


Genetic Diversity and Growth-Promoting Functions of Endophytic Nitrogen-Fixing Bacteria in Apple

 and
and
Abstract
1. Introduction
2. Results
2.1. Diversity and Community Structure of Endophytic Nitrogen-Fixing Bacteria in Apple Plants
2.1.1. Alpha Diversity of Endophytic Nitrogen-Fixing Bacteria in Apple Plants
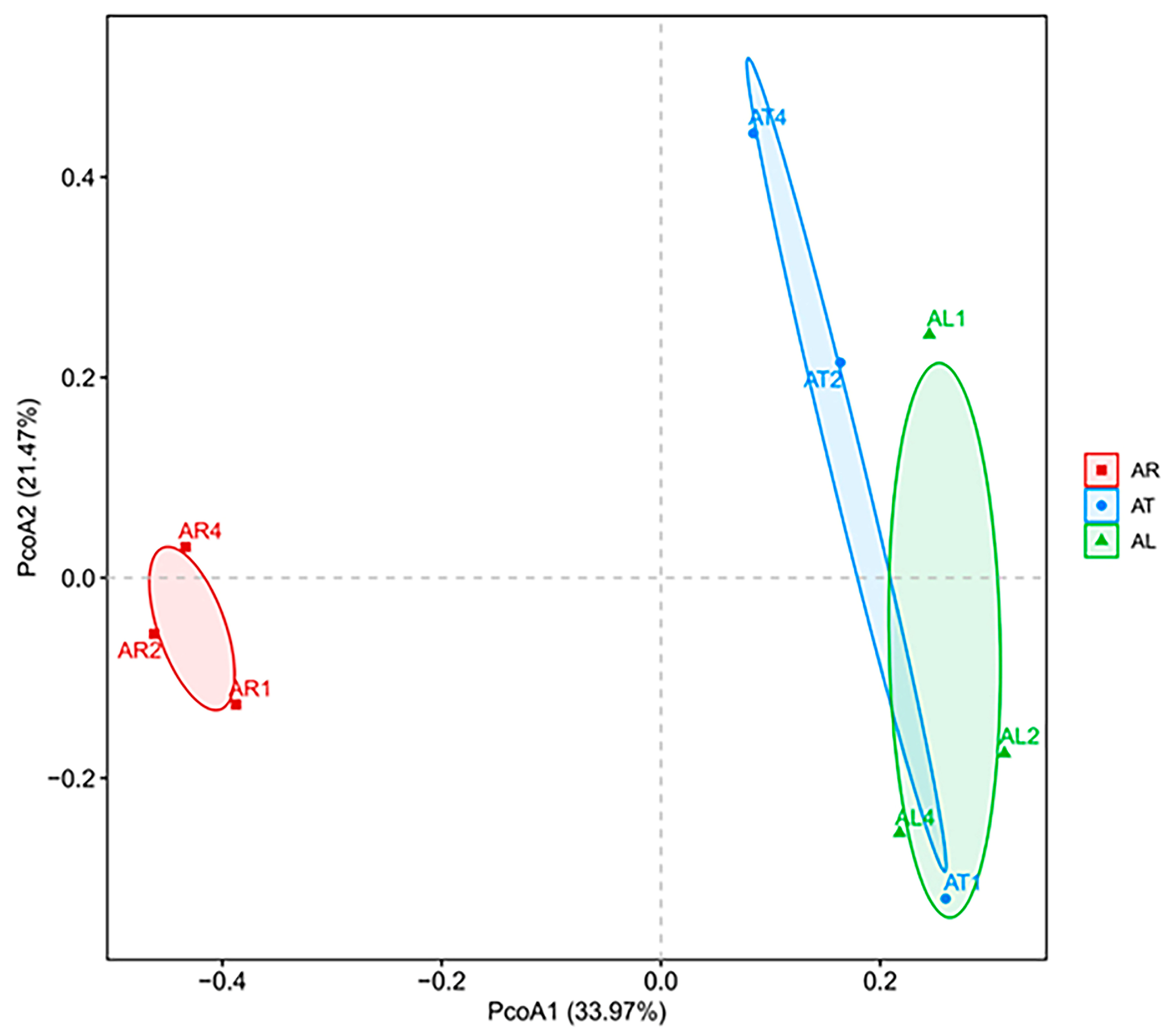
2.1.2. Beta Diversity of Endophytic Nitrogen-Fixing Bacteria in Apple Plants
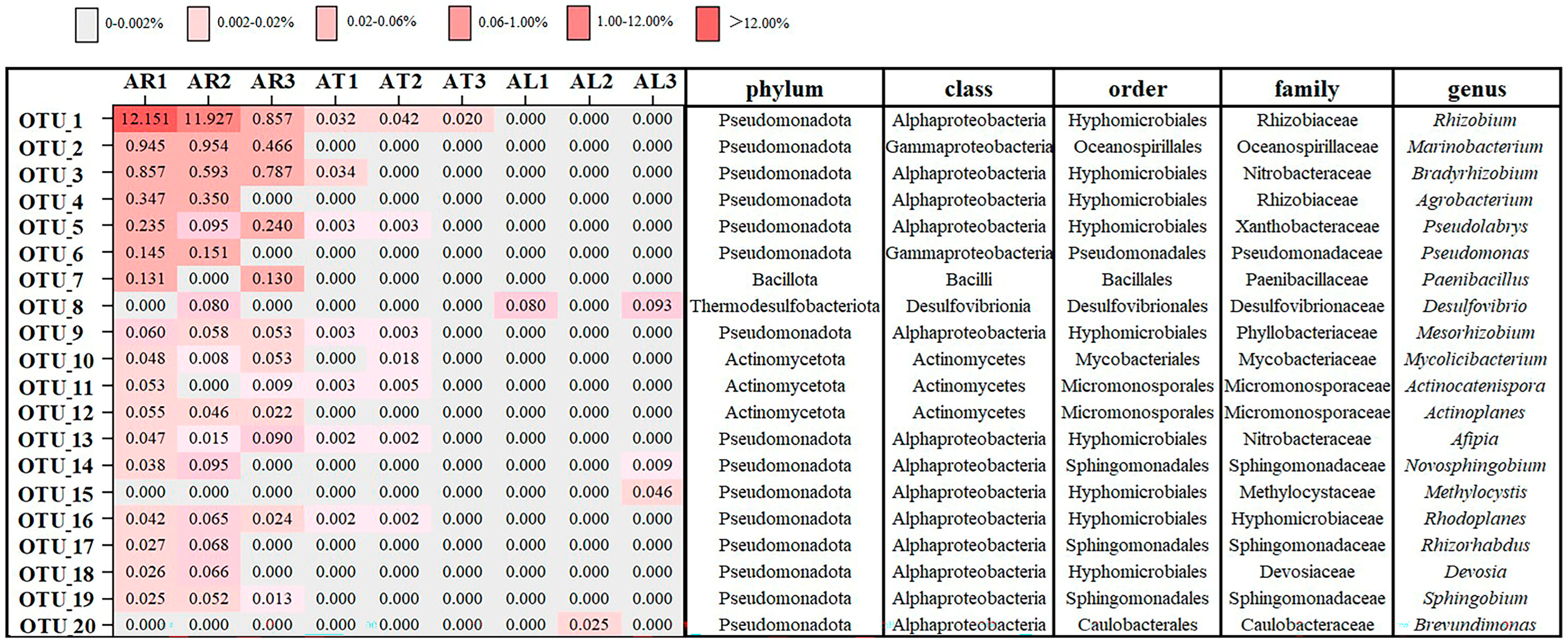
2.1.3. Community Composition of Endophytic Nitrogen-Fixing Bacteria in Apple Plants
2.2. Isolation and Identification of Endophytic Nitrogen-Fixing Bacteria in Apple
2.3. Nitrogenase Activity of Endophytic Nitrogen-Fixing Bacteria
2.4. Assessment of Other Growth-Promoting Potential of Apple Endophytic Nitrogen-Fixing Bacteria
2.5. Growth-Promoting Effects of Apple Endophytic Nitrogen-Fixing Bacteria
3. Discussion
3.1. Diversity of Endophytic Nitrogen-Fixing Bacteria in Apple Plants
3.2. Community Composition of Endophytic Nitrogen-Fixing Bacterial in Apple Plants
3.3. Isolation of Endophytic Nitrogen-Fixing Bacteria from Apple and Its Nitrogenase Activity
3.4. Growth Promoting Characteristics of Endophytic Nitrogen-Fixing Bacteria in Apple
4. Materials and Methods
4.1. Site Description and Sampling
4.2. DNA Extraction, PCR Amplification, and High-Throughput Sequencing
4.3. Isolation and Identification of Endophytic Nitrogen-Fixing Bacteria
4.4. Nitrogenase Activity Assay
4.5. Growth Promotion Assay
4.5.1. Preparation of Tissue Culture Seedlings
4.5.2. Preparation of Bacterial Inoculum
4.6. Assessment of Plant Growth-Promoting Traits of Apple Endophytic Nitrogen-Fixing Bacteria
4.6.1. IAA Production
4.6.2. Phosphorus Solubilization
4.6.3. Siderophore Production
4.6.4. Potassium Solubilization
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- ReferencesWisniewski, M.; Artlip, T.; Liu, J.; Ma, J.; Burchard, E.; Norelli, J.; Dardick, C. Fox Hunting in Wild Apples: Searching for Novel Genes in Malus Sieversii. Int. J. Mol. Sci. 2020, 21, 9516. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xu, X.; Jia, Z.; Hou, X.; Chen, Q.; Sha, J.; Liu, Z.; Zhu, Z.; Jiang, Y.; Ge, S. Nitrification Inhibitor 3,4-Dimethylpyrazole Phosphate Application During the Later Stage of Apple Fruit Expansion Regulates Soil Mineral Nitrogen and Tree Carbon-Nitrogen Nutrition, and Improves Fruit Quality. Front. Plant Sci. 2020, 11, 764. [Google Scholar] [CrossRef]
- Wojciech, K.; Dariusz, W.; Sebastian, P. Effect of Nitrogen Fertilization of Apple Orchard on Soil Mineral Nitrogen Content, Yielding of the Apple Trees and Nutritional Status of Leaves and Fruits. Agriculture 2022, 12, 2169. [Google Scholar] [CrossRef]
- Mosa, W.F.A.; Sas-Paszt, L.; Głuszek, S.; Górnik, K.; Anjum, M.A.; Saleh, A.A.; Abada, H.S.; Awad, R.M. Effect of Some Biostimulants on the Vegetative Growth, Yield, Fruit Quality Attributes and Nutritional Status of Apple. Horticulturae 2022, 9, 32. [Google Scholar] [CrossRef]
- Kilby, S.L. Nitrous Oxide Emissions from Agricultural Soils in Southern Ontario: Influence of Nitrogen Fertilization and Topography. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2006. [Google Scholar]
- Zahra, R.; Jaber, H.S.; Hemmatollah, P.; Saeid, H. Physiological and biochemical traits in coriander affected by plant growth-promoting rhizobacteria under salt stress. Heliyon 2020, 6, e5321. [Google Scholar]
- Tsukanova, K.A.; Chebоtar, V.; Meyer, J.J.M.; Bibikova, T.N. Effect of plant growth-promoting Rhizobacteria on plant hormone homeostasis. S. Afr. J. Bot. 2017, 113, 91–102. [Google Scholar] [CrossRef]
- Reinhold-Hurek, B.; Hurek, T. Living inside plants: Bacterial endophytes. Curr. Opin. Plant Biol. 2011, 14, 435–443. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, J.; Yang, R.; Xu, X.; Lian, D.; Xu, Y.; Shen, H.; Zhang, H.; Xu, J.; Liang, M. Stenotrophomonas sp. SI-NJAU-1 and Its Mutant Strain with Excretion-Ammonium Capability Promote Plant Growth Through Biological Nitrogen Fixation. J. Agric. Food Chem. 2025, 73, 3874–3886. [Google Scholar] [CrossRef]
- Cavalcante, V.A.; Dobereiner, J. A new acid-tolerant nitrogen-fixing bacterium associated with sugarcane. Plant Soil 1988, 108, 23–31. [Google Scholar] [CrossRef]
- Shah, C.; Mali, H.; Mesara, S.; Dhameliya, H.; Subramanian, R.B. Combined inoculation of phosphate solubilizing bacteria with mycorrhizae to alleviate the phosphate deficiency in Banana. Biologia 2022, 77, 2657–2666. [Google Scholar] [CrossRef]
- Silva, L.C.D.; Barbosa, C.K.R.; Junior, K.S.F. Evaluation of the effect of Azospirillum brasilense inoculation on arabic coffee seedlings. Coff. Sci. 2020, 15, 151678. [Google Scholar]
- González-Rodríguez, R.M.; Serrato, R.; Molina, J.; Aragón, C.E.; Olalde, V.; Pulido, L.E.; Dibut, B.; Lorenzo, J.C. Biochemical and physiological changes produced by Azotobacter chroococcum (INIFAT5 strain) on pineapple in vitro-plantlets during acclimatization. Acta Physiol. Plant. 2013, 35, 3483–3487. [Google Scholar] [CrossRef]
- Delaporte-Quintana, P.; Grillo-Puertas, M.; Lovaisa, N.C.; Teixeira, K.R.; Rapisarda, V.A.; Pedraza, R.O. Contribution of Gluconacetobacter diazotrophicus to phosphorus nutrition in strawberry plants. Plant Soil 2017, 419, 335–347. [Google Scholar] [CrossRef]
- El-Serafy, R.S.; El-Sheshtawy, A.A. Effect of nitrogen fixing bacteria and moringa leaf extract on fruit yield, estragole content and total phenols of organic fennel. Sci. Hortic. 2020, 265, 209. [Google Scholar] [CrossRef]
- Munusamy, G.; Jacques, B.; Soon-Wo, K.; Hang-Yeon, W.; Cunthipuram, L. Effects of the inoculation of Burkholderia vietnamensis and related endophytic diazotrophic bacteria on grain yield of rice. Microb. Ecol. 2008, 55, 21–37. [Google Scholar]
- Patel, J.K.; Archana, G. Diverse culturable diazotrophic endophytic bacteria from Poaceae plants show cross-colonization and plant growth promotion in wheat. Plant Soil 2017, 417, 99–116. [Google Scholar] [CrossRef]
- Riggs, P.J.; Chelius, M.K.; Iniguez, A.L.; Kaeppler, S.M.; Triplett, E.W. Enhanced maize productivity by inoculation with diazotrophic bacteria. Funct. Plant Biol. 2001, 28, 829–836. [Google Scholar] [CrossRef]
- Higdon, S.M.; Pozzo, T.; Tibbett, E.J.; Chiu, C.; Jeannotte, R.; Weimer, B.C.; Bennett, A.B. Diazotrophic bacteria from maize exhibit multifaceted plant growth promotion traits in multiple hosts. PLoS ONE 2020, 15, e0239081. [Google Scholar] [CrossRef]
- Pham, V.T.K.; Rediers, H.; Ghequire, M.G.K.; Nguyen, H.H.; De Mot, R.; Vanderleyden, J.; Spaepen, S. The plant growth-promoting effect of the nitrogen-fixing endophyte Pseudomonas stutzeri A15. Arch. Microbiol. 2017, 199, 513–517. [Google Scholar] [CrossRef]
- Wang, H.; Wen, K.; Zhao, X.; Wang, X.; Li, A.; Hong, H. The inhibitory activity of endophytic Bacillus sp. strain CHM1 against plant pathogenic fungi and its plant growth-promoting effect. Crop Prot. 2009, 28, 634–639. [Google Scholar] [CrossRef]
- Rana, K.L.; Kour, D.; Kaur, T.; Negi, R.; Devi, R.; Yadav, N.; Rai, P.K.; Singh, S.; Rai, A.K.; Yadav, A. Endophytic nitrogen-fixing bacteria: Untapped treasurer for agricultural sustainability. J. Appl. Biol. Biotechnol. 2023, 11, 75–93. [Google Scholar] [CrossRef]
- Lima, E.; Boddey, R.M.; Döbereiner, J. Quantification of biological nitrogen fixation associated with sugar cane using a 15N aided nitrogen balance. Soil Biol. Biochem. 1987, 19, 165–170. [Google Scholar] [CrossRef]
- Padda, K.P.; Puri, A.; Chanway, C. Endophytic nitrogen fixation—A possible ‘hidden’ source of nitrogen for lodgepole pine trees growing at unreclaimed gravel mining sites. FEMS Microbiol. Ecol. 2019, 95, 172. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, S.; Wang, G.; Chen, P.; Hu, Z.; Sun, X. Composition and diversity of endophytic diazotrophs within the pioneer plants in a newly formed glacier floodplain on the eastern Tibetan Plateau. Plant Soil 2022, 481, 253–267. [Google Scholar] [CrossRef]
- Compant, S.; Saikkonen, K.; Mitter, B.; Campisano, A.; Mercado-Blanco, J. Editorial special issue: Soil, plants and endophytes. Plant Soil 2016, 405, 1–11. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, M.; Huang, S.; Li, L.; Gao, Q.; Wang, Y.; Zhang, S.; Huang, S.; Yuan, L.; Wen, Y.; et al. A highly conserved core bacterial microbiota with nitrogen-fixation capacity inhabits the xylem sap in maize plants. Nat. Commun. 2022, 1, 3361. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Ahmed, A.; John, N.; Erik, B.; Leonardo, S.; Samir, D.; Michael, W. Apple endophytic microbiota of different rootstock/scion combinations suggests a genotype-specific influence. Microbiome 2018, 6, 18. [Google Scholar]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Karlidag, H.; Esitken, A.; Turan, M.; Sahin, F. Effects of root inoculation of plant growth promoting rhizobacteria (PGPR) on yield, growth and nutrient element contents of leaves of apple. Sci. Hortic. 2007, 114, 16–20. [Google Scholar] [CrossRef]
- Roesch, L.F.W.; Camargo, F.A.O.; Bento, F.M.; Triplett, E.W. Biodiversity of diazotrophic bacteria within the soil, root and stem of field-grown maize. Plant Soil 2008, 302, 91–104. [Google Scholar] [CrossRef]
- Aslantaş, R.; Çakmakçi, R.; Şahin, F. Effect of plant growth promoting rhizobacteria on young apple tree growth and fruit yield under orchard conditions. Sci. Hortic. 2007, 111, 371–377. [Google Scholar] [CrossRef]
- Alquéres, S.M.C.; Oliveira, J.H.M.; Nogueira, E.M.; Guedes, H.V.; Oliveira, P.L.; Câmara, F.; Baldani, J.I.; Martins, O.B. Antioxidant pathways are up-regulated during biological nitrogen fixation to prevent ROS-induced nitrogenase inhibition in Gluconacetobacter diazotrophicus. Arch. Microbiol. 2010, 192, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Doty, S.L.; Dosher, M.R.; Singleton, G.L.; Moore, A.L.; Van Aken, B.; Stettler, R.F.; Strand, S.E.; Gordon, M.P. Identification of an endophytic Rhizobium in stems of Populus. Symbiosis 2005, 39, 27–35. [Google Scholar]
- Wall, J.D.; Rapp-Giles, B.J.; Brown, M.F.; White, J.A. Response of Desulfovibrio desulfuricans colonies to oxygen stress. Can. J. Microbiol. 1990, 36, 70. [Google Scholar] [CrossRef]
- Collavino, M.M.; Cabrera, E.V.R.; Bruno, C.; Aguilar, O.M. Effect of soil chemical fertilization on the diversity and composition of the tomato endophytic diazotrophic community at different stages of growth. Braz. J. Microbiol. 2020, 51, 1965–1975. [Google Scholar] [CrossRef]
- Bahulikar, R.A.; Chaluvadi, S.R.; Torres-Jerez, I.; Mosali, J.; Bennetzen, J.L.; Udvardi, M. Nitrogen Fertilization Reduces Nitrogen Fixation Activity of Diverse Diazotrophs in Switchgrass Roots. Phytobiomes J. 2020, 5, 80–87. [Google Scholar] [CrossRef]
- Qian, T.; Puri, A.; Padda, K.P.; Chanway, C.P. Biological nitrogen fixation and plant growth promotion of lodgepole pine by an endophytic diazotroph Paenibacillus polymyxa and its GFP-tagged derivative. Botany 2017, 95, 611–619. [Google Scholar]
- Liu, X.; Yang, C.; Yu, X.; Yu, H.; Zhuang, W.; Gu, H.; Xu, K.; Zheng, X.; Wang, C.; Xiao, F.; et al. Revealing structure and assembly for rhizophyte-endophyte diazotrophic community in mangrove ecosystem after introduced Sonneratia apetala and Laguncularia racemosa. Sci. Total Environ. 2020, 721, 137807. [Google Scholar] [CrossRef]
- Leardwiriyakool, C.; Paiboolya, G.; Neung, T.; Wasu, P.; Amornrat, C.; Rungroch, S.; Saisamorn, L. Enhancing Teak (Tectona grandis) Seedling Growth by Rhizosphere Microbes: A Sustainable Way to Optimize Agroforestry. Microorganisms 2021, 9, 1990. [Google Scholar] [CrossRef]
- Huang, X.; Zhou, W.; Zhang, Y.; Yang, Q.; Yang, B.; Liang, T.; Ling, J.; Dong, J. Keystone PGPR ecological effect: An inoculation case study of diazotrophic Novosphingobium sp. N034 on mangrove plant Kandelia obovate. Appl. Soil Ecol. 2024, 202, 105567. [Google Scholar] [CrossRef]
- Rangjaroen, C.; Rerkasem, B.; Teaumroong, N.; Noisangiam, R.; Lumyong, S. Promoting plant growth in a commercial rice cultivar by endophytic diazotrophic bacteria isolated from rice landraces. Ann. Microbiol. 2015, 65, 253–266. [Google Scholar] [CrossRef]
- Xu, J.; Kloepper, J.W.; Huang, P.; McInroy, J.A.; Hu, C.H. Isolation and characterization of N2 -fixing bacteria from giant reed and switchgrass for plant growth promotion and nutrient uptake. J. Basic Microbiol. 2018, 58, 459–471. [Google Scholar] [CrossRef]
- James, E.K.; Reis, V.M.; Olivares, F.L.; Baldani, J.I.; Döbereiner, J. Infection of sugar cane by the nitrogen-fixing bacterium Acetobacter diazotrophicus. J. Exp. Bot. 1994, 45, 757–766. [Google Scholar] [CrossRef]
- Rhona; Xiao, H.; Russon; Zhao, H.; Luo, Q. Isolation, identification and diversity analysis of endophytic azotobacter of capsicum. Crop J. 2014, 52–56. [Google Scholar] [CrossRef]
- Ma, J. Isolation, Identification, Growth Characteristics and Tieback Effect of Endophytic Nitrogen-Fixing Bacteria from Vetiver. Master’s Thesis, Fujian Normal University, Fuzhou, China, 2009. [Google Scholar]
- Gao, C.; Huang, S.; Hu, L.; Wang, Z.; Cao, Y. Diversity and plant growth promotion Of endophytic bacteria isolated from Oryza nivara. J. Appl. Environ. Biol. 2018, 24, 33–38. [Google Scholar]
- Cappelletti, M.; Presentato, A.; Piacenza, E.; Firrincieli, A.; Turner, R.J.; Zannoni, D. Biotechnology of Rhodococcus for the production of valuable compounds. Appl. Microbiol. Biot. 2020, 104, 8567–8594. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.D.; Divya, J.; Saurabh, K.; Ravindra, S.; Reeta, G. Differential protein profiling of soil diazotroph Rhodococcus qingshengii S10107 towards low-temperature and nitrogen deficiency. Sci. Rep. 2019, 9, 20378. [Google Scholar]
- Miliute, I.; Buzaite, O.; Gelvonauskiene, D.; Sasnauskas, A.; Stanys, V.; Baniulis, D. Plant growth promoting and antagonistic properties of endophytic bacteria isolated from domestic apple. Zemdirbyste-Agriculture 2016, 103, 77–82. [Google Scholar] [CrossRef]
- Roberto, D.; Anna, A.; Carmen, B. The Overproduction of Indole-3-Acetic Acid (IAA) in Endophytes Upregulates Nitrogen Fixation in Both Bacterial Cultures and Inoculated Rice Plants. Microb. Ecol. 2017, 74, 441–452. [Google Scholar]
- Abderrahim, A.; Adnane, B.; Kaoutar, Y.; Abderraouf, H.; Iman, B.; Youssef, Z.; Issam, M.K. Nitrogen Fixing Azotobacter Species as Potential Soil Biological Enhancers for Crop Nutrition and Yield Stability. Front. Microbiol. 2021, 12, 628379. [Google Scholar]
- Urquiaga, S.; Cruz, K.H.S.; Boddey, R.M. Contribution of Nitrogen Fixation to Sugar Cane: Nitrogen-15 and Nitrogen-Balance Estimates. Soil Sci. Soc. Am. J. 1992, 56, 105–114. [Google Scholar] [CrossRef]
- Li, Y.; Li, Q.; Guan, G.; Chen, S. Phosphate solubilizing bacteria stimulate wheat rhizosphere and endosphere biological nitrogen fixation by improving phosphorus content. PeerJ 2020, 8, e9062. [Google Scholar] [CrossRef]
- Łucja, B.K.; Dariusz, W.; Sebastian, P. The Effect of Microbial Inoculation under Various Nitrogen Regimes on the Uptake of Nutrients by Apple Trees. Agronomy 2021, 11, 2348. [Google Scholar] [CrossRef]
- Nimisha, P.; Yogesh, P.; Archana, M. Bio Fertilizer: A Promising Tool for Sustainable Farming. Int. J. Innov. Res. Sci. Eng. Technol. 2014, 3, 15838–15842. [Google Scholar]
- Pang, Z.; Mao, X.; Zhou, S.; Yu, S.; Liu, G.; Lu, C.; Wan, J.; Hu, L.; Xu, P. Microbiota-mediated nitrogen fixation and microhabitat homeostasis in aerial root-mucilage. Microbiome 2023, 11, 85. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Ehsan, M.; Sin, Y.; Paek, J.; Khalid, N.; Hayat, R.; Chang, Y.H. Sphingobacterium pakistanensis sp. nov., a novel plant growth promoting rhizobacteria isolated from rhizosphere of Vigna mungo. Ant. Van Leeuwenhoek. 2014, 105, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Vaishnav, A.; Singh, J.; Singh, P.; Rajput, R.S.; Singh, H.B.; Sarma, B.K. Sphingobacterium sp. BHU-AV3 Induces Salt Tolerance in Tomato by Enhancing Antioxidant Activities and Energy Metabolism. Front. Microbiol. 2020, 11, 443. [Google Scholar] [CrossRef]
- Oubohssaine, M.; Sbabou, L.; Aurag, J. Potential of the plant growth-promoting rhizobacterium Rhodococcus qingshengii LMR356 in mitigating lead stress impact on Sulla spinosissima L. Environ. Sci. Pollut. Res. Int. 2024, 31, 46002–46022. [Google Scholar] [CrossRef]
- Wen, S.; Cui, N.; Gong, D.; Xing, L.; Wu, Z.; Zhang, Y.; Wang, Z.; Wang, J. Optimizing nitrogen fertilizer application for achieving high yield with low environmental risks in apple orchard. Agric. Water Manag. 2023, 289, 108501. [Google Scholar] [CrossRef]
- Russell, A.E.; Cambardella, C.A.; Laird, D.A.; Jaynes, D.B.; Meek, D.W. Nitrogen fertilizer effects on soil carbon balances in midwestern U.S. agricultural systems. Ecol. Appl. 2009, 19, 1102–1113. [Google Scholar] [CrossRef]
- Poly, F.; Monrozier, L.J.; Bally, R. Improvement in the RFLP procedure for studying the diversity of nifH genes in communities of nitrogen fixers in soil. Res. Microbiol. 2001, 152, 95–103. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2013, 30, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.-Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [PubMed]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 5056. [Google Scholar] [CrossRef] [PubMed]
- Gaby, J.C.; Buckley, D.H. A comprehensive evaluation of PCR primers to amplify the nifH gene of nitrogenase. PLoS ONE 2017, 7, e42149. [Google Scholar] [CrossRef]
- Saiz, E.; Sgouridis, F.; Drijfhout, F.P.; Ullah, S. Biological nitrogen fixation in peatlands: Comparison between acetylene reduction assay and 15 N 2 assimilation methods. Soil Biol. Biochem. 2019, 131, 157–165. [Google Scholar] [CrossRef]
- Lee, K.; Yoshida, T. An assay technique of measurement of nitrogenase activity in root zone of rice for varietal screening by the acetylene reduction method. Plant Soil 1977, 46, 127–134. [Google Scholar] [CrossRef]
- Guo, H.L.; Shi, D.W.; Li, Z.; Liang, J.J.; Liang, B. Summary of apple tissue culture production technology. Hebei Fruits J. 2010, 17–18. [Google Scholar] [CrossRef]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef]
- Li, Y.; Yao, T.; Rong, L. Characteristics of IAA secretion and phosphate dissolving of phosphate-solubilizing bacteria and its effect on alfalfa growth. Acta Agrest. Sin. 2010, 18, 84–88. [Google Scholar]
- Kowalenko, C.G.; Babuin, D. Interference Problems with Phosphoantimonylmolybdenum Colorimetric Measurement of Phosphorus in Soil and Plant Materials. Commun. Soil Sci. Plant Anal. 2007, 38, 1299–1316. [Google Scholar] [CrossRef]
- Machuca, A.; Milagres, A.M.F. Use of CAS-agar plate modified to study the effect of different variables on the siderophore production by Aspergillus. Lett. Appl. Microbiol. 2003, 36, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhao, Z.; Zhao, Z.; Zhang, Y.; Li, M.; Yu, Q. Analysis of the Potassium-Solubilizing Priestia megaterium Strain NK851 and Its Potassium Feldspar-Binding Proteins. Int. J. Mol. Sci. 2023, 24, 14226. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Shannon Index Mean ± SE | Simpson Index | Chao Index | Ace Index | Goods Coverage |
|---|---|---|---|---|---|
| Root | 4.98 ± 0.49 A | 0.95 ± 0.02 A | 1582.93 ± 278.21 a | 1585.42 a | 1.00 |
| Twig | 3.29 ± 0.4 AB | 0.90 ± 0.04 AB | 508.01 ± 417.94 ab | 756.23 ± 775.31 ab | 0.98 |
| Leaf | 2.25 ± 0.85 B | 0.71 ± 0.26 B | 100.6 ± 24.23 b | 100.96 ± 23.98 b | 1.00 |
| Isolate | Class | Closest Match (Biocloud) | Accession Number | Query Coverage (%) | Tissue | |
|---|---|---|---|---|---|---|
| 1 | RA-07 | Pseudomonadota | Agrobacterium deltaense | YTC 4121 | 99.68 | R, L |
| 2 | RC-15 | Pseudomonadota | A. pusense | LMG 25623 | 99.85 | R, L |
| 3 | RC-14 | Pseudomonadota | A. radiobacter | ATCC 19358 | 99.93 | R, L |
| 4 | RC-09 | Pseudomonadota | Rhizobium-AKKA_s | CF122 | 99.13 | R |
| 5 | RB-08 | Pseudomonadota | Ancylobacter defluvii | SK15 | 99.61 | R |
| 6 | LD-05 | Pseudomonadota | Brucella anthropi | ATCC 49188 | 99.78 | L |
| 7 | LA-01 | Pseudomonadota | B. ciceri | Ca-34 | 99.65 | L |
| 8 | ZD-02 | Pseudomonadota | Methylobacterium brachiatum | B0021 | 99.64 | Z |
| 9 | ZC-08 | Pseudomonadota | M. goesingense | iEII3 | 99.18 | Z |
| 10 | ZC-10 | Pseudomonadota | M. komagatae | 002-079 | 96.86 | Z |
| 11 | RD-01 | Pseudomonadota | Novosphingobium barchaimii | LL02 | 99.06 | R |
| 12 | RC-16 | Pseudomonadota | N. resinovorum | NCIMB 8767 | 99.88 | R, Z |
| 13 | ZC-04 | Pseudomonadota | N.-SODQ_s | PhB55 | 99.13 | R, Z |
| 14 | LD-10 | Pseudomonadota | Pseudomonas coleopterorum | Esc2Am | 99.65 | L |
| 15 | LB-03 | Pseudomonadota | P. graminis | DSM 11363 | 98.81 | L |
| 16 | LD-07 | Pseudomonadota | P. putida | EU348741.1 | 99.02 | L |
| 17 | RB-02 | Pseudomonadota | Pseudorhodoplanes sinuspersici | RIPI 110 | 99.49 | R |
| 18 | RC-04 | Pseudomonadota | Sphingobium aquiterrae | SKLS-A10 | 97.17 | R |
| 19 | RA-01 | Pseudomonadota | S. scionense | WP01 | 99.7 | R |
| 20 | LD-09 | Pseudomonadota | S. yanoikuyae | ATCC 51230 | 99.7 | L |
| 21 | ZC-03 | Pseudomonadota | Sphingomonas taxi | ATCC 55669 | 99.12 | Z |
| 22 | LD-06 | Pseudomonadota | S. yabuuchiae | GTC 868 | 100 | L |
| 23 | ZC-11 | Pseudomonadota | S. yunnanensis | YIM 003 | 99.03 | Z |
| 24 | RA-02 | Pseudomonadota | Ancylobacter novellus | DSM 506 | 99.33 | R |
| 25 | ZC-01 | Bacillota | Bacillus velezensis | CR-502 | 99.14 | Z |
| 26 | ZC-12 | Bacillota | B. zanthoxyli | 1433 | 99.65 | Z |
| 27 | ZD-05 | Deinococcota | Deinococcus depolymerans | TDMA-24 | 98.91 | Z |
| 28 | LC-01 | Bacteroidota | Sphingobacterium multivorum | IAM 14316 | 99.67 | L |
| 29 | LD-02 | Actinomycetota | Curtobacterium flaccumfaciens | LMG 3645 | 99.88 | L |
| 30 | LD-11 | Actinomycetota | Microbacterium aurum | KACC 15219 | 98.25 | L |
| 31 | LB-01 | Actinomycetota | Nocardioides phosphati | WYH11-7 | 97.52 | L |
| 32 | ZC-06 | Actinomycetota | Rhodococcus qingshengii | JCM 15477 | 99.72 | Z |
| Strain | Nitrogenase Activity (nmol/h·mL) | Strain | Nitrogenase Activity (nmol/h·mL) | Strain | Nitrogenase Activity (nmol/h·mL) |
|---|---|---|---|---|---|
| RD-01 | 425.3 ± 13.69 a | ZC-06 | 982.28 ± 11.52 a | LC-01 | 543.95 ± 27.53 a |
| RC-15 | 399.91 ± 10.48 a | ZC-11 | 951.91 ± 20.30 a | LD-10 | 513.43 ± 8.52 a |
| RA-07 | 228.03 ± 11.2 b | ZD-04 | 524.9 ± 6.32 b | LD-09 | 512.82 ± 50.36 ab |
| RC-14 | 205.66 ± 5.31 b | ZD-02 | 318.61 ± 2.51 c | LB-03 | 473.63 ± 5.21 b |
| RA-01 | 166.26 ± 4.32 bc | ZC-08 | 283.45 ± 6.21 cd | LD-02 | 428.23 ± 13.69 c |
| RB-08 | 123.77 ± 6.02 c | ZC-12 | 227.05 ± 4.28 d | LB-01 | 420.66 ± 7.30 c |
| RC-09 | 121.1 ± 3.10 c | ZC-10 | 189.7 ± 1.52 de | LA-01 | 412.6 ± 6.33 c |
| RB-02 | 74.95 ± 4.10 d | ZD-05 | 115.36 ± 4.52 e | LD-11 | 338.19 ± 25.41 d |
| RA-01 | 48.58 ± 1.94 de | ZC-03 | 104.36 ± 2.64 e | LD-07 | 308.35 ± 5.74 d |
| RC-04 | 47.12 ± 1.22 e | ZC-01 | 74.95 ± 4.25 ef | LD-05 | 44.68 ± 5.9 f |
| RC-16 | 27.1 ± 0.15 f | LD-06 | 26.86 ± 4.25 f |
| Treatment | Height (cm) | Leaf Number | Chlorophyll Content (mg/g) | Root Length (cm) |
|---|---|---|---|---|
| RD-01 | 4.4 ± 0.2 b | 7 ± 3.46 a | 16.43 ± 0.27 b | 2.9 ± 0.17 d |
| RC-16 | 3.3 ± 0.1 c | 3.33 ± 0.58 c | 10.21 ± 0.22 cd | 2.2 ± 0.1 d |
| RD-01 + RC-16 | 5.3 ± 0.26 a | 6.33 ± 1.15 ab | 21.34 ± 0.43 a | 2.1 ± 0.17 d |
| ZC-11 | 1.73 ± 0.06 e | 5 ± 1 bc | 6.03 ± 0.02 e | 7.5 ± 0.46 a |
| ZC-06 | 1.93 ± 0.23 e | 3.33 ± 0.58 c | 3.59 ± 0.18 f | 5.8 ± 0.35 b |
| ZC-11 + ZC-06 | 2.07 ± 0.49 de | 6.33 ± 0.58 ab | 9.03 ± 0.16 d | 5 ± 0.17 b |
| LC-01 | 3.07 ± 0.76 c | 4 ± 0 bc | 11.24 ± 0.35 c | 4 ± 0.46 c |
| LD-06 | 2.8 ± 0.3 cd | 5.33 ± 0.58 abc | 14.75 ± 0.65 b | 2.13 ± 0.15 d |
| LC-01 + LD-06 | 2.07 ± 0.25 de | 6 ± 0 ab | 20.01 ± 1.76 a | 5.4 ± 0.35 b |
| CK | 2.17 ± 0.83 de | 3.33 ± 0.58 c | 5.13 ± 2.65 ef | 2.23 ± 1.25 d |
| Ingredients (g/L) | Jensen | A4 | Ash | Jnfb |
|---|---|---|---|---|
| C12H22O11 | 20.0 | 20.0 | - | - |
| K2HPO43H2O | 1.31 | - | - | 0.785 |
| NaCl | 0.5 | - | 0.2 | 0.1 |
| CaCO3 | 2.0 | 0.1 | 5.0 | - |
| FeSO4·7H2O | 0.18 | - | - | - |
| MgSO4·7H2O | 1.0 | - | - | - |
| Na2MnO4 | 0.005 | - | - | - |
| MgSO47H2O | - | 0.5 | 0.2 | 0.2 |
| Na2HPO412H2O | - | 5.0 | - | - |
| FeCl3 | - | 0.005 | - | - |
| Mannitol | - | - | 10.0 | - |
| KH2PO4 | - | - | 0.2 | - |
| CaSO42H2O | - | - | 0.1 | - |
| C4H6O5 | - | - | - | 5.0 |
| CaCl2 | - | - | - | 0.02 |
| Fe3-EDTA | - | - | - | 0.066 |
| Biotin | - | - | - | 1.0 |
| VB6 | - | - | - | 0.1 |
| KOH | - | - | - | 4.5 |
| Na2MoO42H2O | - | - | - | 0.2 |
| MnSO4H2O | - | - | - | 0.235 |
| H3BO3 | - | - | - | 0.28 |
| CuSO45H2O | - | - | - | 0.008 |
| ZnSO47H2O | - | - | - | 0.024 |
| Agar | 20.0 | 20.0 | 20.0 | 20.0 |
| Source Tissue | High Nitrogenase Activity Strain | Plant Growth-Promoting Strain | Consortium |
|---|---|---|---|
| Root | RD-01 | RC-16 | RD01 + RC16 |
| Twig | ZC-06 | ZC-11 | ZC11 + ZC06 |
| Leaf | LC-01 | LD-06 | LC01 + LD06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Cheng, H.; Xu, S.; Zhang, D.; Wu, J.; Li, Z.; Fu, B.; Liu, L. Genetic Diversity and Growth-Promoting Functions of Endophytic Nitrogen-Fixing Bacteria in Apple. Plants 2025, 14, 1235. https://doi.org/10.3390/plants14081235
Liu H, Cheng H, Xu S, Zhang D, Wu J, Li Z, Fu B, Liu L. Genetic Diversity and Growth-Promoting Functions of Endophytic Nitrogen-Fixing Bacteria in Apple. Plants. 2025; 14(8):1235. https://doi.org/10.3390/plants14081235
Chicago/Turabian StyleLiu, Hongshan, Huan Cheng, Suwen Xu, Donghua Zhang, Jianrong Wu, Zongyan Li, Benzhong Fu, and Li Liu. 2025. "Genetic Diversity and Growth-Promoting Functions of Endophytic Nitrogen-Fixing Bacteria in Apple" Plants 14, no. 8: 1235. https://doi.org/10.3390/plants14081235
APA StyleLiu, H., Cheng, H., Xu, S., Zhang, D., Wu, J., Li, Z., Fu, B., & Liu, L. (2025). Genetic Diversity and Growth-Promoting Functions of Endophytic Nitrogen-Fixing Bacteria in Apple. Plants, 14(8), 1235. https://doi.org/10.3390/plants14081235