Optimizing Valerianella locusta L. Growth and Metabolism by Combining Red and Blue LED Light: Insights into Plant Physiology, Biochemistry, and Nutraceutical Value

 , ,
, ,  , , , ,
, , , ,  ,
,  and
and
Abstract
1. Introduction
2. Results
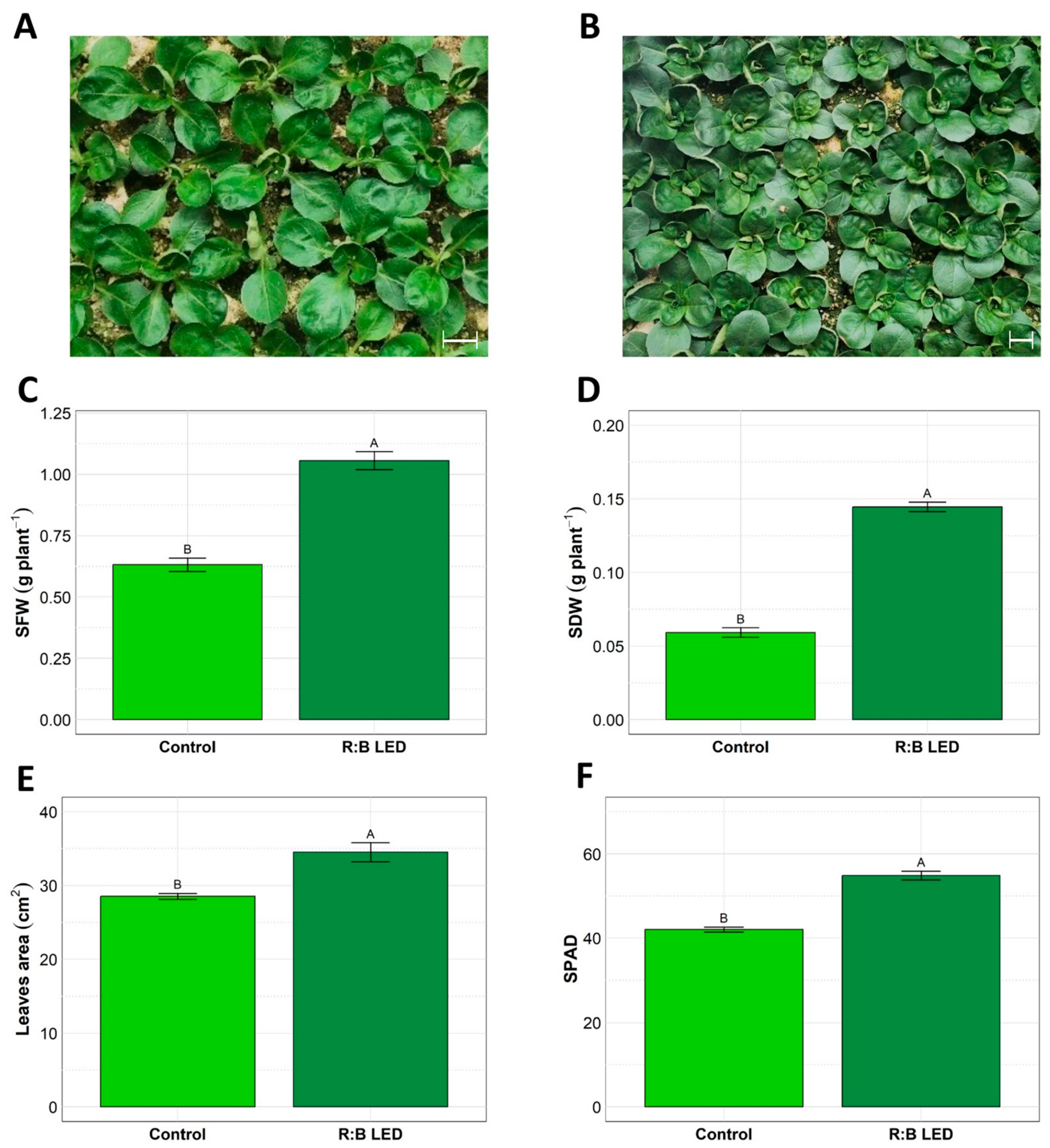
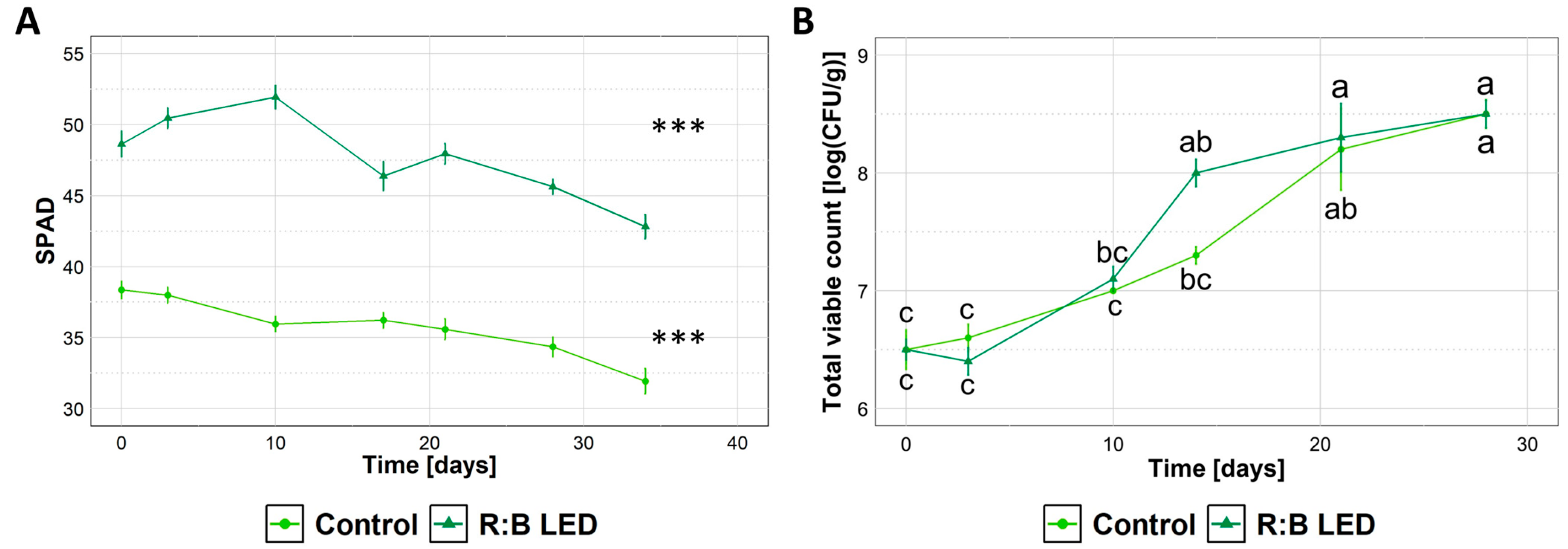
2.1. Plants Growth Evaluation
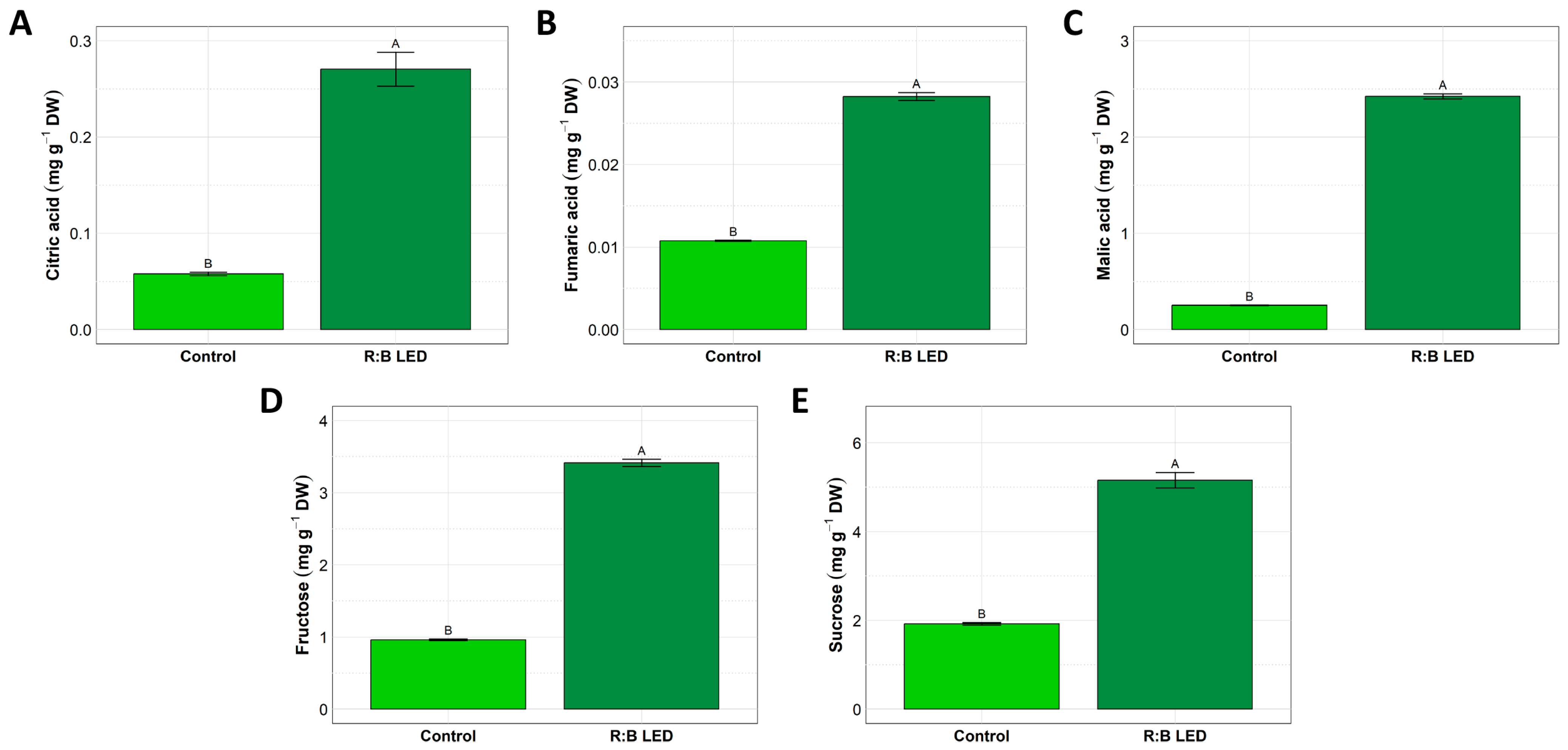
2.2. Organic Acids, Sugars, and Phenolic Compounds Concentrations
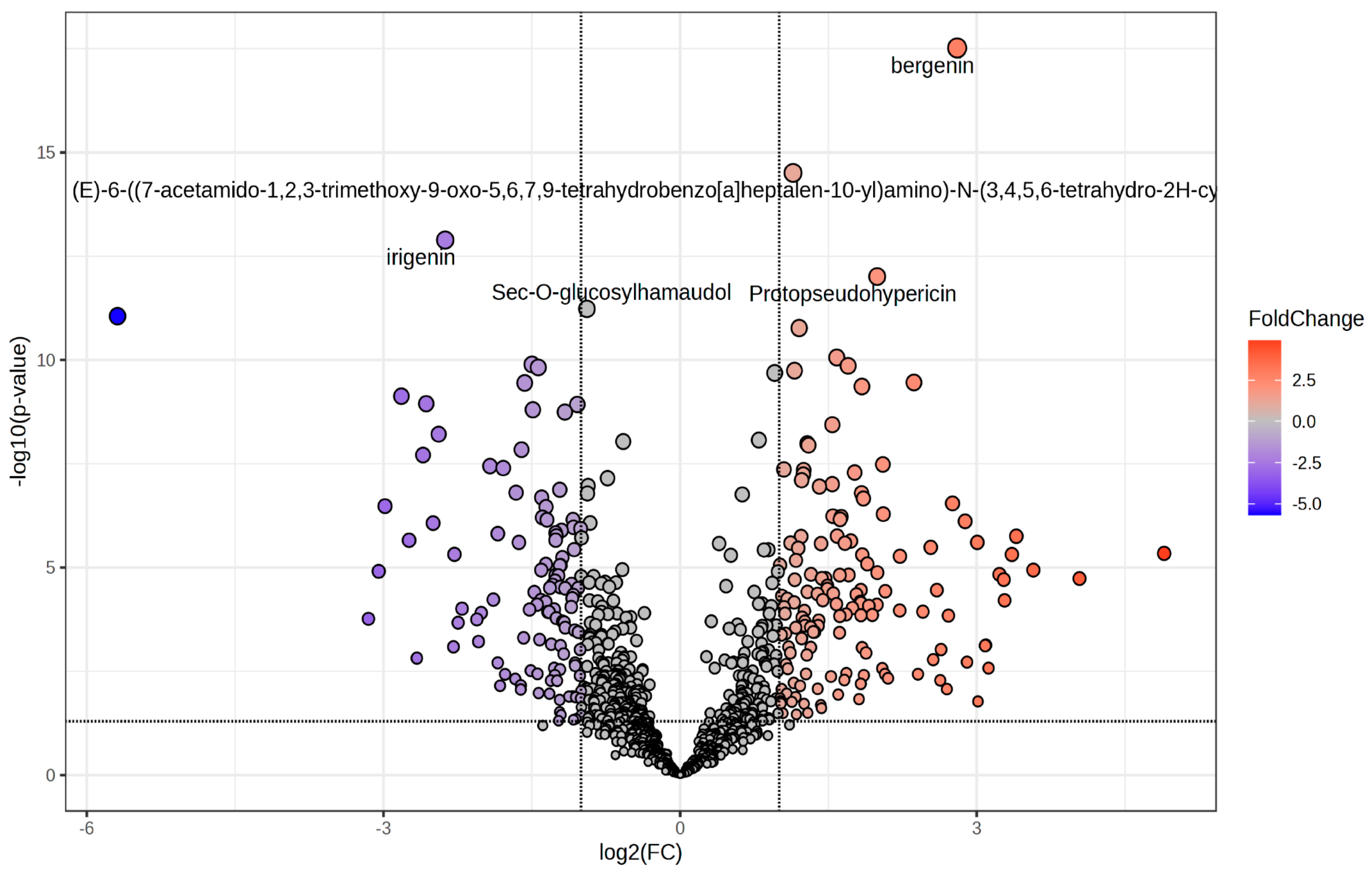
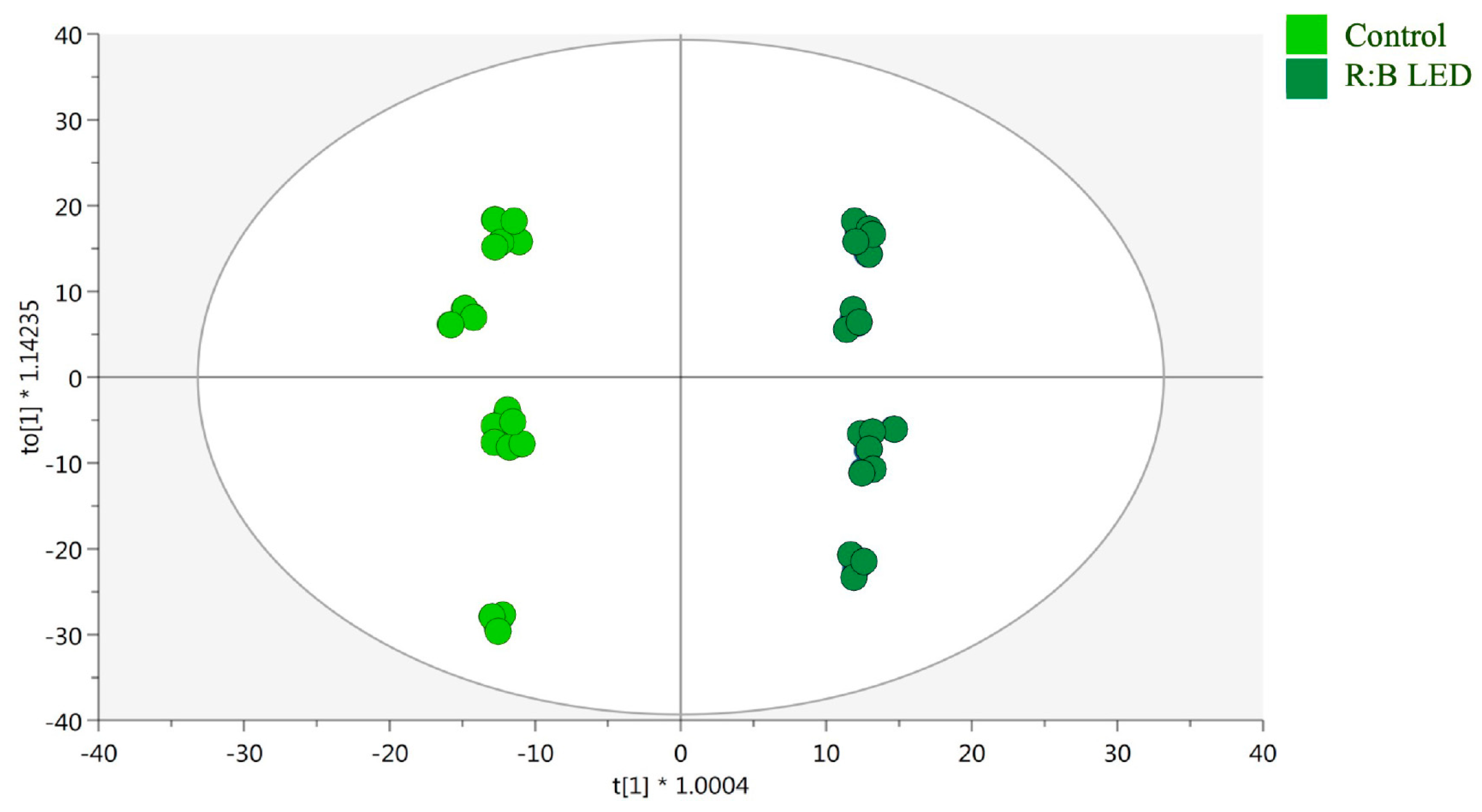
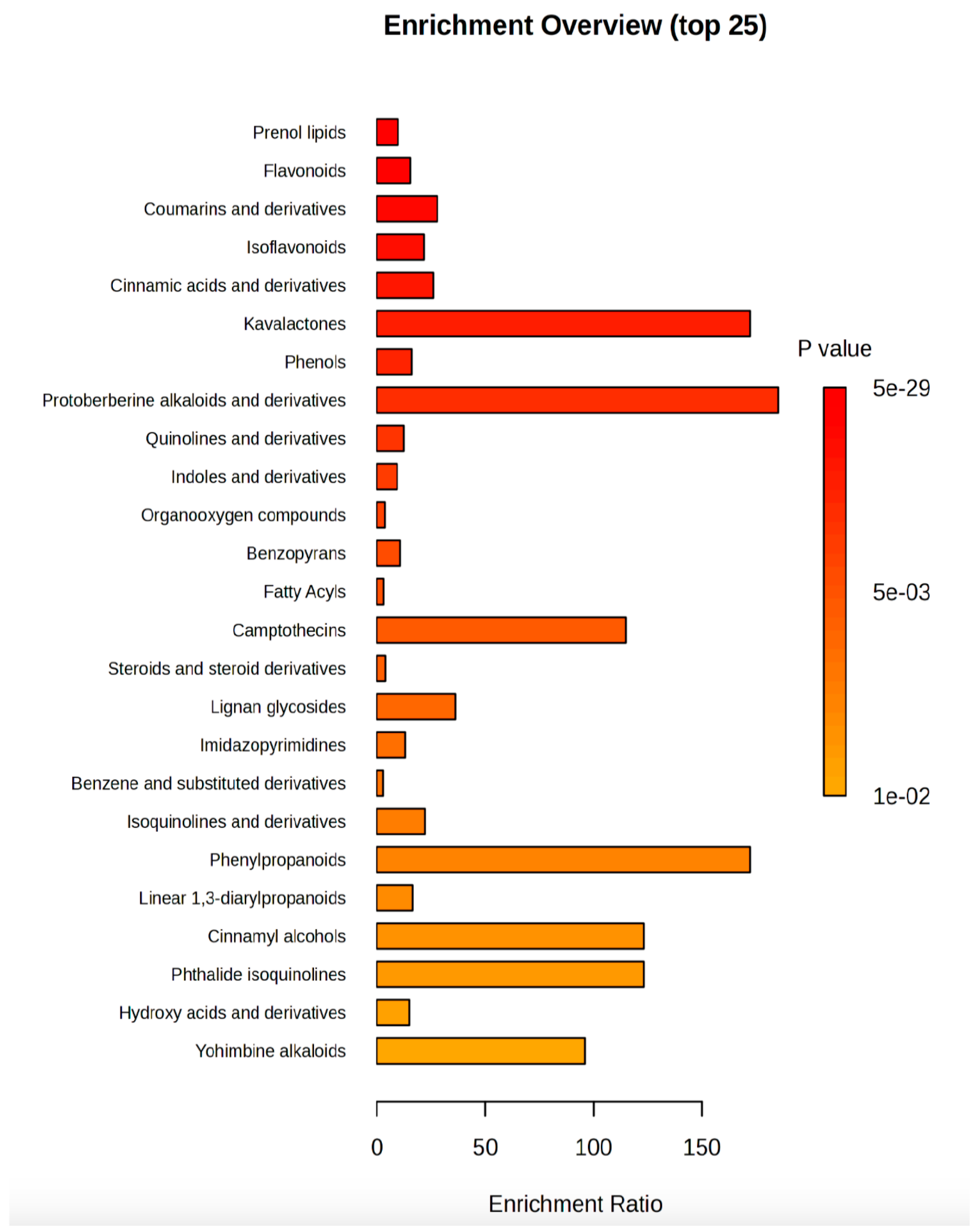
2.3. Untargeted Phenolic Compounds Profiling
2.4. Antioxidant Properties
2.5. Enzyme Inhibitory Activity
2.6. Quality Evolution of Ready-to-Eat Lamb’s Lettuce During Refrigerated Storage
3. Discussion
3.1. Plant Morphology and Growth Parameters
3.2. Content of Organic Acids, Sugars, and Phenolic Compounds
3.3. Profiling of Phenolic Compounds
3.4. Antioxidant Properties and Enzyme Inhibitory Activity
3.5. Post-Harvest Quality
3.6. Conclusions
4. Materials and Methods
4.1. Plant Growth Conditions
4.2. Plant Growth Evaluation
4.3. Preparation of Fresh-Cut Lamb’s Lettuce
4.4. Organic Acids and Sugars Content
4.5. Total Phenolic Compounds Content
4.6. Untargeted Phenolic Compounds Profiling by HRMS Metabolomics
4.7. Antioxidants and Enzyme Inhibitory Activities
4.8. Weight Loss and Firmness
4.9. Microbiological Analyses
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABTS | 2,2’-Azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) |
| ACAE | acarbose equivalents |
| ACH | air changes per hour |
| AChE | acetylcholinesterase |
| BChE | butyrylcholinesterase |
| BOPP | bioriented polypropylene |
| CUPRAC | Cupric Reducing Antioxidant Capacity |
| DPPH | 1,1-Diphenyl-2-picrylhydrazyl |
| EDTAE | EDTA equivalents |
| FRAP | Ferric Reducing Antioxidant Power |
| GAE | gallic acid equivalents |
| GALAE | galanthamine equivalents |
| HESI | heated electrospray ionization |
| HPLC | High-Performance Liquid Chromatography |
| KAE | kojic acid equivalents |
| LA | leaf area |
| LED | Light-Emitting Diode |
| MCA | metal-chelating activity |
| MNM | micronutrient malnutrition |
| MRD | maximum recovery diluent |
| PBD | phosphomolybdenum activity |
| PNPG | p-nitrophenyl-β-glucopyranoside |
| PPFD | photosynthetic photon flux density |
| R:B | red/blue |
| RH | relative humidity |
| SDW | shoot dry weight |
| SFW | shoot fresh weight |
| TE | Trolox equivalents |
| UHPLC-HRMS | ultra-high-performance liquid chromatography–high-resolution mass spectrometry |
| UV | ultraviolet |
References
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2023. Urbanization, Agrifood Systems Transformation and Healthy Diets Across the Rural–Urban Continuum; FAO: Rome, Italy, 2023; ISBN 978-92-5-137226-5. [Google Scholar]
- Kiani, A.K.; Dhuli, K.; Donato, K.; Aquilanti, B.; Velluti, V.; Matera, G.; Iaconelli, A.; Connelly, S.T.; Bellinato, F.; Gisondi, P.; et al. Main Nutritional Deficiencies. J. Prev. Med. Hyg. 2022, 63, E93–E101. [Google Scholar] [CrossRef] [PubMed]
- Yahia, Y.; Benabderrahim, M.A.; Tlili, N.; Bagues, M.; Nagaz, K. Bioactive Compounds, Antioxidant and Antimicrobial Activities of Extracts from Different Plant Parts of Two Ziziphus mill. Species. PLoS ONE 2020, 15, e0232599. [Google Scholar] [CrossRef] [PubMed]
- Sadeer, N.B.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The Versatility of Antioxidant Assays in Food Science and Safety—Chemistry, Applications, Strengths, and Limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef] [PubMed]
- Babbar, N.; Oberoi, H.S.; Sandhu, S.K.; Bhargav, V.K. Influence of Different Solvents in Extraction of Phenolic Compounds from Vegetable Residues and Their Evaluation as Natural Sources of Antioxidants. J. Food Sci. Technol. 2014, 51, 2568–2575. [Google Scholar] [CrossRef]
- Martínez, V.; Mitjans, M.; Vinardell, M.P. Cytoprotective Effects of Polyphenols against Oxidative Damage. Polyphen. Hum. Health Dis. 2014, 1, 275–288. [Google Scholar] [CrossRef]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The Role of Polyphenols in Abiotic Stress Response: The Influence of Molecular Structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Fortier, E.; Desjardins, Y.; Tremblay, N.; Bélec, C.; Côté, M. Influence of Irrigation and Nitrogen Fertilization on Broccoli Polyphenolics Concentration. Acta Hortic. 2010, 856, 55–62. [Google Scholar] [CrossRef]
- Takahashi, S.; Badger, M.R. Photoprotection in Plants: A New Light on Photosystem II Damage. Trends Plant Sci. 2011, 16, 53–60. [Google Scholar] [CrossRef]
- Klimek-Szczykutowicz, M.; Prokopiuk, B.; Dziurka, K.; Pawłowska, B.; Ekiert, H.; Szopa, A. The Influence of Different Wavelengths of LED Light on the Production of Glucosinolates and Phenolic Compounds and the Antioxidant Potential in in Vitro Cultures of Nasturtium officinale (Watercress). Plant Cell Tissue Organ Cult. 2022, 149, 113–122. [Google Scholar] [CrossRef]
- D’Souza, C.; Yuk, H.G.; Khoo, G.H.; Zhou, W. Application of Light-Emitting Diodes in Food Production, Postharvest Preservation, and Microbiological Food Safety. Compr. Rev. Food Sci. Food Saf. 2015, 14, 719–740. [Google Scholar] [CrossRef]
- Dutta Gupta, S.; Jatothu, B. Fundamentals and Applications of Light-Emitting Diodes (LEDs) in Vitro Plant Growth and Morphogenesis. Plant Biotechnol. Rep. 2013, 7, 211–220. [Google Scholar] [CrossRef]
- Livadariu, O.; Maximilian, C.; Rahmanifar, B.; Cornea, C.P. LED Technology Applied to Plant Development for Promoting the Accumulation of Bioactive Compounds: A Review. Plants 2023, 12, 1075. [Google Scholar] [CrossRef] [PubMed]
- Cope, K.R.; Snowden, M.C.; Bugbee, B. Photobiological Interactions of Blue Light and Photosynthetic Photon Flux: Effects of Monochromatic and Broad-Spectrum Light Sources. Photochem. Photobiol. 2014, 90, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xin, G.; Liu, C.; Shi, Q.; Yang, F.; Wei, M. Effects of Red and Blue Light on Leaf Anatomy, CO2 Assimilation and the Photosynthetic Electron Transport Capacity of Sweet Pepper (Capsicum annuum L.) Seedlings. BMC Plant Biol. 2020, 20, 318. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Sams, C.E.; Morrow, R.C. Blue Wavelengths from LED Lighting Increase Nutritionally Important Metabolites in Specialty Crops. HortScience 2015, 50, 1285–1288. [Google Scholar] [CrossRef]
- Lillo, C.; Appenroth, K.J. Light Regulation of Nitrate Reductase in Higher Plants: Which Photoreceptors Are Involved? Plant Biol. 2001, 3, 455–465. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue Light Dose–Responses of Leaf Photosynthesis, Morphology, and Chemical Composition of Cucumis sativus Grown under Different Combinations of Red and Blue Light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Ma, Y.; Xu, A.; Cheng, Z.M. (Max) Effects of Light Emitting Diode Lights on Plant Growth, Development and Traits a Meta-Analysis. Hortic. Plant J. 2021, 7, 552–564. [Google Scholar] [CrossRef]
- Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, M.O.K. Blue Light Added with Red LEDs Enhance Growth Characteristics, Pigments Content, and Antioxidant Capacity in Lettuce, Spinach, Kale, Basil, and Sweet Pepper in a Controlled Environment. Plants 2019, 8, 93. [Google Scholar] [CrossRef]
- Hosseini, A.; Zare Mehrjerdi, M.; Aliniaeifard, S. Alteration of Bioactive Compounds in Two Varieties of Basil (Ocimum basilicum) Grown under Different Light Spectra. J. Essent. Oil Bear. Plants 2018, 21, 913–923. [Google Scholar] [CrossRef]
- Piovene, C.; Orsini, F.; Bosi, S.; Sanoubar, R.; Bregola, V.; Dinelli, G.; Gianquinto, G. Optimal Red:Blue Ratio in Led Lighting for Nutraceutical Indoor Horticulture. Sci. Hortic. 2015, 193, 202–208. [Google Scholar] [CrossRef]
- Min, Q.; Marcelis, L.F.M.; Nicole, C.C.S.; Woltering, E.J. High Light Intensity Applied Shortly before Harvest Improves Lettuce Nutritional Quality and Extends the Shelf Life. Front. Plant Sci. 2021, 12, 615355. [Google Scholar] [CrossRef] [PubMed]
- Godínez-Mendoza, P.L.; Rico-Chávez, A.K.; Ferrusquía-Jimenez, N.I.; Carbajal-Valenzuela, I.A.; Villagómez-Aranda, A.L.; Torres-Pacheco, I.; Guevara-González, R.G. Plant Hormesis: Revising of the Concepts of Biostimulation, Elicitation and Their Application in a Sustainable Agricultural Production. Sci. Total Environ. 2023, 894, 164883. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Hernández, M.C.; Parola-Contreras, I.; Montoya-Gómez, L.M.; Torres-Pacheco, I.; Schwarz, D.; Guevara-González, R.G. Eustressors: Chemical and Physical Stress Factors Used to Enhance Vegetables Production. Sci. Hortic. 2019, 250, 223–229. [Google Scholar] [CrossRef]
- Johansson, M.; Jägerstad, M.; Frølich, W. Folates in Lettuce: A Pilot Study. Food Nutr. Res. 2007, 51, 22–30. [Google Scholar] [CrossRef]
- Długosz-Grochowska, O.; Kołton, A.; Wojciechowska, R. Modifying Folate and Polyphenol Concentrations in Lamb’s Lettuce by the Use of LED Supplemental Lighting during Cultivation in Greenhouses. J. Funct. Foods 2016, 26, 228–237. [Google Scholar] [CrossRef]
- Długosz-Grochowska, O.; Wojciechowska, R.; Kruczek, M.; Habela, A. Supplemental Lighting with LEDs Improves the Biochemical Composition of Two Valerianella locusta (L.) Cultivars. Hortic. Environ. Biotechnol. 2017, 58, 441–449. [Google Scholar] [CrossRef]
- Wojciechowska, R.; Kołton, A.; Długosz-Grochowska, O.; Knop, E. Nitrate Content in Valerianella locusta L. Plants Is Affected by Supplemental LED Lighting. Sci. Hortic. 2016, 211, 179–186. [Google Scholar] [CrossRef]
- Wojciechowska, R.; Kurpaska, S.; Malinowski, M.; Sikora, J.; Krakowiak-Bal, A.; Długosz-Grochowska, O. Effect of Supplemental LED Lighting on Growth and Quality of Valerianella locusta L. and Economic Aspects of Cultivation in Autumn Cycle. Acta Sci. Pol. Hortorum Cultus 2016, 15, 233–244. [Google Scholar]
- Manzocco, L.; Foschia, M.; Tomasi, N.; Maifreni, M.; Dalla Costa, L.; Marino, M.; Cortella, G.; Cesco, S. Influence of Hydroponic and Soil Cultivation on Quality and Shelf Life of Ready-to-Eat Lamb’s Lettuce (Valerianella locusta L. Laterr). J. Sci. Food Agric. 2011, 91, 1373–1380. [Google Scholar] [CrossRef]
- Mirzahosseini, Z.; Shabani, L.; Sabzalian, M.R. LED Lights Increase an Antioxidant Capacity of Arabidopsis thaliana under Wound-Induced Stresses. Funct. Plant Biol. 2020, 47, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Monostori, I.; Heilmann, M.; Kocsy, G.; Rakszegi, M.; Ahres, M.; Altenbach, S.B.; Szalai, G.; Pál, M.; Toldi, D.; Simon-Sarkadi, L.; et al. LED Lighting—Modification of Growth, Metabolism, Yield and Flour Composition in Wheat by Spectral Quality and Intensity. Front. Plant Sci. 2018, 9, 605. [Google Scholar] [CrossRef] [PubMed]
- Folta, K.M.; Childers, K.S. Light as a Growth Regulator: Controlling Plant Biology with Narrow-Bandwidth Solid-State Lighting Systems. HortScience Horts 2008, 43, 1957–1964. [Google Scholar] [CrossRef]
- Parks, B.M.; Hoecker, U.; Spalding, E.P. Light-Induced Growth Promotion by SPA1 Counteracts Phytochrome-Mediated Growth Inhibition during de-Etiolation. Plant Physiol. 2001, 126, 1291–1298. [Google Scholar] [CrossRef]
- Edesi, J.; Kotkas, K.; Pirttilä, A.M.; Häggman, H. Does Light Spectral Quality Affect Survival and Regeneration of Potato (Solanum tuberosum L.) Shoot Tips after Cryopreservation? Plant Cell Tissue Organ Cult. (PCTOC) 2014, 119, 599–607. [Google Scholar] [CrossRef]
- Lian, M.-L.; Murthy, H.N.; Paek, K.-Y. Effects of Light Emitting Diodes (LEDs) on the in Vitro Induction and Growth of Bulblets of Lilium Oriental Hybrid ‘Pesaro.’. Sci. Hortic. 2002, 94, 365–370. [Google Scholar] [CrossRef]
- Poudel, P.R.; Kataoka, I.; Mochioka, R. Effect of Red- and Blue-Light-Emitting Diodes on Growth and Morphogenesis of Grapes. Plant Cell Tissue Organ Cult. 2008, 92, 147–153. [Google Scholar] [CrossRef]
- Wu, H.-C.; Lin, C.-C. Red Light-Emitting Diode Light Irradiation Improves Root and Leaf Formation in Difficult-to-Propagate Protea cynaroides L. Plantlets in Vitro. HortScience Horts 2012, 47, 1490–1494. [Google Scholar] [CrossRef]
- Hung, C.D.; Hong, C.-H.; Kim, S.-K.; Lee, K.-H.; Park, J.-Y.; Nam, M.-W.; Choi, D.-H.; Lee, H.-I. LED Light for in Vitro and Ex Vitro Efficient Growth of Economically Important Highbush Blueberry (Vaccinium corymbosum L.). Acta Physiol. Plant. 2016, 38, 152. [Google Scholar] [CrossRef]
- Silva, M.M.A.; de Oliveira, A.L.B.; Oliveira-Filho, R.A.; Gouveia-Neto, A.S.; Camara, T.J.R.; Willadino, L.G. Effect of Blue/Red LED Light Combination on Growth and Morphogenesis of Saccharum Officinarum Plantlets in Vitro. In Proceedings of the Proceedings Volume 8947, Imaging, Manipulation, and Analysis of Biomolecules, Cells, and Tissues XII, San Francisco, CA, USA, 3–6 February 2014; p. 89471X. [Google Scholar]
- Saradhuldhat, P.; Paull, R.E. Pineapple Organic Acid Metabolism and Accumulation during Fruit Development. Sci. Hortic. 2007, 112, 297–303. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Eprintsev, A.T. Organic Acids: The Pools of Fixed Carbon Involved in Redox Regulation and Energy Balance in Higher Plants. Front. Plant Sci. 2016, 7, 1042. [Google Scholar] [CrossRef] [PubMed]
- Vallarino, J.G.; Osorio, S. Organic Acids. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E.M., Ed.; Woodhead Publishing: Sawston, UK, 2019; pp. 207–224. ISBN 978-0-12-813278-4. [Google Scholar]
- Annunziata, M.G.; Apelt, F.; Carillo, P.; Krause, U.; Feil, R.; Mengin, V.; Lauxmann, M.A.; Köhl, K.; Nikoloski, Z.; Stitt, M.; et al. Getting Back to Nature: A Reality Check for Experiments in Controlled Environments. J. Exp. Bot. 2017, 68, 4463–4477. [Google Scholar] [CrossRef] [PubMed]
- Cioć, M.; Tokarz, K.; Dziurka, M.; Pawłowska, B. Energy-Saving LED Light Affects the Efficiency of the Photosynthetic Apparatus and Carbohydrate Content in Gerbera Jamesonii Bolus Ex Hook. f. Axillary Shoots Multiplied in Vitro. Biology 2021, 10, 1035. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xin, G.; Wei, M.; Shi, Q.; Yang, F.; Wang, X. Carbohydrate Accumulation and Sucrose Metabolism Responses in Tomato Seedling Leaves When Subjected to Different Light Qualities. Sci. Hortic. 2017, 225, 490–497. [Google Scholar] [CrossRef]
- Shi, L.; Cao, S.; Shao, J.; Chen, W.; Yang, Z.; Zheng, Y. Chinese Bayberry Fruit Treated with Blue Light after Harvest Exhibit Enhanced Sugar Production and Expression of Cryptochrome Genes. Postharvest Biol. Technol. 2016, 111, 197–204. [Google Scholar] [CrossRef]
- Li, H.; Tang, C.; Xu, Z.; Liu, X.; Han, X. Effects of Different Light Sources on the Growth of Non-Heading Chinese Cabbage (Brassica campestris L.). J. Agric. Sci. 2012, 4, 262–273. [Google Scholar] [CrossRef]
- Chen, X.; Yang, Q. Effects of Intermittent Light Exposure with Red and Blue Light Emitting Diodes on Growth and Carbohydrate Accumulation of Lettuce. Sci. Hortic. 2018, 234, 220–226. [Google Scholar] [CrossRef]
- Wang, S.; Fang, H.; Xie, J.; Wu, Y.; Tang, Z.; Liu, Z.; Lv, J.; Yu, J. Physiological Responses of Cucumber Seedlings to Different Supplemental Light Duration of Red and Blue LED. Front. Plant Sci. 2021, 12, 709313. [Google Scholar] [CrossRef]
- Al Murad, M.; Razi, K.; Jeong, B.R.; Samy, P.M.A.; Muneer, S. Light Emitting Diodes (LEDs) as Agricultural Lighting: Impact and Its Potential on Improving Physiology, Flowering, and Secondary Metabolites of Crops. Sustainability 2021, 13, 1985. [Google Scholar] [CrossRef]
- Qian, H.; Liu, T.; Deng, M.; Miao, H.; Cai, C.; Shen, W.; Wang, Q. Effects of Light Quality on Main Health-Promoting Compounds and Antioxidant Capacity of Chinese Kale Sprouts. Food Chem. 2016, 196, 1232–1238. [Google Scholar] [CrossRef]
- Huang, J.; Xu, Y.; Duan, F.; Du, X.; Yang, Q.; Zheng, Y. Improvement of the Growth and Nutritional Quality of Two-Leaf-Color Pak Choi by Supplemental Alternating Red and Blue Light. HortScience Horts 2021, 56, 118–125. [Google Scholar] [CrossRef]
- Doukani, K.; Selles, A.S.M.; Bouhenni, H. Chapter 3.1.11—Hypericin and Pseudohypericin. In Naturally Occurring Chemicals Against Alzheimer’s Disease; Belwal, T., Nabavi, S.M., Nabavi, S.F., Dehpour, A.R., Shirooie, S., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 155–165. ISBN 978-0-12-819212-2. [Google Scholar]
- Samuolienė, G.; Brazaitytė, A.; Jankauskienė, J.; Viršilė, A.; Sirtautas, R.; Novičkovas, A.; Sakalauskienė, S.; Sakalauskaitė, J.; Duchovskis, P. LED Irradiance Level Affects Growth and Nutritional Quality of Brassica Microgreens. Cent. Eur. J. Biol. 2013, 8, 1241–1249. [Google Scholar] [CrossRef]
- Pennisi, G.; Blasioli, S.; Cellini, A.; Maia, L.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C.; et al. Unraveling the Role of Red:Blue LED Lights on Resource Use Efficiency and Nutritional Properties of Indoor Grown Sweet Basil. Front. Plant Sci. 2019, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Huyskens-Keil, S.; Eichholz-Dündar, I.; Hassenberg, K.; Herppich, W.B. Impact of Light Quality (White, Red, Blue Light and UV-C Irradiation) on Changes in Anthocyanin Content and Dynamics of PAL and POD Activities in Apical and Basal Spear Sections of White Asparagus after Harvest. Postharvest Biol. Technol. 2020, 161, 111069. [Google Scholar] [CrossRef]
- Paik, I.; Huq, E. Plant Photoreceptors: Multi-Functional Sensory Proteins and Their Signaling Networks. Semin. Cell Dev. Biol. 2019, 92, 114–121. [Google Scholar] [CrossRef]
- Patel, D.K. Medicinal Importance, Pharmacological Activities, and Analytical Aspects of Engeletin in Medicine: Therapeutic Benefit Through Scientific Data Analysis. Endocr. Metab. Immune Disord. Drug Targets 2023, 23, 273–282. [Google Scholar] [CrossRef]
- Chen, A.Y.; Chen, Y.C. A Review of the Dietary Flavonoid, Kaempferol on Human Health and Cancer Chemoprevention. Food Chem. 2013, 138, 2099–2107. [Google Scholar] [CrossRef]
- Wang, S.; Wang, N.; Huang, X.; Yang, B.; Zheng, Y.; Zhang, J.; Wang, X.; Lin, Y.; Wang, Z. Baohuoside i Suppresses Breast Cancer Metastasis by Downregulating the Tumor-Associated Macrophages/C-X-C Motif Chemokine Ligand 1 Pathway. Phytomedicine 2020, 78, 153331. [Google Scholar] [CrossRef]
- Liu, Y.; Singh, S.K.; Pattanaik, S.; Wang, H.; Yuan, L. Light Regulation of the Biosynthesis of Phenolics, Terpenoids, and Alkaloids in Plants. Commun. Biol. 2023, 6, 1055. [Google Scholar] [CrossRef]
- Rajput, A.; Sharma, R.; Bharti, R. Pharmacological Activities and Toxicities of Alkaloids on Human Health. Mater. Today Proc. 2022, 48, 1407–1415. [Google Scholar] [CrossRef]
- Rihan, H.Z.; Aldarkazali, M.; Mohamed, S.J.; McMulkin, N.B.; Jbara, M.H.; Fuller, M.P. A Novel New Light Recipe Significantly Increases the Growth and Yield of Sweet Basil (Ocimum basilicum) Grown in a Plant Factory System. Agronomy 2020, 10, 934. [Google Scholar] [CrossRef]
- Ghaffari, Z.; Rahimmalek, M.; Sabzalian, M.R. Variation in the Primary and Secondary Metabolites Derived from the Isoprenoid Pathway in the Perovskia Species in Response to Different Wavelengths Generated by Light Emitting Diodes (LEDs). Ind. Crops Prod. 2019, 140, 111592. [Google Scholar] [CrossRef]
- Khurshid, R.; Ullah, M.A.; Tungmunnithum, D.; Drouet, S.; Shah, M.; Zaeem, A.; Hameed, S.; Hano, C.; Abbasi, B.H. Lights Triggered Differential Accumulation of Antioxidant and Antidiabetic Secondary Metabolites in Callus Culture of Eclipta alba L. PLoS ONE 2020, 15, e0233963. [Google Scholar] [CrossRef] [PubMed]
- Samuolienė, G.; Urbonavičiūtė, A.; Brazaitytė, A.; Šabajevienė, G.; Sakalauskaitė, J.; Duchovskis, P. The Impact of LED Illumination on Antioxidant Properties of Sprouted Seeds. Cent. Eur. J. Biol. 2011, 6, 68–74. [Google Scholar] [CrossRef]
- Liu, Y.; Roof, S.; Ye, Z.; Barry, C.; Van Tuinent, A.; Vrebalov, J.; Bowler, C.; Giovannoni, J. Manipulation of Light Signal Transduction as a Means of Modifying Fruit Nutritional Quality in Tomato. Proc. Natl. Acad. Sci. USA 2004, 101, 9897–9902. [Google Scholar] [CrossRef]
- Rauf, A.; Jehan, N. Natural Products as a Potential Enzyme Inhibitors from Medicinal Plants. In Enzyme Inhibitors and Activators; Senturk, M., Ed.; IntechOpen: Rijeka, Croatia, 2017. [Google Scholar]
- Lankatillake, C.; Luo, S.; Flavel, M.; Lenon, G.B.; Gill, H.; Huynh, T.; Dias, D.A. Screening Natural Product Extracts for Potential Enzyme Inhibitors: Protocols, and the Standardisation of the Usage of Blanks in α-Amylase, α-Glucosidase and Lipase Assays. Plant Methods 2021, 17, 3. [Google Scholar] [CrossRef]
- Fontana Pereira, D.; Cazarolli, L.H.; Lavado, C.; Mengatto, V.; Figueiredo, M.S.R.B.; Guedes, A.; Pizzolatti, M.G.; Silva, F.R.M.B. Effects of Flavonoids on α-Glucosidase Activity: Potential Targets for Glucose Homeostasis. Nutrition 2011, 27, 1161–1167. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, G.; Liao, Y.; Gong, D. Inhibitory Kinetics and Mechanism of Kaempferol on α-Glucosidase. Food Chem. 2016, 190, 207–215. [Google Scholar] [CrossRef]
- Adhikari, B. Roles of Alkaloids from Medicinal Plants in the Management of Diabetes Mellitus. J. Chem. 2021, 2021, 2691525. [Google Scholar] [CrossRef]
- Santos, M.I.; Grácio, M.; Silva, M.C.; Pedroso, L.; Lima, A. One Health Perspectives on Food Safety in Minimally Processed Vegetables and Fruits: From Farm to Fork. Microorganisms 2023, 11, 2990. [Google Scholar] [CrossRef]
- USDA. Fresh-Cut Produce Shipping Point and Market Inspection Instructions; USDA: Washington, DC, USA, 2016.
- Easlon, H.M.; Bloom, A.J. Easy Leaf Area: Automated Digital Image Analysis for Rapid and Accurate Measurement of Leaf Area. Appl. Plant Sci. 2014, 2, 1400033. [Google Scholar] [CrossRef] [PubMed]
- Pii, Y.; Graf, H.; Valentinuzzi, F.; Cesco, S.; Mimmo, T. The Effects of Plant Growth-Promoting Rhizobacteria (PGPR) on the Growth and Quality of Strawberries. Acta Hortic. 2018, 1217, 231–238. [Google Scholar] [CrossRef]
- Valentinuzzi, F.; Pii, Y.; Mimmo, T.; Savini, G.; Curzel, S.; Cesco, S. Fertilization Strategies as a Tool to Modify the Organoleptic Properties of Raspberry (Rubus idaeus L.) Fruits. Sci. Hortic. 2018, 240, 205–212. [Google Scholar] [CrossRef]
- Uysal, S.; Zengin, G.; Locatelli, M.; Bahadori, M.B.; Mocan, A.; Bellagamba, G.; Luca, E.D.; Mollica, A.; Aktumsek, A. Cytotoxic and Enzyme Inhibitory Potential of Two Potentilla Species (P. speciosa L. and P. reptans Willd.) and Their Chemical Composition. Front. Pharmacol. 2017, 8, 249316. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.; Ikeda, K.; Kanazawa, M.; VanderGheynst, J.; Fiehn, O.; Arita, M. MS-DIAL: Data-Independent MS/MS Deconvolution for Comprehensive Metabolome Analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The Role of Reporting Standards for Metabolite Annotation and Identification in Metabolomic Studies. GigaScience 2013, 2, 2047-2217X-2–13. [Google Scholar] [CrossRef]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies—Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef]
- Grochowski, D.M.; Uysal, S.; Aktumsek, A.; Granica, S.; Zengin, G.; Ceylan, R.; Locatelli, M.; Tomczyk, M. In Vitro Enzyme Inhibitory Properties, Antioxidant Activities, and Phytochemical Profile of Potentilla thuringiaca. Phytochem. Lett. 2017, 20, 365–372. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | ||
|---|---|---|
| Control | R:B LED | |
| Total Phenolic compounds (mg GAE g−1) | 33.61 ± 0.91 b | 43.77 ± 1.24 a |
| DPPH (mg TE g−1) | 62.29 ± 1.54 b | 69.71 ± 0.22 a |
| ABTS (mg TE g−1) | 104.34 ± 5.10 b | 148.81 ± 9.82 a |
| CUPRAC (mg TE g−1) | 120.59 ± 8.26 b | 170.76 ± 6.16 a |
| FRAP (mg TE g−1) | 78.73 ± 2.69 b | 114.04 ± 5.86 a |
| PBD (mmol TE g−1) | 1.79 ± 0.07 b | 2.11 ± 0.12 a |
| MCA (mg EDTAE g−1) | 25.56 ± 0.85 b | 28.72 ± 0.96 a |
| Treatment | AChE (mg GALAE g−1) | BChE (mg GALAE g−1) | Tyrosinase (mg KAE g−1) | α-amylase (mmol ACAE g−1) | α-glucosidase (mmol ACAE g−1) |
|---|---|---|---|---|---|
| Control | 1.77 ± 0.19 a | 0.44 ± 0.20 a | 55.44 ± 4.36 a | 0.29 ± 0.01 a | 0.08 ± 0.05 b |
| R:B LED | 1.99 ± 0.11 a | 0.51 ± 0.12 a | 53.04 ± 3.32 a | 0.28 ± 0.01 a | 0.32 ± 0.08 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monterisi, S.; Rebollo Vicioso, C.; Alzate Zuluaga, M.Y.; Melchior, S.; Senizza, B.; Zengin, G.; Fattorini, R.; Lanza, U.; de Oliveira Caretta, T.; Manzocco, L.; et al. Optimizing Valerianella locusta L. Growth and Metabolism by Combining Red and Blue LED Light: Insights into Plant Physiology, Biochemistry, and Nutraceutical Value. Plants 2025, 14, 1887. https://doi.org/10.3390/plants14121887
Monterisi S, Rebollo Vicioso C, Alzate Zuluaga MY, Melchior S, Senizza B, Zengin G, Fattorini R, Lanza U, de Oliveira Caretta T, Manzocco L, et al. Optimizing Valerianella locusta L. Growth and Metabolism by Combining Red and Blue LED Light: Insights into Plant Physiology, Biochemistry, and Nutraceutical Value. Plants. 2025; 14(12):1887. https://doi.org/10.3390/plants14121887
Chicago/Turabian StyleMonterisi, Sonia, Carmen Rebollo Vicioso, Monica Yorlady Alzate Zuluaga, Sofia Melchior, Biancamaria Senizza, Gokhan Zengin, Roberto Fattorini, Umberto Lanza, Talita de Oliveira Caretta, Lara Manzocco, and et al. 2025. "Optimizing Valerianella locusta L. Growth and Metabolism by Combining Red and Blue LED Light: Insights into Plant Physiology, Biochemistry, and Nutraceutical Value" Plants 14, no. 12: 1887. https://doi.org/10.3390/plants14121887
APA StyleMonterisi, S., Rebollo Vicioso, C., Alzate Zuluaga, M. Y., Melchior, S., Senizza, B., Zengin, G., Fattorini, R., Lanza, U., de Oliveira Caretta, T., Manzocco, L., Lucini, L., Cesco, S., & Pii, Y. (2025). Optimizing Valerianella locusta L. Growth and Metabolism by Combining Red and Blue LED Light: Insights into Plant Physiology, Biochemistry, and Nutraceutical Value. Plants, 14(12), 1887. https://doi.org/10.3390/plants14121887