
Metabolome and Transcriptome Analysis Reveals the Regulatory Effect of Magnesium Treatment on EGCG Biosynthesis in Tea Shoots (Camellia sinensis)


 ,
,
Abstract
1. Introduction
2. Results
2.1. Evaluation of Main Biochemical Components in Tea Shoots Treated by Magnesium
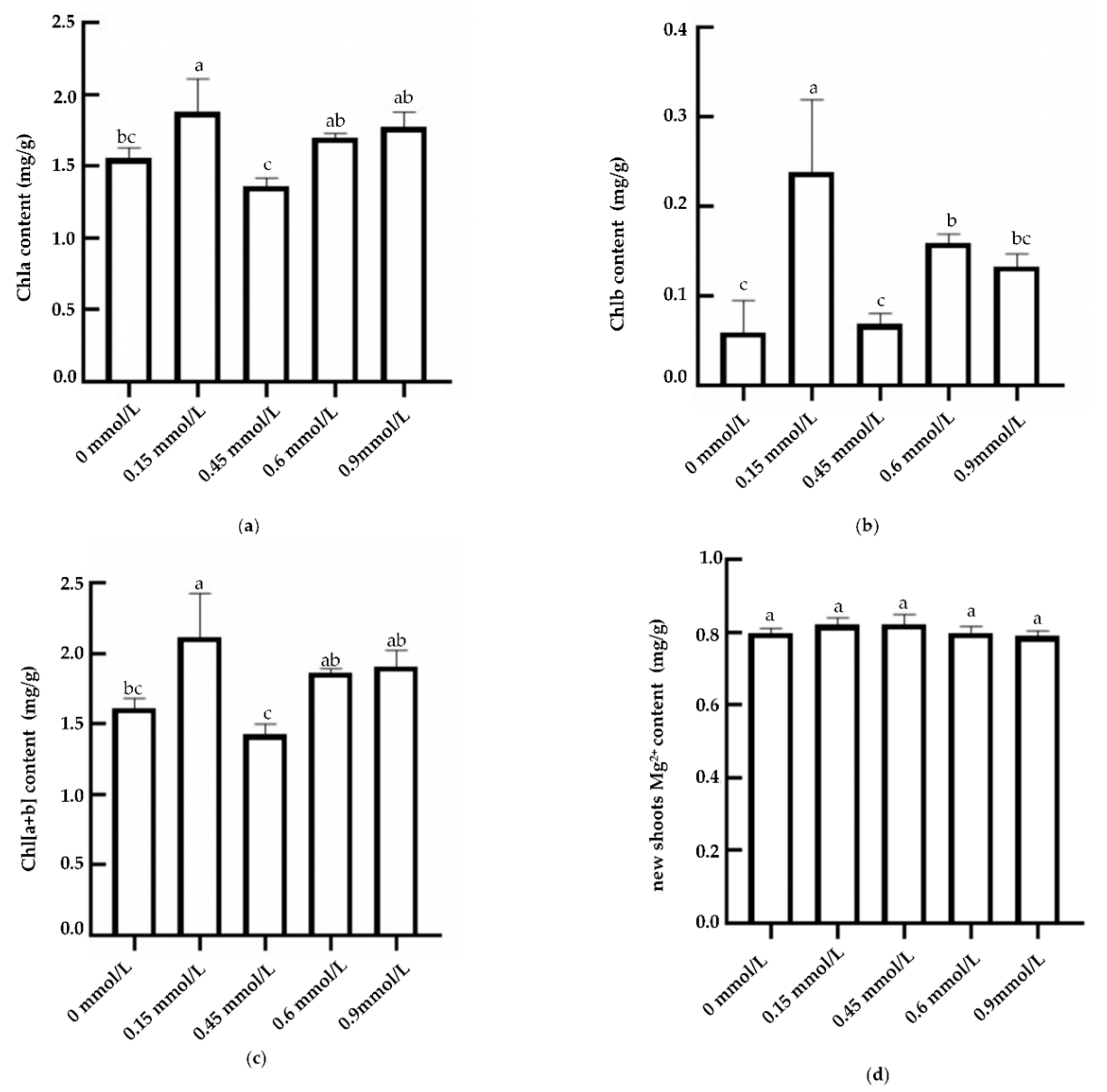
2.2. Evaluation of Magnesium and Chlorophyll Contents in Tea Shoots Treated by Magnesium
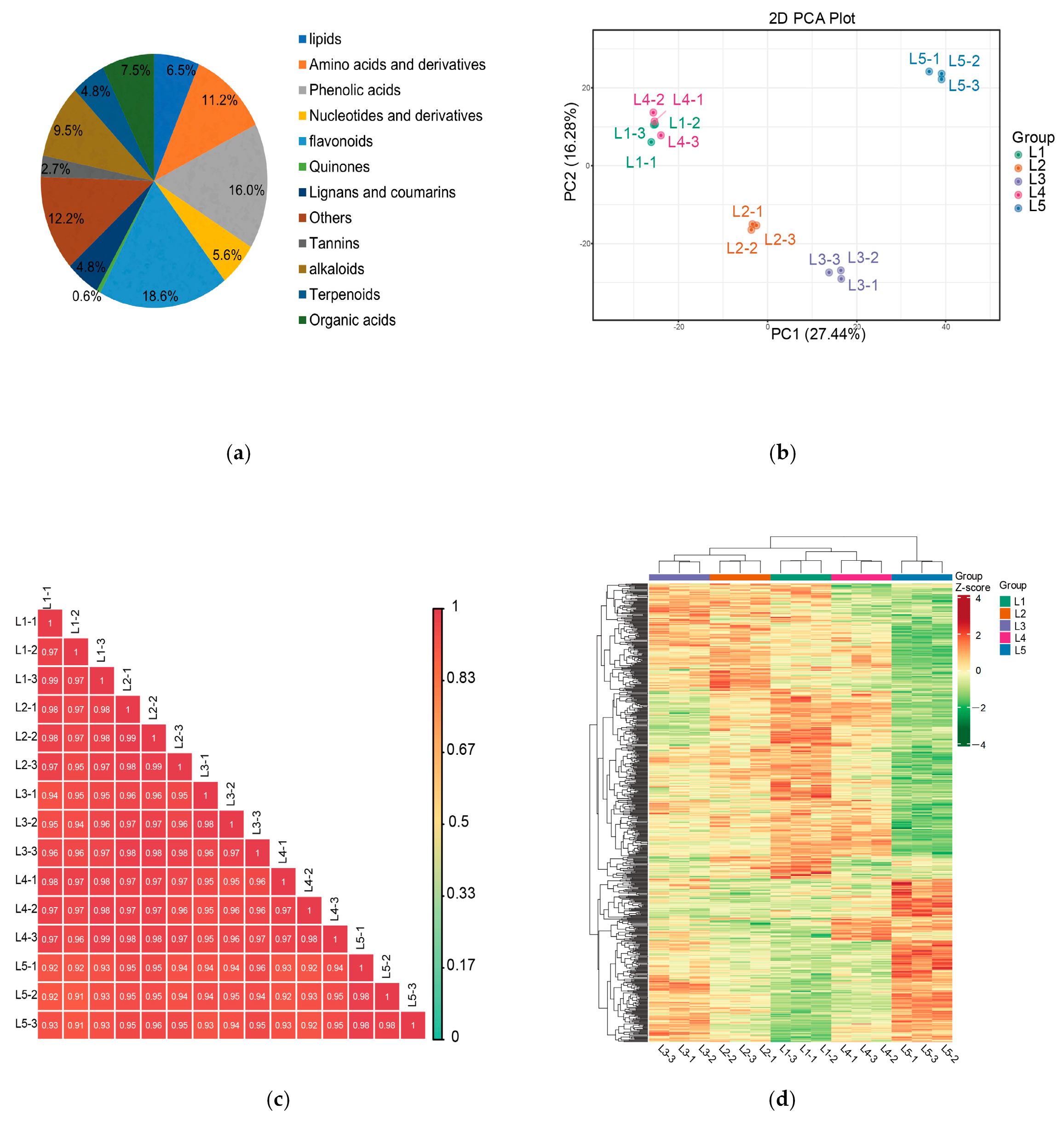
2.3. Analysis of Widely Target Metabolomic in Tea Shoots Treated by Magnesium
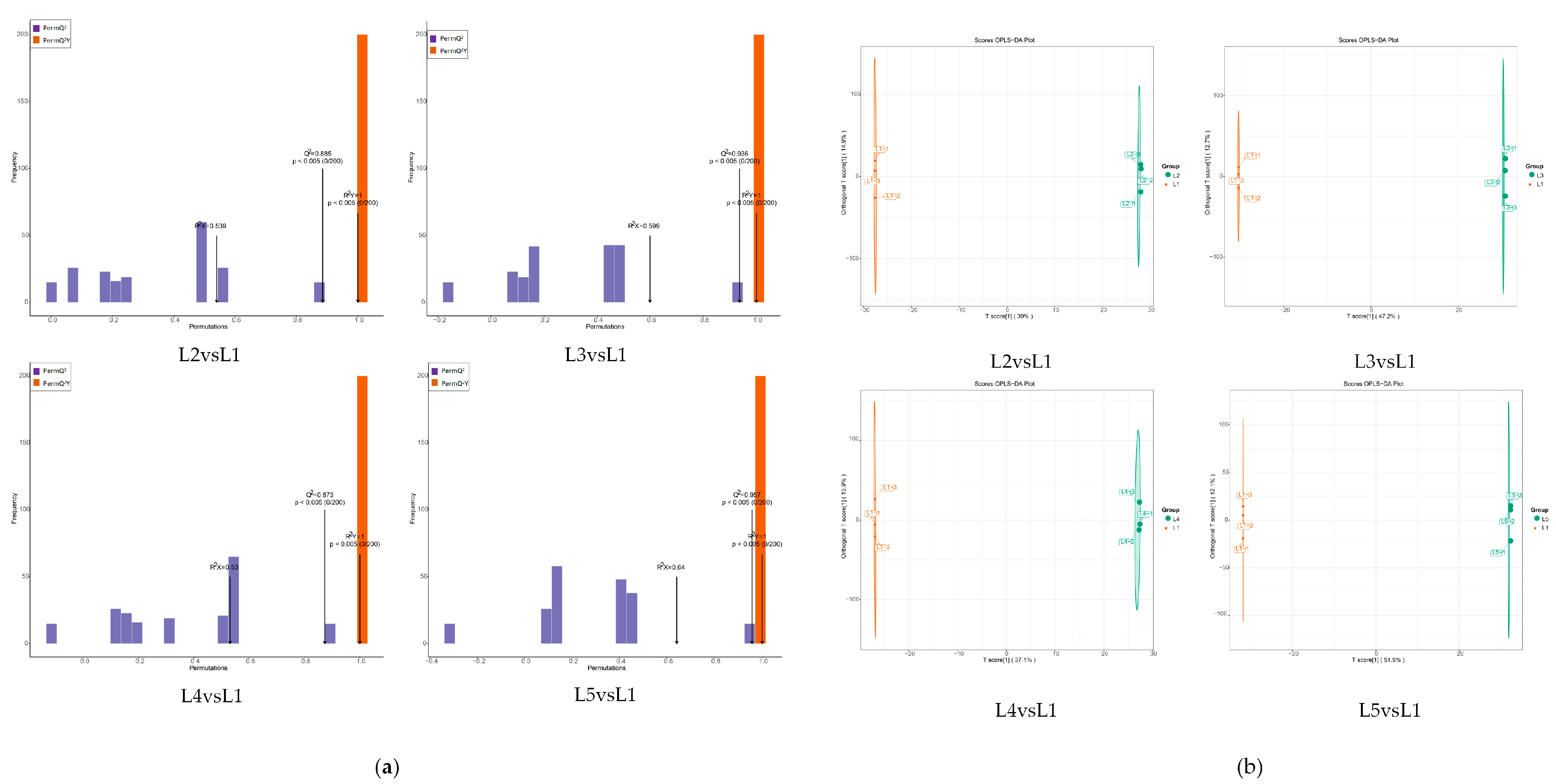
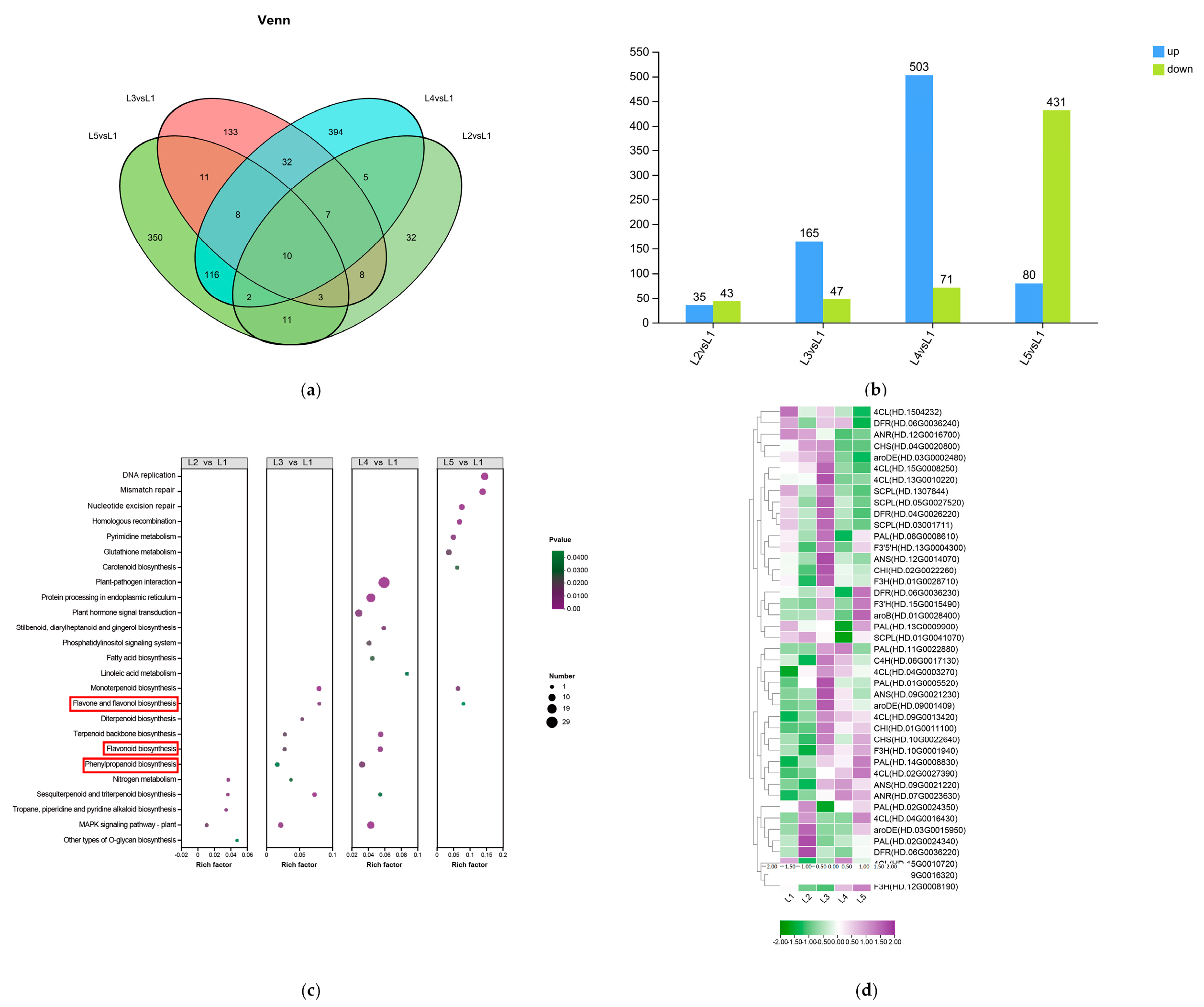
2.4. Analysis of Non-Volatile Metabolites from Differential Accumulation in Tea Shoots Treated by Magnesium
2.5. Identification and Analysis of Key Differential Metabolites in Tea Shoots Treated by Magnesium
2.6. Transcriptomic Analysis in Tea Shoots Treated by Magnesium
2.7. Correlation Analysis of Related Metabolites and Their Synthesis-Related Gene Expressions in Tea Shoots Treated by Magnesium
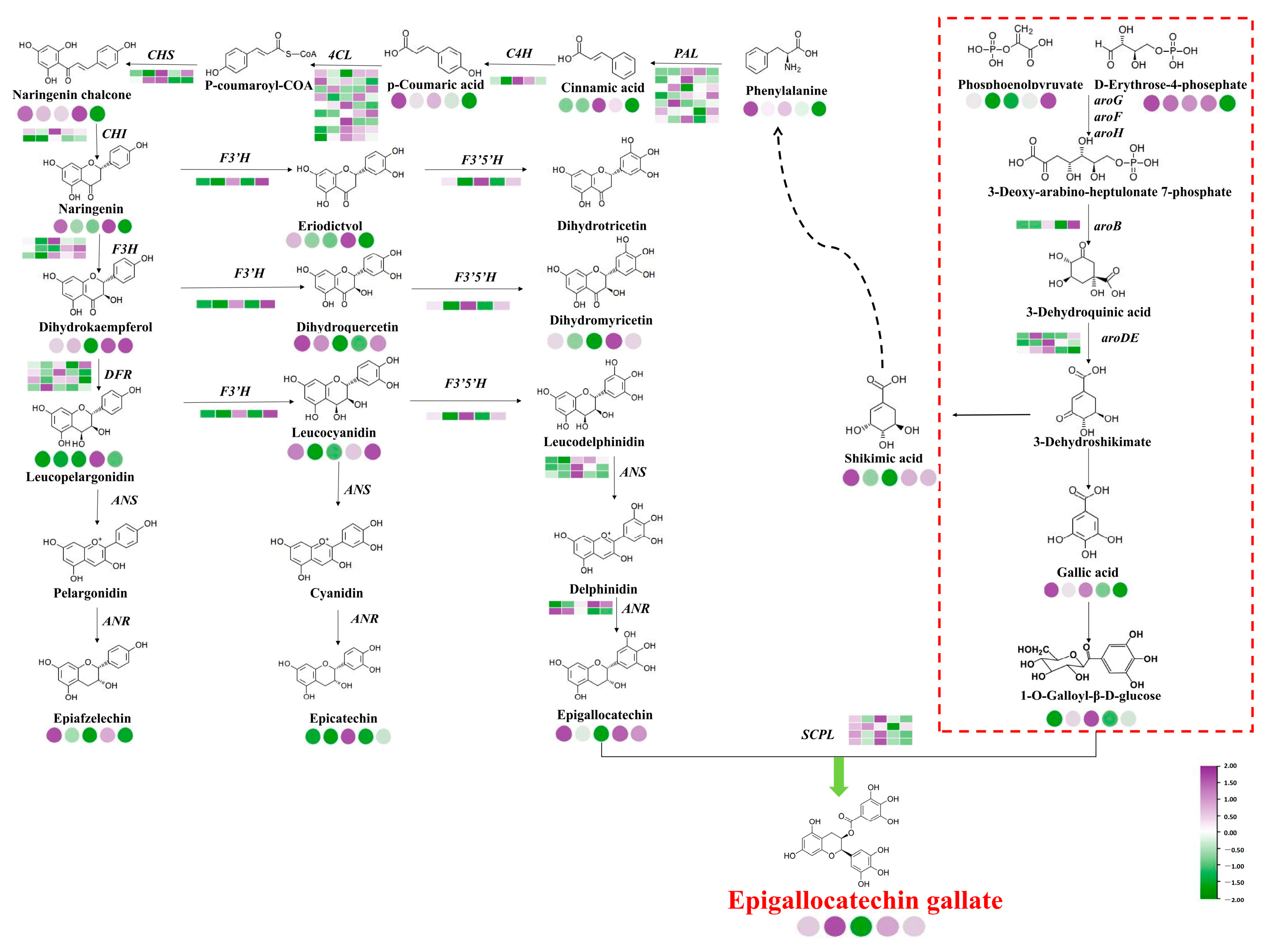
2.8. Pathways of EGCG Biosynthesis in Tea Shoots Treated by Magnesium
2.9. Identification of Transcription Factors in Response to Magnesium Treatment
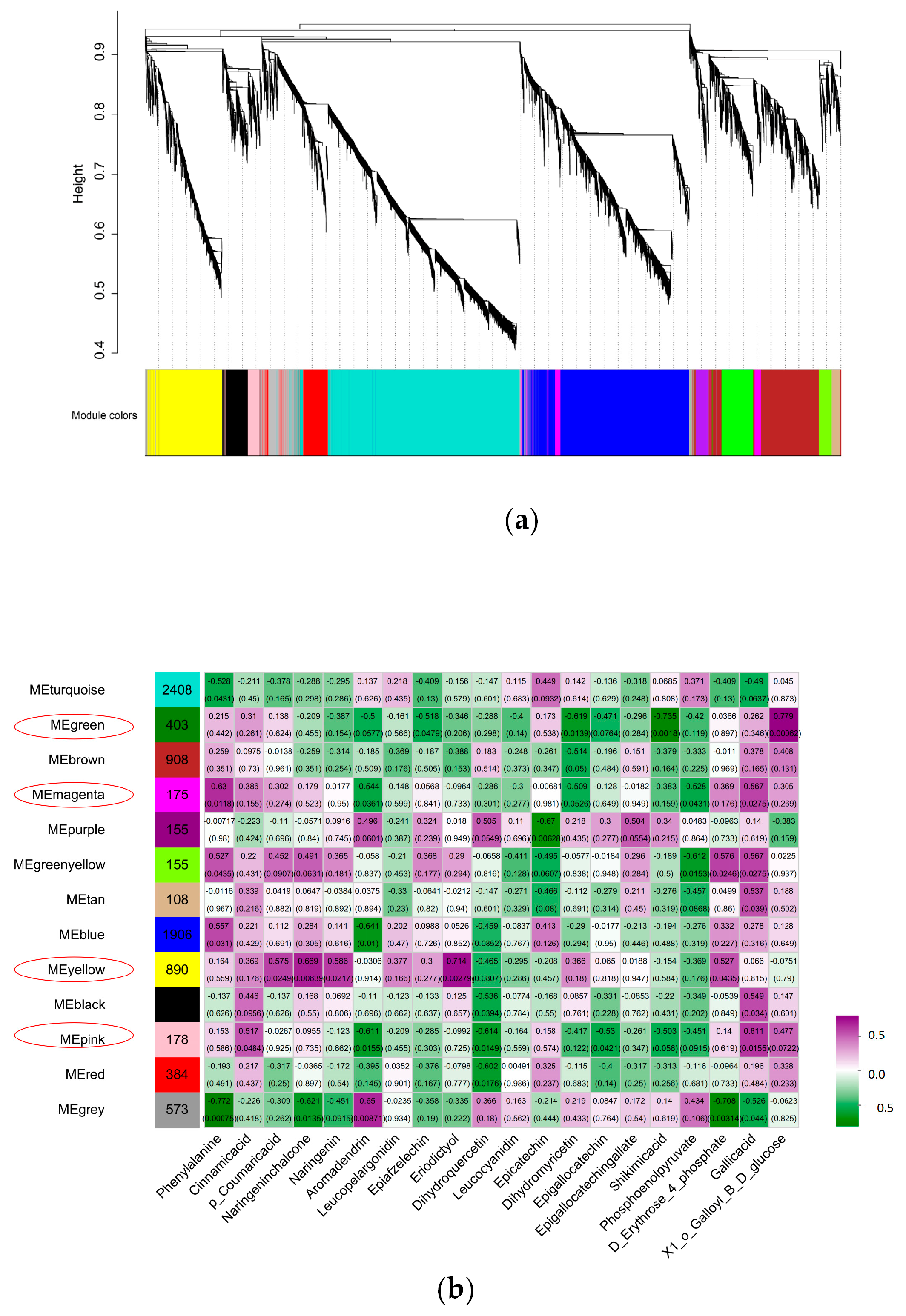
2.10. WGCNA and Hub Gene Identification
2.11. Verification of the Expression Pattern of DEGs
3. Discussion
3.1. Magnesium Treatment Affected the Characteristics, Metabolic Components, and Quality of Tea Plants
3.2. Magnesium Treatment Regulates Metabolites in EGCG Biosynthesis
3.3. Magnesium Treatment Regulates the Expression of Key Genes in EGCG Biosynthesis
3.4. Transcription Factors Play an Important Role in EGCG Biosynthesis
4. Materials and Methods
4.1. Tea Plant Materials
4.2. Determination of the Main Biochemical Components
4.3. Analysis of Non-Volatile Metabolites by Widely Targeted Metabolomics
4.4. Transcriptomics Analysis
4.5. Redundancy Analysis
4.6. Co-Expression Analysis
4.7. Validation of Quantitative Real-Time PCR
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lei, S.; Li, Y. Study on health efficacy and value enhancement strategy of six kinds of tea. Food Saf. Guide 2025, 142–144. [Google Scholar] [CrossRef]
- Daglia, M.; Antiochia, R.; Sobolev, A.P.; Mannina, L. Untargeted and targeted methodologies in the study of tea (Camellia sinensis L.). Food Res. Int. 2014, 63, 275–289. [Google Scholar] [CrossRef]
- Wan, X. Chemical composition and properties of tea. In Tea Biochemistry, 3rd ed.; Wan, X., Huang, J., Zhang, Z., Shen, S., Zhou, C., Shao, W., Huang, J., Liu, Q., Eds.; China Agriculture Press: Beijing, China, 2003; pp. 9–21. [Google Scholar]
- Del Seppia, C.; Federighi, G.; Lapi, D.; Gerosolimo, F.; Scuri, R. Effects of a catechins-enriched diet associated with moderate physical exercise in the prevention of hypertension in spontaneously hypertensive rats. Sci. Rep. 2022, 12, 17303. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Dou, Q.P.; Li, B.; Wang, Y.; Wang, X.; Guan, H.; Li, D.; Li, F. MicroRNAs as novel molecular targets of green tea polyphenol epigallocatechin-3-gallate (EGCG): Relevance and importance to nutrition sciences and cancer prevention. Food Sci. Hum. Well. 2024, 1–27. [Google Scholar] [CrossRef]
- Lu, X. Study on Synergistic Regulation of EGCG Biosynthesis by Endogenous Ethylene and Chrysolactone in Tea Plant. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2024. [Google Scholar]
- Liu, L. Study on Regulation of EGCG Biosynthesis in Tea Plant by Polybulobutenazole and Lovastatin Mediated by Endogenous Hormones. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2024. [Google Scholar]
- Xie, D.; Tong, C.; Chen, W.; Hua, B.; Li, G.; Zhu, J.; Huang, K.; Wei, Z.; Liu, S. Effects of selenium on the growth, quality and nutrients of tea plants. J. Tea Sci. 2023, 64, 1–9. [Google Scholar] [CrossRef]
- Malyukova, L.S.; Koninskaya, N.G.; Orlov, Y.L.; Samarina, L.S. Effects of exogenous calcium on the drought response of the tea plant (Camellia sinensis (L.) Kuntze). PeerJ 2022, 10, e13997. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Q.-H.; Zhang, X.-Y.; Zhang, L.-Y.; Zhao, P.-L.; Wen, T.; Zhang, J.-Q.; Xu, W.-L.; Guo, F.; Zhao, H.; et al. Exploring the Effects of Magnesium Deficiency on the Quality Constituents of Hydroponic-Cultivated Tea (Camellia sinensis L.) Leaves. J. Agr. Food Chem. 2021, 69, 14278–14286. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Liu, X.; Ye, Z.; Ye, Z. Research progress on the response of tea plants to soil pH and nutrients. South China Agric. 2024, 18, 22–26. [Google Scholar] [CrossRef]
- Liu, X. Mechanism by Which Magnesium Fertilizer Regulates Citrus Peel Coloration. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2024. [Google Scholar]
- Masao, I. The Transcription Factor Gene RDD3 Improves Ammonium and Magnesium Uptake and Accumulation and Drought Tolerance in Rice. Plant Mol. Biol. Rep. 2023, 42, 297–307. [Google Scholar]
- Xiong, B.; Li, Q.; Yao, J.; Liu, Z.; Yang, X.; Yu, X.; Li, Y.; Liao, L.; Wang, X.; Deng, H.; et al. Widely targeted metabolomic profiling combined with transcriptome analysis sheds light on flavonoid biosynthesis in sweet orange ‘Newhall’ (C. sinensis) under magnesium stress. Front. Plant Sci. 2023, 14, 1182284. [Google Scholar] [CrossRef] [PubMed]
- Jin, X. Physiological Characteristics of Mulberry in Response to Magnesium Stress and Identification of MmNIA Gene Function. Master’s Thesis, Jiangsu University of Science and Technology, Zhenjiang, China, 2023. [Google Scholar]
- Guo, L. Effects of Different Levels of Potassium and Magnesium on the Contents of Low Terpenoids and Catechins in Tea. Master’s Thesis, Anhui Agricultural University, Hefei, China, 2014. [Google Scholar]
- Liu, X.; Hu, C.; Liu, X.; Riaz, M.; Liu, Y.; Dong, Z.; Tan, Q.; Sun, X.; Wu, S.; Tan, Z. Effect of magnesium application on the fruit coloration and sugar accumulation of navel orange (Citrus sinensis Osb.). Sci. Hortic. 2022, 304, 111282. [Google Scholar] [CrossRef]
- Scott, E.R.; Li, X.; Kfoury, N.; Morimoto, J.; Han, W.-Y.; Ahmed, S.; Cash, S.B.; Griffin, T.S.; Stepp, J.R.; Robbat, A.; et al. Interactive effects of drought severity and simulated herbivory on tea (Camellia sinensis) volatile and non-volatile metabolites. Environ. Exp. Bot. 2019, 157, 283–292. [Google Scholar] [CrossRef]
- Kristine M, O.; Rune, S.; Unni, S.L.; Cato, B.; Trond, L.; Peter, R.; Michel, V.; Cathrine, L. Temperature and nitrogen effects on regulators and products of the flavonoid pathway: Experimental and kinetic model studies. Plant Cell Environ. 2009, 32, 286–299. [Google Scholar]
- Ruan, J.; Ma, L.; Shi, Y. Potassium management in tea plantations: Its uptake by field plants, status in soils, and efficacy on yields and quality of teas in China. J. Plant Nutr. Soil Sci. 2013, 176, 450–459. [Google Scholar] [CrossRef]
- Zhu, Q.; Liu, L.; Lu, X.; Du, X.; Xiang, P.; Cheng, B.; Tan, M.; Huang, J.; Wu, L.; Kong, W.; et al. The biosynthesis of EGCG, theanine and caffeine in response to temperature is mediated by hormone signal transduction factors in tea plant (Camellia sinensis L.). Front. Plant Sci. 2023, 14, 1149182. [Google Scholar] [CrossRef]
- Zhang, Q.; Tang, D.; Yang, X.; Geng, S.; He, Y.; Chen, Y.; Yi, X.; Ni, K.; Liu, M.; Ruan, J. Plant Availability of Magnesium in Typical Tea Plantation Soils. Front. Plant Sci. 2021, 12, 641501. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Chen, X.; Zhang, Y.; Huang, Z.; Yin, J.; Weng, X.; Yang, W.; Wu, H.; Zhang, F.; Wu, L. Magnesium is a nutritional tool for the yield and quality of oolong tea (Camellia sinensis L.) and reduces reactive nitrogen loss. Sci. Hortic. 2023, 308, 111590. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, Y.; Hu, H.; Shi, Y.; Tang, D.; Ruan, J.; Fernie, A.R.; Liu, M.-Y. Magnesium promotes tea plant growth via enhanced glutamine synthetase-mediated nitrogen assimilation. Plant Physiol. 2023, 192, 1321–1337. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Xiao, L.; Huang, Y.; Meng, Y.; Xiao, B.; Yang, Y. Effects of different concentrations of magnesium on photosynthetic and photoprotective ability of tea plants. Northwest Agric. For. J. 2021, 30, 1210–1219. [Google Scholar]
- Liu, Y.; Tian, J.; Liu, B.; Zhuo, Z.; Shi, C.; Xu, R.; Xu, M.; Liu, B.; Ye, J.; Sun, L.; et al. Effects of pruning on mineral nutrients and untargeted metabolites in fresh leaves of Camellia sinensis cv. Shuixian. Front. Plant Sci. 2022, 13, 1016511. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, L.; Shan, Y.; Liu, Y.; Tian, Y.; Xia, T. Influence of shade on flavonoid biosynthesis in tea (Camellia sinensis (L.) O. Kuntze). Sci. Hortic. 2012, 141, 7–16. [Google Scholar] [CrossRef]
- Livigni, S.; Lucini, L.; Sega, D.; Navacchi, O.; Pandolfini, T.; Zamboni, A.; Varanini, Z. The different tolerance to magnesium deficiency of two grapevine rootstocks relies on the ability to cope with oxidative stress. BMC Plant Biol. 2019, 19, 148. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zeng, L.; Chen, Y.; Wang, X.; Liao, Y.; Xiao, Y.; Fu, X.; Yang, Z. Metabolism of Gallic Acid and Its Distributions in Tea (Camellia sinensis) Plants at the Tissue and Subcellular Levels. Int. J. Mol. Sci. 2020, 21, 5684. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, S.; Yang, Y.; Zeng, H.; Yu, L. Research progress on the influence of organic acids in tea on tea quality and its detection methods. Food Technol. 2021, 46, 254–259. [Google Scholar] [CrossRef]
- Xiang, P.; Zhu, Q.; Zhang, L.; Xu, P.; Liu, L.; Li, Y.; Cheng, B.; Wang, X.; Liu, J.; Shi, Y.; et al. Integrative analyses of transcriptome and metabolome reveal comprehensive mechanisms of Epigallocatechin-3-gallate (EGCG) biosynthesis in response to ecological factors in tea plant (Camellia sinensis). Food Res. Int. 2023, 166, 112591. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Zhang, X.; He, Y.; Li, L.; Wang, Y.; Hong, G.; Xu, P. Transcriptomic Analysis Reveals the Molecular Adaptation of Three Major Secondary Metabolic Pathways to Multiple Macronutrient Starvation in Tea (Camellia sinensis). Genes 2020, 11, 241. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Yang, H.; Wang, S.; Zhao, J.; Liu, C.; Gao, L.; Xia, E.; Lu, Y.; Tai, Y.; She, G.; et al. Draft genome sequence of Camellia sinensis var. sinensis provides insights into the evolution of the tea genome and tea quality. Proc. Natl. Acad. Sci. USA 2018, 115, E4151–E4158. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Xia, E.; Fu, J.; Xu, Y.; Zhao, X.; Tong, W.; Tang, Q.; Tadege, M.; Fernie, A.R.; Zhao, J. Diverse roles of MYB transcription factors in regulating secondary metabolite biosynthesis, shoot development, and stress responses in tea plants (Camellia sinensis). Plant J. 2022, 110, 1144–1165. [Google Scholar] [CrossRef]
- Lin, J.; Wilson, I.W.; Ge, G.; Sun, G.; Xie, F.; Yang, Y.; Wu, L.; Zhang, B.; Wu, J.; Zhang, Y.; et al. Whole transcriptome analysis of three leaf stages in two cultivars and one of their F1 hybrid of Camellia sinensis L. with differing EGCG content. Tree Genet. Genomes 2017, 13, 13. [Google Scholar] [CrossRef]
- GB/T 8313-2018; Determination of Total Polyphenols and Catechins Content in Tea. State Administration for Market Regulation and Administration Standardization Administration of the People’s Republic of China: Beijing, China, 2018.
- GB/T 8314-2013; Tea—Determination of Free Amino Acids Content. General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China and Administration Standardization Administration of the People’s Republic of China: Beijing, China, 2013.
- GB 5009.241-2017; Determination of Magnesium in Food of National Standard for Food Safety. The National Health and Family Planning Commission of the People’s Republic of China and the State Food and Drug Administration: Beijing, China, 2017.
- Lin, X.; Chen, X.; Wang, P.; Zheng, Y.; Guo, Y.; Hong, Y.; Yang, R.; Ye, N. Metabolite profiling in albino tea mutant Camellia sinensis ‘Fuyun 6’ using LC–ESI–MS/MS. Trees 2021, 36, 261–272. [Google Scholar] [CrossRef]
- Yang, N.; Li, Z.; Liu, M.; Ma, J.; Shi, Y.; Wei, X.; Liu, Y.; Mao, Y.; Gao, S. Study on the regulatory effect of potassium nutrition on EGCG biosynthesis in tea plants (Camellia sinensis). Tea Sci. 2024, 44, 887–900. [Google Scholar] [CrossRef]
- Gao, T.; Hou, B.-H.; Shao, S.-X.; Xu, M.-T.; Zheng, Y.-C.; Jin, S.; Wang, P.-J.; Ye, N.-X. Differential metabolites and their transcriptional regulation in seven major tea cultivars (Camellia sinensis) in China. J. Integr. Agr. 2023, 22, 3346–3363. [Google Scholar] [CrossRef]
- Wang, P.; Yu, J.; Jin, S.; Chen, S.; Yue, C.; Wang, W.; Gao, S.; Cao, H.; Zheng, Y.; Gu, M.; et al. Genetic basis of high aroma and stress tolerance in the oolong tea cultivar genome. Hortic. Res. 2021, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Chen, S.; Gu, M.; Chen, X.; Chen, X.; Yang, J.; Zhao, F.; Ye, N. Exploration of the Effects of Different Blue LED Light Intensities on Flavonoid and Lipid Metabolism in Tea Plants via Transcriptomics and Metabolomics. Int. J. Mol. Sci. 2020, 21, 4606. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Treated Magnesium Concentration (mmol/L) | ||||
|---|---|---|---|---|---|
| 0 | 0.15 | 0.45 | 0.6 | 0.9 | |
| Gallic acid (GA) | 1.69 ± 0.03 ab | 1.78 ± 0.12 a | 1.71 ± 0.02 ab | 1.66 ± 0.02 b | 1.67 ± 0.05 b |
| Caffeine (CA) | 5.18 ± 0.20 a | 4.29 ± 1.85 a | 4.99 ± 0.48 a | 4.92 ± 0.14 a | 4.71 ± 0.20 a |
| Catechin (C) | 1.55 ± 0.03 a | 1.45 ± 0.05 b | 1.47 ± 0.09 b | 1.42 ± 0.19 b | 1.34 ± 0.02 c |
| Epicatechin (EC) | 1.59 ± 0.38 a | 1.59 ± 0.12 a | 1.52 ± 0.39 a | 1.50 ± 0.27 a | 1.49 ± 0.08 a |
| Epigallocatechin gallate (EGCG) | 60.84 ± 2.72 ab | 63.82 ± 2.19 a | 63.56 ± 1.33 ab | 56.87 ± 4.67 ab | 59.12 ± 3.36 b |
| Epigallocatechin (EGC) | 48.80 ± 4.17 ab | 52.74 ± 2.43 ab | 54.68 ± 1.69 a | 45.76 ± 2.47 b | 47.34 ± 1.88 ab |
| Gallocatechin (GC) | 23.31 ± 0.7 ab | 14.68 ± 0.2 a | 26.91 ± 1.22 a | 19.95 ± 3.38 bc | 23.91 ± 1.29 ab |
| Epicatechin gallate (ECG) | 14.14 ± 0.48 b | 27.01 ± 0.89 a | 14.39 ± 0.29 b | 13.07 ± 0.47 b | 13.85 ± 0.55 b |
| Total catechins | 151.96 ± 6.47 ab | 163.11 ± 5.61 a | 164.27 ± 4.48 a | 140.29 ± 3.93 b | 153.767 ± 3.12 ab |
| Tea polyphenols | 12.12 ± 0.62 c | 12.73 ± 0.33 bc | 12.30 ± 0.29 bc | 13.82 ± 0.62 a | 13.78 ± 0.14 a |
| Free amino acids | 0.63 ± 0.022 ab | 0.69 ± 0.099 a | 0.62 ± 0.037 ab | 0.63 ± 0.015 ab | 0.60 ± 0.049 ab |
| Phenolammonia ratio | 19.28 ± 1.54 bc | 18.77 ± 3.08 bc | 20.63 ± 0.85 b | 22.12 ± 0.98 ab | 23.14 ± 2.11 ab |
| Gene Name | Upstream Primer (5′-3′) | Downstream Primers (5′-3′) |
|---|---|---|
| CsHD.15G0015490-F3′H | GCCCAATGCTGATGTTAGGG | ATGGACCAAGGTCGCAGTTA |
| CsHD.13G0004300-F3′5′H | TTTCTCGACGTTGTGATGGC | CCCATTCAACTGTGCTTGCT |
| CsHD.11G0000850-WRKY | GAGCCGAGATTTGCGTTCAT | TTTGGCCGTACTTCCTCCAT |
| CsHD.03G0018510-MYB | AGGCTCATGGAGAAGGCAAT | TCATCCACCTTAGCCTGCAA |
| CsHD.02G0004890-XTH7 | ACGAAGCAAGAGGAATCCCA | CAGTCATCGGCTTCCCAAAG |
| CsHD.02G0004890-VTC | AGAGGCCGACTGAGTTTCAA | GCATTCCGGAAGAACTGGAC |
| CsHD.01G0001670-VDE | ACGCATGGGATGGATATGGT | CTCCAGTCTCTCCACAAGGG |
| CsHD.07G0004130-PSAK | AGCAAACAGGAAAGCCACAG | AGAGCCACAAGCCAAGGTAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Z.; Li, Z.; Yan, R.; Yang, N.; Liu, M.; Bai, Y.; Mao, Y.; Zhou, C.; Guo, Y.; Zeng, Y.; et al. Metabolome and Transcriptome Analysis Reveals the Regulatory Effect of Magnesium Treatment on EGCG Biosynthesis in Tea Shoots (Camellia sinensis). Plants 2025, 14, 684. https://doi.org/10.3390/plants14050684
Feng Z, Li Z, Yan R, Yang N, Liu M, Bai Y, Mao Y, Zhou C, Guo Y, Zeng Y, et al. Metabolome and Transcriptome Analysis Reveals the Regulatory Effect of Magnesium Treatment on EGCG Biosynthesis in Tea Shoots (Camellia sinensis). Plants. 2025; 14(5):684. https://doi.org/10.3390/plants14050684
Chicago/Turabian StyleFeng, Zixuan, Zhuan Li, Rui Yan, Nan Yang, Meichen Liu, Yueting Bai, Yuyuan Mao, Chengzhe Zhou, Yuqiong Guo, Yulin Zeng, and et al. 2025. "Metabolome and Transcriptome Analysis Reveals the Regulatory Effect of Magnesium Treatment on EGCG Biosynthesis in Tea Shoots (Camellia sinensis)" Plants 14, no. 5: 684. https://doi.org/10.3390/plants14050684
APA StyleFeng, Z., Li, Z., Yan, R., Yang, N., Liu, M., Bai, Y., Mao, Y., Zhou, C., Guo, Y., Zeng, Y., Ji, Y., Lin, Y., Chen, J., & Gao, S. (2025). Metabolome and Transcriptome Analysis Reveals the Regulatory Effect of Magnesium Treatment on EGCG Biosynthesis in Tea Shoots (Camellia sinensis). Plants, 14(5), 684. https://doi.org/10.3390/plants14050684