
The Status of Wild Grapevine (Vitis vinifera L. subsp. sylvestris (C.C. Gmel.) Hegi) Populations in Georgia (South Caucasus)

 ,
,  , , , , , , ,
, , , , , , ,
Abstract
1. Introduction
2. Results
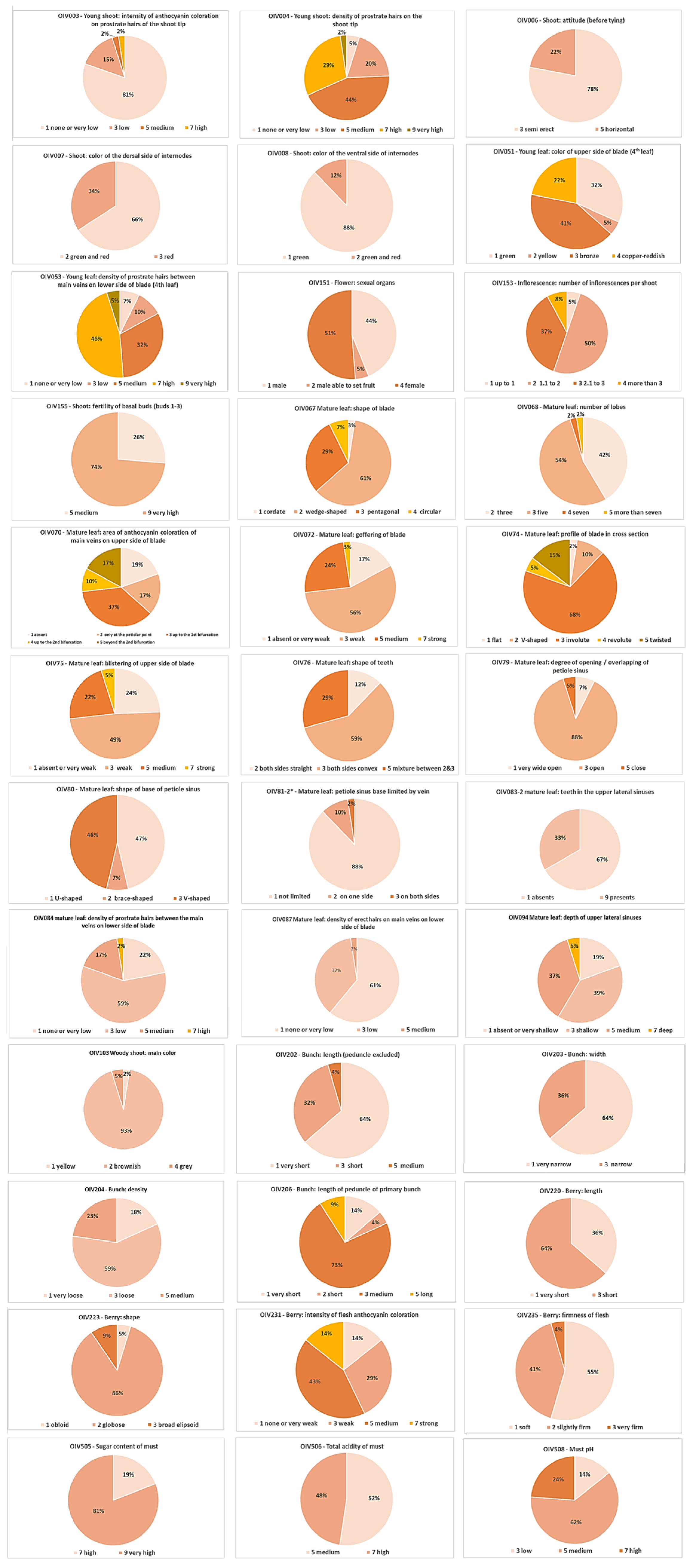
2.1. Ampelography
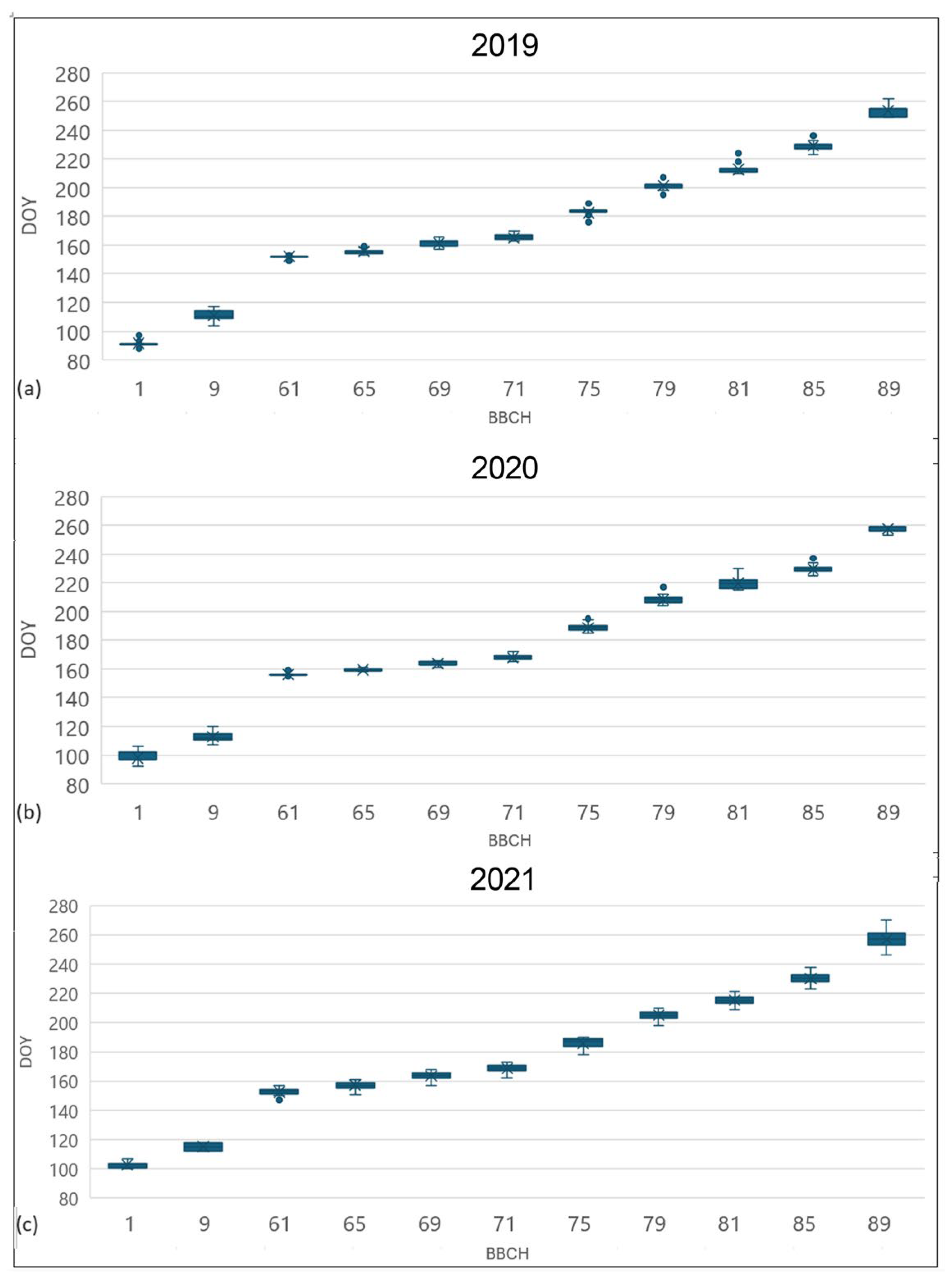
2.2. Phenology
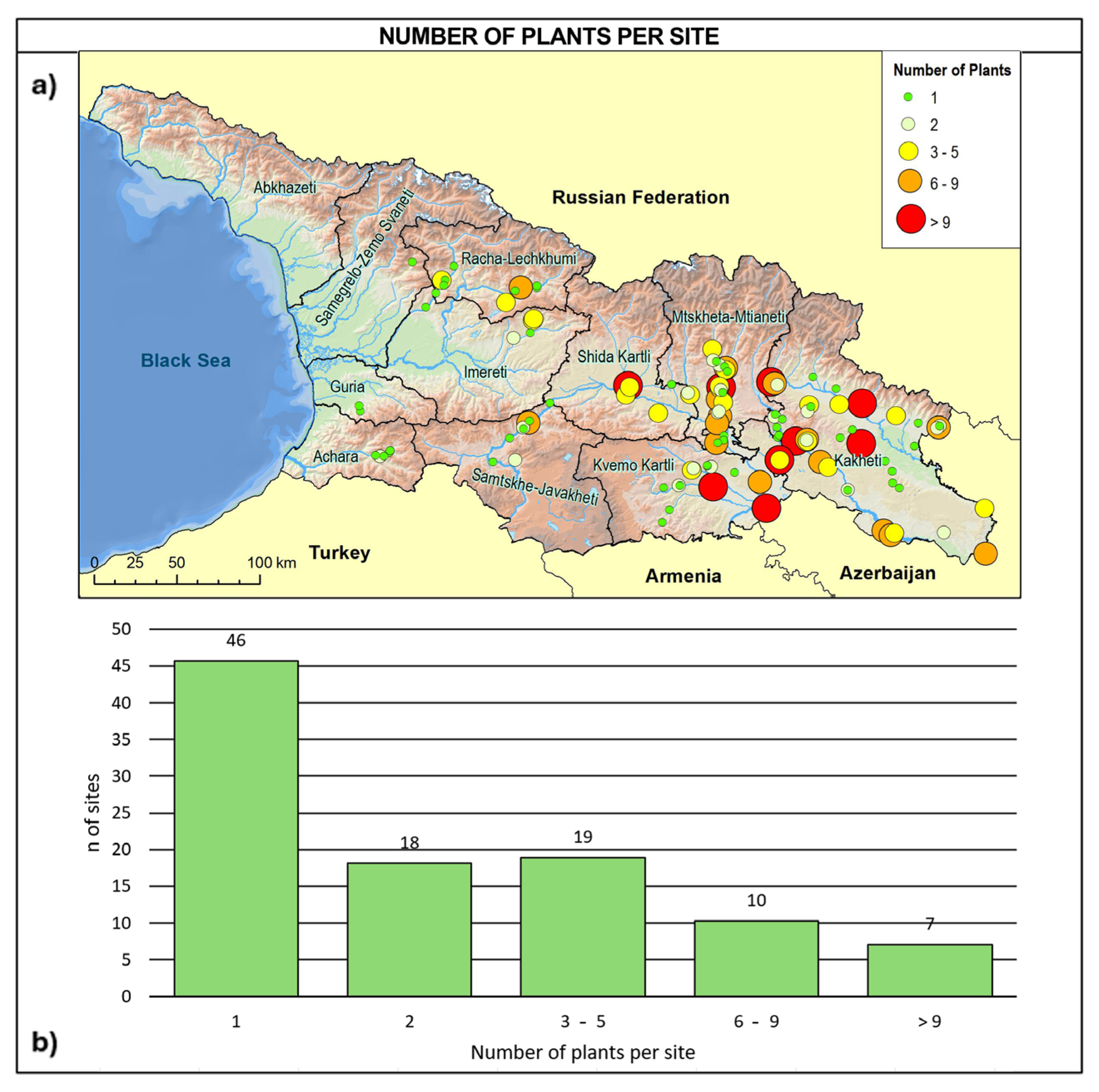
2.3. Consistency and Ecology of the Georgian Wild Grapevine Populations
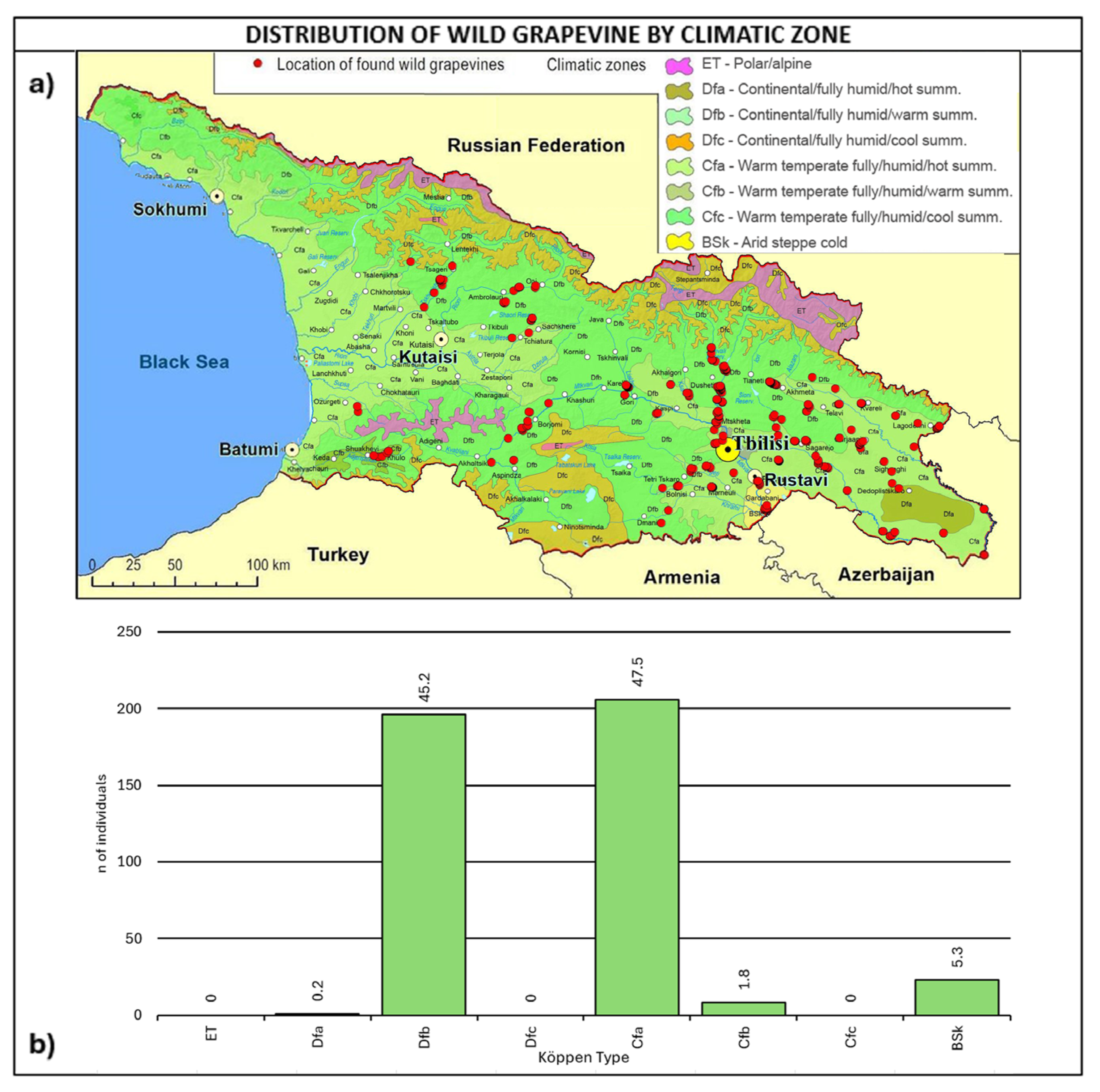
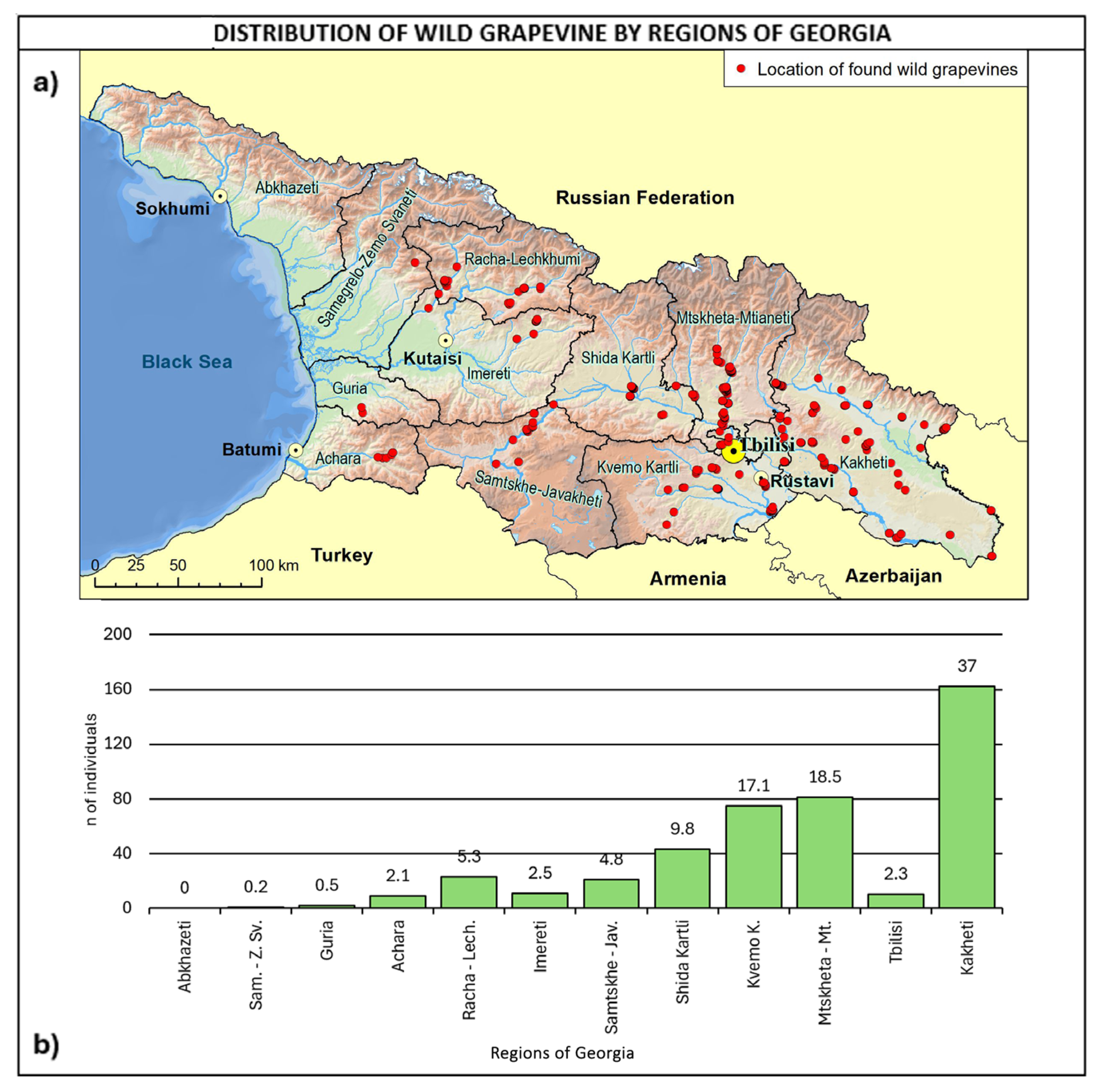
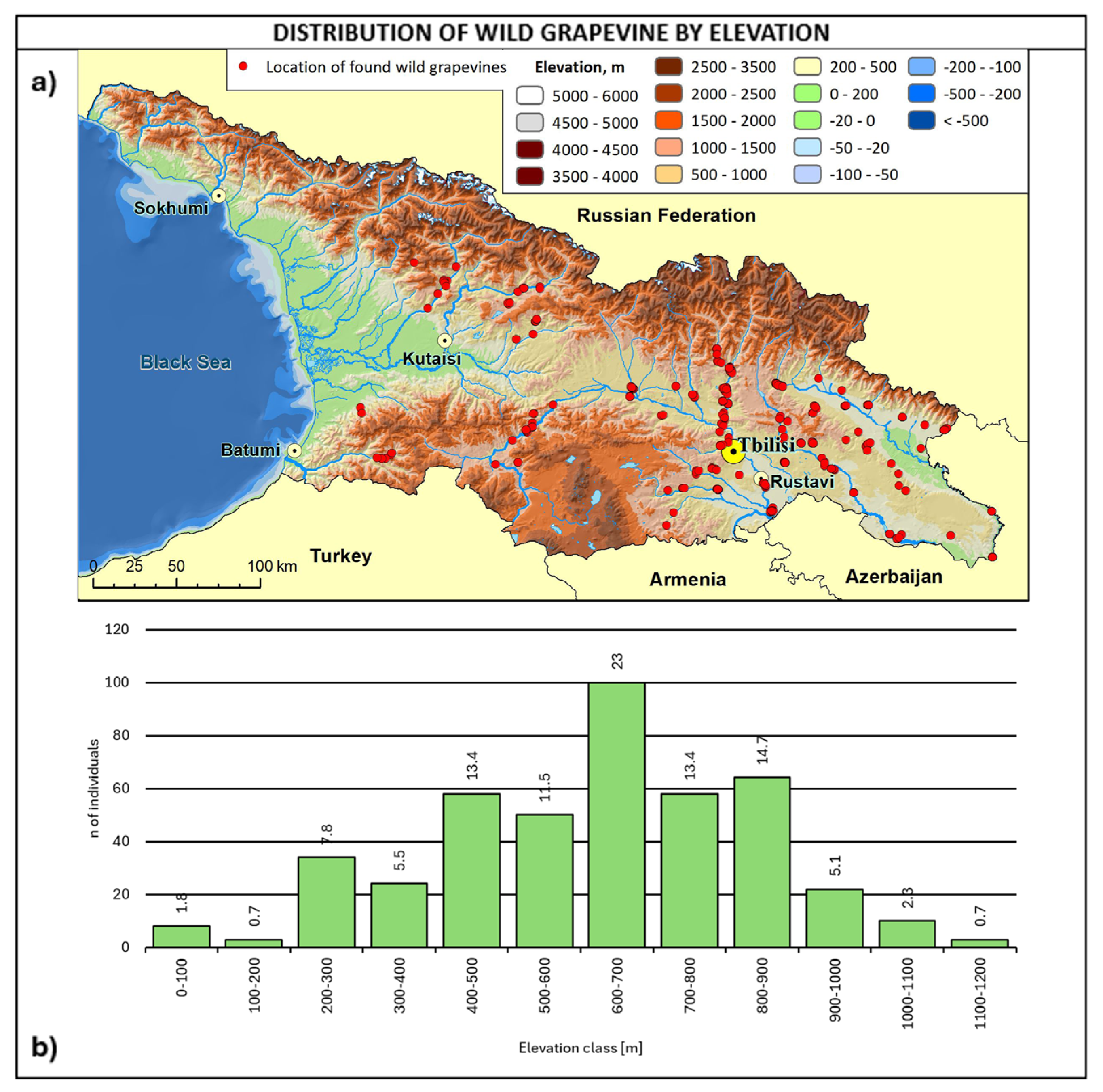
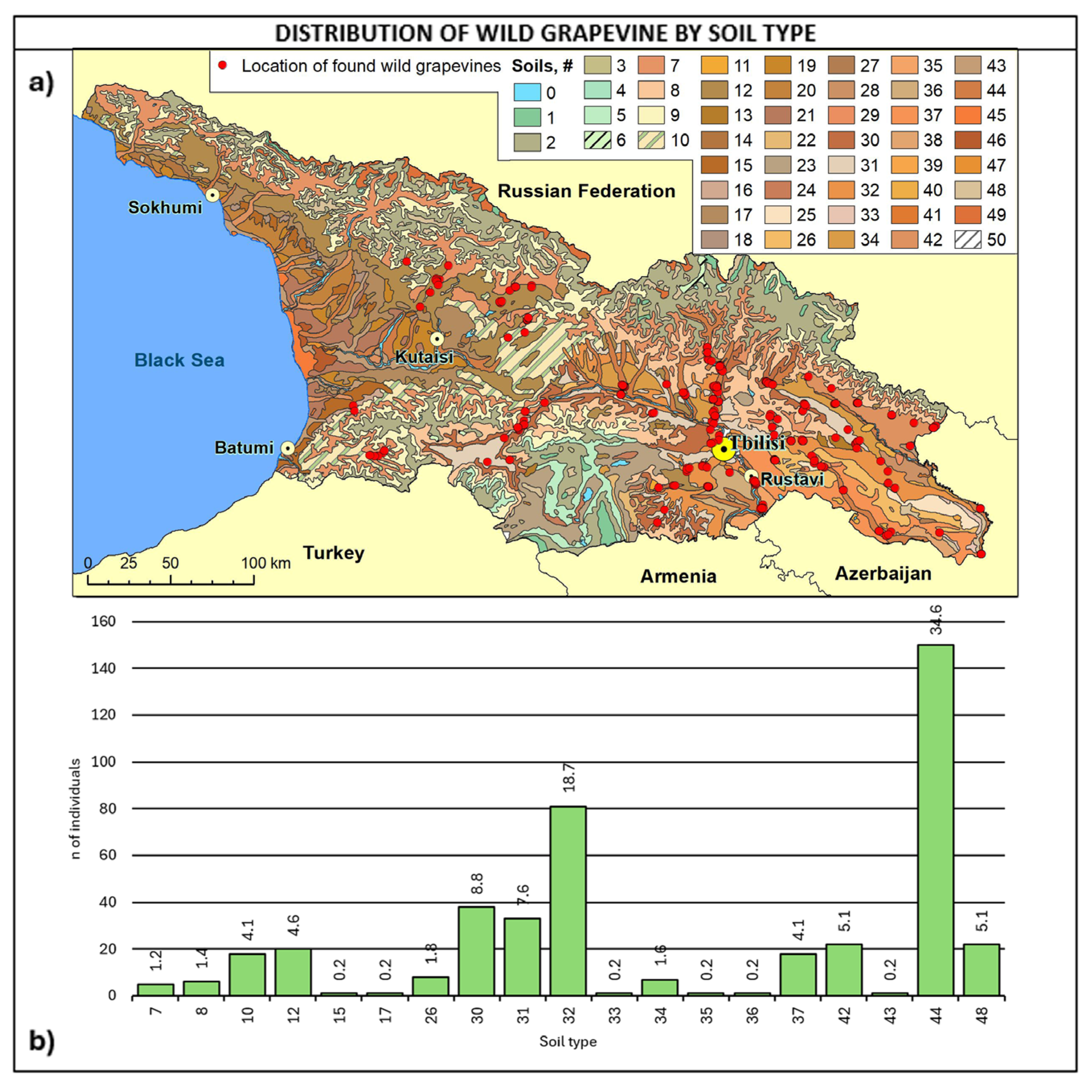
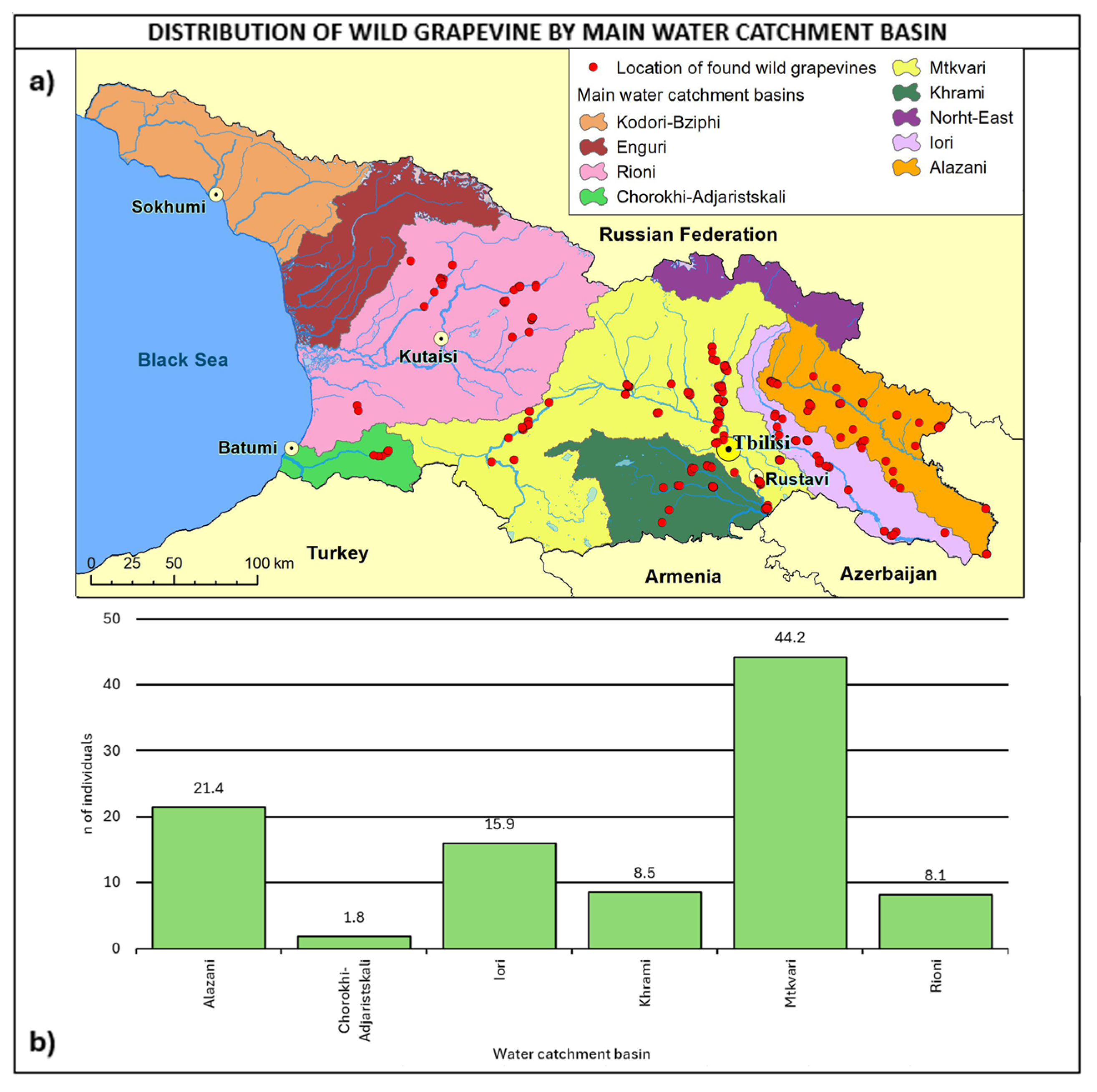
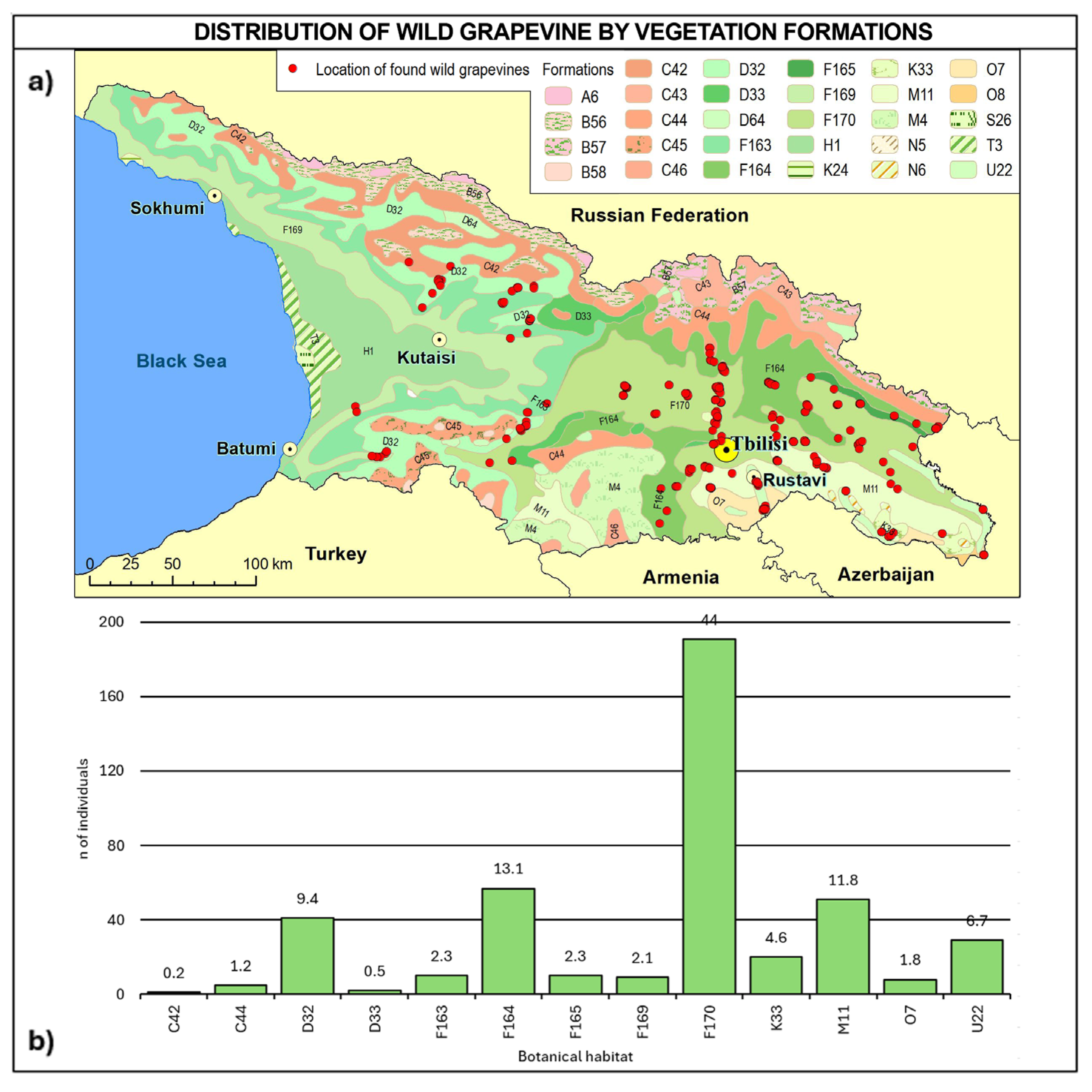
3. Discussion
4. Materials and Methods
4.1. Wild Grapevine Survey
4.2. The Jighaura Collection
4.3. Ampelographical and Phenological Records
4.4. The Georgian Wild Grapevine Populations Database
- Administrative region
- Elevation
- Main water catchment basin
- Vegetation formation
- Soil type
- Köppen climate type
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Descriptor | Code | Descriptor | Code | Descriptor |
|---|---|---|---|---|---|
| 001 | Young Shoot: aperture of tip | 075 | Mature leaf: blistering of upper side of blade | 209 | Bunch: number of wings of the primary bunch |
| 003 | 003 Young shoot: intensity of anthocyanin coloration on prostrate hairs of the shoot tip | 076 | Mature leaf: shape of teeth | 220 | Berry: length |
| 004 | Young shoot: density of prostrate hairs on the shoot tip | 079 | Mature leaf: degree of opening/overlapping of petiole sinus | 221 | Berry: width |
| 006 | Shoot: attitude (before tying) | 080 | Mature leaf: shape of base of petiole sinus | 223 | Berry: shape |
| 007 | Shoot: color of the dorsal side of internodes | 081-1 | Mature leaf: teeth in the petiole sinus | 225 | Berry: color of skin |
| 008 | Shoot: color of the ventral side of internodes | 081-2 | Mature leaf: petiole sinus base limited by veins | 231 | Berry: intensity of the anthocyanin coloration of flesh |
| 016 | Shoot: number of consecutive tendrils | 083-3 | Mature leaf: teeth in the upper lateral sinuses | 235 | Berry: firmness of flesh |
| 051 | Young leaf: color of upper side of blade (4th leaf) | 084 | Mature leaf: density of prostrate hairs between the main veins on lower side of blade | 236 | Berry: particularity of flavor |
| 053 | Young leaf: density of prostrate hairs between main veins on lower side of blade (4th leaf) | 087 | Mature leaf: density of erect hairs on main veins on lower side of blade | 241 | Berry: formation of seeds |
| 151 | Flower: sexual organs | 094 | Mature leaf: depth of upper lateral sinuses | 502 | Bunch: weight of a single bunch |
| 153 | Inflorescence: number of inflorescences per shoot | 103 | Woody shoot: main color | 503 | Berry: single berry weight |
| 155 | Shoot: fertility of basal buds (buds 1–3) | 202 | Bunch: length (peduncle excluded) | 505 | Sugar content of must |
| 067 | Mature leaf: shape of blade | 203 | Bunch: width | 506 | Total acidity of must |
| 070 | Mature leaf: area of anthocyanin coloration of main veins on upper side of blade | 204 | Bunch: density | 508 | Must pH |
| 072 | Mature leaf: goffering of blade | 206 | Bunch: length of peduncle of primary bunch | ||
| 074 | Mature leaf: profile of blade in cross section | 208 | Bunch: shape |
| Gender | Occurrences (N) | Frequency (%) |
|---|---|---|
| Female | 29 | 6.7 |
| Probable female | 29 | 6.7 |
| Male | 34 | 7.8 |
| Probable male | 14 | 3.2 |
| Unknown | 328 | 75.6 |
| Total | 434 | 100.0 |
| Gender | Occurrences (N) | Frequency (%) |
|---|---|---|
| Female and probable female | 57 | 54.3 |
| Male and probable male | 48 | 45.7 |
| Total | 105 | 100.0 |
| Soil N | Soil Type | Soil N | Soil Type |
|---|---|---|---|
| 7 | Brown forest acid-(umbric cambisols) | 33 | Cinnamonic light |
| 8 | Brown forest weakly unsaturated-(eutric cambisols) | 34 | Meadow-cinnamonic |
| 10 | Yellow-brown forest | 35 | Meadow gray-cinnamonic |
| 12 | Raw-humus calcareus (calcaric)-(rendzinas) | 36 | Gray-cinnamonic dark |
| 15 | Red soils | 37 | Gray-cinnamonic-(ermic cambisols) |
| 17 | Yellow soils-(haplic alisoils) | 42 | Alluvial acid-(dystric iluvisols) |
| 26 | Black calcareus | 43 | Alluvial saturated-(eutric fluvisols) |
| 30 | Cinnamonic leached | 44 | Alluvial calcareous-(calcaric fluvisoils) |
| 31 | Cinnamonic-(chromic cambisols) | 48 | Strongly eroded soils and bare rocks (dystric or eutric lithosols) |
| 32 | Cinnamonic calcareus |
| Code | Description |
|---|---|
| C | Subarctic, boreal, and nemoral-montane open woodlands as well as subalpine and oro-Mediterranean vegetation |
| C42 | Southwest Caucasian krummholz and open woodlands (Betula litwinowii, Fagus sylvatica subsp. orientalis, Acer trautvetteri) with Betula megrelica, Quercus pontica, scrub (Rhododendron caucasicum) with Rhamnus imeretina (on carbonate rocks with Corylus colchica), tall-forb communities (Heracleum ponticum) with Delphinium pyramidatum (on carbonate rocks with Heracleum aconitifolium, Ligusticum arafoe), and grasslands (Calamagrostis arundinacea, Stachys macrantha, on carbonate rocks with Geum speciosum, Carex pontica) |
| C44 | Southeast and low Caucasian krummholz and open woodlands (Betula litwinowii, Acer trautvetteri, Quercus macranthera), scrub (Rhododendron caucasicum), tall-forb communities (Heracleum sosnowskyi, Aconitum orientale), and grasslands (Festuca woronowii, Calamagrostis arundinacea, Geranium ibericum) |
| D | Mesophytic and hygromesophytic coniferous and mixed broad-leaved–coniferous forests |
| D32 | West Caucasian fir, spruce–fir and beech–fir forests (Abies nordmanniana, Picea orientalis, Fagus sylvatica subsp. orientalis) with evergreen understorey (Rhododendron ponticum, Prunus laurocerasus, Ilex colchica), often alternating with Oriental beech forests (Fagus sylvatica subsp. orientalis) |
| D33 | Caucasian fir, spruce–fir and beech–fir forests (Abies nordmanniana, Picea orientalis, Fagus sylvatica subsp. orientalis) without evergreen understorey, partly alternating with Oriental beech forests (Fagus sylvatica subsp. orientalis) |
| D64 | Caucasian pine forests (Pinus kochiana), partly alternating with birch forests (Betula litwinowii, B. raddeana) and spruce forests (Picea orientalis) |
| F | Mesophytic deciduous broad-leaved and mixed coniferous–broad-leaved forests |
| F163 | East Euxinian-Caucasian Oriental beech forests (Fagus sylvatica subsp. orientalis), partly with Picea orientalis, mostly with evergreen understorey (Prunus laurocerasus, Rhododendron ponticum, Daphne pontica) with Hedera colchica, Ilex colchica, Ruscus colchicus |
| F164 | Caucasian Oriental beech forests (Fagus sylvatica subsp. orientalis) with Carpinus betulus, partly with Picea orientalis, without evergreen understorey, partly alternating with oak-hornbeam forests (Carpinus betulus, Quercus iberica) |
| F165 | East Caucasian submontane to montane hornbeam–maple–Oriental beech forests (Fagus sylvatica subsp. orientalis, Acer velutinum, Carpinus betulus) with Hedera pastuchowii in combination with hornbeam–chestnut–oak forests (Quercus iberica, Castanea sativa, Carpinus betulus) |
| F169 | East Euxinian oak and hornbeam–oak forests (Quercus iberica, Carpinus orientalis, C. betulus), alternating with hornbeam–chestnut–Oriental beech forests (Carpinus betulus, Fagus sylvatica subsp. orientalis, Castanea sativa) with evergreen understorey |
| F170 | Transcaucasian oak forests (Quercus iberica), hornbeam–oak forests (Quercus iberica, Carpinus betulus) and Oriental hornbeam–oak forests (Quercus iberica, Carpinus orientalis), with Acer cappadocicum, Sorbus torminalis, partly in combination with shibliak communities (scrub) |
| H | Hygro-thermophilous mixed deciduous broad-leaved forests |
| H1 | Colchic lowland to submontane mixed oak forests (Quercus imeretina, Q. hartwissiana, Zelkova carpinifolia, Carpinus betulus, Castanea sativa, Fagus sylvatica subsp. orientalis) with evergreen understorey species (Rhododendron ponticum, Prunus laurocerasus), alternating with oak and hornbeam–oak forests (Quercus iberica, Carpinus betulus) in the submontane belt |
| K | Xerophytic coniferous forests, woodlands, and scrub |
| K33 | Transcaucasian colline-montane juniper open woodlands (Juniperus polycarpos, J. foetidissima), partly in combination with Pistacia mutica-open woodlands |
| M | Steppes |
| M11 | Pre- and Transcaucasian Stipa-steppes (Stipa tirsa, S. pulcherrima, S. daghestanica, S. transcaucasica, S. araxensis, S. pontica, S. holosericea, S. hohenackeriana) with Elytrigia gracillima, Astragalus haesitabundus, A. gjunaicus, Medicago daghestanica, Onobrychis transcaucasica, Linaria megrica and Bothriochloa ischaemum-steppes with Onobrychis ruprechtii, O. kachetica, Salvia daghestanica, Hyssopus officinalis subsp. angustifolius, Medicago caerulea, Polygala transcaucasica, alternating with tomillares (Thymus daghestanicus, T. karamarjanicus, T. tiflisiensis, Thymus kotschyanus, Scutellaria orientalis) and thorn-cushion communities (Astragalus aureus, A. denudatus, A. microcephalus, A. uraniolimneus) |
| O | Deserts |
| O7 | East Transcaucasian wormwood deserts (Artemisia fragrans) with ephemeroids (Poa bulbosa, Catabrosella humilis) |
| U | Vegetation of flood-plains, estuaries and fresh-water polders, and other moist or wet sites |
| U22 | Transcaucasian hardwood alluvial forests (Quercus pedunculiflora, Ulmus minor) in combination with poplar and willow alluvial forests (Populus × canescens, P. nigra, Salix excelsa) as well as Tamarix ramosissima-scrub |
References
- Ardenghi, N.M.G.; Galasso, G.; Banfi, E. Vitaceae. Euro+Med Plantbase—The Information Resource for Euro-Mediterranean Plant Diversity. 2017. Available online: http://ww2.bgbm.org/EuroPlusMed/PTaxonDetail.asp?NameId=34507&PTRefFk=7100000 (accessed on 26 June 2024).
- POWO “Plants of the World Online”. Facilitated by the Royal Botanic Gardens, Kew. 2024. Available online: http://www.plantsoftheworldonline.org/ (accessed on 26 June 2024).
- Zohary, D.; Weiss, E.; Hopf, M. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin. Fruit Trees and Nuts, 4th ed.; Oxford Academic: Oxford, UK, 2012. [Google Scholar]
- Arnold, C.; Guillet, F.; Gobat, J.M. Situation de la vigne sauvage Vitis vinifera ssp. silvestris en Europe. Vitis J. Grapevine Res. 1998, 37, 159–170. [Google Scholar]
- Arnold, C. Ecologie de la vigne sauvage, Vitis vinífera L. ssp. sylvestris (Gmelin) Hegi, dans les forêts alluviales et colluviales d’Europe. Geobot. Helv. 2002, 76, 1–256. [Google Scholar]
- Arroyo Garía, R.; Revilla, E. The Current Status of Wild Grapevine Populations (Vitis vinifera ssp. sylvestris) in the Mediterranean Basin. In The Mediterranean Genetic Code; Poljuha, D., Sladonja, B., Eds.; IntechOpen: Rijeka, Croatia, 2013. [Google Scholar]
- Grassi, F.; Labra, M.; Imazio, S.; Ocete Rubio, R.; Failla, O.; Scienza, A.; Sala, F. Phylogeographical structure and conservation genetics of wild grapevine. Conserv. Genet. 2006, 7, 837–845. [Google Scholar] [CrossRef]
- Benito, A.; Muñoz-Organero, G.; de Andrés, M.T.; Ocete, R.; García-Muñoz, S.; López, M.; Arroyo-García, R.; Cabello, F. Ex situ ampelographical characterization of wild Vitis vinifera from fifty-one Spanish populations. Aust. J. Grape Wine Res. 2017, 23, 143–152. [Google Scholar] [CrossRef]
- Cunha, J.; Baleiras-Couto, M.; Cunha, J.P.; Banza, J.; Soveral, A.; Carneiro, L.C.; Eiras-Dias, J.E. Characterization of Portuguese populations of Vitis vinifera L. ssp. sylvestris (Gmelin) Hegi. Genet. Resour. Crop Evol. 2007, 54, 981–988. [Google Scholar] [CrossRef]
- De Andrés, M.T.; Benito, A.; Pérez-Rivera, G.; Ocete, R.; López, M.A.; Gaforio, L.; Muñoz, G.; Cabello, F.; Martínez-Zapater, J.M.; Arroyo-García, R. Genetic diversity of wild grapevine populations in Spain and their genetic relationships with cultivated grapevines. Mol. Ecol. 2012, 21, 800–816. [Google Scholar] [CrossRef]
- Iriarte-Chiapusso, M.J.; Ocete-Pérez, C.A.; Hernández-Beloqui, B.; Ocete-Rubio, R. Vitis vinifera in the Iberian Peninsula: A review. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2016, 151, 245–257. [Google Scholar] [CrossRef]
- Ocete, R.; Arroyo-Garcia, R.; Morales, M.L.; Cantos, M.; Gallardo, A.; Pérez, M.A.; Gómez, I.; López, M.A. Characterization of Vitis vinifera L. subspecies sylvestris (Gmelin) Hegi in the Ebro river Basin (Spain). Vitis J. Grapevine Res. 2011, 50, 11–16. [Google Scholar]
- Zinelabidine, L.H.; Haddioui, A.; Bravo, G.; Arroyo-García, R.; Martínez Zapater, J.M. Genetic origins of cultivated and wild grapevines from Morocco. Am. J. Enol. Vitic. 2010, 61, 83–90. [Google Scholar] [CrossRef]
- Selmi, I.; Pacifico, D.; Lehad, A.; Garfi, G.; Carimi, F.; Mahfoudhi, N. Occurrence of grapevine virus a in wild grapevines (Vitis vinifera subsp. sylvestris) in Tunisia. J. Plant Pathol. 2017, 99, 817. [Google Scholar]
- Arnold, C.; Schnitzler, A.; Parisot, C.; Maurin, A. Historical Reconstruction of a Relictual Population of Wild Grapevines (Vitis vinifera ssp. sylvestris, Gmelin, Hegi), in a Floodplain Forest of the Upper Seine Valley, France. River Res. Appl. 2010, 26, 904–914. [Google Scholar] [CrossRef]
- Lacombe, T.; Laucou, V.; Di Vecchi, M.; Bordenave, L.; Bourse, T.; Siret, R.; David, J.; Boursiquot, J.M.; Bronner, A.; Merdinoglu, D.; et al. Inventory and characterization of Vitis vinifera ssp. silvestris in France. Acta Hortic. 2003, 603, 553–557. [Google Scholar] [CrossRef]
- Biagini, B.; De Lorenzis, G.; Imazio, S.; Failla, O.; Scienza, A. Italian wild grapevine (Vitis vinifera L. subsp. sylvestris) population: Insights into eco-geographical aspects and genetic structure. Tree Genet. Genomes 2014, 5, 1369–1385. [Google Scholar] [CrossRef]
- Schneider, A.; Boccacci, P.; Ruffa, P.; Torello Marinoni, D.; Cavallo, L.; Festari, I.; Rotti, G.; Raimondi, S. Identification and characterization of Vitis vinifera subsp. sylvestris populations in north-western Italy. Vitis J. Grapevine Res. 2015, 54, 223–225. [Google Scholar]
- Arnold, C.; Schnitzler, A.; Douard, A.; Peter, R.; Gillet, F. Is there a future for wild grapevine (Vitis vinifera subsp. silvestris) in the Rhine Valley? Biodivers. Conserv. 2005, 14, 1507–1523. [Google Scholar] [CrossRef]
- Regner, F.; Hack, R.; Gangl, H.; Leitner, G.; Mandl, K.; Tiefenbrunner, W. Genetic variability and incidence of systemic diseases in wild vines (Vitis vinifera ssp. silvestris) along the Danube. Vitis J. Grapevine Res. 2004, 43, 123–130. [Google Scholar]
- Zdunić, G.; Maul, E.; Hančević, K.; Leko, M.; Butorac, L.; Mucalo, A.; Radić, T.; Šimon, S.; Budić-Leto, I.; Mihaljević, M.Ž.; et al. Genetic diversity of wild grapevine [Vitis vinifera L. subsp. sylvestris (Gmel.) Hegi] in the eastern Adriatic region. Am. J. Enol. Vitic. 2017, 68, 252–257. [Google Scholar] [CrossRef]
- Zdunić, G.; Lukšić, K.; Nagy, Z.A.; Mucalo, A.; Hančević, K.; Radić, T.; Butorac, L.; Jahnke, G.G.; Kiss, E.; Ledesma-Krist, G.; et al. Genetic structure and relationships among wild and cultivated grapevines from central Europe and part of the western Balkan peninsula. Genes 2020, 11, 962. [Google Scholar] [CrossRef]
- Perko, A.; Trapp, O.; Maul, E.; Röckel, F.; Piltaver, A.; Vršič, S. Monitoring and Genotyping of Wild Grapevine (Vitis vinifera L. subsp. sylvestris) in Slovenia. Plants 2024, 13, 9. [Google Scholar] [CrossRef]
- Kullaj, E.; Ŝimon, S.; Pejic, I.; Çakalli, A. Molecular characterization of wild grapevine (Vitis vinifera ssp. sylvestris) populations of northern Albania. Acta Hortic. 2012, 948, 129–134. [Google Scholar] [CrossRef]
- Susaj, L.; Susaj, E.; Ferraj, B.; Dragusha, B. Identification of the main characters and accompanying plants of wild type grapevine [Vitis vinifera L. ssp. sylvestris (gmelin) Hegi], through Shkrelis Valley, Malësia e Madhe. In Proceedings of the 5th Annual International Conference “Albanians and Europa 2020—Statement and Prospects”, Prishtina, Kosovo, 1–3 October 2010; Volume: Abstract Book, p. 111. [Google Scholar]
- Bodor, P.; Höhn, M.; Pedryc, A.; Deák, T.; Dücso, I.; Uzun, I.; Cseke, K.; Böhm, É.I.; Bisztray, G.D. Conservation value of the native Hungarian wild grape (Vitis sylvestris Gmel.) evaluated by microsatellite markers. Vitis J. Grapevine Res. 2010, 49, 23–27. [Google Scholar]
- Bartha, D.; Kevey, B.; Tiborcz, V. Current and 20th century distributions of Vitis sylvestris in Hungary. Folia Oecolog. 2012, 39, 99–106. [Google Scholar]
- Jahnke, G.; Nagy, Z.A.; Koltai, G.; Hajdu, E.; Májer, J. Preservation and Characterization of Woodland Grape (Vitis vinifera ssp. sylvestris GMEL.) Genotypes of the Szigetköz, Hungary. In Germplasm: Characteristics, Diversity and Preservation; Nova Science Publishers: Hauppauge, NY, USA, 2016; pp. 27–45. [Google Scholar]
- Popescu, C.F.; Dejeu, L.C.; Ocete Rubio, R. Preliminary Characterization of Wild Grapevine Populations (Vitis vinifera ssp. sylvestris) Grown Along the Danube River. Not. Bot. Horti Agrobot. 2013, 41, 472–477. [Google Scholar] [CrossRef]
- Dzhambazova, T.; Tsvetkov, I.; Atanassov, I.; Rusanov, K.; Zapater JM, M.; Atanassov, A.; Hvarleva, T. Genetic diversity in native Bulgarian grapevine germplasm (Vitis vinifera L.) based on nuclear and chloroplast microsatellite polymorphisms. Vitis J. Grapevine Res. 2009, 48, 115–121. [Google Scholar]
- Ergül, A.; Perez-Rivera, G.; Söylemezoğlu, G.; Kazan, K.; Arroyo-Garcia, R. Genetic diversity in Anatolian wild grapes (Vitis vinifera subsp. sylvestris) estimated by SSR markers. Plant Genet. Resour. 2011, 9, 375–383. [Google Scholar] [CrossRef]
- Karataş, D.D.; Karataş, H.; Laucou, V.; Sarikamiş, G.; Riahi, L.; Bacilieri, R.; This, P. Genetic diversity of wild and cultivated grapevine accessions from southeast Turkey. Hereditas 2014, 151, 73–80. [Google Scholar] [CrossRef]
- Karataş, D.D.; Karataş, H.; García-Muñoz, S. Morphological characterization of endangered wild grapevine Vitis vinifera ssp. silvestris in Eastern Turkey. J. Am. Pomol. Soc. 2014, 68, 14–23. [Google Scholar]
- Amanov, M.V. Wild Grapevine of Azerbaijan; Azerneshr: Baku, Azerbaijan, 1998. (In Azerbaijani) [Google Scholar]
- Ocete Rubio, R.; Ocete Rubio, E.; Ocete Pérez, C.; Ángeles Pérez Izquierdo, M.; Rustioni, L.; Failla, O.; Chipashvili, R.; Maghradze, D. Ecological and Sanitary Characteristics of the Eurasian Wild Grapevine (Vitis vinifera L. ssp. sylvestris (Gmelin) Hegi) in Georgia (Caucasian Region). Plant Genet. Resour. 2012, 10, 155–162. [Google Scholar] [CrossRef]
- Maghradze, D.; Salimov, V.; Melyan, G.; Musayev, M.; Ocete, C.A.; Chipashvili, R.; Failla, O.; Ocete, R. Sanitary status of the Eurasian wild grapevine in the South Caucasian region. Vitis 2015, 54, 203–205. [Google Scholar]
- Maghradze, D.; Kikilashvili, S.; Mamasakhlisashvili, L.; Mdinaradze, I.; Chipashvili, R.; Giorgobiani, S.; Kenchiashvili, S.; Ujmajuridze, L.; Gotsiridze, O.; Bitsadze, N.; et al. Ampelographic description of wild grape accessions. In Wild Grapevine in Georgia, Multidisciplinary Comparative Research to Unravel the Mystery of Its Domestication; Maghradze, D., Failla, O., Eds.; Shota Rustaveli National Science Foundation of Georgia (SRNCFG): Tbilisi, Georgia, 2022; pp. 254–345. [Google Scholar]
- Kikvadze, M.; Kikilashvili, S.; Bitsadze, N.; Maghradze, T.; De Lorenzis, G.; Rubio, R.O.; Rivera, D.; Bacilieri, R.; Failla, O.; Maghradze, D. Wildly growing Eurasian grapevine (Vitis vinifera L.) in Georgia: Composition, research and efforts for the preservation. Acta Hortic. 2024, 1385, 19–24. [Google Scholar] [CrossRef]
- Rahimi, O.; Berger, J.Z.; Shtein, I.; Kher, M.M.; Frumin, S.; Hübner, S.; Weiss, E.; Drori, E. Wild Grapevine (Vitis vinifera L. subsp. sylvestris (C.C. Gmelin) Hegi)—Novel Species to the Israeli Flora. Horticulturae 2023, 9, 998. [Google Scholar] [CrossRef]
- Naqinezhad, A.; Ramezani, E.; Djamali, M.; Schnitzler, A.; Arnold, C. Wild grapevine (Vitis vinifera subsp. sylvestris) in the Hyrcanian relict forests of northern Iran: An overview of current taxonomy, ecology and palaeorecords. J. For. Res. 2018, 29, 1757–1768. [Google Scholar] [CrossRef]
- Marrano, A.; Grzeskowiak, L.; Moreno Sanz, P.; Lorenzi, S.; Prazzoli, M.L.; Arzumanov, A.; Amanova, M.; Failla, O.; Maghradze, D.; Grando, M.S. Genetic diversity and relationships in the grapevine germplasm collection from Central Asia. Vitis 2015, 54, 233–237. [Google Scholar]
- Anzani, R.; Failla, O.; De Micheli, L.; Scienza, A. Individuazione e conservazione del germoplasma di vite selvatica (Vitis vinifera silvestris) in Italia. Vignevini 1993, 20, 51–60. [Google Scholar]
- Mc Govern, P. (University of Pennsylvania, Philadelphia, PA, USA). Survey Along the Tigris River. Personal communication, 2024. [Google Scholar]
- Ocete, R.; Fevereiro, P.; Failla, O. Proposal for the wild grapevine (Vitis vinifera L. subsp. sylvestris (Gmelin) Hegi) conservation in the European countries. Vitis 2015, 54, 281–282. [Google Scholar]
- Petitpierre, B.; Arnold, C.; Phelps, L.N.; Guisan, A. A tale of three vines: Current and future threats to wild Eurasian grapevine by vineyards and invasive rootstocks. Divers. Distrib. 2023, 29, 1594–1608. [Google Scholar] [CrossRef]
- Schneider, A.; Ruffa, P.; Tumino, G.; Fontana, M.; Boccacci, P.; Raimondi, S. Genetic relationships and introgression events between wild and cultivated grapevines (Vitis vinifera L.): Focus on Italian Lambruscos. Sci. Rep. 2024, 14, 12392. [Google Scholar] [CrossRef]
- D’Onofrio, C. Introgression among cultivated and wild grapevine in Tuscany. Front. Plant Sci. 2020, 11, 202. [Google Scholar] [CrossRef]
- Imazio, S.; Labra, M.; Grassi, F.; Scienza, A.; Failla, O. Chloroplast microsatellites to investigate the origin of grapevine. Genet. Resour. Crop Evol. 2006, 53, 1003–1011. [Google Scholar] [CrossRef]
- Imazio, S.; Maghradze, D.; De Lorenzis, G.; Bacilieri, R.; Laucou, V.; This, P.; Scienza, A.; Failla, O. From the cradle of grapevine domestication: Molecular overview and description of Georgian grapevine (Vitis vinifera L.) germplasm. Tree Genet. Genomes 2013, 9, 641–658. [Google Scholar] [CrossRef]
- McGovern, P.; Jalabadze, M.; Batiuk, S.; Callahan, M.P.; Smith, K.E.; Hall, G.R.; Kvavadze, E.; Maghradze, D.; Rusishvili, N.; Bouby, L.; et al. Early Neolithic wine of Georgia in the South Caucasus. Proc. Natl. Acad. Sci. USA 2017, 114, E10309–E10318. [Google Scholar] [CrossRef] [PubMed]
- Riaz, S.; De Lorenzis, G.; Velasco, D.; Koehmstedt, A.; Maghradze, D.; Bobokashvili, Z.; Musayev, M.; Zdunic, G.; Laucou, V.; Walker, M.A.; et al. Genetic diversity analysis of cultivated and wild grapevine (Vitis vinifera L.) accessions around the Mediterranean basin and Central Asia. BMC Pant Biol. 2018, 18, 137. [Google Scholar] [CrossRef] [PubMed]
- Magris, G.; Jurman, I.; Fornasiero, A.; Paparelli, E.; Schwope, R.; Marroni, F.; Di Gaspero, G.; Morgante, M. The genomes of 204 Vitis vinifera accessions reveal the origin of European wine grapes. Nat. Commun. 2021, 12, 7240. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Duan, S.; Xia, Q.; Liang, Z.; Dong, X.; Margaryan, K.; Musayev, M.; Goryslavets, S.; Zdunić, G.; Bert, P.F.; et al. Dual domestications and origin of traits in grapevine evolution. Science 2023, 379, 892–901. [Google Scholar] [CrossRef]
- Kolenati, F.A. Versuch einer systematischen Anordnung der in Grusien einheimischen Reben, nebst einem oekonomisch-technischen Anhange. Bull. Naturforschenden Ges. Moscau 1846, 19, 1–104. [Google Scholar]
- Negrul, A. Origin of cultivated grapevine and its classification. In Ampelography of the Soviet Union; Frollov-Bagreev, A.M., Ed.; Pishepromizdat: Moscow, Russia, 1946; Volume 1, pp. 159–216. (In Russian) [Google Scholar]
- Ramishvili, R. Dikorastushchii Vinograd Zakavkazia (Wild Grape of the South Caucasus); Publishing House ‘Ganatleba’: Tbilisi, Georgia, 1988. (In Russian) [Google Scholar]
- Ramishvili, R. History of Georgian Grapevine and Wine; Tbilisi, Georgia, 2001; p. 240. (In Georgian) [Google Scholar]
- Ekhvaia, J.; Akhalkatsi, M. Morphological variation and relationships of Georgian populations of Vitis vinifera L. subsp. sylvestris (C.C. Gmel.) Hegi. Flora Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 608–617. [Google Scholar] [CrossRef]
- Ekhvaia, J.; Gurushidze, M.; Blattner, F.R.; Akhalkatsi, M. Genetic diversity of Vitis vinifera in Georgia: Relationships between local cultivars and wild grapevine, V. vinifera L. subsp. sylvestris. Genet. Resour. Crop Evol. 2014, 61, 1507–1521. [Google Scholar] [CrossRef]
- Maghradze, D.; Failla, O.; Turok, J.; Amanov, M.; Avidzba, A.; Chkhartishvili, N.; Costantini, L.; Cornea, V.; Hausman, J.F.; Gasparian, S.; et al. East-West collaboration for grapevine diversity exploration and mobilization of adaptive traits for breeding: A four years story. Vitis 2015, 54, 1–4. [Google Scholar]
- Maghradze, D.; Imazio, S.; Failla, O. Conservation and sustainable use grapevine genetic resources in the Caucasus and Northern Black Sea area. Acta Hortic. 2009, 827, 155–158. [Google Scholar] [CrossRef]
- OIV. The OIV (International Organization of Vine and Wine) Ampelographic Descriptors; OIV: Paris, France, 2007. [Google Scholar]
- Meier, U. (Ed.) Growth Stages of Mono-and Dicotyledonous Plants, 2nd ed.; BBCH Monograph; Federal Biological Research Centre for Agriculture and Forestry: Braunschweig, Germany, 2009; 158p. [Google Scholar]
- European Environment Agency. Corine Land Cover 2018 [Data Set]. 2019. Available online: https://www.eea.europa.eu/data-and-maps/data/copernicus-land-monitoring-service-corine (accessed on 12 March 2024).
- Köppen, W.; Geiger, G. Handbuch der Klimatologie; Verlag von Gebrüder Borntraeger: Berlin, Germany, 1936; Volume 1, pp. 1–44. [Google Scholar]
- Geiger, R. Klassifikation der Klimate nach W. Köppen. In Landolt-Börnstein—Zahlenwerte und Funktionen aus Physik, Chemie, Astronomie, Geophysik und Technik; Alte Serie 3; Springer: Berlin/Heidelberg, Germany, 1954; pp. 603–607. [Google Scholar]
- Urushadze, T. (Ed.) Soil Map of Georgia with a Scale of 1:500000, Prepared Georgian State Project and Scientific-Research Institut of Land Organization; Joint-Stock Company “Cartography”: Tbilisi, Georgia, 1991. [Google Scholar]
- Bohn, U.; Zazanashvili, N.; Nakhutsrishvili, G. The Map of the Natural Vegetation of Europe and its application in the Caucasus Ecoregion. Georgian Acad. Sci. 2007, 175, 112–121. [Google Scholar]
- Ocete, R.; Arnoldi, C.; Failla, O.; Lovicy, G.; Biagini, B.; Imazio, S.; Lara, M.; Maghradze, D.; Angeles López, M. Considerations on the European wild grapevine (Vitis vinifera L. ssp. sylvestris (Gmelin) Hegi) and Phylloxera infestation. Vitis 2011, 50, 97–98. [Google Scholar]
- Röckel, F.; Margaryan, K.; Merkouropoulos, G.; Laucou, V.; De, G.; Tello, J.; Zdunic, G.; Teresa, M.; Baeta, F.; Cunha, J.; et al. A Population Genetic Study of Vitis vinifera L. subsp. sylvestris Gmelin Based on 3.000 Individuals from 20 Countries; IVES Conference Series; OIV: Dijon, France, 2024. [Google Scholar]
- Maghradze, D.; Kikilashvili, S.; Gotsiridze, O.; Maghradze, T.; Fracassetti, D.; Failla, O.; Rustioni, L. Comparison between the Grape Technological Characteristics of Vitis vinifera subsp. sylvestris and subsp. sativa. Agronomy 2021, 11, 472. [Google Scholar] [CrossRef]
- Ricciardi, V.; Crespan, M.; Maddalena, G.; Migliaro, D.; Brancadoro, L.; Maghradze, D.; Failla, O.; Toffolatti, S.L.; De Lorenzis, G. Novel loci associated with resistance to downy and powdery mildew in grapevine. Front. Plant Sci. 2024, 15, 1386225. [Google Scholar] [CrossRef]
- Bitsadze, N.; Kikilashvili, S.; Chipashvili, R.; Mamasakhlisashvili, L.; Maghradze, T.; Kikvadze, M.; Toffolatti, S.L.; De Lorenzis, G.; Failla, O.; Ocete Rubio, R.; et al. Resistance to downy mildew in wildly growing Eurasian Vitis vinifera L. grapevines. J. Plant Pathol. 2024, 106, 1759–1771. [Google Scholar] [CrossRef]
- De Lorenzis, G.; Chipashvili, R.; Failla, O.; Maghradze, D. Study of genetic variability in Vitis vinifera L. germplasm by high-throughput Vitis18kSNP array: The case of Georgian genetic resources. BMC Plant Biol. 2015, 15, 154. [Google Scholar] [CrossRef]
- Bacilieri, R.; Maghradze, D.; Grando, S.; Pejic, I.; Maul, E.; Munoz, G.; Eiras-Dias, J.; Schneider, A.; Boselli, M.; This, P. Conservation, characterisation and management of grapevine genetic resources: The European project Grapegen06. In Proceedings of the Materials of OIV 33d World Congress on Vine and Wine, Tbilisi, Georgia, 20–27 June 2010; pp. 1–13. [Google Scholar]
- Rustioni, L.; Maghradze, D.; Popescu, C.F.; Cola, G.; Abashidze, E.; Aroutiounian, R.; Brazao, J.; Coletti, S.; Cornea, V.; Dejeu, L.; et al. First results of the European grapevine collections’ collaborative network: Validation of a standard eno-carpological phenotyping method. Vitis J. Grapevine Res. 2014, 53, 219–226. [Google Scholar]
- Sharikadze, K.; Togonidze, N.; Ekhvaia, J. Investigation of Wild Grape (Vitis vinifera L. ssp. sylvestris (C.C. Gmel) Hegi) Biodiversity in Georgia; Science and Technologies: Tbilisi, Georgia, 2010; pp. 38–51. (In Georgian) [Google Scholar]
- Zdunić, G.; Maul, E.; Eiras Dias, J.E.J.; Munoz Organero, G.; Carka, F.; Maletic, E.; Savvides, S.; Jahnke, G.G.; Nagy, Z.A.; Nikolic, D.; et al. Guiding principles for identification, evaluation and conservation of Vitis vinifera subsp. sylvestris. Vitis 2017, 56, 127–131. [Google Scholar]
- Rustioni, L.; Failla, O.; Maul, E.; Maghradze, D.; Chipashvili, R.; Cornea, V. Application of standard methods for the grapevine (Vitis vinifera L.) phenotypic diversity exploration: Phenological traits. Acta Hortic. 2014, 1032, 253–260. [Google Scholar] [CrossRef]
- Pipia, I.; Gogniashvili, M.; Tabidze, V.; Beridze, T.; Gamkrelidze, M.; Gotsiridze, V.; Melyan, G.; Musaev, M.; Salimov, V.; Benk, J.; et al. Plastid DNA sequence diversity in wild grapevine samples (Vitis vinifera subsp. sylvestris) from the Caucasus region. Vitis 2012, 51, 119–124. [Google Scholar]
- NASA JPL. NASA Shuttle Radar Topography Mission Global 1 Arc Second [Data Set]. 2024. Available online: https://data.nasa.gov/dataset/NASA-Shuttle-Radar-Topography-Mission-Global-1-arc/4mak-tnnx/data (accessed on 10 February 2024).


| Descriptor | N | Mode | Variance | Min. | Max. |
|---|---|---|---|---|---|
| 001 Young Shoot: aperture of tip | 41 | 5 | 0.000 | 5 | 5 |
| 003 Young shoot: intensity of anthocyanin coloration on prostrate hairs of the shoot tip | 41 | 1 | 1.605 | 1 | 7 |
| 004 Young shoot: density of prostrate hairs on the shoot tip | 41 | 5 | 3.190 | 1 | 9 |
| 006 Shoot: attitude (before tying) | 41 | 3 | 0.702 | 3 | 5 |
| 007 Shoot: color of the dorsal side of internodes | 41 | 2 | 0.230 | 2 | 3 |
| 008 Shoot: color of the ventral side of internodes | 41 | 1 | 0.110 | 1 | 2 |
| 016 Shoot: number of consecutive tendrils | 41 | 1 | 0.000 | 1 | 1 |
| 051 Young leaf: color of upper side of blade (4th leaf) | 41 | 3 | 1.355 | 1 | 4 |
| 053 Young leaf: density of prostrate hairs between main veins on lower side of blade (4th leaf) | 41 | 7 | 3.888 | 1 | 9 |
| 151 Flower: sexual organs | 41 | 4 | 2.199 | 1 | 4 |
| 153 Inflorescence: number of inflorescences per shoot | 37 | 2 | 0.646 | 1 | 5 |
| 155 Shoot: fertility of basal buds (buds 1–3) | 40 | 9 | 3.272 | 5 | 9 |
| 067 Mature leaf: shape of blade | 41 | 2 | 0.449 | 1 | 4 |
| 068 Mature leaf: number of lobes | 41 | 3 | 0.430 | 2 | 5 |
| 070 Mature leaf: area of anthocyanin coloration of main veins on upper side of blade | 41 | 3 | 1.760 | 1 | 5 |
| 072 Mature leaf: goffering of blade | 41 | 3 | 2.039 | 1 | 7 |
| 074 Mature leaf: profile of blade in cross section | 41 | 3 | 0.811 | 1 | 5 |
| 075 Mature leaf: blistering of upper side of blade | 41 | 3 | 2.678 | 1 | 7 |
| 076 Mature leaf: shape of teeth | 41 | 3 | 1.105 | 2 | 5 |
| 079 Mature leaf: degree of opening/overlapping of petiole sinus | 41 | 3 | 0.498 | 1 | 5 |
| 080 Mature leaf: shape of base of petiole sinus | 41 | 1–3 | 0.950 | 1 | 3 |
| 081-1 Mature leaf: teeth in the petiole sinus | 41 | 1 | 0.000 | 1 | 1 |
| 081-2 Mature leaf: petiole sinus base limited by veins | 41 | 1 | 0.422 | 1 | 3 |
| 083-2 Mature leaf: teeth in the upper lateral sinuses | 41 | 1 | 3.901 | 1 | 9 |
| 084 Mature leaf: density of prostrate hairs between the main veins on lower side of blade | 41 | 3 | 2.000 | 1 | 7 |
| 087 Mature leaf: density of erect hairs on main veins on lower side of blade | 41 | 1 | 1.195 | 1 | 5 |
| 094 Mature leaf: depth of upper lateral sinuses | 41 | 3 | 2.805 | 1 | 7 |
| 103 Woody shoot: main color | 41 | 2 | 0.220 | 1 | 4 |
| 202 Bunch: length (peduncle excluded) | 22 | 1 | 1.394 | 1 | 5 |
| 203 Bunch: width | 22 | 1 | 0.970 | 1 | 3 |
| 204 Bunch: density | 22 | 3 | 1.706 | 1 | 5 |
| 206 Bunch: length of peduncle of primary bunch | 22 | 3 | 0.981 | 1 | 5 |
| 208 Bunch: shape | 22 | 1 | 0.000 | 1 | 1 |
| 209 Bunch: number of wings of the primary bunch | 22 | 2 | 0.000 | 2 | 2 |
| 220 Berry: length | 22 | 3 | 0.970 | 1 | 3 |
| 221 Berry: width | 22 | 3 | 1.013 | 1 | 3 |
| 223 Berry: shape | 21 | 2 | 0.148 | 1 | 3 |
| 225 Berry: color of skin | 22 | 6 | 0.000 | 6 | 6 |
| 231 Berry: intensity of the anthocyanin coloration of flesh | 21 | 5 | 3.429 | 1 | 7 |
| 235 Berry: firmness of flesh | 22 | 1 | 0.357 | 1 | 3 |
| 236 Berry: particularity of flavor | 22 | 1 | 0.000 | 1 | 1 |
| 241 Berry: formation of seeds | 22 | 3 | 0.000 | 3 | 3 |
| 502 Bunch: weight of a single bunch | 22 | 1 | 0.000 | 1 | 1 |
| 503 Berry: single berry weight | 22 | 1 | 0.000 | 1 | 1 |
| 505 Sugar content of must | 21 | 9 | 0.648 | 7 | 9 |
| 506 Total acidity of must | 21 | 5 | 1.048 | 5 | 7 |
| 508 Must pH | 21 | 5 | 1.562 | 3 | 7 |
| BBCH | Statistics | 2019 | 2020 | 2021 |
|---|---|---|---|---|
| 9 bud break | Average [DOY] | 111.1 | 112.7 | 115.4 |
| Minimum [DOY] | 104 | 107 | 112 | |
| Maximum [DOY] | 117 | 120 | 118 | |
| Standard deviation [n days] | 3.7 | 3.3 | 2.7 | |
| 65 full flowering | Average [DOY] | 155.7 | 159.3 | 157.1 |
| Minimum [DOY] | 153 | 157 | 151 | |
| Maximum [DOY] | 159 | 162 | 161 | |
| Standard deviation [n days] | 1.3 | 1.0 | 2.4 | |
| 75 berries pea-sized | Average [DOY] | 182.5 | 188.8 | 185.8 |
| Minimum [DOY] | 176 | 185 | 178 | |
| Maximum [DOY] | 189 | 195 | 190 | |
| Standard deviation [n days] | 3.4 | 2.2 | 3.5 | |
| 85 softening of berries | Average [DOY] | 229.1 | 229.4 | 230.1 |
| Minimum [DOY] | 223 | 220 | 223 | |
| Maximum [DOY] | 236 | 237 | 238 | |
| Standard deviation [n days] | 2.4 | 3.4 | 4.1 |
| Model | Sum of Squares | gl | Quadratic Mean | F | Sign. | ||
|---|---|---|---|---|---|---|---|
| Regression | 91.343 | 2 | 45.672 | 4.824 | 0.010 | ||
| Residual | 880.532 | 93 | 9.468 | ||||
| Total | 971.875 | 95 | |||||
| Model | Unstandardized coefficients | Standardized coefficients | t | Sign. | |||
| B | Standard Error | Beta | |||||
| (Costant) | 61.885 | 19.986 | 3.09 | 0.003 | |||
| Elevation (m) | 0.004 | 0.003 | 0.168 | 1.38 | 0.168 | ||
| Longitude (°) | −1.447 | 0.471 | −0.371 | −3.07 | 0.003 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cola, G.; De Lorenzis, G.; Failla, O.; Kvaliashvili, N.; Kikilashvili, S.; Kikvadze, M.; Mamasakhlisashvili, L.; Mdinaradze, I.; Chipashvili, R.; Maghradze, D. The Status of Wild Grapevine (Vitis vinifera L. subsp. sylvestris (C.C. Gmel.) Hegi) Populations in Georgia (South Caucasus). Plants 2025, 14, 232. https://doi.org/10.3390/plants14020232
Cola G, De Lorenzis G, Failla O, Kvaliashvili N, Kikilashvili S, Kikvadze M, Mamasakhlisashvili L, Mdinaradze I, Chipashvili R, Maghradze D. The Status of Wild Grapevine (Vitis vinifera L. subsp. sylvestris (C.C. Gmel.) Hegi) Populations in Georgia (South Caucasus). Plants. 2025; 14(2):232. https://doi.org/10.3390/plants14020232
Chicago/Turabian StyleCola, Gabriele, Gabriella De Lorenzis, Osvaldo Failla, Nikoloz Kvaliashvili, Shengeli Kikilashvili, Maia Kikvadze, Londa Mamasakhlisashvili, Irma Mdinaradze, Ramaz Chipashvili, and David Maghradze. 2025. "The Status of Wild Grapevine (Vitis vinifera L. subsp. sylvestris (C.C. Gmel.) Hegi) Populations in Georgia (South Caucasus)" Plants 14, no. 2: 232. https://doi.org/10.3390/plants14020232
APA StyleCola, G., De Lorenzis, G., Failla, O., Kvaliashvili, N., Kikilashvili, S., Kikvadze, M., Mamasakhlisashvili, L., Mdinaradze, I., Chipashvili, R., & Maghradze, D. (2025). The Status of Wild Grapevine (Vitis vinifera L. subsp. sylvestris (C.C. Gmel.) Hegi) Populations in Georgia (South Caucasus). Plants, 14(2), 232. https://doi.org/10.3390/plants14020232