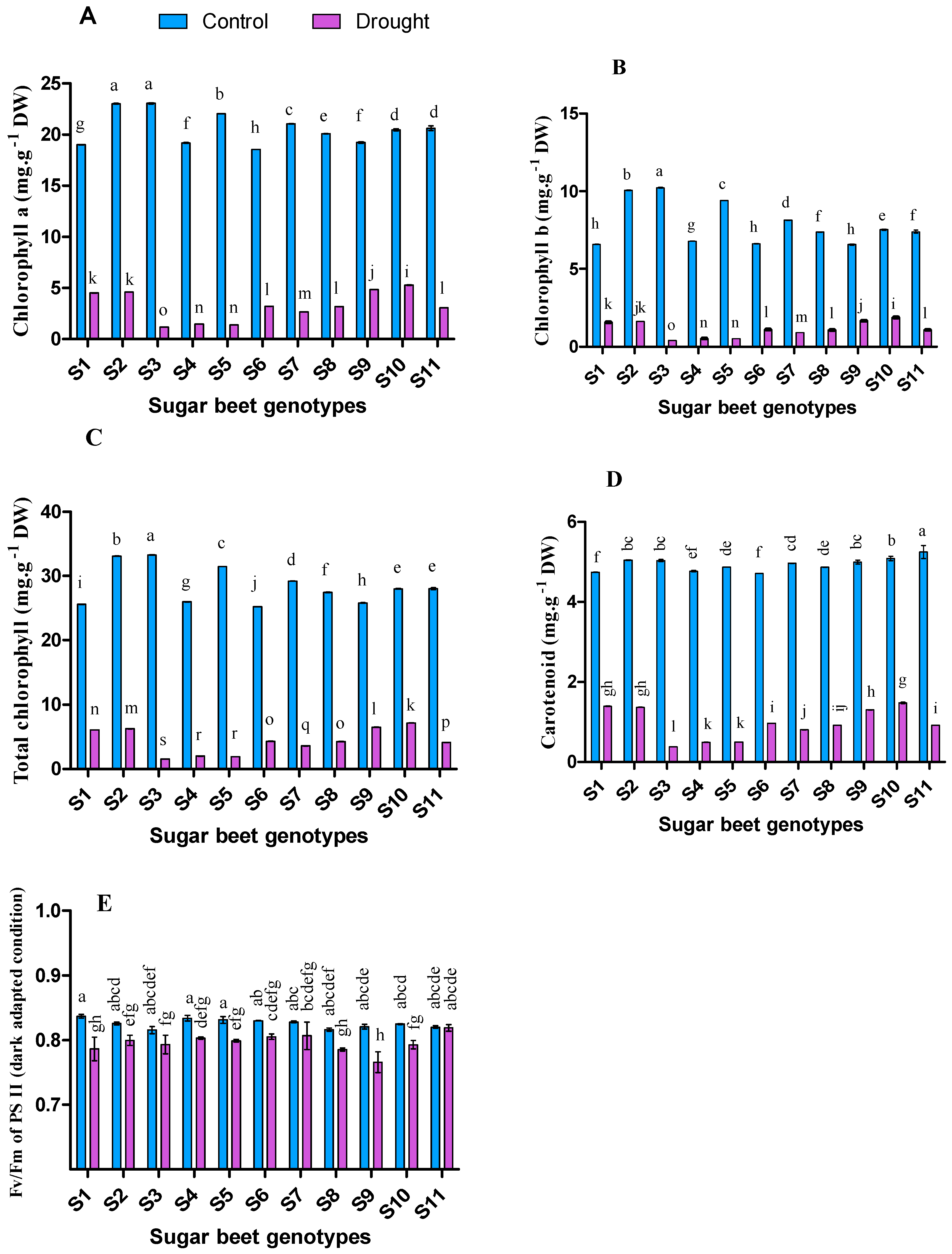
3.2. Influence of Drought on Photosynthetic Pigments and Fluorescence
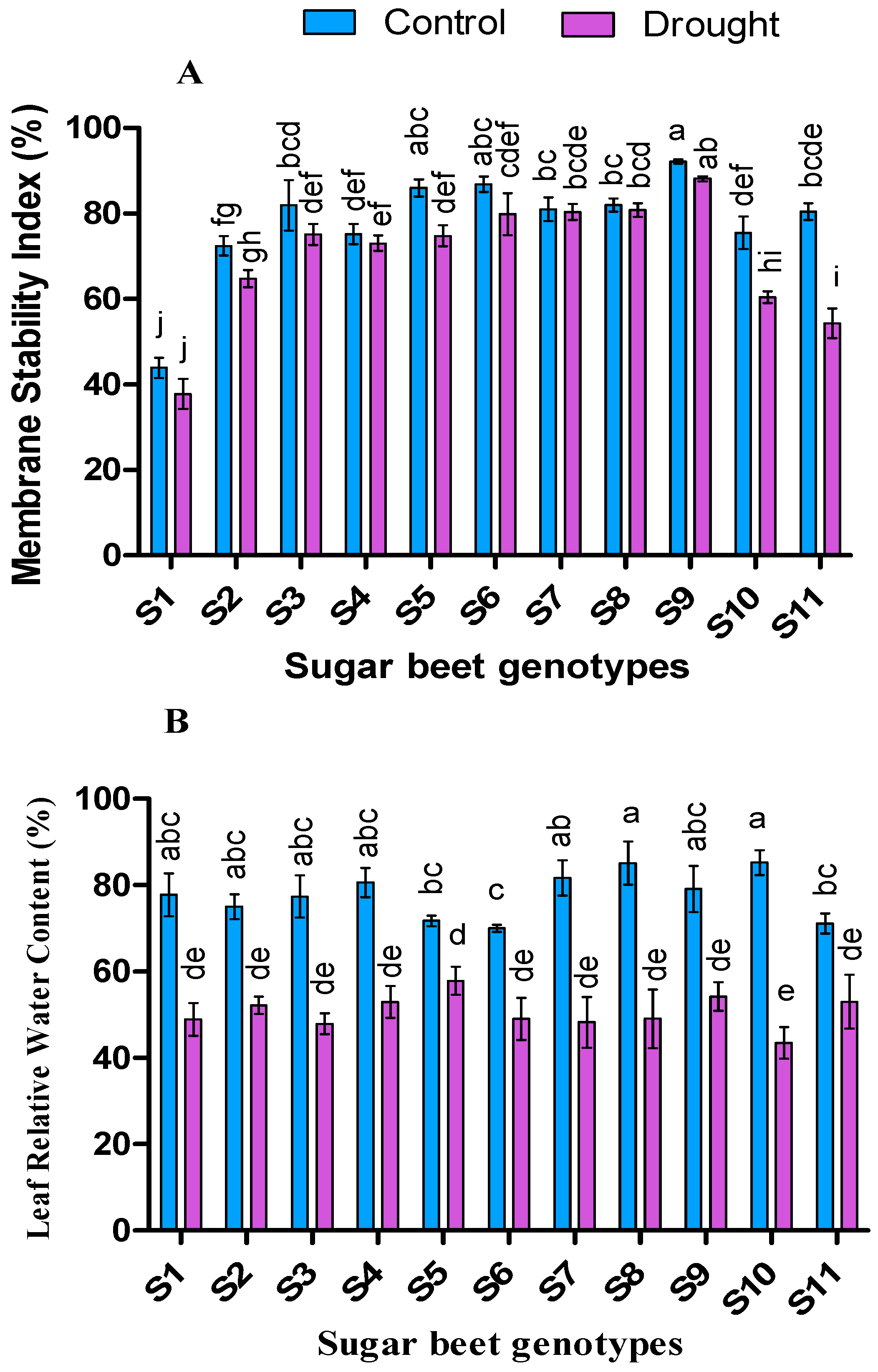
3.3. Effect of Drought on Drought Tolerance Index, Relative Water Content (%), and Membrane Stability Index (%)
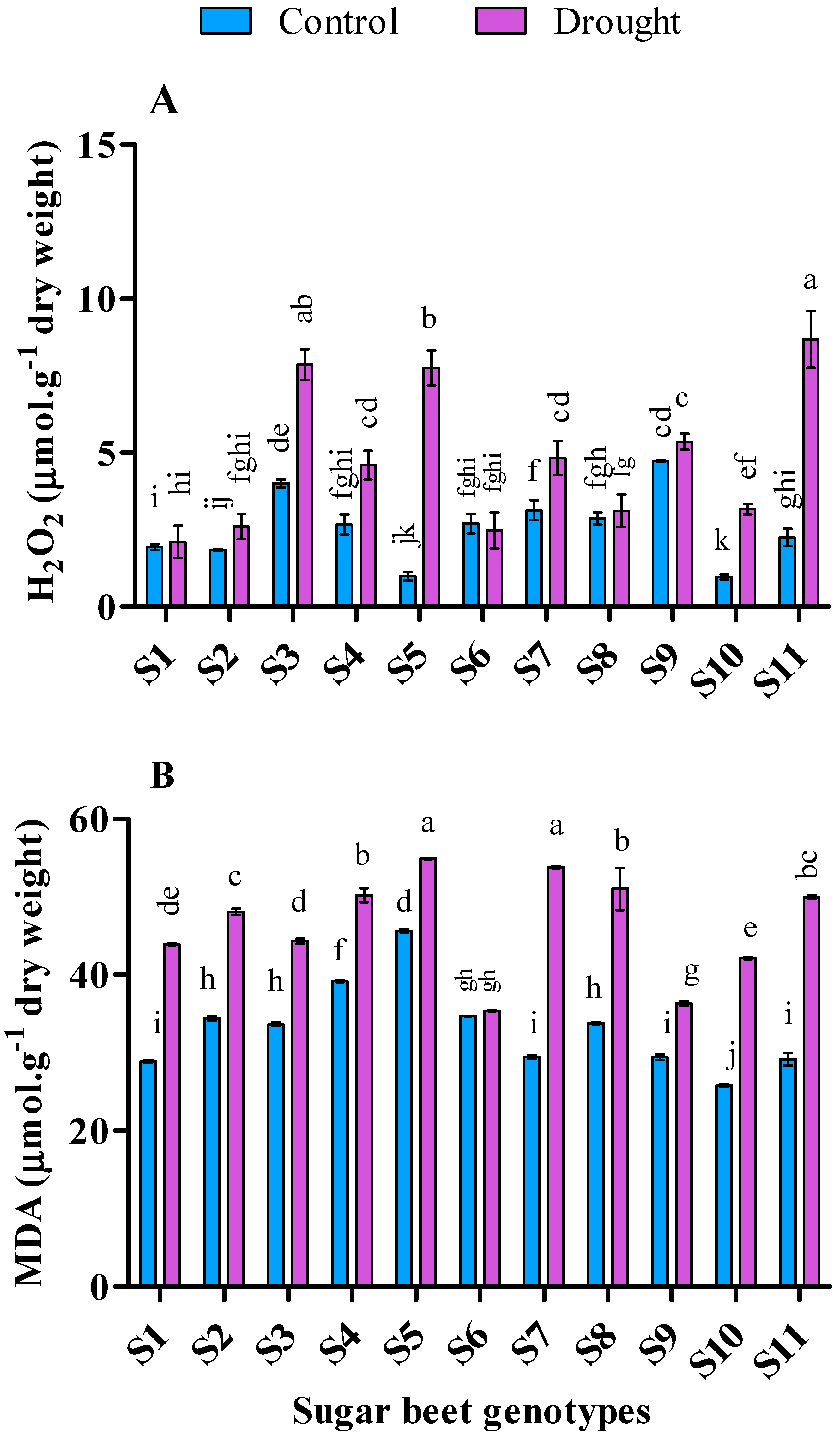
3.4. Influence of Drought Stress on Lipid Peroxidation and Hydrogen Peroxide
3.5. Effect of Drought on Osmotic Adjustment Molecules
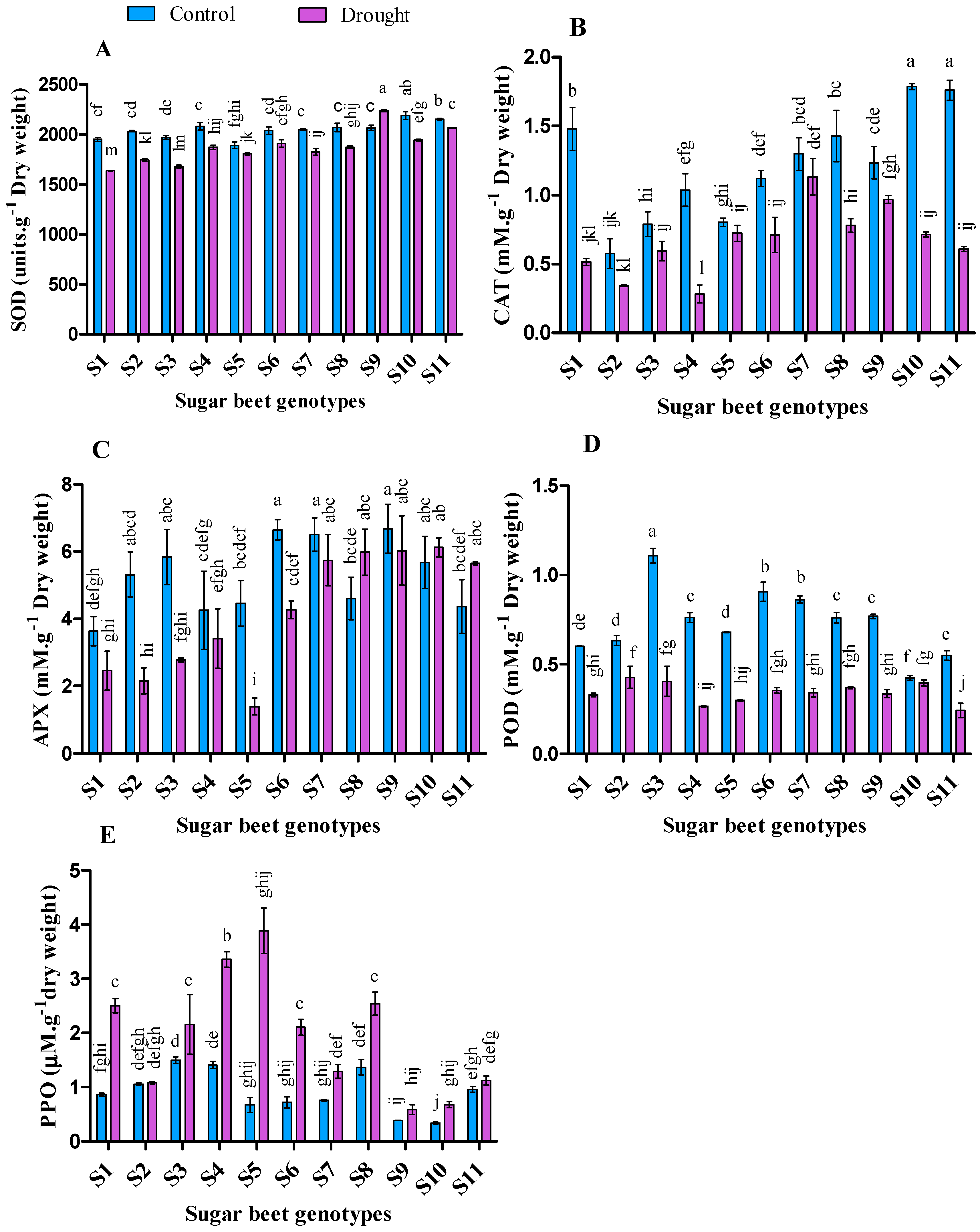
3.6. Effect of Drought Stresses on Antioxidant Enzymes Activities
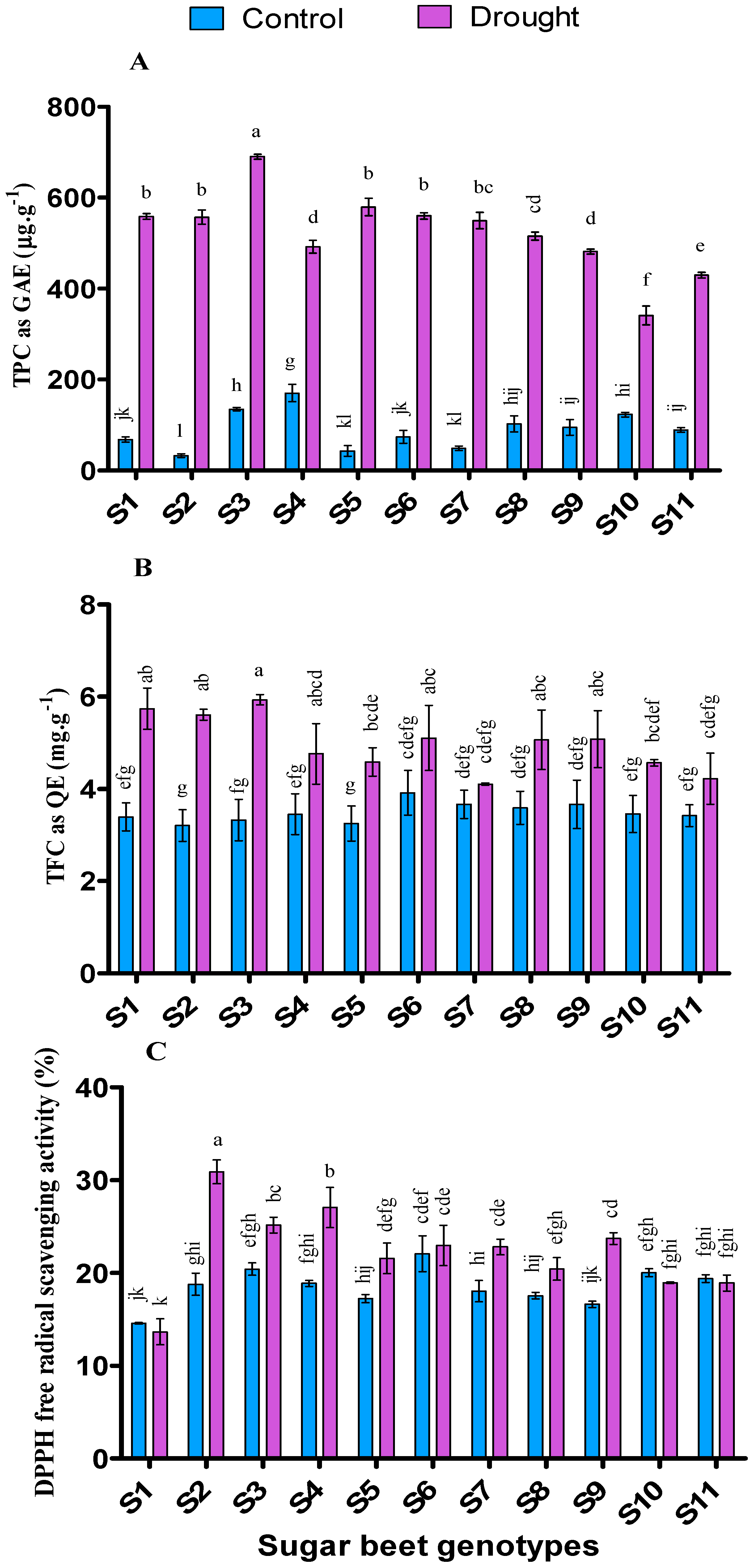
3.7. Influence of Drought Stresses on Antioxidant Activities
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- He, M.; He, C.-Q.; Ding, N.-Z. Abiotic stresses: General defenses of land plants and chances for engineering multistress tolerance. Front. Plant Sci. 2018, 9, 1771. [Google Scholar] [CrossRef] [PubMed]
- Jeandroz, S.; Lamotte, O. Plant responses to biotic and abiotic stresses: Lessons from cell signaling. Front. Plant Sci. 2017, 8, 1772. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Achievements and challenges in understanding plant abiotic stress responses and tolerance. Plant Cell Physiol. 2011, 52, 1569–1582. [Google Scholar] [CrossRef] [PubMed]
- Luković, J.; Maksimović, I.; Zorić, L.; Nagl, N.; Perčić, M.; Polić, D.; Putnik-Delić, M. Histological characteristics of sugar beet leaves potentially linked to drought tolerance. Ind. Crops Prod. 2009, 30, 281–286. [Google Scholar] [CrossRef]
- Islam, J.; Choi, S.P.; Azad, O.K.; Kim, J.W.; Lim, Y.-S. Evaluation of tuber yield and marketable quality of newly developed thirty-two potato varieties grown in three different ecological zones in South Korea. Agriculture 2020, 10, 327. [Google Scholar] [CrossRef]
- Chołuj, D.; Wiśniewska, A.; Szafrański, K.M.; Cebula, J.; Gozdowski, D.; Podlaski, S. Assessment of the physiological responses to drought in different sugar beet genotypes in connection with their genetic distance. J. Plant Physiol. 2014, 171, 1221–1230. [Google Scholar] [CrossRef]
- Sadeghian, S.Y.; Fazli, H.; Mohammadian, R.; Taleghani, D.F.; Mesbah, M. Genetic variation for drought stress in sugarbeet. J. Sugar Beet Res. 2000, 37, 55–78. [Google Scholar] [CrossRef]
- Zhang, L.; Zhou, T. Drought over East Asia: A review. J. Clim. 2015, 28, 3375–3399. [Google Scholar] [CrossRef]
- Kwon, H.; Lall, U.; Kim, S. The unusual 2013–2015 drought in South Korea in the context of a multicentury precipitation record: Inferences from a nonstationary, multivariate, Bayesian copula model. Geophys. Res. Lett. 2016, 43, 8534–8544. [Google Scholar] [CrossRef]
- Kim, C.J.; Park, M.J.; Lee, J.H. Analysis of climate change impacts on the spatial and frequency patterns of drought using a potential drought hazard mapping approach. Int. J. Climatol. 2014, 34, 61–80. [Google Scholar] [CrossRef]
- Schuppler, U.; He, P.-H.; John, P.C.L.; Munns, R. Effect of water stress on cell division and Cdc2-like cell cycle kinase activity in wheat leaves. Plant Physiol. 1998, 117, 667–678. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Manivannan, P.; Lakshmanan, G.M.A.; Gomathinayagam, M.; Panneerselvam, R. Alterations in morphological parameters and photosynthetic pigment responses of Catharanthus roseus under soil water deficits. Colloids Surf. B Biointerfaces 2008, 61, 298–303. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Gopi, R.; Manivannan, P.; Gomathinayagam, M.; Sridharan, R.; Panneerselvam, R. Antioxidant potential and indole alkaloid profile variations with water deficits along different parts of two varieties of Catharanthus roseus. Colloids Surf. B Biointerfaces 2008, 62, 312–318. [Google Scholar] [CrossRef]
- Rampino, P.; Pataleo, S.; Gerardi, C.; Mita, G.; Perrotta, C. Drought stress response in wheat: Physiological and molecular analysis of resistant and sensitive genotypes. Plant. Cell Environ. 2006, 29, 2143–2152. [Google Scholar] [CrossRef]
- Nouri, A.; Etminan, A.; Teixeira da Silva, J.A.; Mohammadi, R. Assessment of yield, yield-related traits and drought tolerance of durum wheat genotypes (Triticum turgidum var. durum Desf.). Aust. J. Crop Sci. 2011, 5, 8. [Google Scholar]
- Fukai, S.; Fischer, K.S. Field phenotyping strategies and breeding for adaptation of rice to drought. Front. Physiol. 2012, 3, 282. [Google Scholar]
- Munjal, R.; Dhanda, S.S. Assessment of drought resistance in Indian wheat cultivars for morpho-physiological traits. Ekin J. Crop Breed. Genet. 2016, 2, 74–81. [Google Scholar]
- Lebaudy, A.; Vavasseur, A.; Hosy, E.; Dreyer, I.; Leonhardt, N.; Thibaud, J.-B.; Véry, A.-A.; Simonneau, T.; Sentenac, H. Plant adaptation to fluctuating environment and biomass production are strongly dependent on guard cell potassium channels. Proc. Natl. Acad. Sci. USA 2008, 105, 5271–5276. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Neményi, A.; Bőcs, A.; Pék, Z.; Helyes, L. Physiological factors and their relationship with the productivity of processing tomato under different water supplies. Water 2019, 11, 586. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Molnár, K.; Pék, Z.; Helyes, L. Effect of water supply on the water use-related physiological traits and yield of snap beans in dry seasons. Irrig. Sci. 2018, 36, 143–158. [Google Scholar] [CrossRef]
- Mundim, F.M.; Pringle, E.G. Whole-plant metabolic allocation under water stress. Front. Plant Sci. 2018, 9, 852. [Google Scholar] [CrossRef]
- Chaves, M.M.; Oliveira, M.M. Mechanisms underlying plant resilience to water deficits: Prospects for water-saving agriculture. J. Exp. Bot. 2004, 55, 2365–2384. [Google Scholar] [CrossRef]
- Chan, K.X.; Wirtz, M.; Phua, S.Y.; Estavillo, G.M.; Pogson, B.J. Balancing metabolites in drought: The sulfur assimilation conundrum. Trends Plant Sci. 2013, 18, 18–29. [Google Scholar] [CrossRef]
- Zivcak, M.; Brestic, M.; Sytar, O. Osmotic adjustment and plant adaptation to drought stress. In Drought Stress Tolerance in Plants; Springer: Cham, Switzerland, 2016; Volume 1, pp. 105–143. [Google Scholar]
- Nemeskeri, E.; Kovacs-Nagy, E.; Nyeki, J.; Sardi, E. Responses of apple tree cultivars to drought: Carbohydrate composition in the leaves. Turk. J. Agric. For. 2015, 39, 949–957. [Google Scholar] [CrossRef]
- Guo, R.; Shi, L.; Jiao, Y.; Li, M.; Zhong, X.; Gu, F.; Liu, Q.; Xia, X.; Li, H. Metabolic responses to drought stress in the tissues of drought-tolerant and drought-sensitive wheat genotype seedlings. AoB Plants 2018, 10, ply016. [Google Scholar] [CrossRef]
- Zabalza, A.; Gálvez, L.; Marino, D.; Royuela, M.; Arrese-Igor, C.; González, E.M. The application of ascorbate or its immediate precursor, galactono-1, 4-lactone, does not affect the response of nitrogen-fixing pea nodules to water stress. J. Plant Physiol. 2008, 165, 805–812. [Google Scholar] [CrossRef]
- Smirnoff, N. Tansley review no. 52. The role of active oxygen in the response of plants to water deficit and desiccation. New Phytol. 1993, 125, 27–58. [Google Scholar] [CrossRef]
- Bi, A.; Fan, J.; Hu, Z.; Wang, G.; Amombo, E.; Fu, J.; Hu, T. Differential acclimation of enzymatic antioxidant metabolism and photosystem II photochemistry in tall fescue under drought and heat and the combined stresses. Front. Plant Sci. 2016, 7, 453. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.; Corpas, F.J.; Borsani, O.; Barroso, J.B.; Monza, J. Water stress induces a differential and spatially distributed nitro-oxidative stress response in roots and leaves of Lotus japonicus. Plant Sci. 2013, 201, 137–146. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. The roles of reactive oxygen metabolism in drought: Not so cut and dried. Plant Physiol. 2014, 164, 1636–1648. [Google Scholar] [CrossRef]
- Scandalios, J.G.; Guan, L.; Polidoros, A.N. Catalases in plants: Gene structure, properties, regulation, and expression. Cold Spring Harb. Monogr. Ser. 1997, 34, 343–406. [Google Scholar]
- Ali, B.; Hasan, S.A.; Hayat, S.; Hayat, Q.; Yadav, S.; Fariduddin, Q.; Ahmad, A. A role for brassinosteroids in the amelioration of aluminium stress through antioxidant system in mung bean (Vigna radiata L. Wilczek). Environ. Exp. Bot. 2008, 62, 153–159. [Google Scholar] [CrossRef]
- Rubio, M.C.; González, E.M.; Minchin, F.R.; Webb, K.J.; Arrese-Igor, C.; Ramos, J.; Becana, M. Effects of water stress on antioxidant enzymes of leaves and nodules of transgenic alfalfa overexpressing superoxide dismutases. Physiol. Plant. 2002, 115, 531–540. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, J. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Liu, C.-J.; Tschaplinski, T.J.; Zhao, N. Genomics of secondary metabolism in Populus: Interactions with biotic and abiotic environments. Crit. Rev. Plant Sci. 2009, 28, 375–392. [Google Scholar] [CrossRef]
- Ncube, B.; Van Staden, J. Tilting plant metabolism for improved metabolite biosynthesis and enhanced human benefit. Molecules 2015, 20, 12698–12731. [Google Scholar] [CrossRef]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Islam, M.J.; Hassan, M.K.; Sarker, S.R.; Rahman, A.B.; Fakir, M.S.A. Light and temperature effects on sprout yield and its proximate composition and vitamin C content in lignosus and mung beans. J. Bangladesh Agric. Univ. 2017, 15, 248–254. [Google Scholar] [CrossRef]
- Bharti, N.; Yadav, D.; Barnawal, D.; Maji, D.; Kalra, A. Exiguobacterium oxidotolerans, a halotolerant plant growth promoting rhizobacteria, improves yield and content of secondary metabolites in Bacopa monnieri (L.) Pennell under primary and secondary salt stress. World J. Microbiol. Biotechnol. 2013, 29, 379–387. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.-M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef]
- Ferrat, L.; Pergent-Martini, C.; Roméo, M. Assessment of the use of biomarkers in aquatic plants for the evaluation of environmental quality: Application to seagrasses. Aquat. Toxicol. 2003, 65, 187–204. [Google Scholar] [CrossRef]
- Abo-Elwafa, S.F.; Abdel-Rahim, H.M.; Abou-Salama, A.M.; Teama, E.A. Sugar beet floral induction and fertility: Effect of vernalization and day-length extension. Sugar Tech 2006, 8, 281–287. [Google Scholar] [CrossRef]
- Brar, N.S.; Dhillon, B.S.; Saini, K.S.; Sharma, P.K. Agronomy of sugar beet cultivation—A review. Agric. Rev. 2015, 36, 184–197. [Google Scholar] [CrossRef]
- Mutasa-Göttgens, E.S.; Qi, A.; Zhang, W.; Schulze-Buxloh, G.; Jennings, A.; Hohmann, U.; Müller, A.E.; Hedden, P. Bolting and flowering control in sugar beet: Relationships and effects of gibberellin, the bolting gene B and vernalization. AoB Plants 2010, 2010, plq012. [Google Scholar] [CrossRef]
- Publication. Available online: https://web.kma.go.kr/eng/aboutkma/webzine.jsp (accessed on 15 March 2020).
- Sunrise and sunset in South Korea. Available online: https://www.worlddata.info/asia/south-korea/sunset.php (accessed on 15 March 2020).
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1987; Volume 148, pp. 350–382. ISBN 0076-6879. [Google Scholar]
- Meng, L.-L.; Song, J.-F.; Wen, J.; Zhang, J.; Wei, J.-H. Effects of drought stress on fluorescence characteristics of photosystem II in leaves of Plectranthus scutellarioides. Photosynthetica 2016, 54, 414–421. [Google Scholar] [CrossRef]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- Fernandez, G.C.J. Effective selection criteria for assessing plant stress tolerance. In Proceedings of the International Symposium on Adaptation of Vegetables and other Food Crops in Temperature and Water Stress, Shanhua, Taiwan, 13–16 August 1992; pp. 257–270. [Google Scholar]
- Ober, E.S.; Clark, C.J.A.; Le Bloa, M.; Royal, A.; Jaggard, K.W.; Pidgeon, J.D. Assessing the genetic resources to improve drought tolerance in sugar beet: Agronomic traits of diverse genotypes under droughted and irrigated conditions. F. Crop. Res. 2004, 90, 213–234. [Google Scholar] [CrossRef]
- Sairam, R.K.; Rao, K.V.; Srivastava, G.C. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Singh, N.; Ma, L.Q.; Srivastava, M.; Rathinasabapathi, B. Metabolic adaptations to arsenic-induced oxidative stress in Pteris vittata L. and Pteris ensiformis L. Plant Sci. 2006, 170, 274–282. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Xu, Z.; Sun, M.; Jiang, X.; Sun, H.; Dang, X.; Cong, H.; Qiao, F. Glycinebetaine biosynthesis in response to osmotic stress depends on jasmonate signaling in watermelon suspension cells. Front. Plant Sci. 2018, 9, 1469. [Google Scholar] [CrossRef]
- Khoyerdi, F.F.; Shamshiri, M.H.; Estaji, A. Changes in some physiological and osmotic parameters of several pistachio genotypes under drought stress. Sci. Hortic. (Amsterdam) 2016, 198, 44–51. [Google Scholar] [CrossRef]
- Van Handel, E. Direct microdetermination of sucrose. Anal. Biochem. 1968, 22, 280–283. [Google Scholar] [CrossRef]
- Slabbert, M.M.; Krüger, G.H.J. Antioxidant enzyme activity, proline accumulation, leaf area and cell membrane stability in water stressed Amaranthus leaves. South Afr. J. Bot. 2014, 95, 123–128. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Zhang, X.Z. The measurement and mechanism of lipid peroxidation and SOD, POD and CAT activities in biological system. Res. Methodol. Crop Physiol. Agric. Press. Beijing 1992, 15, 208–211. [Google Scholar]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Tagele, S.B.; Kim, S.W.; Lee, H.G.; Lee, Y.S. Aggressiveness and fumonisins production of Fusarium Subglutinans and Fusarium Temperatum on Korean maize cultivars. Agronomy 2019, 9, 88. [Google Scholar] [CrossRef]
- Mayer, A.M.; Harel, E.; Ben-Shaul, R. Assay of catechol oxidase—A critical comparison of methods. Phytochemistry 1966, 5, 783–789. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Adnan, M.; Azad, M.O.K.; Ju, H.S.; Son, J.M.; Park, C.H.; Shin, M.H.; Alle, M.; Cho, D.H. Development of biopolymer-mediated nanocomposites using hot-melt extrusion to enhance the bio-accessibility and antioxidant capacity of kenaf seed flour. Appl. Nanosci. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Braca, A.; Fico, G.; Morelli, I.; De Simone, F.; Tomè, F.; De Tommasi, N. Antioxidant and free radical scavenging activity of flavonol glycosides from different Aconitum species. J. Ethnopharmacol. 2003, 86, 63–67. [Google Scholar] [CrossRef]
- Shaw, B.; Thomas, T.H.; Cooke, D.T. Responses of sugar beet (Beta vulgaris L.) to drought and nutrient deficiency stress. Plant Growth Regul. 2002, 37, 77–83. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.H.; Terry, N.; Sears, T.; van Dun, K. Enhanced drought resistance in fructan-producing sugar beet. Plant Physiol. Biochem. 1999, 37, 313–317. [Google Scholar] [CrossRef]
- Ribaut, J.-M.; Betran, J.; Monneveux, P.; Setter, T. Drought tolerance in maize. In Handbook of Maize: Its Biology; Springer: New York, NY, USA, 2009; pp. 311–344. [Google Scholar]
- Lobell, D.B.; Roberts, M.J.; Schlenker, W.; Braun, N.; Little, B.B.; Rejesus, R.M.; Hammer, G.L. Greater sensitivity to drought accompanies maize yield increase in the US Midwest. Science 2014, 344, 516–519. [Google Scholar] [CrossRef]
- Mou, B.; He, B.-J.; Zhao, D.-X.; Chau, K. Numerical simulation of the effects of building dimensional variation on wind pressure distribution. Eng. Appl. Comput. Fluid Mech. 2017, 11, 293–309. [Google Scholar] [CrossRef]
- Efeoğlu, B.; Ekmekçi, Y.; Çiçek, N. Physiological responses of three maize cultivars to drought stress and recovery. South Afr. J. Bot. 2009, 75, 34–42. [Google Scholar] [CrossRef]
- Hosseini, S.A.; Réthoré, E.; Pluchon, S.; Ali, N.; Billiot, B.; Yvin, J.-C. Calcium application enhances drought stress tolerance in sugar beet and promotes plant biomass and beetroot sucrose concentration. Int. J. Mol. Sci. 2019, 20, 3777. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Mibei, E.K.; Ambuko, J.; Giovannoni, J.J.; Onyango, A.N.; Owino, W.O. Carotenoid profiling of the leaves of selected African eggplant accessions subjected to drought stress. Food Sci. Nutr. 2017, 5, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Kaewsuksaeng, S. Chlorophyll degradation in horticultural crops. Walailak J. Sci. Technol. 2011, 8, 9–19. [Google Scholar]
- Ekmekci, Y.; Bohms, A.; Thomson, J.A.; Mundree, S.G. Photochemical and antioxidant responses in the leaves of Xerophyta viscosa Baker and Digitaria sanguinalis L. under water deficit. Z. Nat. C 2005, 60, 435–443. [Google Scholar] [CrossRef]
- Souza, R.P.; Machado, E.C.; Silva, J.A.B.; Lagôa, A.; Silveira, J.A.G. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery. Environ. Exp. Bot. 2004, 51, 45–56. [Google Scholar] [CrossRef]
- Akhkha, A. The effect of water stress on photosynthesis, respiration and relative chlorophyll index of the desert plant Calotropis procera. Biosci. Biotechnol. Res. Asia 2016, 6, 653–658. [Google Scholar]
- Hasheminasab, H.; Aliakbari, A.; Baniasadi, R. Optimizing the relative water protection (RWP) as novel approach for monitoring drought tolerance in Iranian pistachio cultivars using graphical analysis. Int. J. Biosci. 2014, 4, 194–204. [Google Scholar]
- Min, Z.; Li, R.; Chen, L.; Zhang, Y.; Li, Z.; Liu, M.; Ju, Y.; Fang, Y. Alleviation of drought stress in grapevine by foliar-applied strigolactones. Plant Physiol. Biochem. 2019, 135, 99–110. [Google Scholar] [CrossRef]
- Van Ha, C.; Leyva-González, M.A.; Osakabe, Y.; Tran, U.T.; Nishiyama, R.; Watanabe, Y.; Tanaka, M.; Seki, M.; Yamaguchi, S.; Van Dong, N. Positive regulatory role of strigolactone in plant responses to drought and salt stress. Proc. Natl. Acad. Sci. USA 2014, 111, 851–856. [Google Scholar]
- Yeilaghi, H.; Arzani, A.; Ghaderian, M.; Fotovat, R.; Feizi, M.; Pourdad, S.S. Effect of salinity on seed oil content and fatty acid composition of safflower (Carthamus tinctorius L.) genotypes. Food Chem. 2012, 130, 618–625. [Google Scholar] [CrossRef]
- Ghaffari, H.; Tadayon, M.R.; Nadeem, M.; Cheema, M.; Razmjoo, J. Proline-mediated changes in antioxidant enzymatic activities and the physiology of sugar beet under drought stress. Acta Physiol. Plant. 2019, 41, 23. [Google Scholar] [CrossRef]
- Al-Jbawi, E.; Abbas, F. The effect of length during drought stress on sugar beet (Beta vulgaris L.) yield and quality. Persian Gulf Crop Prot. 2013, 2, 35–43. [Google Scholar]
- Lin, C.C.; Kao, C.H. Effect of NaCl stress on H2O2 metabolism in rice leaves. Plant Growth Regul. 2000, 30, 151–155. [Google Scholar] [CrossRef]
- Hernández, J.A.; Almansa, M.S. Short-term effects of salt stress on antioxidant systems and leaf water relations of pea leaves. Physiol. Plant. 2002, 115, 251–257. [Google Scholar] [CrossRef]
- Scandalios, J.G. Oxygen stress and superoxide dismutases. Plant Physiol. 1993, 101, 7. [Google Scholar] [CrossRef]
- Borišev, M.; Borišev, I.; Župunski, M.; Arsenov, D.; Pajević, S.; Ćurčić, Ž.; Vasin, J.; Djordjevic, A. Drought impact is alleviated in sugar beets (Beta vulgaris L.) by foliar application of fullerenol nanoparticles. PLoS ONE 2016, 11, e0166248. [Google Scholar] [CrossRef]
- Zhao, L.; Peng, B.; Hernandez-Viezcas, J.A.; Rico, C.; Sun, Y.; Peralta-Videa, J.R.; Tang, X.; Niu, G.; Jin, L.; Varela-Ramirez, A. Stress response and tolerance of Zea mays to CeO2 nanoparticles: Cross talk among H2O2, heat shock protein, and lipid peroxidation. ACS Nano 2012, 6, 9615–9622. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought stress effects on growth, ROS markers, compatible solutes, phenolics, flavonoids, and antioxidant activity in Amaranthus tricolor. Appl. Biochem. Biotechnol. 2018, 186, 999–1016. [Google Scholar] [CrossRef]
- Putnik-Delić, M.I.; Maksimović, I.V.; Nikolić-Đorić, E.B.; Nagl, N.M. Analyses of statistical transformations of row data describing free proline concentration in sugar beet exposed to drought. Zb. Matice Srp. Prir. Nauk. 2010, 2010, 7–16. [Google Scholar] [CrossRef]
- Szabados, L.; Savoure, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Hong-Bo, S.; Xiao-Yan, C.; Li-Ye, C.; Xi-Ning, Z.; Gang, W.; Yong-Bing, Y.; Chang-Xing, Z.; Zan-Min, H. Investigation on the relationship of proline with wheat anti-drought under soil water deficits. Colloids Surf. B Biointerfaces 2006, 53, 113–119. [Google Scholar] [CrossRef]
- Farooq, A.; Bukhari, S.A.; Akram, N.A.; Ashraf, M.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Exogenously applied ascorbic acid-mediated changes in osmoprotection and oxidative defense system enhanced water stress tolerance in different cultivars of safflower (Carthamus tinctorious L.). Plants 2020, 9, 104. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Anjum, S.A.; Wang, L.C.; Farooq, M.; Hussain, M.; Xue, L.L.; Zou, C.M. Brassinolide application improves the drought tolerance in maize through modulation of enzymatic antioxidants and leaf gas exchange. J. Agron. Crop Sci. 2011, 197, 177–185. [Google Scholar] [CrossRef]
- Genard, H.; Le Saos, J.; Billard, J.-P.; Tremolieres, A.; Boucaud, J. Effect of salinity on lipid composition, glycine betaine content and photosynthetic activity in chloroplasts of Suaeda maritima. Plant Physiol. Biochem. 1991, 29, 421–427. [Google Scholar]
- Giri, J. Glycinebetaine and abiotic stress tolerance in plants. Plant Signal. Behav. 2011, 6, 1746–1751. [Google Scholar] [CrossRef]
- Hoekstra, F.A.; Golovina, E.A.; Buitink, J. Mechanisms of plant desiccation tolerance. Trends Plant Sci. 2001, 6, 431–438. [Google Scholar] [CrossRef]
- Karimi, S.; Yadollahi, A.; Arzani, K. Responses of almond genotypes to osmotic stress induced in vitro. J. Nuts 2013, 4, 1–7. [Google Scholar]
- Ashraf, M. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv. 2009, 27, 84–93. [Google Scholar] [CrossRef]
- Wang, W.-B.; Kim, Y.-H.; Lee, H.-S.; Kim, K.-Y.; Deng, X.-P.; Kwak, S.-S. Analysis of antioxidant enzyme activity during germination of alfalfa under salt and drought stresses. Plant Physiol. Biochem. 2009, 47, 570–577. [Google Scholar] [CrossRef]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2002, 53, 1331–1341. [Google Scholar] [CrossRef]
- Jaleel, C.A.; Riadh, K.; Gopi, R.; Manivannan, P.; Inès, J.; Al-Juburi, H.J.; Chang-Xing, Z.; Hong-Bo, S.; Panneerselvam, R. Antioxidant defense responses: Physiological plasticity in higher plants under abiotic constraints. Acta Physiol. Plant. 2009, 31, 427–436. [Google Scholar] [CrossRef]
- DaCosta, M.; Huang, B. Changes in antioxidant enzyme activities and lipid peroxidation for bentgrass species in response to drought stress. J. Am. Soc. Hortic. Sci. 2007, 132, 319–326. [Google Scholar] [CrossRef]
- Almeselmani, M.; Deshmukh, P.S.; Sairam, R.K.; Kushwaha, S.R.; Singh, T.P. Protective role of antioxidant enzymes under high temperature stress. Plant Sci. 2006, 171, 382–388. [Google Scholar] [CrossRef]
- Shen, C.; Zhang, Q.; Li, J.; Bi, F.; Yao, N. Induction of programmed cell death in Arabidopsis and rice by single-wall carbon nanotubes. Am. J. Bot. 2010, 97, 1602–1609. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Sekmen, A.H.; Ozgur, R.; Uzilday, B.; Turkan, I. Reactive oxygen species scavenging capacities of cotton (Gossypium hirsutum) cultivars under combined drought and heat induced oxidative stress. Environ. Exp. Bot. 2014, 99, 141–149. [Google Scholar] [CrossRef]
- Alam, M.M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Exogenous jasmonic acid modulates the physiology, antioxidant defense and glyoxalase systems in imparting drought stress tolerance in different Brassica species. Plant Biotechnol. Rep. 2014, 8, 279–293. [Google Scholar] [CrossRef]
- Boeckx, T.; Winters, A.L.; Webb, K.J.; Kingston-Smith, A.H. Polyphenol oxidase in leaves: Is there any significance to the chloroplastic localization? J. Exp. Bot. 2015, 66, 3571–3579. [Google Scholar] [CrossRef]
- Lee, B.-R.; Kim, K.-Y.; Jung, W.-J.; Avice, J.-C.; Ourry, A.; Kim, T.-H. Peroxidases and lignification in relation to the intensity of water-deficit stress in white clover (Trifolium repens L.). J. Exp. Bot. 2007, 58, 1271–1279. [Google Scholar] [CrossRef]
- Thipyapong, P.; Melkonian, J.; Wolfe, D.W.; Steffens, J.C. Suppression of polyphenol oxidases increases stress tolerance in tomato. Plant Sci. 2004, 167, 693–703. [Google Scholar] [CrossRef]
- Mayer, A.M.; Harel, E. Polyphenol oxidases in plants. Phytochemistry 1979, 18, 193–215. [Google Scholar] [CrossRef]
- Vaughn, K.C.; Duke, S.O. Function of polyphenol oxidase in higher plants. Physiol. Plant. 1984, 60, 106–112. [Google Scholar] [CrossRef]
- Halliwell, B. Hydroxylation of p-Coumaric acid by illuminated chloroplasts. The role of superoxide. Eur. J. Biochem. 1975, 55, 355–360. [Google Scholar] [CrossRef]
- Steffens, J.C.; Harel, E.; Hunt, M.D. Polyphenol oxidase. In Genetic Engineering of Plant Secondary Metabolism; Springer: Boston, MA, USA, 1994; pp. 275–312. [Google Scholar]
- Sairam, R.K.; Srivastava, G.C.; Saxena, D.C. Increased antioxidant activity under elevated temperatures: A mechanism of heat stress tolerance in wheat genotypes. Biol. Plant. 2000, 43, 245–251. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef]
- Bartwal, A.; Mall, R.; Lohani, P.; Guru, S.K.; Arora, S. Role of secondary metabolites and brassinosteroids in plant defense against environmental stresses. J. Plant Growth Regul. 2013, 32, 216–232. [Google Scholar] [CrossRef]
- Gharibi, S.; Tabatabaei, B.E.S.; Saeidi, G.; Goli, S.A.H. Effect of drought stress on total phenolic, lipid peroxidation, and antioxidant activity of Achillea species. Appl. Biochem. Biotechnol. 2016, 178, 796–809. [Google Scholar] [CrossRef]
- Easwar Rao, D.; Viswanatha Chaitanya, K. Changes in the antioxidant intensities of seven different soybean (Glycine max (L.) Merr.) cultivars during drought. J. Food Biochem. 2019, 44, e13118. [Google Scholar] [CrossRef]
- Espinoza, A.; San Martín, A.; López-Climent, M.; Ruiz-Lara, S.; Gómez-Cadenas, A.; Casaretto, J.A. Engineered drought-induced biosynthesis of α-tocopherol alleviates stress-induced leaf damage in tobacco. J. Plant Physiol. 2013, 170, 1285–1294. [Google Scholar] [CrossRef]
- Bettaieb, I.; Hamrouni-Sellami, I.; Bourgou, S.; Limam, F.; Marzouk, B. Drought effects on polyphenol composition and antioxidant activities in aerial parts of Salvia officinalis L. Acta Physiol. Plant. 2011, 33, 1103–1111. [Google Scholar] [CrossRef]
- Lin, K.-H.; Chao, P.-Y.; Yang, C.-M.; Cheng, W.-C.; Lo, H.-F.; Chang, T.-R. The effects of flooding and drought stresses on the antioxidant constituents in sweet potato leaves. Bot. Stud. 2006, 47, 417–426. [Google Scholar]







{kind=link}
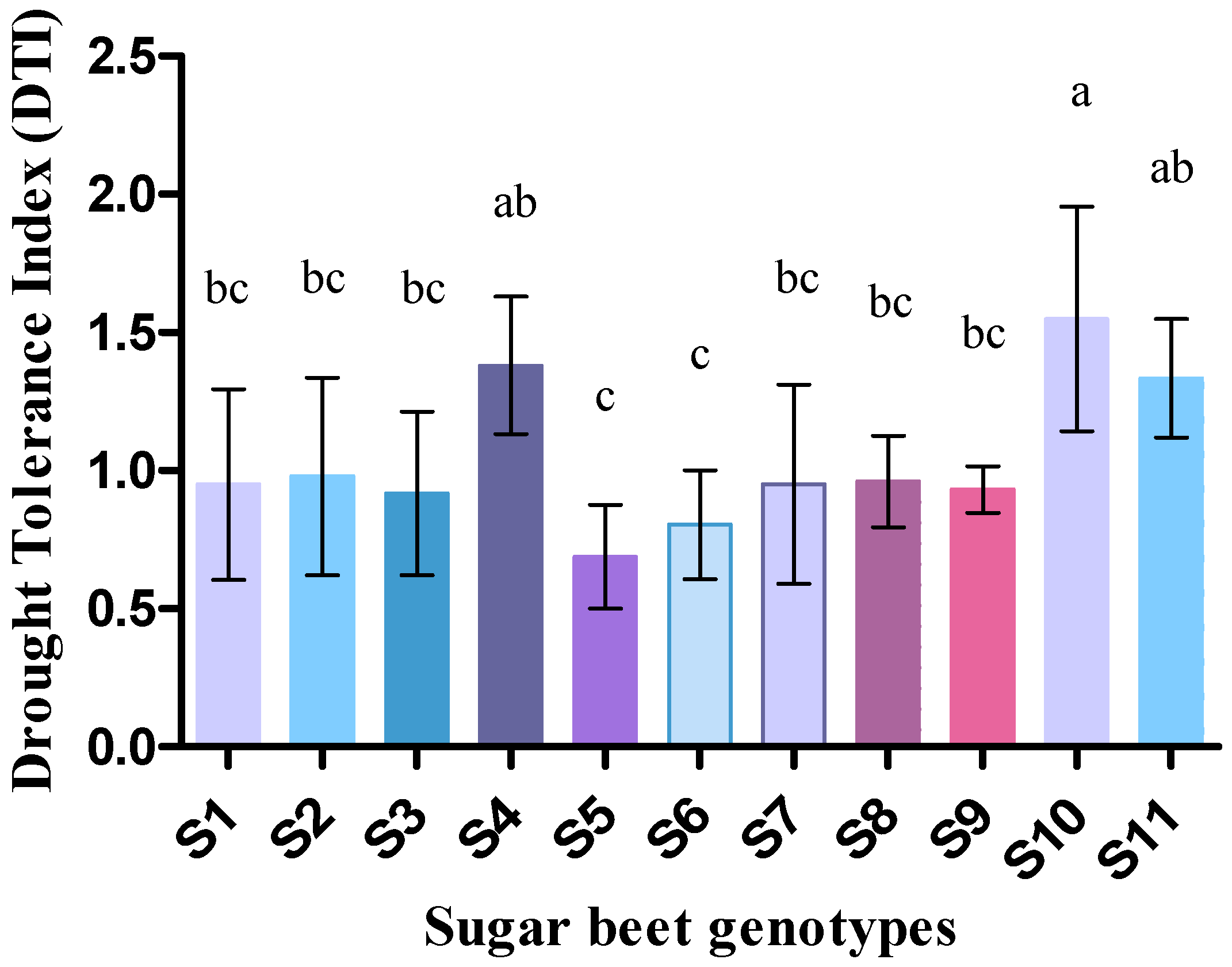{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.N. | Name of the Cultivar | Origin | Code Name |
|---|---|---|---|
| 1 | MAXIMELLA | Germany | S1 |
| 2 | HELENIKA | Germany | S2 |
| 3 | TERRANOVA | Germany | S3 |
| 4 | TOLERANZA | Germany | S4 |
| 5 | BORNITA | Germany | S5 |
| 6 | RECODDINA | Germany | S6 |
| 7 | GREGOIA | Germany | S7 |
| 8 | SV2347 | Belgium | S8 |
| 9 | SV2348 | Belgium | S9 |
| 10 | BSRI Sugarbeet 1 | Bangladesh | S10 |
| 11 | BSRI Sugarbeet 2 | Bangladesh | S11 |
| Cultivars | Plant Height (cm) | Fresh Weight/Plant (gm) | Dry Weight/Plant (gm) | |||
|---|---|---|---|---|---|---|
| Control | Drought | Control | Drought | Control | Drought | |
| S1 | 23.33 ± 1.53 bc | 17.67 ± 0.58 de | 5.09 ± 1.52 abc | 1.72 ± 0.38 gh | 0.28 ± 0.09 bcd | 0.19 ± 0.03 eh |
| S2 | 24.33 ± 1.53 bc | 17.17 ± 0.29 e | 5.94 ± 0.77 a | 1.99 ± 0.43 gh | 0.36 ± 0.04 a | 0.27 ± 0.09 b–e |
| S3 | 23.83 ± 3.82 bc | 18.33 ± 1.15 de | 3.38 ± 1.20 ef | 1.10 ± 0.13 h | 0.20 ± 0.09 d–h | 0.13 ± 0.04 h |
| S4 | 23.33 ± 3.79 bc | 18.43 ± 3.19 de | 4.00 ± 0.50 cde | 2.07 ± 0.41 gh | 0.25 ± 0.02 b–f | 0.23 ± 0.02 c–g |
| S5 | 25.00 ± 1.00 ab | 17.17 ± 2.93 e | 4.72 ± 1.30 bcd | 1.38 ± 0.42 gh | 0.28 ± 0.06 abc | 0.14 ± 0.03 h |
| S6 | 27.67 ± 0.76 a | 21.00 ± 3.04 cd | 4.55 ± 1.51 cde | 1.38 ± 0.30 gh | 0.26 ± 0.08 b–e | 0.15 ± 0.02 gh |
| S7 | 24.67 ± 2.08 ab | 18.17 ± 2.75 de | 4.12 ± 1.12 cde | 1.71 ± 0.29 gh | 0.26 ± 0.08 b–e | 0.18 ± 0.05 fgh |
| S8 | 18.67 ± 0.58 de | 17.00 ± 2.18 e | 4.71 ± 0.19 bcd | 1.79 ± 0.37 gh | 0.27 ± 0.005 b–e | 0.19 ± 0.02 e–h |
| S9 | 18.33 ± 1.53 de | 17.33 ± 2.25 e | 5.88 ± 0.12 ab | 1.85 ± 0.06 gh | 0.33 ± 0.03 ab | 0.23 ± 0.01 c–g |
| S10 | 18.10 ± 0.36 de | 16.67 ± 1.53 e | 4.18 ± 0.19 cde | 2.50 ± 0.29 fg | 0.23 ± 0.01 c–g | 0.23 ± 0.01 c–g |
| S11 | 19.33 ± 2.02 de | 16.00 ± 1.73 e | 3.54 ± 0.53 def | 1.86 ± 0.37 gh | 0.18 ± 0.02 gh | 0.18 ± 0.03 fgh |
| LSD (0.05) | 3.49 | 1.18 | 0.08 | |||
| DTI | Chl a | Chl b | Car | RWC | MSI | MDA | Fv/Fm | Plant ht. | Plant fr. Wt. | Plant dry wt. | H2O2 | SOD | CAT | APX | POD | PPO | Proline | GB | TSC | Sucrose | TPC | TFC | DPPH | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DTI | 1 | |||||||||||||||||||||||
| Chl a | 0.005 | 1 | ||||||||||||||||||||||
| Chl b | −0.036 | 0.988 ** | 1 | |||||||||||||||||||||
| Car | 0.038 | 0.994 ** | 0.967 ** | 1 | ||||||||||||||||||||
| RWC | 0.025 | 0.926 ** | 0.893 ** | 0.932 ** | 1 | |||||||||||||||||||
| MSI | −0.256 | 0.276 | 0.282 | 0.262 | 0.286 | 1 | ||||||||||||||||||
| MDA | −0.133 | −0.758 ** | −0.705 ** | −0.786 ** | −0.728 ** | −0.176 | 1 | |||||||||||||||||
| Fv/Fm | 0.073 | 0.767 ** | 0.739 ** | 0.772 ** | 0.762 ** | 0.065 | −0.463 * | 1 | ||||||||||||||||
| Plant ht. | −0.333 | 0.699 ** | 0.721 ** | 0.669 ** | 0.570 ** | 0.239 | −0.444 * | 0.681 ** | 1 | |||||||||||||||
| Plant fr. Wt. | −0.142 | 0.709 ** | 0.731 ** | 0.676 ** | 0.606 ** | 0.125 | −0.374 | 0.717 ** | 0.927 ** | 1 | ||||||||||||||
| Plant dry wt. | −0.202 | 0.391 | 0.42 | 0.356 | 0.315 | −0.026 | −0.105 | 0.495 * | 0.811 ** | 0.90 ** | 1 | |||||||||||||
| H2O2 | −0.079 | −0.580 ** | −0.565 ** | −0.587 ** | −0.426 * | 0.015 | 0.498 * | −0.365 | −0.465 * | −0.429 * | −0.346 | 1 | ||||||||||||
| SOD | 0.365 | 0.560 ** | 0.514 * | 0.582 ** | 0.595 ** | 0.415 | −0.643 ** | 0.342 | 0.197 | 0.265 | −0.05 | −0.174 | 1 | |||||||||||
| CAT | 0.178 | 0.609 ** | 0.518 * | 0.659 ** | 0.668 ** | 0.257 | −0.693 ** | 0.465 * | 0.194 | 0.151 | −0.134 | −0.343 | 0.640 ** | 1 | ||||||||||
| APX | 0.18 | 0.391 | 0.38 | 0.392 | 0.285 | 0.450 * | −0.414 | 0.204 | 0.253 | 0.219 | −0.024 | −0.141 | 0.648 ** | 0.385 | 1 | |||||||||
| POD | −0.229 | 0.831 ** | 0.835 ** | 0.817 ** | 0.752 ** | 0.388 | −0.584 ** | 0.604 ** | 0.760 ** | 0.713 ** | 0.492 * | −0.356 | 0.332 | 0.36 | 0.406 | 1 | ||||||||
| PPO | −0.197 | −0.596 ** | −0.558 ** | −0.614 ** | −0.461 * | −0.175 | 0.625 | −0.412 | −0.301 | −0.329 | −0.045 | 0.389 | −0.610 ** | −0.531 * | −0.642 ** | −0.429 * | 1 | |||||||
| Proline | −0.054 | −0.867 ** | −0.840 ** | −0.873 ** | −0.744 ** | −0.282 | 0.835 ** | −0.530* | −0.594 ** | −0.590 ** | −0.293 | 0.606 ** | −0.589 ** | −0.561 ** | −0.484 * | −0.750 ** | 0.704 ** | 1 | ||||||
| GB | 0.094 | −0.366 | −0.301 | −0.404 | −0.443 * | −0.513 * | 0.42 | −0.231 | −0.177 | −0.071 | 0.108 | −0.059 | −0.461 * | −0.702 ** | −0.539 ** | −0.473 * | 0.347 | 0.295 | 1 | |||||
| TSC | −0.327 | −0.500 * | −0.480 * | −0.507 * | −0.494 * | −0.440 * | 0.355 | −0.583 ** | −0.266 | −0.275 | −0.023 | 0.205 | −0.590 ** | −0.470 * | −0.428 * | −0.243 | 0.364 | 0.33 | 0.221 | 1 | ||||
| Sucrose | −0.402 | −0.425 * | −0.416 | −0.419 | −0.486 * | −0.268 | 0.278 | −0.535 * | −0.203 | −0.276 | −0.114 | 0.188 | −0.503 * | −0.276 | −0.288 | −0.185 | 0.226 | 0.296 | −0.011 | 0.801 ** | 1 | |||
| TPC | −0.093 | −0.964 ** | −0.941 ** | −0.965 ** | −0.885 ** | −0.235 | 0.729 ** | −0.790 ** | −0.671 ** | −0.692 ** | −0.354 | 0.553 ** | −0.634 ** | −0.616 ** | −0.486 * | −0.769 ** | 0.634 ** | 0.842 ** | 0.375 | 0.533 * | 0.462 * | 1 | ||
| TFC | −0.132 | −0.863 ** | −0.855 ** | −0.853 ** | −0.832 ** | −0.275 | 0.534 * | −0.820 ** | −0.757 ** | −0.636 ** | −0.301 | 0.377 | −0.635 ** | −0.617 ** | −0.460 * | −0.645 ** | 0.501 * | 0.618 ** | 0.391 | 0.660 ** | 0.536 * | 0.903 ** | 1 | |
| DPPH | 0.003 | −0.506 * | −0.478 * | −0.517 * | −0.455 * | 0.274 | 0.403 | −0.391 | −0.245 | −0.275 | −0.088 | 0.291 | −0.221 | −0.436 * | −0.25 | −0.314 | 0.253 | 0.423 * | 0.217 | −0.031 | 0.035 | 0.549 ** | 0.506 * | 1 |
| Genotypes | Proline (μg∙g−1 DW) | Glycine Betaine (mg∙g−1 DW) | TSC (μg∙g−1 DW) | Sucrose (μg∙g−1 DW) | ||||
|---|---|---|---|---|---|---|---|---|
| Control | Drought | Control | Drought | Control | Drought | Control | Drought | |
| S1 | 12.99 ± 0.44 h | 159.45 ± 0.16 c | 7.35 ± 0.05 g | 8.37 ± 0.03 d | 471.33 ± 13.87 ghi | 670.67 ± 14.39 a | 151.17 ± 1.39 e | 205.59 ± 4.48 a |
| S2 | 9.70 ± 0.44 h | 119.01 ± 12.87 d | 8.91 ± 0.06 b | 8.78 ± 0.06 c | 389.44 ± 63.57 jkl | 533.00 ± 2.17 de | 142.94 ± 0.30 f | 174.09 ± 3.12 c |
| S3 | 8.89 ± 0.22 h | 118.36 ± 15.39 d | 5.91 ± 0.06 k | 7.66 ± 0.06 f | 456.22 ± 24.94 hij | 547.22 ± 18.17 cd | 149.72 ± 7.19 e | 172.77 ± 4.07 cd |
| S4 | 18.92 ± 2.35 h | 217.99 ± 15.21 b | 6.78 ± 0.08 i | 9.10 ± 0.1 a | 444.11 ± 40.21 hij | 388.44 ± 38.83 l | 127.51 ± 1.29 g | 96.15 ± 2.79 j |
| S5 | 12.41 ± 2.69 h | 235.43 ± 0.21 a | 8.34 ± 0.07 d | 6.14 ± 0.05 j | 445.00 ± 9.75 h–k | 579.67 ± 11.07 bc | 124.25 ± 1.59 gh | 206.98 ± 5.08 a |
| S6 | 8.20 ± 1.22 h | 102.49 ± 2.30 e | 4.61 ± 0.05 o | 7.78 ± 0.07 e | 435.22 ± 17.25 h–k | 496.78 ± 2.17 fg | 173.37 ± 2.58 c | 154.44 ± 0.40 e |
| S7 | 8.56 ± 0.71 h | 233.46 ± 3.30 a | 3.09 ± 0.04 q | 4.72 ± 0.05 n | 496.78 ± 26.34 efg | 416.11 ± 2.80 kl | 139.02 ± 2.99 f | 167.28 ± 0.41 d |
| S8 | 16.25 ± 0.34 h | 90.73 ± 7.13 e | 5.36 ± 0.05 l | 7.12 ± 0.05 h | 501.67 ± 18.15 ef | 477.33 ± 8.27 fgh | 139.35 ± 0.60 f | 149.74 ± 2.78 e |
| S9 | 12.70 ± 4.62 h | 75.75 ± 12.19 f | 4.31 ± 0.06 p | 6.16 ± 0.04 j | 394.56 ± 10.56 l | 527.89 ± 27.71 de | 140.28 ± 8.35 f | 172.04 ± 3.87 cd |
| S10 | 10.52 ± 0.09 h | 59.07 ± 4.83 g | 5.84 ± 0.06 k | 7.29 ± 0.04 g | 347.22 ± 27.74 m | 585.78 ± 3.89 b | 117.06 ± 2.70 i | 195.93 ± 2.78 b |
| S11 | 7.62 ± 0.61 h | 172.39 ± 22.07 c | 5.24 ± 0.05 m | 7.76 ± 0.06 e | 320.11 ± 10.24 m | 433.89 ± 25.32 ijk | 125.26 ± 8.38 g | 119.04 ± 1.79 hi |
| LSD (0.05) | 13.43 | 0.097 | 34.16 | 6.08 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.J.; Kim, J.W.; Begum, M.K.; Sohel, M.A.T.; Lim, Y.-S. Physiological and Biochemical Changes in Sugar Beet Seedlings to Confer Stress Adaptability under Drought Condition. Plants 2020, 9, 1511. https://doi.org/10.3390/plants9111511
Islam MJ, Kim JW, Begum MK, Sohel MAT, Lim Y-S. Physiological and Biochemical Changes in Sugar Beet Seedlings to Confer Stress Adaptability under Drought Condition. Plants. 2020; 9(11):1511. https://doi.org/10.3390/plants9111511
Chicago/Turabian StyleIslam, Md. Jahirul, Ji Woong Kim, Mst. Kohinoor Begum, Md. Abu Taher Sohel, and Young-Seok Lim. 2020. "Physiological and Biochemical Changes in Sugar Beet Seedlings to Confer Stress Adaptability under Drought Condition" Plants 9, no. 11: 1511. https://doi.org/10.3390/plants9111511
APA StyleIslam, M. J., Kim, J. W., Begum, M. K., Sohel, M. A. T., & Lim, Y.-S. (2020). Physiological and Biochemical Changes in Sugar Beet Seedlings to Confer Stress Adaptability under Drought Condition. Plants, 9(11), 1511. https://doi.org/10.3390/plants9111511