Abstract
Improving fruit size or weight, firmness, and shelf life is a major target for horticultural crop breeding. It is associated with the depolymerization and rearrangement of cell components, including pectin, hemicellulose, cellulose, and other structural (glyco)proteins. Expansins are structural proteins to loosen plant cell wall polysaccharides in a pH-dependent manner and play pivotal roles in the process of fruit development, ripening, and softening. Rubus chingii Hu, a unique Chinese red raspberry, is a prestigious pharmaceutical and nutraceutical dual-function food with great economic value. Thirty-three RchEXPs were predicted by genome-wide identification in this study, containing twenty-seven α-expansins (EXPAs), three β-expansins (EXPBs), one expansin-like A (EXPLA), and two expansin-like B (EXPLBs). Subsequently, molecular characteristics, gene structure and motif compositions, phylogenetic relationships, chromosomal location, collinearity, and regulatory elements were further profiled. Furthermore, transcriptome sequencing (RNA-seq) and real-time quantitative PCR assays of fruits from different developmental stages and lineages showed that the group of RchEXPA5, RchEXPA7, and RchEXPA15 were synergistically involved in fruit expanding and ripening, while another group of RchEXPA6 and RchEXPA26 might be essential for fruit ripening and softening. They were regulated by both abscisic acid and ethylene and were collinear with phylogenetic relationships in the same group. Our new findings laid the molecular foundation for improving the fruit texture and shelf life of R. chingii medicinal and edible fruit.
1. Introduction
Rubus chingii Hu is an economically important Eastern Chinese raspberry with a prestigious pharmaceutical and nutraceutical dual function [1,2]. The unripe fruit harvested at the stages of big green (BG) or green-to-yellow (GY) has been used in traditional Chinese medicine for tonifying kidneys and eyesight from the ancient Qin and Han Dynasties [1,3]. It has been widely used as a key ingredient in some Chinese Proprietary Medicines, such as Wuzi Yanzong Pill, Guilu Bushen Pill, Shenbao Mixture, and others, or soaked in white wine for medicinal liquor in folk [1]. More than 235 chemical constituents, including terpenoids, flavonoids, phenolics, polysaccharides, and others, have been identified and isolated in recent years [4,5,6]. The pharmacological studies indicated the therapeutical effects of anti-inflammation, antioxidantion, antidiabetic, anti-thrombosis, anti-tumor, neuroprotective, and liver protection of R. chingii fruit and leaves [1,4,7,8,9,10,11]. Simultaneously, the ripe fruit of R. chingii is popular for pleasant fresh fruit, like raspberries and blackberries, and has rich nutrients, including amino acids, phenolic acids, vitamins C and PP, ellagitannins, aromatics, and minerals, such as magnesium, zinc, and potassium, and it is beneficial for human health [2,12,13]. In the past, the utilization of R. chingii mainly depended on wild resources. During the last decade, spurred by increasing consumption, the R. chingii industry flourished in East and South China. The planting area was about 1000 ha in China in 2017 and reached more than 17,000 ha in 2019 with the production of nearly 70 thousand tons of fresh immature and ripe fruits [2]. Nevertheless, the fresh red berry of R. chingii is very perishable with a short shelf life, although it is regarded as non-climacteric like other berries [14]. Furthermore, to meet the demands of higher yield, greater quality, and excellent health benefits, thousands of germplasms from wild habitats were domesticated and crossed to breed superior cultivars [15]. For economic yield, fruit size is one of the major characteristics of breeding. However, there is little knowledge on R. chingii fruit expansion and ripening in the literature to date. Therefore, unveiling the mechanism and vital genes for fruit enlargement and softening of the unique plant, R. chingii, can provide useful information for fruit development, ripening, softening, and molecular breeding. In the genomic age, genomic sequencing results have greatly facilitated the illumination of key genes for traits. The first genome of Rubus was reported in 2016 [16]. R. chingii genomic sequencing was also completed in 2021 [6]. Based on this, several vital genes, such as flavonol synthesis (FLS), dihydroflavonol 4-reductase (DFR), and MYB308, have been investigated by integrated multiple-omics techniques [17,18]. In this study, genome-wide identification and RNA-seq of key genes for fruit ripening and softening were performed.
Fruit enlarging, ripening, and softening processes are closely associated with plant cell wall loosening, degradation, extension, and reconstruction. These changes in the cell wall are attributed to the depolymerization, and hydrolyzation of pectin, hemicellulose, cellulose, and microfibrils, which are induced by a complex set of enzymes, such as pectate lyase (PL or Pel), pectin methyl esterase (PME), polygalacturonase (PG), cellulase (Cel), xyloglucan endotransglucosylase/hydrolase (XTH), and others [19,20]. In addition, non-enzymatic cell wall proteins like expansins (EXPs) are also essentially involved in cell wall loosening and rearrangements in an acid-dependent manner [21]. EXPs are a group of structural proteins. A typical expansin is composed of 250–275 amino acids (aa) and has two functional domains and a signal peptide (SP): (1) the N-terminal 20–30 aa is for an SP to possibly use transmembrane transport for secretion; (2) the N-terminal 120–135 aa contributes to the six-stranded double-psi beta-barrel (DPBB_1) domain (PF03330) with glycoside hydrolase (GH45)-like catalytic activity; and (3) C-terminal 90–120 aa residues contribute to the Pollen_allerg_1 domain (PF01357), named Expansin_C [22,23]. According to phylogenetic analysis, the EXP family members can be divided into four subfamilies: α-expansin (EXPA), β-expansin (EXPB), expansin-like A (EXLA or EXPLA), and expansin-like B (EXLB or EXPLB) [23]. They work in divergent ways by a process of molecular “creep” to loosen the cell wall. EXPA members may promote cellulose microfibrils separation and entrail movement by local dissociation and the slippage of xyloglucans on the cellulose surface, whereas EXPB members may work on other glycans, like xylan, or dissolve the middle lamella of stigma to facilitate pollen tube invasion [21,23]. However, little information is available regarding the action mechanism of EXLA and EXLB.
EXPs exert cell wall loosening activity during seed germination, root growth and development, stem elongation, leaf initiation and growth, stomatal movement, fruit ripening and softening, and abscission, as well as in responses to biotic and abiotic stresses [23,24]. There are thirty-six members in Arabidopsis thaliana, including twenty-six EXPA, six EXPB, three EXLA, and one EXLB [22]. AtEXPA1-promoted stomatal opening, AtEXPA2 accelerated germination, AtEXPA3 enhanced growth and enlarged leaves, and AtEXPA10 increased the sizes of plant cells, leaves, and stems. The overexpression of AtEXPB1 and AtEXPB5 plants have longer petioles and soft stigma, respectively [24]. There are 35 and 114 candidate genes in Fragaria vesca and F. ananassa, respectively. Among these, FvEXPA9 (FveEXP11), FvEXPA12 (FveEXP14), and FvEXPA27 (FveEXP33) were highly expressed in fruit ripening stages [21,25]; FaEXPA2, FaEXPA5, FaEXP88, and FaEXP114 were closely related with fruit softening [21]. In addition, FaEXP7 was regarded as a potential softening activator [20]. However, there is no report about expansins in Rubus, which greatly limits the comprehensive understanding of fruit development and softness for these attractive berries. In this study, we identified 33 expansins encoding genes from R. chingii and systematically analyzed the bioinformation of RchEXPs and their roles in fruit ripening and softening. The results provided valuable insights into the characterization and function of the expansin genes family in R. chingii, an important Chinese herb and red raspberry.
2. Results
2.1. Identification of Expansin Family Members in Rubus chingii
A total of 39 genes and their proteins were initially obtained by the HMM search. Among these, thirty proteins consisting of two conserved domains (DPBB_1 and Expansin_C domains) were selected as expansins candidates in R. chingii. In addition, proteins with one complete DPBB_1 domain, such as LG04.3299, LG04.3300, and LG04.3301, were also retained according to strawberry expansins [21,25]. Therefore, 33 RchEXPs were identified in this study (Table 1 and Table S1). Furthermore, expansin genes in its sister plants, red raspberry (Rubus idaeus) and black raspberry (Rubus occidentalis), were also extracted from the genomic data, and a similar number of members were obtained (Tables S1 and S2), indicating the similarity of the same Rubus. The amino acids of RchEXPs ranged from 125 aa (RchEXPA8, molecular weight MW = 12,970.17) to 482 aa (RchEXPA4, MW = 53,196.43) and the isoelectric point (pI) of most members (26/33) were greater than 7, indicating these members might be rich in alkaline amino acids. The prediction of the subcellular location showed that RchEXPs were all located on the cell wall.

Table 1.
Characteristics of RchEXP family members in Rubus chingii Hu.
Phylogenetic analyses showed RchEXPs had relatively distant relatives with AtEXPs (Figure S1a), while they were more closely related to FvEXPs, RiEXPs, and RoEXPs, which belonged to the same Rosaceae (Figure 1). Based on the phylogenetic relationships with FvEXPs, all RchEXPs were divided into four subfamilies, including twenty-seven (81.8%) members of EXPA, three members of EXPB, one member of EXPLA, and two members of EXPLB (Table 1 and Table S1). Twenty-three orthologous pairs in R. chingii and F. vesca, and five paralogous pairs in R. chingii, were identified based on the high bootstrap values (>90%) (Figure 2 and Figure S1b).
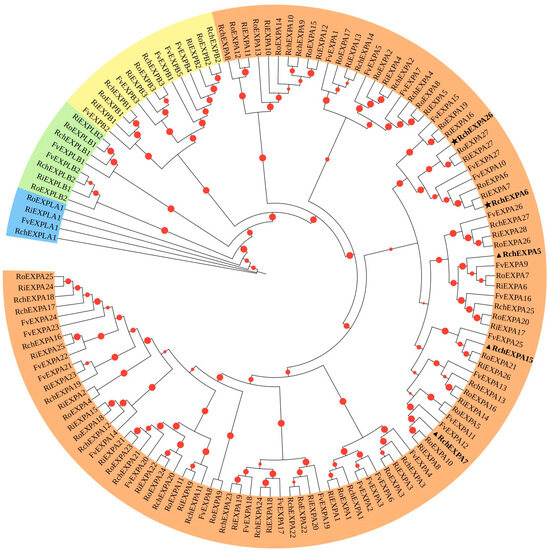
Figure 1.
Phylogenetic tree of expansins from Rubus chingii, Fragaria vesca, Rubus idaeus, and R. occidentalis. EXPA, EXPB, EXPLA, and EXPLB subfamilies were presented in light orange, yellow, blue, and green. “★”indicates the same expression tendency during fruit development stages which was highest at the red stage, while “▲” indicates that these gene expressions were initiated in earlier stages of fruit.
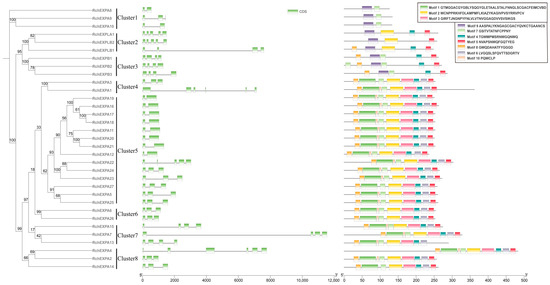
Figure 2.
Phylogenetic relationships, gene structures of exon/intron, and motif compositions of expansins in Rubus chingii.
2.2. Gene Structures and Protein Motifs of RchEXPs
The analyses of gene structures of RchEXPs exhibited the members of exons and varied from two to six, and introns varied from one to five (Figure 2). All RchEXPBs and RchEXPLB1 had four exons, whereas RchEXPLA and RchEXPLB2 contained five exons. Among the EXPA subfamily, 37% of the members contained two exons and 40.7% of the members contained three exons, whereas the others had four, five, or six exons. They could be divided into six clusters, and most members in the same cluster showed consistent exon numbers. All these results indicated the similarities of the same subfamily in structure.
Meanwhile, we provided the organization of the conserved motifs in Figure 2. Most members (77.8%) of the EXPA subfamily in R. chingii were composed of nine motifs, including Motifs 8, 1, 10, 7, 2, 3, 4, 6, and 5, whereas RchEXPA3, RchEXPA14, and RchEXPA27 contained the above eight motifs, except for Motif 10. Unexpectedly, RchEXPA8–10 contained only Motifs 9 and 7. RchEXPBs consisted of Motifs 8, 9, 7, 4, and 5. Motif 7 was found in all EXPAs and EXPBs (90.9% of the total) in R. chingii, except for EXPLA and EXPLBs. Oppositely, Motif 9 was not founded in most RchEXPAs, except for RchEXPA8–10. In addition, Motif 2 was not founded in RchEXPBs. All these results suggested the conserved structural evolution and consistency of the same subfamily.
2.3. Chromosomal Location and Synteny Analyses of RchEXPs
RchEXPs were unevenly located on the seven chromosomes (Figure 3). The distribution pattern was similar to F. vesca [25]. Chromosome 6 (LG06) possessed the largest number (12/33) of RchEXPs encoding genes, including eleven members of the EXPA subfamily and one EXPB. Among these, three tandem duplication events occurred, including RchEXPA16–19, RchEXPA20–21, and RchEXPA22–24. Another tandem duplication was observed among RchEXPA8, A9, and A10 in chromosome 4 (LG04). Chromosome 1 (LG01) had only one expansin gene (RchEXPA1). Chromosomes 2 (LG02) and 3 (LG03) both contained three RchEXP genes, whereas Chromosomes 4 and 5 (LG05) contained five RchEXP genes. RchEXPBs were distributed on chromosomes 5 and 7 (LG07). RchEXPLA was located on chromosome 2, whereas RchEXPLBs were located on chromosome 5, and both were in line with F. vesca. All these results supported the conserved evolution of expansins in berries, like strawberries and Rubus. Furthermore, synteny analysis indicated that only five collinear EXP gene pairs were found in R. chingii, including RchEXPA6 and RchEXPA26, RchEXPA5 and RchEXPA27, RchEXPA5 and RchEXPA25, RchEXPA7 and RchEXPA13, and RchEXPA7 and RchEXPA15 (Figure 4a). In addition, 33 collinear EXP gene pairs were identified between R. chingii and F. vesca (Figure 4b). Interestingly, the collinear events mainly occurred in RchEXPA5, RchEXPA6, RchEXPA7, RchEXPA13, RchEXPA15, and RchEXPA26. They all had three syntenic relationships with corresponding expansin genes in wild strawberry F. vesca (Table S3). For example, both RchEXPA6 and RchEXPA26 had collinear relationships with FvEXPA10, FvEXPA15, and FvEXPA27; both RchEXPA7 and RchEXPA25 had collinear relationships with FvEXPA12, FvEXPA13, and FvEXPA25; while RchEXPA5 had collinear relationships with FvEXPA9, FvEXPA16, and FvEXPA26 (Figure 4b and Table S3). All these results suggested that the orthologous and paralog gene pairs and roles of EXPs are found in berry fruit.
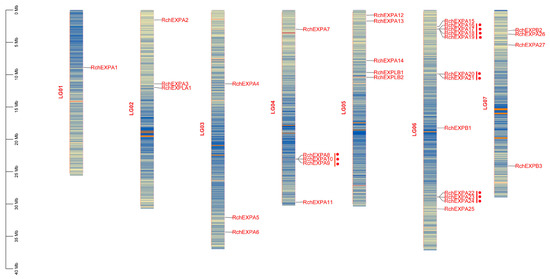
Figure 3.
Chromosomal location of expansin encoding genes in R. chingii. The red dot means the tandem duplication events, including RchEXPA8–10, RchEXPA16–19, RchEXPA20–21, and RchEXPA22–24.
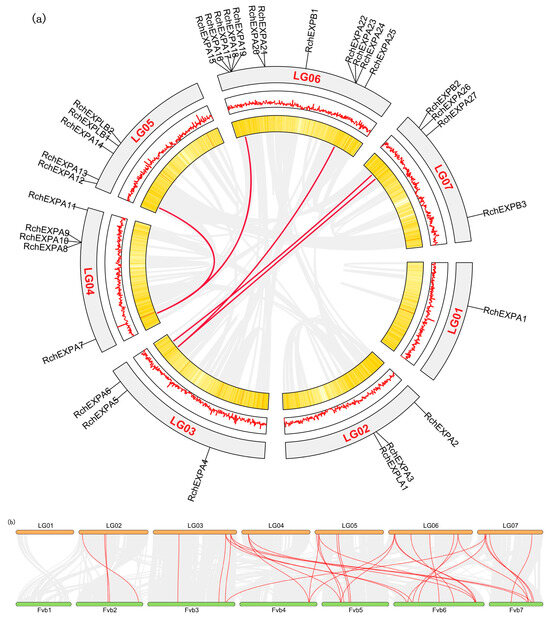
Figure 4.
Synteny analyses of R. chingii and F. vesa genomes. (a) Synteny analysis of R. chingii genome. The gene density was displayed in the form of heat maps and lines. Syntenic blocks were linked by gray lines, and syntenic relationships of EXP members were highlighted by red color. (b) Syntenic relationships of R. chingii and F. vesa genomes. Syntenic EXP gene pairs between R. chingii and F. vesa were highlighted by red color.
2.4. Cis-Regulatory Elements and Interaction Analyses of RchEXPs
Analysis of cis-acting elements on the promoter sequences (2 kb upstream of ATG) could predict the regulatory factors and temporal–spatial expression patterns of RchEXPs (Figure 5a). First, cis-regulatory elements of a phytohormone response, such as abscisic acid-responsive elements (ABREs and AAGAA-motif), ethylene-responsive elements (EREs), auxin-responsive elements (AuxRR-core and TGA-element), gibberellin (GA)-responsive elements (GARE-motif, P-box and CARE), salicylic acid (SA)-responsive element (TCA-element) SA, auxin-response element (as-1), and jasmonic acid methyl ester (MeJA) responsive elements (TGACG-motif and CGTCA-motif) were detected in this study. The results showed promoters of most RchEXPs possessed ABRE sites, except for EXPA1, A8, A22, and A24. Moreover, the majority of them contained multiple ABRE sites, such as nine sites of the RchEXPA13 promoter, six sites of the RchEXPA4 promoter, and four sites of the RchEXPA5, RchEXPA6, RchEXPA26, RchEXPA3, RchEXPA9, RchEXPA14, RchEXPA20, RchEXPB1, and RchEXPLB1 promoters. In addition, promoters of major RchEXPs had 1–4 AAGAA-motif sites. Both ABRE and AAGAA-motif sites indicated the possible ABA regulation of RchEXPs. Promoters of some RchEXPs had ERE sites, such as RchEXPA26, RchEXPA1–2, RchEXPA7–9, RchEXPA11–12, and others. There was an auxin-responsive element (TGA-element) in the promoter of RchEXPA5, RchEXPA6, and RchEXPA7. Most RchEXPs were associated with SA and MeJA, but may not respond to GA. Taken together, the five genes that were highly expressed in fruit (Figure 6), including RchEXPA5, RchEXPA6, RchEXPA7, RchEXPA15, and RchEXPA26, all could respond to the ABA signal, while only RchEXPA7 and RchEXPA26 were putatively regulated by Eth (Figure 5a). These results predicted the main regulation of ABA in non-climacteric fruits [14].
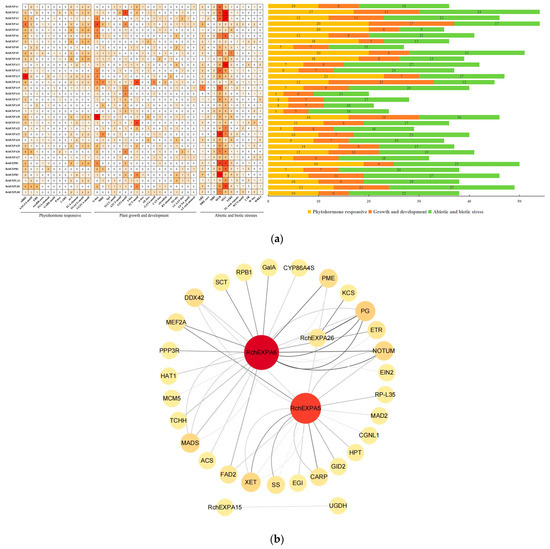
Figure 5.
Predicted cis-acting elements and protein–protein interaction networks of expansins in R. chingii. (a) Predicted cis-acting elements and their numbers in the promoters of expansin genes in R. chingii. (b) Construction of protein–protein interaction networks of expansins and other proteins in R. chingii.
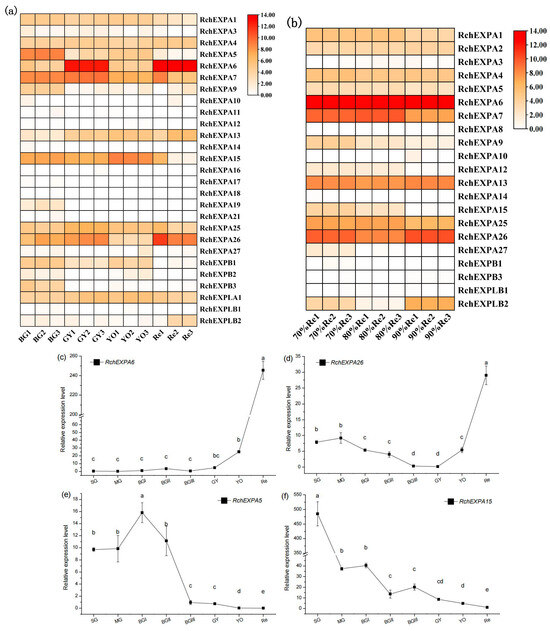
Figure 6.
Expression profile of RchEXPs during fruit ripening and softening. (a) RchEXPs expression levels at the four representative stages, BGI, GY, YO, and Re, by RNA-seq. (b) RchEXPs expression levels at the red stages of the different degrees of ripeness by RNA-seq. (c–f) qPCR of RchEXPA6, RchEXPA26, RchEXPA5, and RchEXPA15. Different lower cases mean the significant difference at p < 0.05 by a one-way analysis of variance with the LSD method.
Secondly, cis-regulatory elements of plant growth and development were analyzed, including elements involved in light responsiveness (MRE, Sp1, TCCC-motif, ATCT-motif, GT1-motif, I-box, TCT-motif, A-box, AE-box and GATA-motif), meristem expression (CAT-box), circadian control (circadian), seed-specific regulation (RY-element), zein metabolism regulation (O2-site), endosperm expression (GCN4_motif), and MYBHv1 and AT-rich DNA binding protein (ATBP-1) binding sites (CCAAT-box and AT-rich element). The results indicated the action of RchEXPs was mainly regulated by light in divergent ways (Figure 5a). Finally, RchEXPs might actively participate in stress responses because of the most abundant sites in their promoters, including cis-acting elements of STRE (wound-responsive element), ARE (for anaerobic induction), LTR (involved in low-temperature responsiveness), MBS (involved in drought inducibility), TC-rich repeats (involved in defense and stress responsiveness), WUN-motif (wound-responsive element), DRE core, W box, and WRE3, especially MYB and MYC. Five genes, RchEXPA5, RchEXPA6, RchEXPA7, RchEXPA15, and RchEXPA26, all could respond to anaerobic and wound stress and could be regulated by MYBs and MYCs, whereas RchEXPA6, RchEXPA15, and RchEXPA26 could respond to drought (Figure 5a).
Protein–protein interaction networks (PPIs) were constructed by a cytoscape tool based on transcriptome data (Figure 5b). RchEXPA6 was predicted to strongly interact with cell wall hydrolases, including PME and PG. It also could interact with an ethylene receptor (ETR) and ethylene-insensitive protein 2 (EIN2) and then result in ethylene-induced responses. Meanwhile, RchEXPA26 also could tightly interact with PG. These results suggest that RchEXPA6 and RchEXPA26 were apt in their involvement in fruit softening. RchEXPA5 was predicted to interact with PME, xyloglucan/xyloglucosyl transferase (XET), sucrose synthase (SS), and others. Hence, it might work earlier in fruit development. No PPI was found between RchEXPA7 and other proteins. Only UDP glucose 6-dehydrogenase (UGDH) was found to interact with RchEXPA15.
2.5. Expression Profiles of RchEXPs in Different Fruit Ripening and Softening Stages of R. chingii
The fresh weight of the eight stages (SG—small green, MG—middle green, BGI—big green I, BGII—big green II, BGIII—big green III, GY—green to yellow, YO—yellow to orange, Re—red) during R. chingii fruit development were 0.25, 0.48, 0.97, 1.02, 1.02, 1.43, 1.80, and 4.54 g, respectively [5]. Fruit firmness before the 60% red stages was all >15 kg/cm2, whereas fruit firmness of the 70% red, 80% red, and 90% red stages were 10.81, 8.47, 6.48 kg/cm2, respectively, indicating that the firmness was gradually reduced with fruit ripening. The data of transcriptomic sequencing showed the divergent expression pattern of all the expansin genes in R. chingii. Most RchEXPs were not detected or had very low expression levels in fruit, such as RchEXPA2–3, RchEXPA8, RchEXPA10–12, RchEXPA14, RchEXPA16–24, RchEXPA27, RchEXPB2, and RchEXPLB1. They might play vital roles in the root, stem, leaf, and flowers, or respond to different kinds of stresses. Nevertheless, some RchEXPs exhibited remarkable effects on fruit ripening and softening (Figure 6). Especially, RchEXPA6 and RchEXPA26 might be notably involved in fruit expanding and softening. The expression level of RchEXPA6 at the immature BGI stage was 25.66 (FPKM, fragments per kilobase per million mapped fragments), while it rapidly increased to 6774.05 at the turning stage of GY, decreased at the YO stage, and then finally dramatically elevated to 14360.67 at the red stage, which was 559.65-fold in comparison with BGI (Figure 6a). The qPCR result verified its strong expression in the red stage (Figure 6c). The expression levels of RcEXPA6 were related to fruit size, fresh weight, and firmness throughout the whole fruit development. In addition, RchEXPA6 kept the highest expression level through the softening process from 70% to 90% ripe (Figure 6b). Simultaneously, the expression pattern of RchEXPA26 was in line with RchEXPA6 (Figure 6a–d). Oppositely, the expression levels of RchEXPA5 and RchEXPA15 were at high values at the beginning of the fruit set and earlier stages and decreased at later stages (Figure 6a,b,e,f). The FPKM of RchEXPA5 and RchEXPA15 at the BGI stage were 348.06 (449.34/1.29) and 146.04 (186.93/1.28) folds in comparison with the red stage, respectively. The RchEXPA7 had relatively high expression levels from BG to 80% Re and then sharply reduced to a low level at 90% Re. Therefore, RchEXPA5, RchEXPA7, and RchEXPA15 mainly participated in the process of cell expansion and fruit ripening. Therefore, red fruit from different lineages with different fruit sizes and different hardness were further analyzed. The fresh weights and firmness of L1, L2, L3, L6, L7, L14, L20, L26, L20⊗ (selfing), L3 × L7, L3 × L20, and L14 × L3 were measured (Figure 7a,b). The fruit of L3 × L7 was the smallest, and the fruit of L20⊗ was the biggest (Figure 7a). The 90% red fruit of L26, L20⊗, L1, L3 × L20, and L2 were relative hard, while the fruit of L3 × L7, L3, L6, L7, and L14 × L3 were softer (Figure 7b). The expression levels of RchEXP6 and RchEXP26 were relatively higher in softer fruits, while the expression levels of RchEXP7 and RchEXP15 were relatively higher in bigger fruits (Figure 7c–f). Pearson correlation analysis showed that RchEXP6 and RchEXP26 had significantly negative correlations with hardness, while RchEXP7 and RchEXP15 positively correlated with fresh weight (Figure 7g). These results verified the essential functions of RchEXPA6 and RchEXPA26 for fruit expanding and softening, while RchEXP7 and RchEXP15 mainly participated in fruit expansion and ripening.
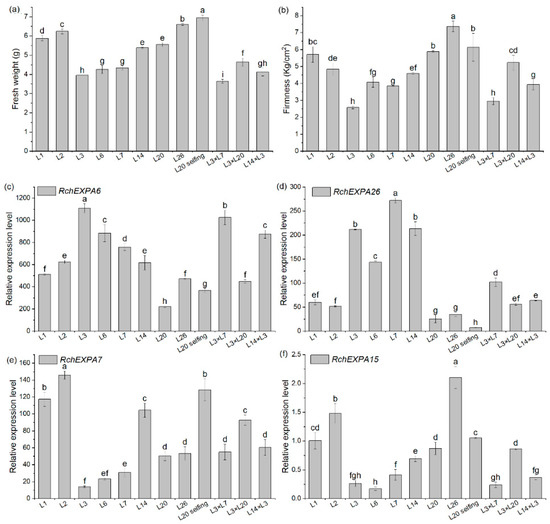
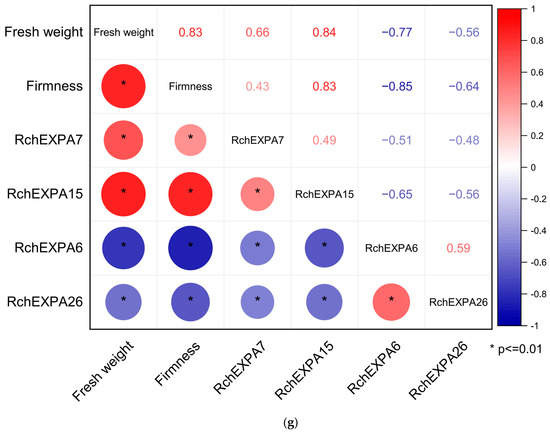
Figure 7.
Biomass and real-time quantitative PCR of RchEXPs in red fruit of different R. chingii lineages. (a) Fresh weight (Fw); (b) fruit firmness; (c–f) qPCR of RchEXPA6, RchEXPA26, RchEXPA7, and RchEXPA15. Different lower cases mean a significant difference at p < 0.05 by a one-way analysis of variance with the LSD method. (g) Pearson correlation analysis of RchEXPs and fruit fresh weight and firmness based on the qPCR data of red fruits from different lineages. “*” means a significant difference at p ≤ 0.05.
These experimental results were greatly consistent with the bioinformatics analysis. In Figure 2, the phylogenetic relationships confirmed that RchEXPA6 and RchEXPA26 were in cluster 6, and this cluster was composed of only these two proteins. Moreover, in Figure 4, syntenic relationships also predicted the collinear and paralog gene pairs of RchEXPA6 and RchEXPA26, suggesting the synergistic and complementary of the two EXPs. Simultaneously, RchEXPA7 and RchEXPA15 were in cluster 7, whereas RchEXPA5 was in cluster 5. In Figure 1, these three proteins were gathered in the same upper cluster. RchEXPA7 and RchEXPA15 showed a syntenic relationship. PPI analysis showed the interaction among RchEXPA6, RchEXPA26, PME, PG, ETR, and EIN2 (Figure 5b). The consistent functions of the two expansin genes RchEXPA6 and RchEXPA26, or the three expansin genes RchEXPA5, RchEXPA7, and RchEXPA15, were first identified in this study. Unveiling the action mechanism and regulatory pathways of these vital EXPs would be vital for genetic modification and biotechnological breeding for fruit quality improvement and softening delay.
3. Discussion
The plant cell wall is pivotal for cell size, shape, and strength, and affects cell division and differentiation, growth rate, and functional realization [23,26]. It can be hydrolyzed, modified, or reconstructed by a series of enzymic and non-enzymic proteins. Expansins (EXPs) are noncatalytic structure proteins that are predominately located in the cell wall. They are involved in almost all aspects of plant growth and development, from germination to fruiting, by loosening the cell walls [22,24,25,27]. They also play vital roles in stress responses [22,28,29]. In total, 35 and 114 EXPs have been identified in woodland strawberry Fragaria vesa and octoploid-cultivated strawberry F. ananassa with distinct expression patterns [21,25]. In this study, we identified 33 EXPs from the R. chingii genome for the first time (Table 1, Figure 1, Figure 2 and Figure 3). Transcriptomic and quantitative PCR profiles verified that RchEXPA5, RchEXPA7, and RchEXPA15 were synergistically involved in fruit expansion and ripening, while RchEXPA6 and RchEXPA26 might be essential for fruit ripening and softening (Figure 6). Our new findings laid a molecular foundation for improving fruit quality, controlling fruit firmness, and extending the storage life of R. chingii edible red fruit.
The expansins can be divided into four subfamilies according to the phylogenetic analysis. EXPA members are the main components of expansins in all kinds of plants, while EXPB members might mainly exist in Gramineae monocotyledons. In dicotyledonous plants, Arabidopsis has twenty-six, six, three, and one members of EXPA, EXPB, EXLA, and EXLB, respectively; polar (Populus) contains twenty-seven, three, two, and four members of the four subfamilies, respectively; and F. vesa consists of twenty-seven EXPAs, five EXPBs, one EXPLA, and two EXPLBs, respectively [25]. The proportion of EXPs in R. chingii is similar to F. vesa. RchEXPs are composed of twenty-seven EXPAs, three EXPBs, one EXPLA, and two EXPLBs (Table 1, Figure 2). These results suggest that the EXPA subfamily in dicotyledons might be expanded far more than the other three subfamilies. Nevertheless, Gramineae plants contain higher numbers of EXPB members. There are 34 EXPAs and 19 EXPBs in rice, 36 EXPAs and 48 EXPBs in maize, and 45 EXPAs and 29 EXPBs in moso bamboo [22,25]. The cell wall is fundamentally made up of different groups of polysaccharides, such as cellulose, hemicellulose, and pectin. Loosening of the polysaccharides network is the direct cause of cell wall looseness and cell expansions [30]. Xyloglucan is the main hemicellulose in the primary wall of dicotyledonous and non-gramineous monocotyledonous plants, comprising up to 20% of the wall dry matter [31]. However, the content of xyloglucan in gramineous plants was very low. EXPAs promote movement and separation of microfibrils by means of molecular creep for the dissociation and slippage of xyloglucans, while EXPBs work on another glycan, maybe xylan [19,23,32]. Among the xylans, arabinoxylan (AX) mainly exists in the grain of gramineous plants, glucuronoarabinoxylan (GAX) exists in vegetative tissues of gramineous plants and the primary cell wall of dicotyledonous, and glucuronoxylan (GX) mainly exists in the secondary wall of dicotyledonous and non-gramineous monocotyledonous plants [33]. Therefore, the various compositions of plant cell wall polysaccharides and the distinct action modes of EXPs might determine the proportion and numbers of EXP subfamilies.
Furthermore, it has been reported that xyloglucan disassembly might be an early event in fruit softening [24], and firmness is one of the decisive indexes of fruit quality and postharvest shelf life. The effects of expansins on fruit ripening and softening have been described in various horticultural plants. For example, the overexpression of tomato Slexp1 hastened the softening process, while Slexp1-6 and Slexp1-7 mutants enhanced fruit firmness and could be stored for longer periods [34]. FaEXP2 and FaEXP5 were predominantly expressed in cultivated octaploid strawberry fruit, and the significant increase in the FaEXP5 expression level was in close correlation with the rapid decrease in fruit firmness [19,35]. The above expression pattern of EXP5 was also observed in Chilean strawberries [36]. In addition, FaEXP7, FaEXP88, and FaEXP114 were also candidate genes for softening activation [20,21]. Our experiment evaluated the expression pattern of RchEXPs during fruit development and ripening and found that three expansin genes of RchEXPA5, RchEXPA7, and RchEXPA15 were highly expressed at earlier stages but decreased at later stages, suggesting that they were predominantly involved in cell expansions and fruit ripening; while the expressions of the two expansin genes, RchEXPA6 and RchEXPA26, were dramatically activated at red stage, indicating their essential role in fruit softening (Figure 6). In F. vesa, both Dong et al. [25] and Mu et al. [21] reported that three FvEXPs were highly expressed in ripening fruit. Among these, FvEXPA9 (FveEXP11) and FvEXPA12 had especially high expression levels in the turning stage, while FvEXPA27 (FveEXP33) had especially high expression levels in the red stage and over-ripening stage. Phylogenetic analysis and synteny profiles showed that different expansins from diverse species belonging to the same clade or collinearity contributed to similar functions [21]. As expected, RchEXPA6 had collinearity with RchEXPA26 and FvEXPA27 (Figure 4, Table S3), and the encoded protein had high structural similarity with FvEXP27 (Figure 2, Figure S1). Therefore, they exhibited the same function (Figure 6). Simultaneously, there were collinear relationships and close evolutionary relationships between RchEXPA5 and FvEXPA9, RchEXPA15 and FvEXPA12, as well as RchEXPA15 and RchEXPA7 (Figure 4), resulting in their consistent function. Overall, our results elucidated the key expansin encoding genes in fruit ripening and softening.
The regulator of expansins in R. chingii was also predicted through promoter and PPI analyses. Promoters of most RchEXPs showed multiple ABA-responsive element sites, and some of them also possessed Eth-responsive elements (Figure 5). Numerous studies have revealed that ethylene and ABA both could regulate fruit ripening in strawberries [14,37]. ERE was found upstream of FvEXPA9 and FvEXPA27 [25]. In this study, ERE was found in the promoter region of RchEXPA7 and RchEXPA26, whereas it was absent in promoters of RchEXPA5, RchEXPA6, and RchEXPA15 (Figure 5). Therefore, the ripening of R. chingii might be controlled by both ABA and Eth, and the RchEXPs act coordinately for fruit texture. It has been reported that the high relative expression of FvEXP12 in the turning stage was attributed to the auxin-responsive element in its promoter [25]. Interestingly, RchEXPA7 had collinearity with FvEXP12 (Table S3), and RchEXPA7 was also the orthologous gene for FvEXP12. Hence, the high expression level of RchEXPA7 in the turning stage consisted of strawberry expansin.
In addition, many pieces of evidence also supported the effects of expansins on stress resistance. TaEXPA2 regulated by MYB transcription factor enhanced drought tolerance in wheat [29]. PeEXPA19, highly expressed in the leaves and roots of moso bamboo, had three MBS elements in its promoter and exhibited drought resistance, whereas PeEXPA44 showed downregulated expression under PEG stress [22]. Therefore, the functions of other expansins in R. chingii need to be further investigated. All expansins are distributed in different plant tissues and have distinct roles, whereas they are synergistically involved in plant growth, development, maturity, senescence, and response to stresses.
4. Conclusions
In conclusion, this study performed the first genome-wide identification of 33 expansins and their encoding genes in the unique Chinese raspberry, Rubus chingii Hu. The molecular characterization, biological function, and evolutionary pattern were elucidated. The results confirmed that three RchEXPs, including RchEXP5, RchEXP7, and RchEXP15, and two RchEXPs, RchEXP6 and RchEXP26, played pivotal roles in fruit ripening and softening, respectively. Our study gives a comprehensive understanding of the expansin family in R. chingii and screens out the candidate genes for molecular breeding to slow down fruit softening and extend storage life.
5. Materials and Methods
5.1. Plant Material
Rubus chingii Hu seedlings were planted in our germplasm resource orchard (28°73′39″ N, 121°09′11″ E) for more than 5 years. Fruits from uniform 3-year seedlings of different superior lineages and crossing progenies were selected. For transcriptomic sequencing, every 20 fruits from the 4 representative stages, including BGI (big green I, 21 DPA, day-post-anthesis), GY (green-to-yellow, 42 DPA), YO (yellow-to-orange, 48 DAP), and the Re (red, 54 DPA, 90% ripe) of L7 (softer lineage with low firmness in red stages; other detailed characteristics of different lineages have been described in our previous work [2]) were harvested. Moreover, the ripening fruits of 70% red, 80% red, and 90% red from L26 (harder lineage with relatively high firmness) in red stages were also harvested for RNA-seq. For further real-time quantitative PCR assays, every 50 fruits in the eight stages from L7 were harvested in 2021, including SG (small green, 7 DPA), MG (medium green, 14 DPA), BGI, BGII (big green II, 28 DPA), BGIII (big green III, 35 DPA), GY, YO, and Re. Meanwhile, red fruit from different lineages were investigated, including L1, L2, L3, L6, L7, L14, L20, L26, L20⊗ (selfing), L3 × L7, L3 × L20, and L14 × L3. The fresh weight and fruit size were measured using electronic balance and vernier calipers. Firmness was determined using a digital durometer (FHT-05, Lantai, China).
5.2. Genome-Wide Identification of Expansin Family in R. chingii and Phylogenetic Analysis
The genome and protein sequences (v1.0) of R. chingii were obtained from the Genome Database for Rosaceae (GDR) (https://www.rosaceae.org/species/rubus/all) (accessed on 27 May 2021) [6]. In addition, Rubus idaeus Joan J Genome v2.0 (accessed on 15 September 2022), Rubus occidentalis whole genome assembly v3.0 (accessed on 30 April 2018), and Fragaria vesca Genome v4.0.a2 (accessed on 9 May 2019)were also downloaded from GDR. The PF03330 (DPBB_1 domain) and PF01357 (Expansin_C) domains were applied for the Hidden Markov Model (HMM) search with E-value < l0−5. The primary gene ID and protein sequences were extracted by TBtools, and then they were examined by the online tools in the Conserved Domain Database (CDD) (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi, accessed on 27 May 2021) and PFAM (https://pfam-legacy.xfam.org/search/sequence, accessed on 27 May 2021). Proteins with incomplete C or N terminal and incomplete domain DPPB_1 or Expansin_C were discarded. A maximum-likelihood phylogeny (ML-tree) of R. chingii, R. idaeus, R. occidentalis, F. vesca, and A. thaliana was constructed by TBtools and visualized by MEGA 11 and iTOL (https://itol.embl.de/, accessed on 27 May 2021). The expansins from the three Rubus species were renamed according to their chromosomal position and their similarity with F. vesca. The isoelectric point (pI) and molecular weight (MW) were calculated through ExPaSy (https://web.expasy.org/compute_pi/, accessed on 27 May 2021), and subcellular localization was predicted using online software on the Plant-mPLoc server (2.0 version) (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/#, accessed on 27 May 2021).
5.3. Motifs, Gene Structures, Chromosomal Location, and Synteny Analysis of Expansins in R. chingii
The gene structures of exon/intron were mapped using TBtools according to the genome annotation “Rubus_chingii_Hu.gff” file. Conserved motifs were identified with the online Multiple Em for Motif Elicitation (MEME, https://meme-suite.org/meme/tools/meme, accessed on 27 May 2021) tool with the parameter setting of the maximum number of motifs equal to 10. Gene density was profiled, and gene locations were visualized by TBtools. The synteny analysis in R. chingii genome and synteny analysis between R. chingii and F. vesa genomes were calculated with MCScanX (CPU for BlastP: 2; E-value: 1 × 10−10; Num of BlastHits:5) [38]. The syntenic diagram was visualized by an advanced circos and dual synteny plot using TBtools.
5.4. Cis-Regulatory Elements Analysis and Protein–Protein Interaction Network Construction of Expansins in R. chingii
The 2000 bp upstream genomic sequences of the expansin genes were extracted as the putative promoters to examine the cis-elements by PlantCARE (https://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 27 May 2021). The predicted cis-acting elements and their numbers for phytohormone response, plant growth, and development, were summarized, respectively. The protein–protein interaction network (PPI) was analyzed based on transcriptome data, and the protein–protein interaction network (PPI) was constructed using cytoscape_v3.10.1.
5.5. Expression Analyses of RchExps during Developmental Stages
Transcriptomic sequencing of fruits from the four representative stages (BG, GY, YO, and Re) of L7 was carried out in 2017 using the Illumina (HiSeq X-Ten, San Diego, CA, USA) platform from the Beijing Genomics Institute (BGI, Wuhan, China). Approximately 536.05 Mb total clean reads with high quality (Q20 > 98.5%, Q30 > 82.5%) were obtained from all samples (4 stages in triplicate) [5]. Then, the de nova sequencing data were reanalyzed in 2021, according to the later published R. chingii genome [6]. The expression levels of RchEXPs were expressed by Fragments Per Kilobase per Million mapped fragments (FPKM) value (=106 C/(NL/103) and mapped as a heat map using TBtools. Moreover, RNA-seq of the ripening fruits of 70% red, 80% red, and 90% red from L26 was performed in 2021 using the DNBseq platform by the Beijing Genomics Institute (BGI, Wuhan, China). A total of 380.41 million total clean reads with high quality were obtained from all 9 samples (3 stages in triplicate), and the FPKM values of RchEXPs were extracted and mapped with the above method.
For a further real-time quantitative PCR (aPCR) assay, fruits of eight stages from L7 (SG, MG, BGI, BGII, BGIII, GY, YO, and Re) were harvested in 2021. A total of about 50 fruits at each stage were collected, and RNA was isolated using an OminiPlant RNA Kit (CW2598, CWBIO, Beijing, China). The cDNA synthesis by reverse transcription and qPCR determination were performed according to the previous methods [5]. The relative expression level was calculated using the 2−∆∆Ct method. All the experiments were carried out in triplicate to quadruplicate, and the LSD test was selected for one-way analysis of variance using SPSS 26. Moreover, red fruits (90% ripe, edible) from different lineages, including L1, L2, L3, L6, L7, L14, L20, L26, L20⊗ (selfing), L3 × L7, L3 × L20, and L14 × L3, were harvested, and the qPCR of RchEXP6, RchEXP7, RchEXP15, and RchEXP26 in these red fruits were determined with the above method.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants13030431/s1.
Author Contributions
Conceptualization, X.L. and W.Z.; investigation, Z.C., D.S., Y.C., H.H., J.J. (Junfeng Jiang), and F.W.; resources, J.J. (Jingyong Jiang); writing-original draft, Z.C. and D.S.; methodology, Y.S. and Z.C.; data curation, X.W.; writing–review and edition, X.L. and W.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Basic Public Welfare Research Project of Zhejiang Province (LGN22C020001), the Taizhou Science and Technology Project (21nya14), and the College Students’ innovation and entrepreneurship training program (202310350029, S202310350160).
Data Availability Statement
Data are contained within the article and supplementary materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sheng, J.Y.; Wang, S.Q.; Liu, K.H.; Zhu, B.; Zhang, Q.Y.; Qin, L.P.; Wu, J.J. Rubus chingii Hu: An overview of botany, traditional uses, phytochemistry, and pharmacology. Chin. J. Nat. Med. 2020, 18, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Jiang, J.; Li, X.; Xie, Y.; Jin, Z.; Wang, X.; Li, Y.; Zhong, Y.; Lin, J.; Yang, W. Bioactive compounds and fruit quality of Chinese raspberry, Rubus chingii Hu varied with genotype and phenological phase. Sci. Hortic. 2021, 281, 109951. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Fupenzi, Rubi Fructus. Pharmacopoeia of the People’s Republic of China (Edition 2020); China Medical Science Press: Beijing, China, 2020. [Google Scholar]
- Yu, G.; Luo, Z.; Wang, W.; Li, Y.; Zhou, Y.; Shi, Y. Rubus chingii Hu: A review of the phytochemistry and pharmacology. Front. Pharmacol. 2019, 10, 799. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Jiang, J.; Shu, L.; Li, X.; Huang, J.; Qian, B.; Wang, X.; Li, X.; Chen, J.; Xu, H. Combined transcriptomic and metabolic analyses reveal potential mechanism for fruit development and quality control of Chinese raspberry (Rubus chingii Hu). Plant Cell Rep. 2021, 40, 1923–1946. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lei, T.; Han, G.; Yue, J.; Zhang, X.; Yang, Q.; Ruan, H.; Gu, C.; Zhang, Q.; Qian, T.; et al. The chromosome-scale reference genome of Rubus chingii Hu provides insight into the biosynthetic pathway of hydrolyzable tannins. Plant J. 2021, 107, 1466–1477. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Hu, Y.; Li, J.; Cai, J.; Qiu, Y.; Dong, C. Anti-inflammatory effect of a novel pectin polysaccharide from Rubus chingii Hu on colitis mice. Front. Nutr. 2022, 9, 868657. [Google Scholar] [CrossRef]
- Wang, J.; Xiao, X.; Zhou, N.; Zhao, M.; Lang, S.; Ren, Q.; Wang, D.; Fu, H. Rubochingosides A—J, labdane-type diterpene glycosides from leaves of Rubus chingii. Phytochemistry 2023, 210, 113670. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, D.; Zhu, B.; Wang, S.; Xu, Y.; Zhang, C.; Yang, H.; Wang, S.; Liu, P.; Qin, L.; et al. Rubus chingii Hu. unripe fruits extract ameliorates carbon tetrachloride-induced liver fibrosis and improves the associated gut microbiota imbalance. Chin. Med. 2022, 17, 56. [Google Scholar] [CrossRef]
- Zhong, J.; Wang, Y.; Li, C.; Yu, Q.; Xie, J.; Dong, R.; Xie, Y.; Li, B.; Tian, J.; Chen, Y. Natural variation on free, esterified, glycosylated and insoluble-bound phenolics of Rubus chingii Hu: Correlation between phenolic constituents and antioxidant activities. Food Res. Int. 2022, 162, 112043. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Z.; Guo, Q.; Gao, X.; Ma, Q.; Xue, Z.; Ferri, N.; Zhang, M.; Chen, H. Identification of ellagitannins in the unripe fruit of Rubus chingii Hu and evaluation of its potential antidiabetic activity. J. Agric. Food Chem. 2019, 67, 7025–7039. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Jin, L.; Chen, Z.; Jiang, J.; Jackson, A. Development of fruit color in Rubus chingii Hu (Chinese raspberry): A story about novel offshoots of anthocyanin and carotenoid biosynthesis. Plant Sci. 2021, 311, 110996. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.M.; Bassil, N.V.; Dossett, M.; Leigh Worthington, M.; Graham, J. Genetic and genomic resources for Rubus breeding: A roadmap for the future. Hortic. Res. 2019, 6, 116. [Google Scholar] [CrossRef]
- Li, B.J.; Grierson, D.; Shi, Y.; Chen, K.S. Roles of abscisic acid in regulating ripening and quality of strawberry, a model non-climacteric fruit. Hortic. Res. 2022, 9, uhac089. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Manghwar, H.; Hu, W. Study on supergenus Rubus L.: Edible, medicinal, and phylogenetic characterization. Plants 2022, 11, 1211. [Google Scholar] [CrossRef]
- VanBuren, R.; Bryant, D.; Bushakra, J.M.; Vining, K.J.; Edger, P.P.; Rowley, E.R.; Priest, H.D.; Michael, T.P.; Lyons, E.; Filichkin, S.A.; et al. The genome of black raspberry (Rubus occidentalis). Plant J. 2016, 87, 535–547. [Google Scholar] [CrossRef]
- Hua, Y.; Dai, B.; Luo, Y.; Ding, Y. Integrated analysis of multiple metabolome and transcriptome revealed the accumulation of flavonoids and associated molecular regulation mechanisms in Rubus chingii Hu at different developmental stages. Plant Physiol. Biochem. 2023, 204, 108085. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Huang, J.; Ruan, H.; Qian, W.; Fang, Z.; Gu, C.; Zhang, N.; Liang, Y.; Wang, Z.; Gao, L.; et al. Competition between FLS and DFR regulates the distribution of flavonols and proanthocyanidins in Rubus chingii Hu. Front. Plant Sci. 2023, 14, 1134993. [Google Scholar] [CrossRef]
- Valenzuela-Riffo, F.; Parra-Palma, C.; Ramos, P.; Morales-Quintana, L. Molecular and structural insights into FaEXPA5, an alpha-expansin protein related with cell wall disassembly during ripening of strawberry fruit. Plant Physiol. Biochem. 2020, 154, 581–589. [Google Scholar] [CrossRef]
- Ren, Y.; Li, B.; Jia, H.; Yang, X.; Sun, Y.; Shou, J.; Jiang, G.; Shi, Y.; Chen, K. Comparative analysis of fruit firmness and genes associated with cell wall metabolisms in three cultivated strawberries during ripening and postharvest. Food Qual. Saf. 2023, 7, fyad020. [Google Scholar] [CrossRef]
- Mu, Q.; Li, X.; Luo, J.; Pan, Q.; Li, Y.; Gu, T. Characterization of expansin genes and their transcriptional regulation by histone modifications in strawberry. Planta 2021, 254, 21. [Google Scholar] [CrossRef]
- Jin, K.M.; Zhuo, R.Y.; Xu, D.; Wang, Y.J.; Fan, H.J.; Huang, B.Y.; Qiao, G.R. Genome-wide identification of the expansin gene family and its potential association with drought stress in moso bamboo. Int. J. Mol. Sci. 2020, 21, 9491. [Google Scholar] [CrossRef] [PubMed]
- Sampedro, J.; Cosgrove, D.J. The expansin superfamily. Genome Biol. 2005, 6, 242. [Google Scholar] [CrossRef] [PubMed]
- Marowa, P.; Ding, A.; Kong, Y. Expansins: Roles in plant growth and potential applications in crop improvement. Plant Cell Rep. 2016, 35, 949–965. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Zou, X.; Gao, Q. Genome-wide identification of expansin in Fragaria vesca and expression profiling analysis of the FvEXPs in different fruit development. Gene 2022, 814, 146162. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Plant expansins: Diversity and interactions with plant cell walls. Curr. Opin. Plant Biol. 2015, 25, 162–172. [Google Scholar] [CrossRef]
- Samalova, M.; Gahurova, E.; Hejatko, J. Expansin-mediated developmental and adaptive responses: A matter of cell wall biomechanics? Quant. Plant Biol. 2022, 13, e11. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, B.; Li, C.; Lei, C.; Kong, C.; Yang, Y.; Gong, M. A comprehensive expression analysis of the expansin gene family in potato (Solanum tuberosum) discloses stress-responsive expansin-like B genes for drought and heat tolerances. PLoS ONE 2019, 14, e0219837. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, G.; An, J.; Li, Q.; Chen, Y.; Zhao, X.; Wu, J.; Wang, Y.; Hao, Q.; Wang, W. Expansin gene TaEXPA2 positively regulates drought tolerance in transgenic wheat (Triticum aestivum L.). Plant Sci. 2020, 298, 110596. [Google Scholar] [CrossRef]
- Bashline, L.; Lei, L.; Li, S.; Gu, Y. Cell wall, cytoskeleton, and cell expansion in higher plants. Mol. Plant 2014, 7, 586–600. [Google Scholar] [CrossRef]
- Culbertson, A.T.; Smith, A.L.; Cook, M.D.; Zabotina, O.A. Truncations of xyloglucan xylosyltransferase 2 provide insights into the roles of the N- and C-terminus. Phytochemistry 2016, 128, 12–19. [Google Scholar] [CrossRef]
- Wang, T.; Park, Y.B.; Caporini, M.A.; Rosay, M.; Zhong, L.; Cosgrove, D.J.; Hong, M. Sensitivity-enhanced solid-state NMR detection of expansin’s target in plant cell walls. Proc. Natl. Acad. Sci. USA 2013, 110, 16444–16449. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Chen, F.; Luo, J.; Zeng, W.; Xu, W. The biosynthesis of plant cell wall xylan and its application. Chin. Bull. Bot. 2023, 58, 316–334. (In Chinese) [Google Scholar] [CrossRef]
- Minoia, S.; Boualem, A.; Marcel, F.; Troadec, C.; Quemener, B.; Cellini, F.; Petrozza, A.; Vigouroux, J.; Lahaye, M.; Carriero, F.; et al. Induced mutations in tomato SlExp1 alter cell wall metabolism and delay fruit softening. Plant Sci. 2016, 242, 195–202. [Google Scholar] [CrossRef]
- Dotto, M.C.; Martínez, G.A.; Civello, P.M. Expression of expansin genes in strawberry varieties with contrasting fruit firmness. Plant Physiol. Biochem. 2006, 44, 301–307. [Google Scholar] [CrossRef]
- Valenzuela-Riffo, F.; Gaete-Eastman, C.; Stappung, Y.; Lizana, R.; Herrera, R.; Moya-León, M.A.; Morales-Quintana, L. Comparative in silico study of the differences in the structure and ligand interaction properties of three alpha-expansin proteins from Fragaria chiloensis fruit. J. Biomol. Struct. Dyn. 2019, 37, 3245–3258. [Google Scholar] [CrossRef]
- Figueroa, N.E.; Gatica-Meléndez, C.; Figueroa, C.R. Ethylene application at the immature stage of Fragaria chiloensis fruit represses the anthocyanin biosynthesis with a concomitant accumulation of lignin. Food Chem 2021, 358, 129913. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).