


Alfalfa Responses to Intensive Soil Compaction: Effects on Plant and Root Growth, Phytohormones and Internal Gene Expression
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Preparation
2.2. Plant Growth
2.3. Sampling and Morphological and Physiological Analysis
2.4. RNA Extraction, Transcriptome Sequencing and Quantitative Real-Time PCR (qRT-PCR) Analysis
2.5. Bioinformatics Analysis
2.6. Root Sectioning and Microscopy
2.7. Extraction and Assay of Phytohormones
2.8. Statistical Analysis
3. Results
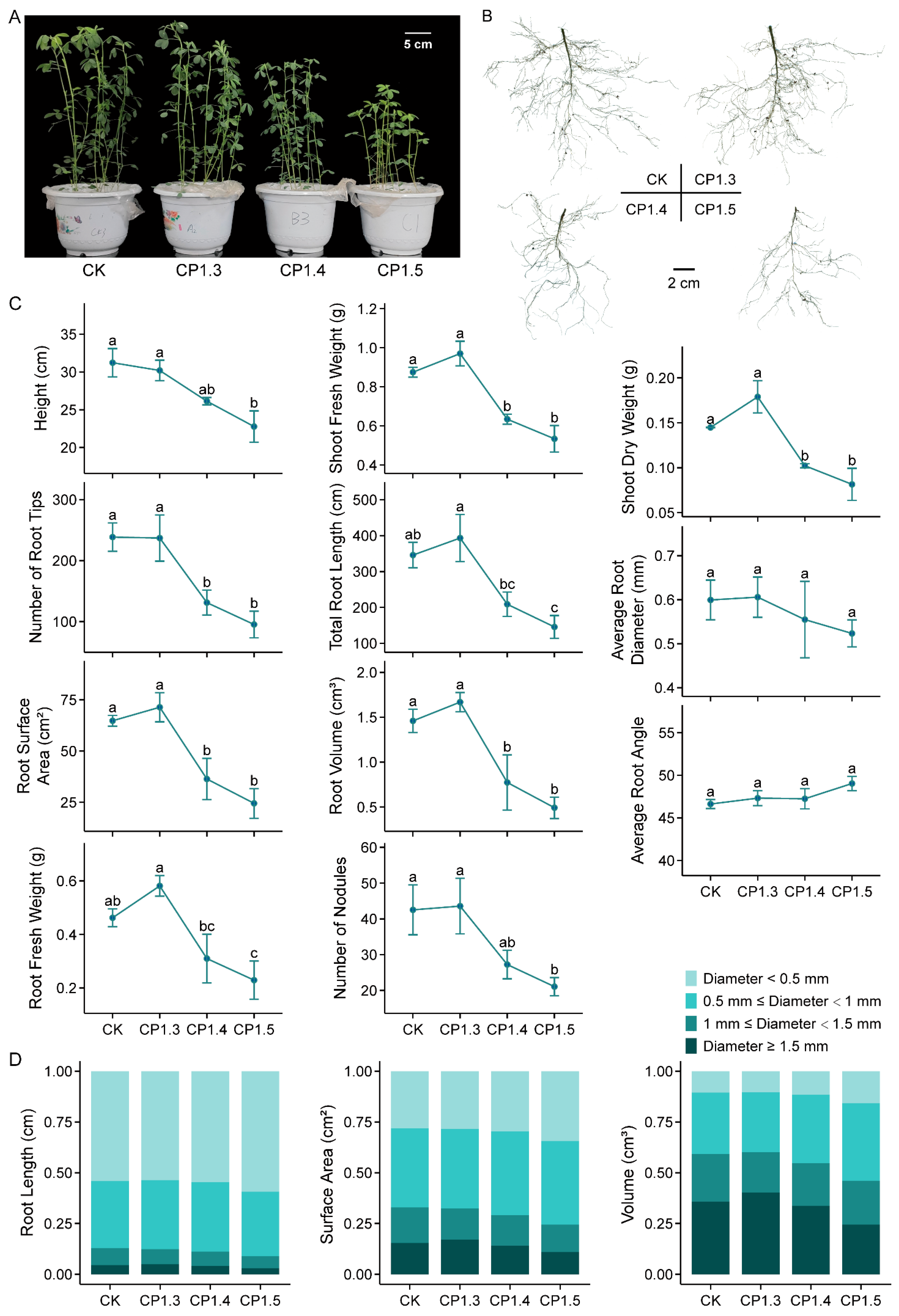
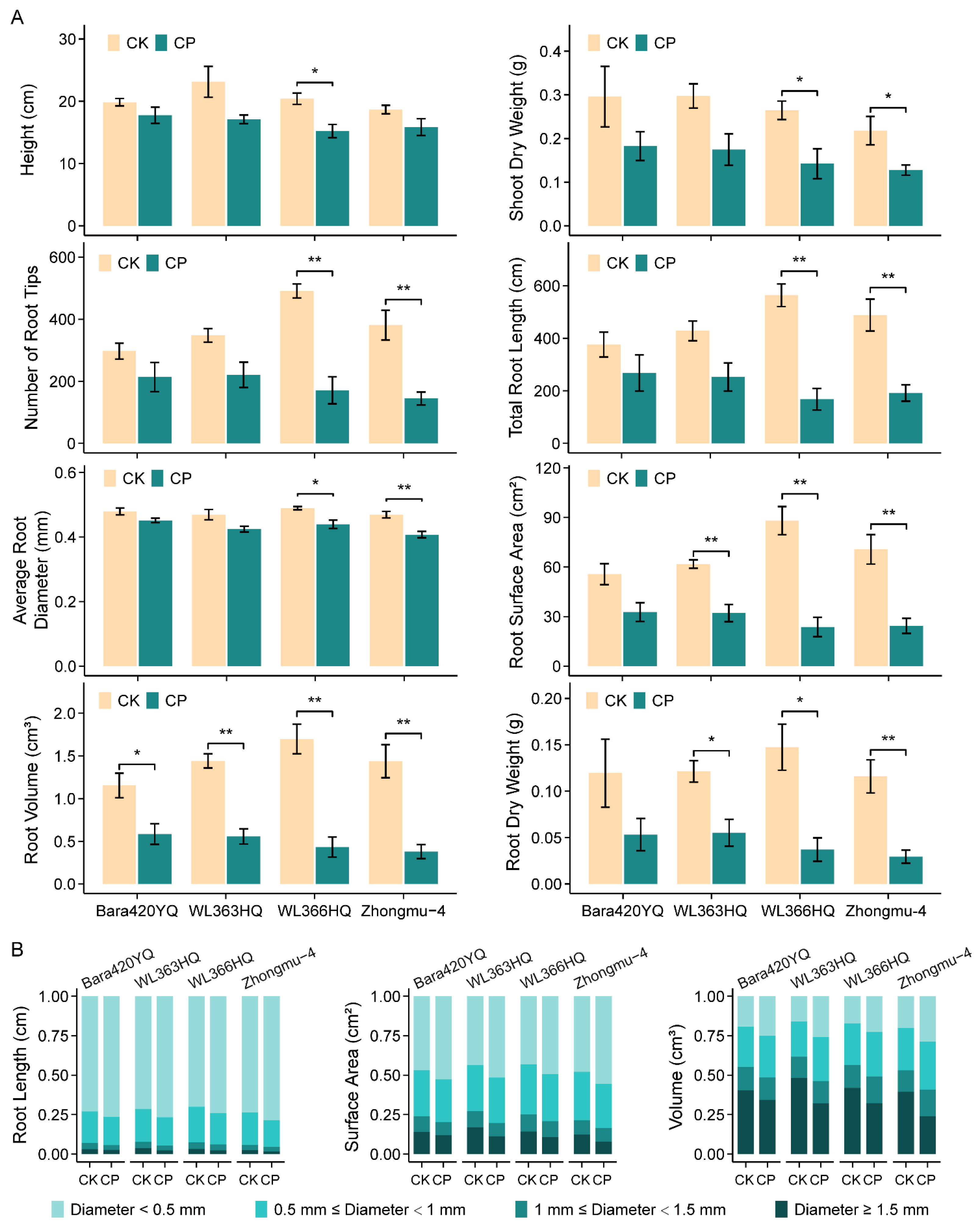
3.1. Effects of Different Levels of Soil Compaction on Alfalfa Growth
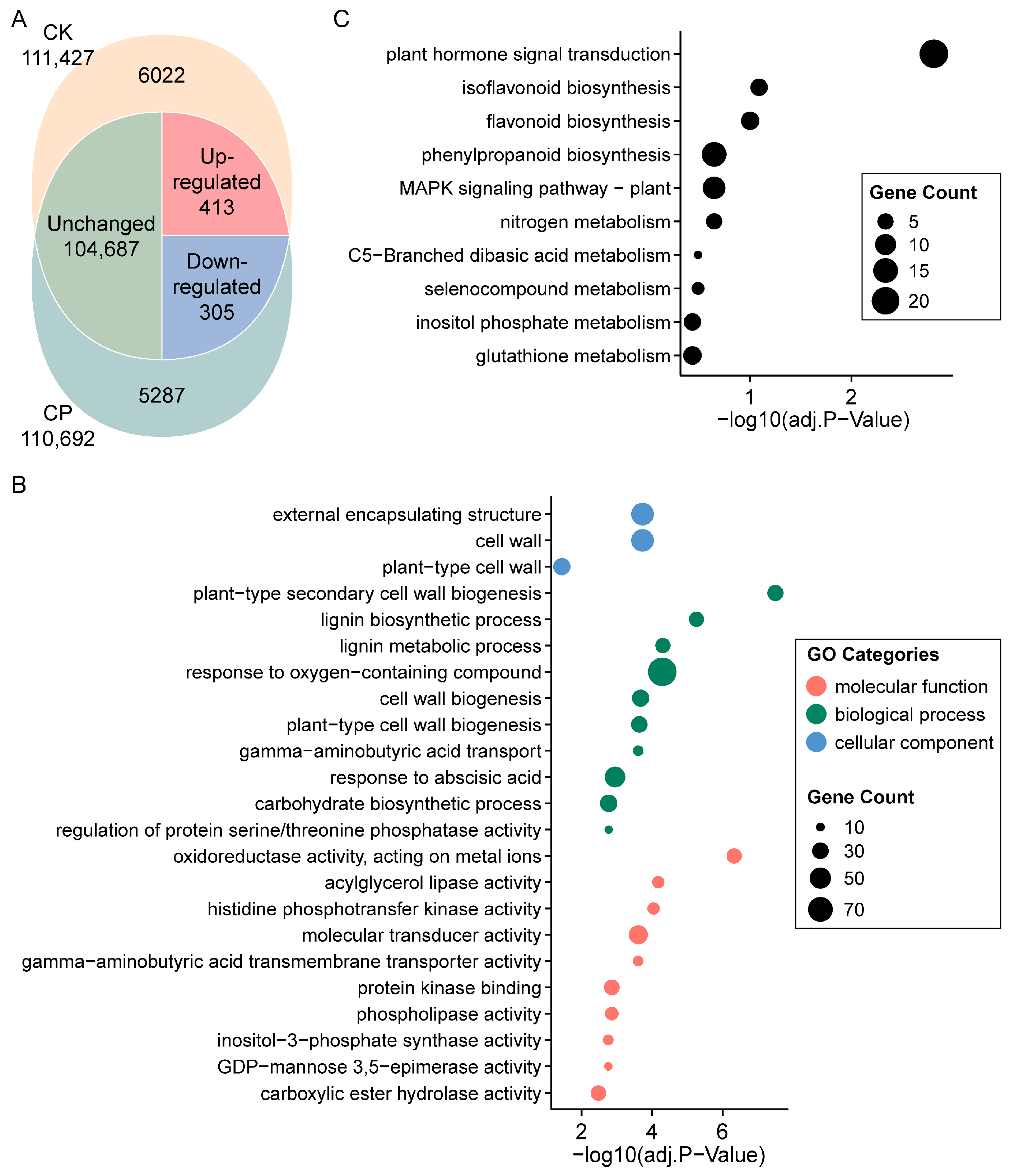
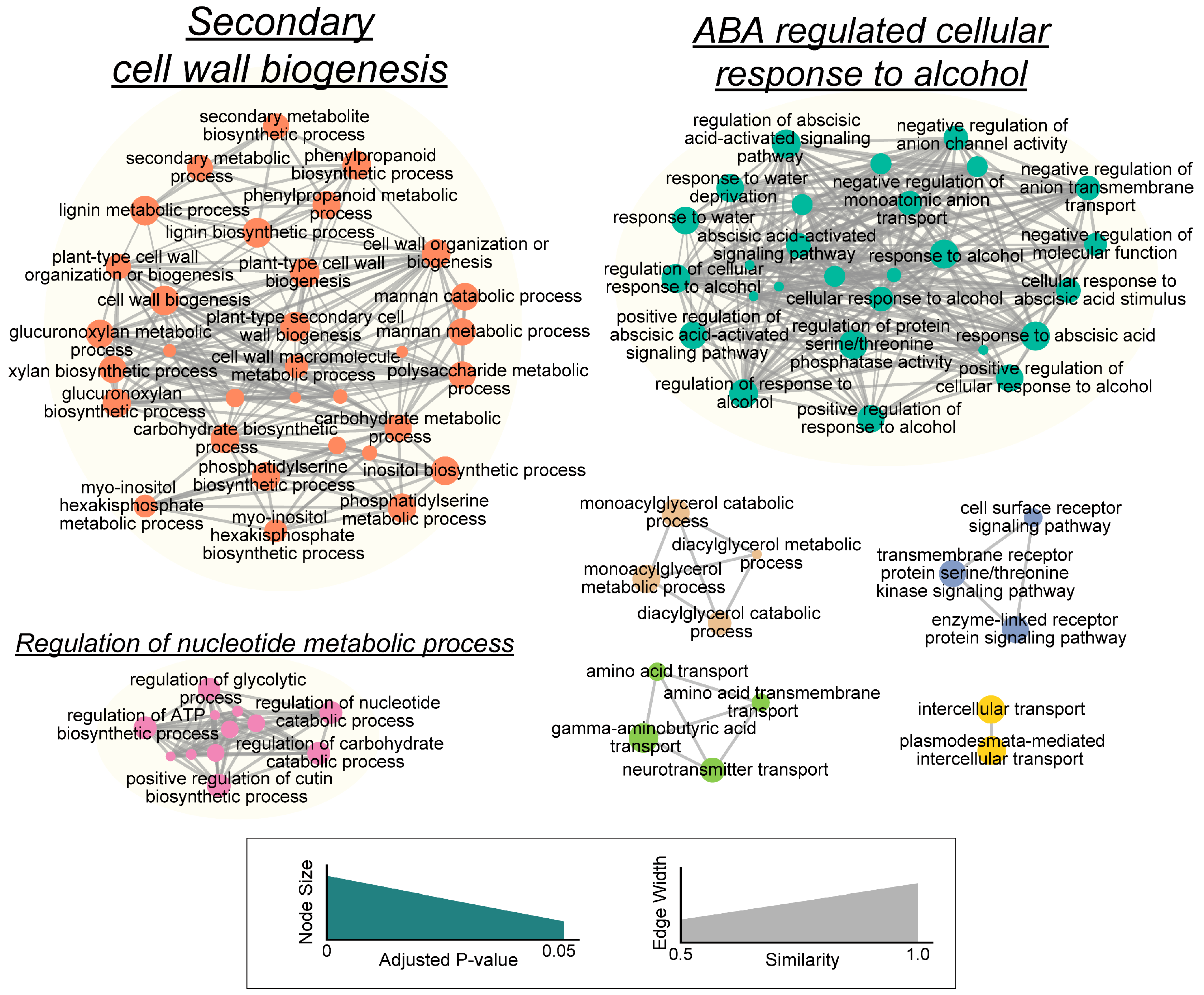
3.2. Transcriptome Profile of Alfalfa Root Response to Soil Compaction
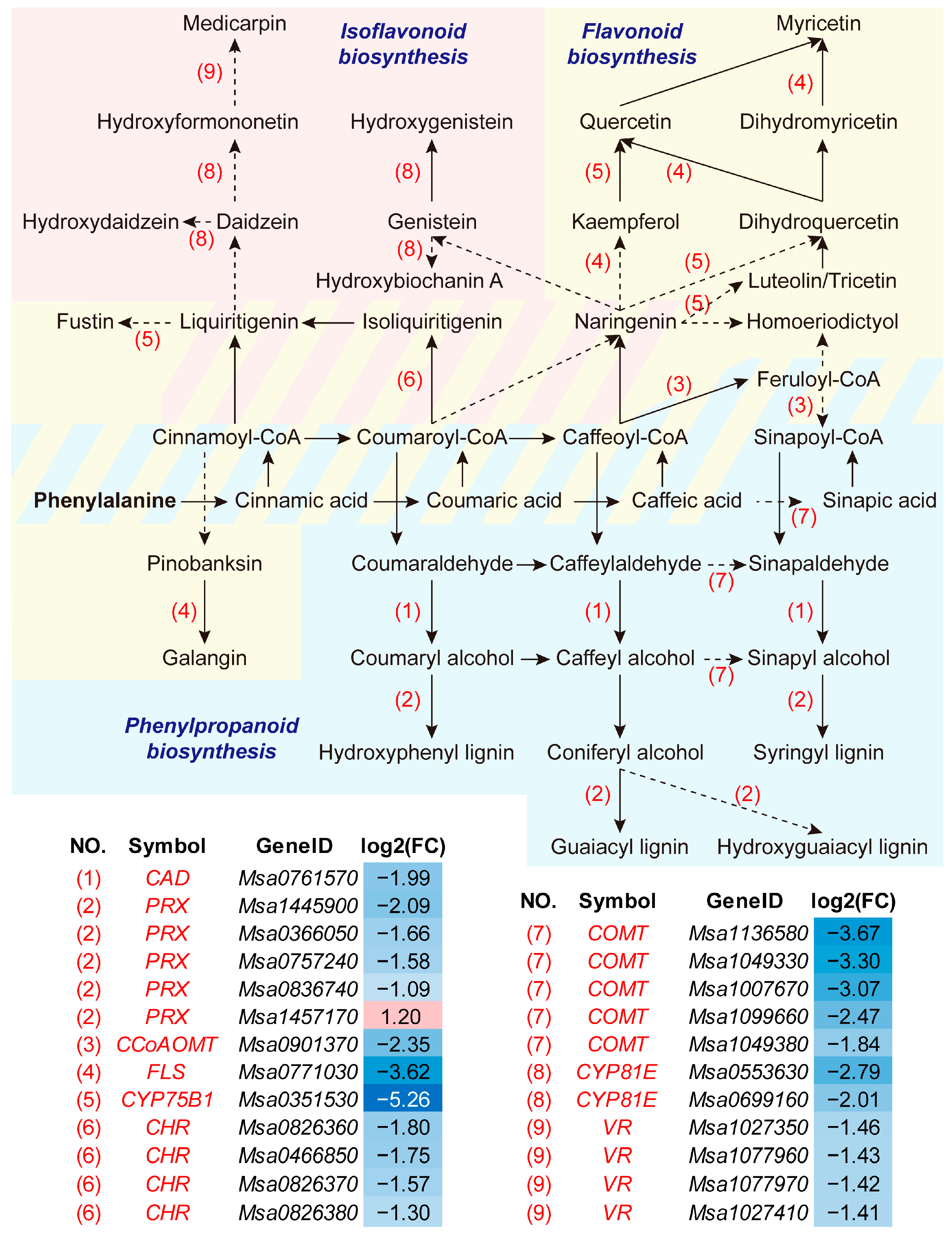
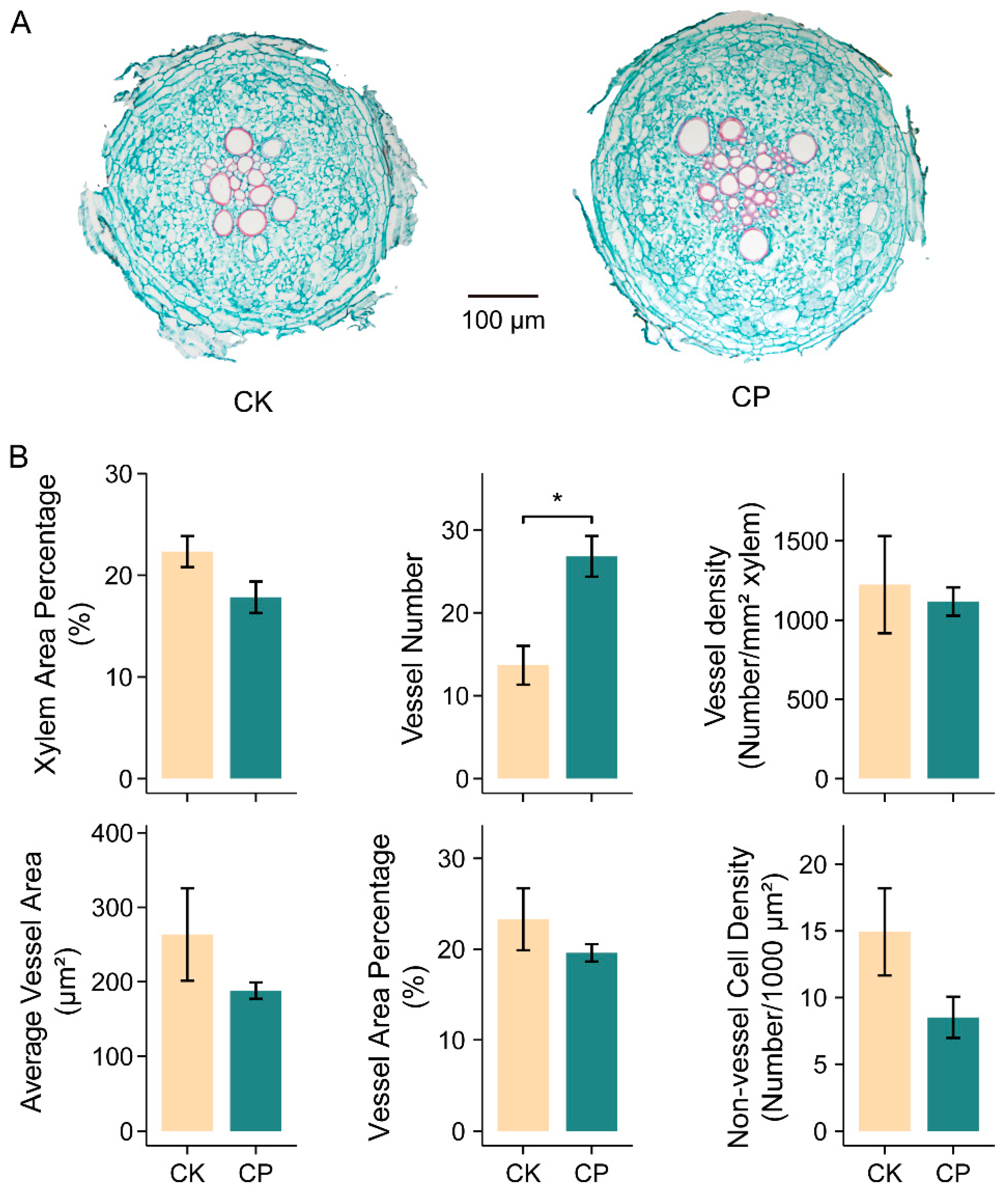
3.3. Adapted Phenylpropanoid-Derived Pathways and Root Anatomy under Soil Compaction
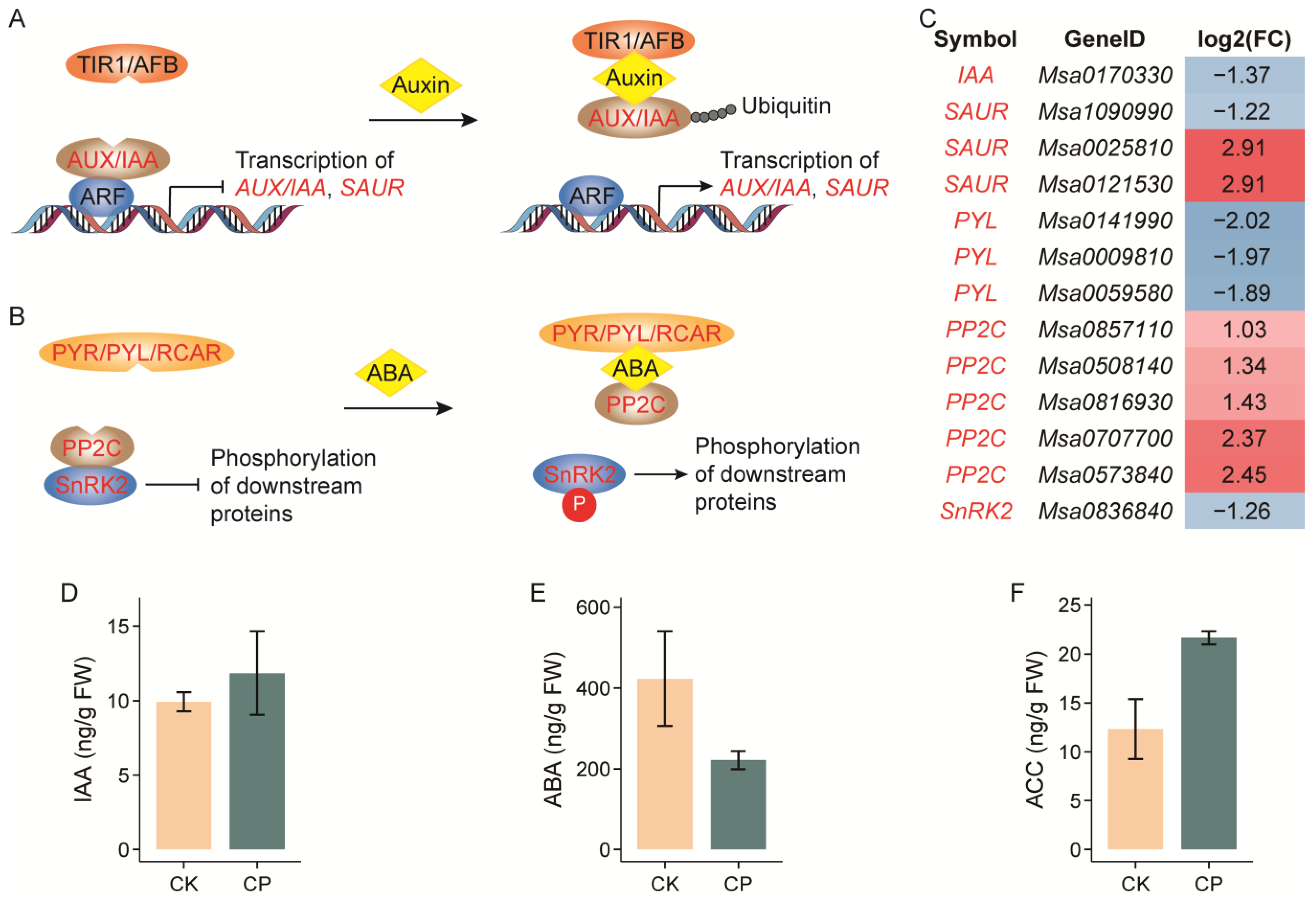
3.4. Alterations in Phytohormone Biosynthesis
4. Discussion
4.1. Effects of Soil Compaction on Alfalfa Growth Depend on Stress Severity and Plant Germplasm
4.2. Phenylpropanoid, Lignin, Flavonoid and Cell Wall Modification in Alfalfa Root Responses to Soil Compaction
4.3. Plant Hormone Networks Involved in Alfalfa Root Responses to Soil Compaction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gao, A.-M.; Wu, J.-F.; Dai, F.; Zhang, F.-W.; Han, Z.-S. Correlation study of soil compaction by mowing machine and alfalfa yield. Acta Prataculturae Sin. 2014, 23, 59. [Google Scholar]
- Shaheb, M.R.; Venkatesh, R.; Shearer, S.A. A review on the effect of soil compaction and its management for sustainable crop production. J. Biosyst. Eng. 2021, 46, 417–439. [Google Scholar] [CrossRef]
- Correa, J.; Postma, J.A.; Watt, M.; Wojciechowski, T. Soil compaction and the architectural plasticity of root systems. J. Exp. Bot. 2019, 70, 6019–6034. [Google Scholar] [CrossRef] [PubMed]
- Pandey, B.K.; Bennett, M.J. Uncovering root compaction response mechanisms: New insights and opportunities. J. Exp. Bot. 2024, 75, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Longepierre, M.; Feola Conz, R.; Barthel, M.; Bru, D.; Philippot, L.; Six, J.; Hartmann, M. Mixed effects of soil compaction on the nitrogen cycle under pea and wheat. Front. Microbiol. 2022, 12, 822487. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Colombi, T.; Jäck, O.; Keller, T.; Weih, M. Effects of soil compaction on grain yield of wheat depend on weather conditions. Sci. Total Environ. 2022, 807, 150763. [Google Scholar] [CrossRef]
- Trükmann, K.; Reintam, E.; Kuht, J.; Nugis, E.; Edesi, L. Effect of soil compaction on growth of narrow–leafed lupine, oilseed rape and spring barley on sandy loam soil. Agron. Res. 2008, 6, 101–108. [Google Scholar]
- Bello-Bello, E.; López-Arredondo, D.; Rico-Chambrón, T.Y.; Herrera-Estrella, L. Conquering compacted soils: Uncovering the molecular components of root soil penetration. Trends Plant Sci. 2022, 27, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, A.G.R.; Jervis, G.; Xu, J.; Topping, J.F.; Lindsey, K. Root growth responses to mechanical impedance are regulated by a network of ROS, ethylene and auxin signalling in Arabidopsis. New Phytol. 2021, 231, 225–242. [Google Scholar] [CrossRef]
- Lee, H.-J.; Kim, H.-S.; Park, J.M.; Cho, H.S.; Jeon, J.H. PIN-mediated polar auxin transport facilitates root—Obstacle avoidance. New Phytol. 2020, 225, 1285–1296. [Google Scholar] [CrossRef]
- Zhang, Y.; Friml, J. Auxin guides roots to avoid obstacles during gravitropic growth. New Phytol. 2020, 225, 1049–1052. [Google Scholar] [CrossRef]
- Shih, H.-W.; Miller, N.D.; Dai, C.; Spalding, E.P.; Monshausen, G.B. The receptor-like kinase FERONIA is required for mechanical signal transduction in Arabidopsis seedlings. Curr. Biol. 2014, 24, 1887–1892. [Google Scholar] [CrossRef]
- Huang, G.; Kilic, A.; Karady, M.; Zhang, J.; Mehra, P.; Song, X.; Sturrock, C.J.; Zhu, W.; Qin, H.; Hartman, S.; et al. Ethylene inhibits rice root elongation in compacted soil via ABA- and auxin-mediated mechanisms. Proc. Natl. Acad. Sci. USA 2022, 119, e2201072119. [Google Scholar] [CrossRef]
- Pandey, B.K.; Huang, G.; Bhosale, R.; Hartman, S.; Sturrock, C.J.; Jose, L.; Martin, O.C.; Karady, M.; Voesenek, L.A.C.J.; Ljung, K.; et al. Plant roots sense soil compaction through restricted ethylene diffusion. Science 2021, 371, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Wang, J.; Zhou, J.; Qiao, J.; Li, Y.; Quan, R.; Huang, R. Abscisic acid promotes auxin biosynthesis to inhibit primary root elongation in rice. Plant Physiol. 2023, 191, 1953–1967. [Google Scholar] [CrossRef]
- Xu, F.; Chen, J.; Li, Y.; Ouyang, S.; Yu, M.; Wang, Y.; Fang, X.; He, K.; Yu, F. The soil emergence-related transcription factor PIF3 controls root penetration by interacting with the receptor kinase FER. Dev. Cell 2024, 59, 434–447.e438. [Google Scholar] [CrossRef] [PubMed]
- Bello-Bello, E.; Herrera-Estrella, L. Breaking new ground: Decoding the root’s molecular circuits to penetrate compacted soil. Dev. Cell 2024, 59, 431–433. [Google Scholar] [CrossRef]
- Yang, L.; Wu, Q.; Liang, H.; Yin, L.; Shen, P. Integrated analyses of transcriptome and metabolome provides new insights into the primary and secondary metabolism in response to nitrogen deficiency and soil compaction stress in peanut roots. Front. Plant Sci. 2022, 13, 948742. [Google Scholar] [CrossRef] [PubMed]
- Schneider, H.M.; Strock, C.F.; Hanlon, M.T.; Vanhees, D.J.; Perkins, A.C.; Ajmera, I.B.; Sidhu, J.S.; Mooney, S.J.; Brown, K.M.; Lynch, J.P. Multiseriate cortical sclerenchyma enhance root penetration in compacted soils. Proc. Natl. Acad. Sci. USA 2021, 118, e2012087118. [Google Scholar] [CrossRef] [PubMed]
- Głąb, T. Effects of tractor wheeling on root morphology and yield of lucerne (Medicago sativa L.). Grass Forage Sci. 2008, 63, 398–406. [Google Scholar] [CrossRef]
- Gląb, T. Effect of soil compaction on root system morphology and productivity of Alfalfa (Medicago sativa L.). Pol. J. Environ. Stud. 2011, 20, 1473–1480. [Google Scholar]
- Głąb, T.; Gondek, K. The influence of soil compaction on chemical properties of mollic fluvisol soil under lucerne (Medicago sativa L.). Pol. J. Environ. Stud. 2013, 22, 107–113. [Google Scholar]
- Zhen, L.; Zhang, Y.; Bai, C.; Yang, J. Comparative studies on soil water content and soil bulk density of lucerne with different cultivating ages. Mod. J. Anim. Husb. Vet. Med. 2011, 7, 58–59. [Google Scholar]
- Gao, A.; Zhang, X.; Wang, Y.; Huang, X.; Han, Z. Experimental study on effect of mechanical compaction on the soil structure in different types of alfalfa fields. For. Mach. Woodwork. Equip. 2019, 47, 53–58. [Google Scholar]
- Seethepalli, A.; Dhakal, K.; Griffiths, M.; Guo, H.; Freschet, G.T.; York, L.M. RhizoVision Explorer: Open-source software for root image analysis and measurement standardization. AoB Plants 2021, 13, plab056. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.-G.; Ye, J.; Yang, S.-N.; Fu, J.-R.; Ma, J.-W.; Sun, W.-C.; Jiang, L.-N.; Wang, Q. Effects of nitrogen application level on rice nutrient uptake and ammonia volatilization. Rice Sci. 2013, 20, 139–147. [Google Scholar] [CrossRef]
- Li, Z.-Y.; Long, R.-C.; Zhang, T.-J.; Yang, Q.-C.; Kang, J.-M. Molecular cloning and characterization of the MsHSP17.7 gene from Medicago sativa L. Mol. Biol. Rep. 2016, 43, 815–826. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Long, R.; Zhang, F.; Zhang, Z.; Li, M.; Chen, L.; Wang, X.; Liu, W.; Zhang, T.; Yu, L.-X.; He, F.; et al. Genome assembly of alfalfa cultivar Zhongmu-4 and identification of SNPs associated with agronomic traits. Genom. Proteom. Bioinform. 2022, 20, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Oh, D.-H.; Dassanayake, M. GOMCL: A toolkit to cluster, evaluate, and extract non-redundant associations of Gene Ontology-based functions. BMC Bioinform. 2020, 21, 139. [Google Scholar] [CrossRef]
- Arvidsson, J.; Håkansson, I. Response of different crops to soil compaction—Short-term effects in Swedish field experiments. Soil Tillage Res. 2014, 138, 56–63. [Google Scholar] [CrossRef]
- Whalley, W.R.; Dumitru, E.; Dexter, A. Biological effects of soil compaction. Soil Tillage Res. 1995, 35, 53–68. [Google Scholar] [CrossRef]
- Mósena, M.; Dillenburg, L.R. Early growth of Brazilian pine (Araucaria angustifolia [Bertol.] Kuntze) in response to soil compaction and drought. Plant Soil 2004, 258, 293–306. [Google Scholar] [CrossRef]
- Bejarano, M.D.; Villar, R.; Murillo, A.M.; Quero, J.L. Effects of soil compaction and light on growth of Quercus pyrenaica Willd. (Fagaceae) seedlings. Soil Tillage Res. 2010, 110, 108–114. [Google Scholar] [CrossRef]
- Alameda, D.; Villar, R. Linking root traits to plant physiology and growth in Fraxinus angustifolia Vahl. seedlings under soil compaction conditions. Environ. Exp. Bot. 2012, 79, 49–57. [Google Scholar] [CrossRef]
- Zhao, Q. Lignification: Flexibility, biosynthesis and regulation. Trends Plant Sci. 2016, 21, 713–721. [Google Scholar] [CrossRef]
- Cesarino, I. Structural features and regulation of lignin deposited upon biotic and abiotic stresses. Curr. Opin. Biotechnol. 2019, 56, 209–214. [Google Scholar] [CrossRef]
- Muro-Villanueva, F.; Mao, X.; Chapple, C. Linking phenylpropanoid metabolism, lignin deposition, and plant growth inhibition. Curr. Opin. Biotechnol. 2019, 56, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.S.S.; Chen, F.; Shadle, G.; Jackson, L.; Aljoe, H.; Dixon, R.A. Targeted down-regulation of cytochrome P450 enzymes for forage quality improvement in alfalfa (Medicago sativa L.). Proc. Natl. Acad. Sci. USA 2005, 102, 16573–16578. [Google Scholar] [CrossRef]
- Gallego-Giraldo, L.; Jikumaru, Y.; Kamiya, Y.; Tang, Y.; Dixon, R.A. Selective lignin downregulation leads to constitutive defense response expression in alfalfa (Medicago sativa L.). New Phytol. 2011, 190, 627–639. [Google Scholar] [CrossRef] [PubMed]
- Hacke, U.G.; Spicer, R.; Schreiber, S.G.; Plavcová, L. An ecophysiological and developmental perspective on variation in vessel diameter. Plant Cell Environ. 2017, 40, 831–845. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, Y.; Wang, X.; Han, X.; An, Y.; Lin, S.; Shen, C.; Wen, J.; Liu, C.; Yin, W.; et al. Root-specific NF-Y family transcription factor, PdNF-YB21, positively regulates root growth and drought resistance by abscisic acid-mediated indoylacetic acid transport in Populus. New Phytol. 2020, 227, 407–426. [Google Scholar] [CrossRef] [PubMed]
- Hummel, I.; Vile, D.; Violle, C.; Devaux, J.; Ricci, B.; Blanchard, A.; Garnier, É.; Roumet, C. Relating root structure and anatomy to whole-plant functioning in 14 herbaceous Mediterranean species. New Phytol. 2007, 173, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Lv, S.; Zhao, L.; Gao, T.; Yu, C.; Hu, J.; Ma, F. Advances in the study of the function and mechanism of the action of flavonoids in plants under environmental stresses. Planta 2023, 257, 108. [Google Scholar] [CrossRef]
- Shah, A.; Smith, D.L. Flavonoids in agriculture: Chemistry and roles in biotic and abiotic stress responses, and microbial associations. Agronomy 2020, 10, 1209. [Google Scholar] [CrossRef]
- Dakora, F.D.; Joseph, C.M.; Phillips, D.A. Alfalfa (Medicago sativa L.) root exudates contain isoflavonoids in the presence of rhizobium meliloti. Plant Physiol. 1993, 101, 819–824. [Google Scholar] [CrossRef]
- Bosse, M.A.; Silva, M.B.d.; Oliveira, N.G.R.M.d.; Araujo, M.A.d.; Rodrigues, C.; Azevedo, J.P.d.; Reis, A.R.d. Physiological impact of flavonoids on nodulation and ureide metabolism in legume plants. Plant Physiol. Biochem. 2021, 166, 512–521. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, J.; Li, H.; Chiang, V.L.; Fu, Y. Cooperative regulation of flavonoid and lignin biosynthesis in plants. Crit. Rev. Plant Sci. 2021, 40, 109–126. [Google Scholar] [CrossRef]
- Gallego-Giraldo, L.; Bhattarai, K.; Pislariu, C.I.; Nakashima, J.; Jikumaru, Y.; Kamiya, Y.; Udvardi, M.K.; Monteros, M.J.; Dixon, R.A. Lignin modification leads to increased nodule numbers in alfalfa. Plant Physiol. 2014, 164, 1139–1150. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, F.; Friml, J.; Ding, Z. Auxin signaling: Research advances over the past 30 years. J. Integr. Plant Biol. 2022, 64, 371–392. [Google Scholar] [CrossRef] [PubMed]
- Spartz, A.K.; Lee, S.H.; Wenger, J.P.; Gonzalez, N.; Itoh, H.; Inzé, D.; Peer, W.A.; Murphy, A.S.; Overvoorde, P.J.; Gray, W.M. The SAUR19 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. Plant J. 2012, 70, 978–990. [Google Scholar] [CrossRef] [PubMed]
- Chae, K.; Isaacs, C.G.; Reeves, P.H.; Maloney, G.S.; Muday, G.K.; Nagpal, P.; Reed, J.W. Arabidopsis SMALL AUXIN UP RNA63 promotes hypocotyl and stamen filament elongation. Plant J. 2012, 71, 684–697. [Google Scholar] [CrossRef]
- Qiu, T.; Qi, M.; Ding, X.; Zheng, Y.; Zhou, T.; Chen, Y.; Han, N.; Zhu, M.; Bian, H.; Wang, J. The SAUR41 subfamily of SMALL AUXIN UP RNA genes is abscisic acid inducible to modulate cell expansion and salt tolerance in Arabidopsis thaliana seedlings. Ann. Bot. 2019, 125, 805–819. [Google Scholar] [CrossRef]
- Fidler, J.; Graska, J.; Gietler, M.; Nykiel, M.; Prabucka, B.; Rybarczyk-Płońska, A.; Muszyńska, E.; Morkunas, I.; Labudda, M. PYR/PYL/RCAR receptors play a vital role in the abscisic-acid-dependent responses of plants to external or internal stimuli. Cells 2022, 11, 1352. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.-F.F.; et al. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, M.; Yang, D.; He, Y.; Ma, Y.; Zhang, X.; Wang, Q.; Gao, J. Alfalfa Responses to Intensive Soil Compaction: Effects on Plant and Root Growth, Phytohormones and Internal Gene Expression. Plants 2024, 13, 953. https://doi.org/10.3390/plants13070953
Yan M, Yang D, He Y, Ma Y, Zhang X, Wang Q, Gao J. Alfalfa Responses to Intensive Soil Compaction: Effects on Plant and Root Growth, Phytohormones and Internal Gene Expression. Plants. 2024; 13(7):953. https://doi.org/10.3390/plants13070953
Chicago/Turabian StyleYan, Mingke, Dongming Yang, Yijun He, Yonglong Ma, Xin Zhang, Quanzhen Wang, and Jinghui Gao. 2024. "Alfalfa Responses to Intensive Soil Compaction: Effects on Plant and Root Growth, Phytohormones and Internal Gene Expression" Plants 13, no. 7: 953. https://doi.org/10.3390/plants13070953
APA StyleYan, M., Yang, D., He, Y., Ma, Y., Zhang, X., Wang, Q., & Gao, J. (2024). Alfalfa Responses to Intensive Soil Compaction: Effects on Plant and Root Growth, Phytohormones and Internal Gene Expression. Plants, 13(7), 953. https://doi.org/10.3390/plants13070953