



Genetic Mapping of Tolerance to Bacterial Stem Blight Caused by Pseudomonas syringae pv. syringae in Alfalfa (Medicago sativa L.)
 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
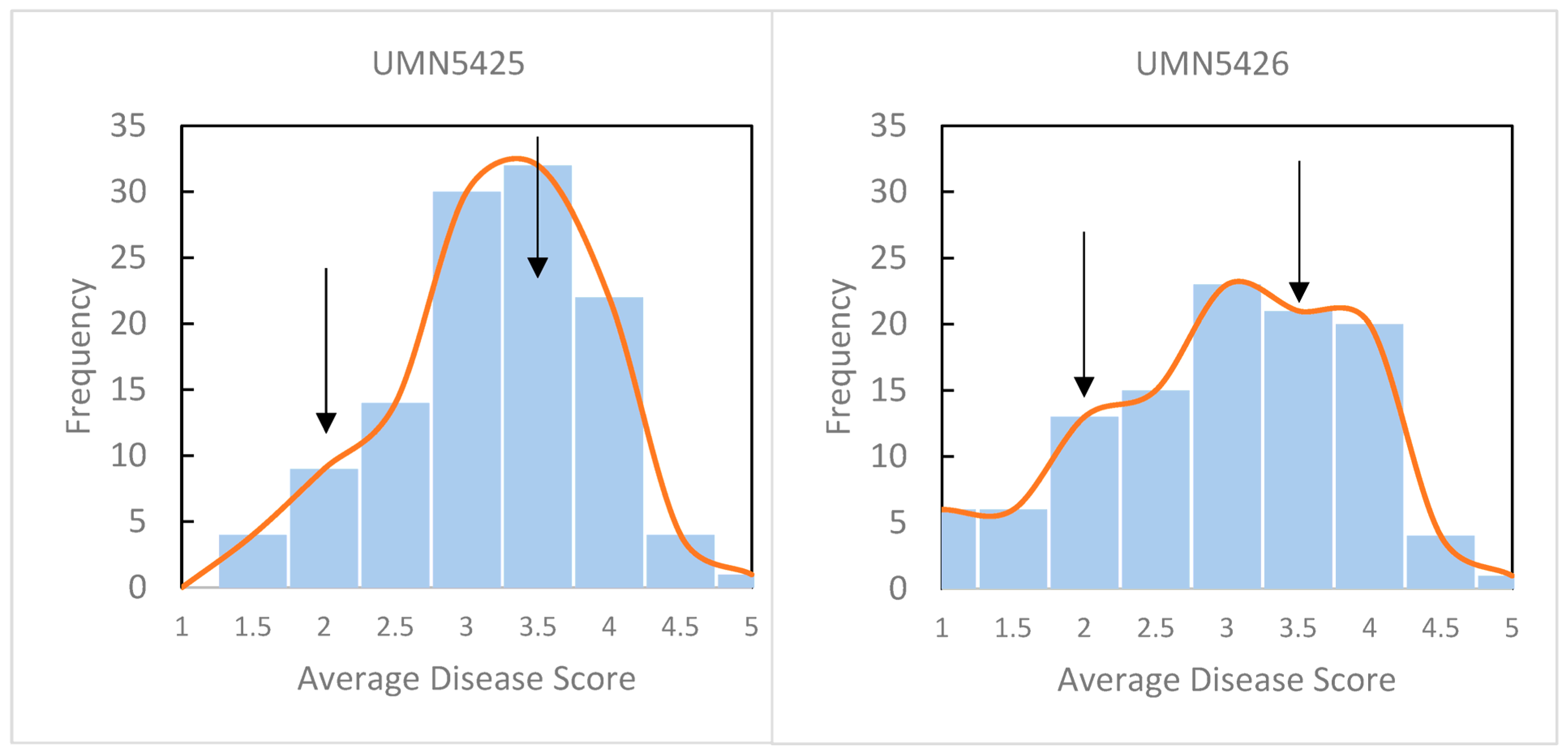
2.1. Disease Phenotypes of F1 Populations
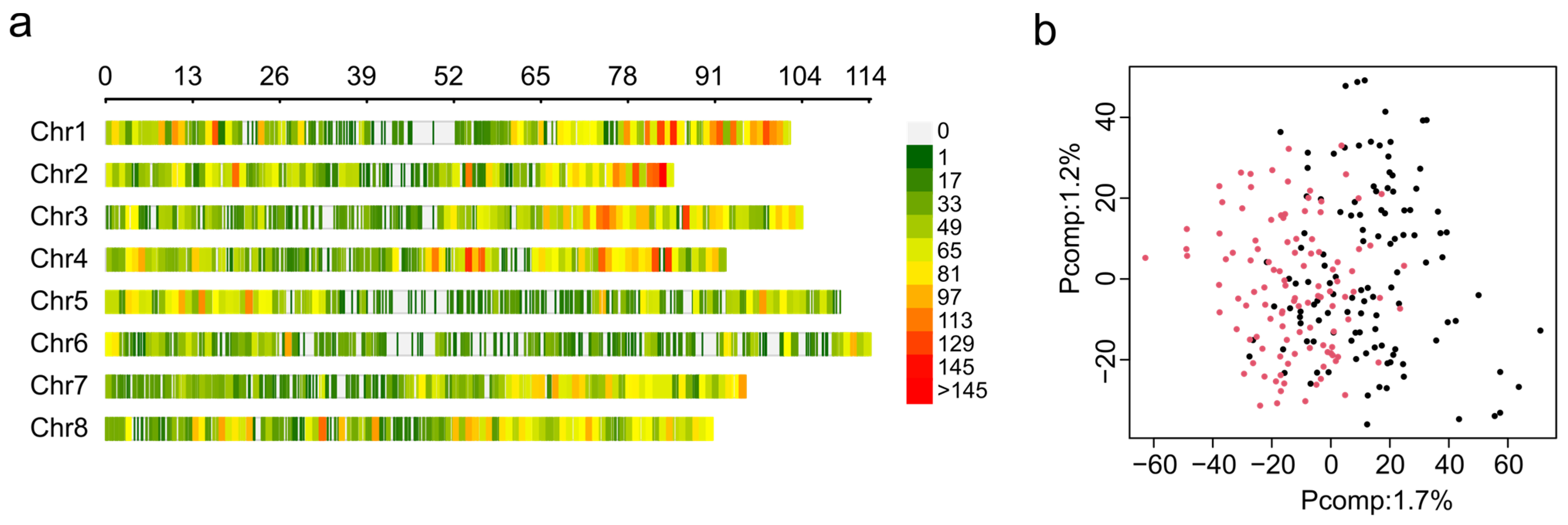
2.2. Genotyping Coverage and Population Structure
2.3. Genome-Wide Association Studies
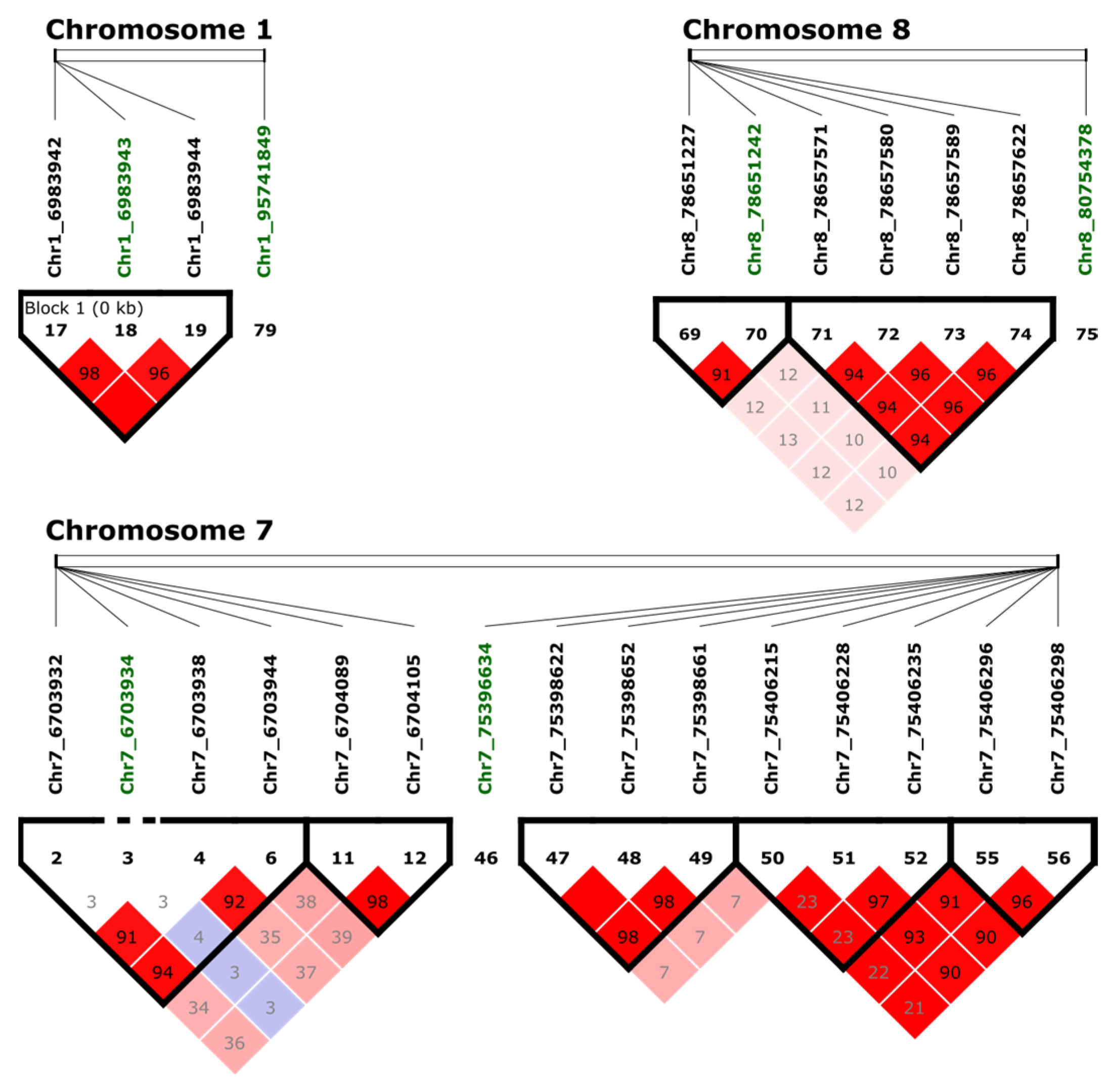
2.4. Examining Linkage Disequilibrium (LD)
3. Discussion
4. Materials and Methods
4.1. Bacterial Culture Conditions
4.2. Development of Mapping Populations and Plant Growth Conditions
4.3. Leaf Inoculation and Disease Scoring
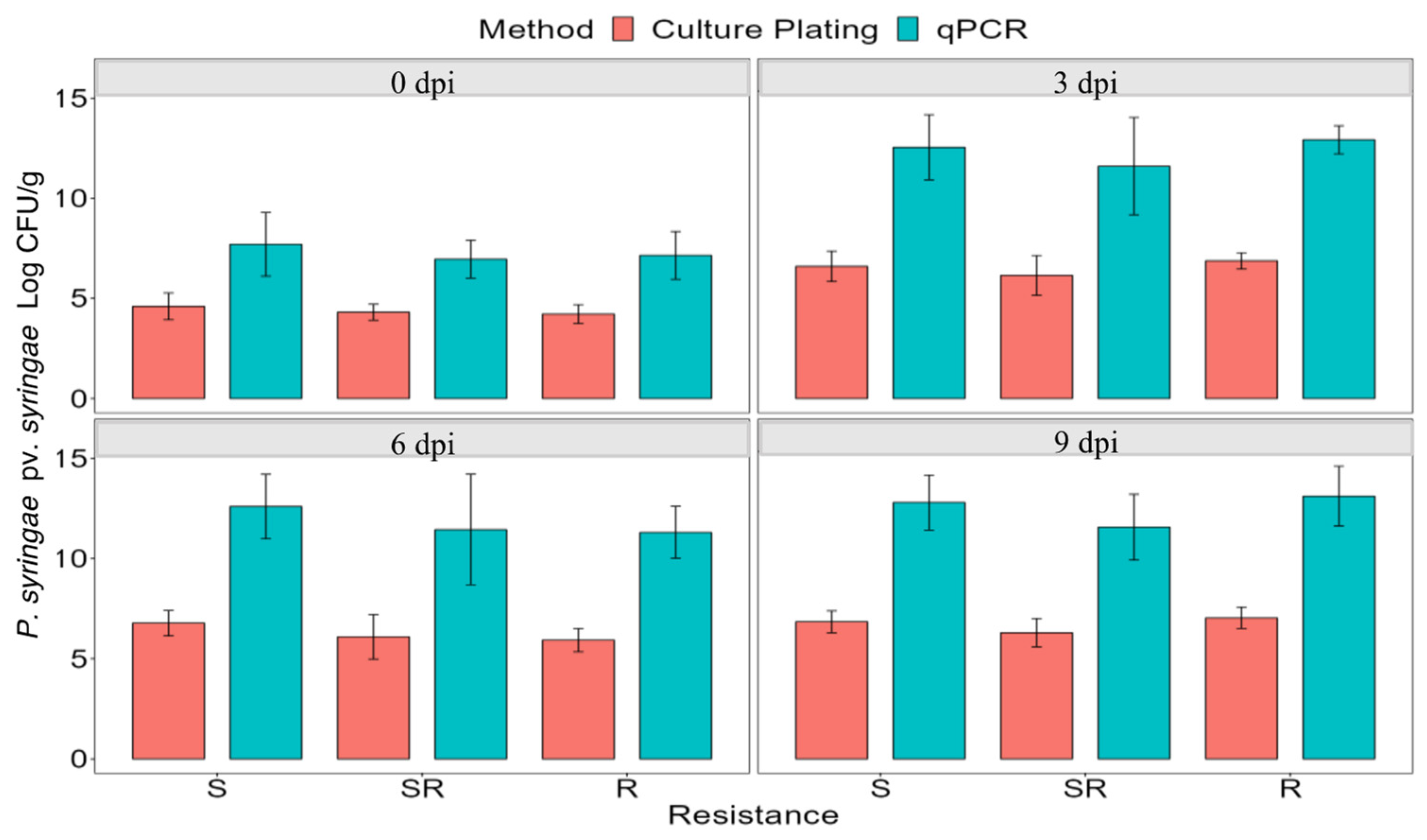
4.4. Pathogen Quantification
4.5. DNA Extraction and Sequencing
4.6. GBS and Variant Calling
4.7. Association Mapping and Annotation
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernandez, A.L.; Sheaffer, C.C.; Tautges, N.E.; Putnam, D.H.; Hunter, M.C. Alfalfa, Wildlife, and the Environment, 2nd ed.; National Alfalfa and Forage Alliance: St. Paul, MN, USA, 2019. [Google Scholar]
- Samac, D.A.; Studholme, D.J.; Ao, S. Characterization of the bacterial stem blight pathogen of alfalfa, Pseudomonas syringae pv. syringae ALF3. Phytopathology 2014, 104 (Suppl. S3), 102. [Google Scholar]
- Lipps, S.M.; Lenz, P.; Samac, D.A. First report of bacterial stem blight of alfalfa caused by Pseudomonas viridiflava in California and Utah. Plant Dis. 2019, 103, 3274. [Google Scholar] [CrossRef]
- Nemchinov, L.G.; Shao, J.; Lee, M.N.; Postnikova, O.A.; Samac, D.A. Resistant and susceptible responses in alfalfa (Medicago sativa) to bacterial stem blight caused by Pseudomonas syringae pv. syringae. PLoS ONE 2017, 12, e0189781. [Google Scholar] [CrossRef] [PubMed]
- Hamedan, E.; Harighi, B. Genetic diversity of Pseudomonas syringae pv. syringae strains, causing bacterial stem blight disease of alfalfa in the Kurdistan province of Iran. J. Plant Pathol. 2014, 96, 303–307. [Google Scholar]
- Lipps, S.M.; Castell-Miller, C.V.; Morris, C.E.; Ishii, S.; Samac, D.A. Diversity in Pseudomonas syringae and Pseudomonas viridiflava strains causing bacterial stem blight of alfalfa (Medicago sativa) in the United States. Phytopathology 2023. [Google Scholar] [CrossRef]
- Gray, F.A.; Hollingsworth, C.R. Bacterial stem blight. In Compendium of Alfalfa Diseases and Pests, 3rd ed.; Samac, D.A., Rhodes, L.H., Lamp, W.O., Eds.; American Phytopathological Society: St. Paul, MN, USA, 2015; pp. 60–62. [Google Scholar]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-Microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Yang, Y.; Wu, D.; Zhang, C. Plant immunity. Plant Signal. Behav. 2011, 6, 794–799. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- González, A.M.; Godoy, L.; Santalla, M. Dissection of resistance genes to Pseudomonas syringae pv. phaseolicola in UI3 common bean cultivar. Int. J. Mol. Sci. 2017, 18, 2503. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, K.H.; Van, K.; Kim, M.Y.; Lee, S.-H. Fine mapping of a resistance gene to bacterial leaf pustule in soybean. Theor. Appl. Genet. 2010, 120, 1443–1450. [Google Scholar] [CrossRef]
- Tahir, J.; Hoyte, S.; Bassett, H.; Brendolise, C.; Chatterjee, A.; Templeton, K.; Deng, C.; Crowhurst, R.; Montefiori, M.; Morgan, E.; et al. Multiple quantitative trait loci contribute to resistance to bacterial canker incited by Pseudomonas syringae pv. actinidiae in kiwifruit (Actinidia chinensis). Hortic. Res. 2019, 6, 101. [Google Scholar] [CrossRef] [PubMed]
- Tahir, J.; Brendolise, C.; Hoyte, S.; Lucas, M.; Thomson, S.; Hoeata, K.; McKenzie, C.; Wotton, A.; Funnell, K.; Morgan, E.; et al. QTL mapping for resistance to cankers induced by Pseudomonas syringae pv. actinidiae (Psa) in a tetraploid Actinidia chinensis kiwifruit population. Pathogens 2020, 9, 967. [Google Scholar] [CrossRef] [PubMed]
- Tanksley, S.D. Mapping polygenes. Annu. Rev. Genet. 1993, 27, 205–233. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Feng, H.; Jia, X.; Ma, S.; Ma, C.; Wang, Y.; Pan, S.; Chen, Q.; Xin, D.; Liu, C. Identifications of QTLs and candidate genes associated with Pseudomonas syringae responses in cultivated soybean (Glycine max) and wild soybean (Glycine soja). Int. J. Mol. Sci. 2023, 24, 4618. [Google Scholar] [CrossRef] [PubMed]
- Whitham, S.A.; Qi, M.; Innes, R.W.; Ma, W.; Lopes-Caitar, V.; Hewezi, T. Molecular soybean-pathogen interactions. Annu. Rev. Phytopathol. 2016, 54, 443–468. [Google Scholar] [CrossRef] [PubMed]
- Bernacki, M.J.; Czarnocka, W.; Szechyńska-Hebda, M.; Mittler, R.; Karpiński, S. Biotechnological potential of LSD1, EDS1, and PAD4 in the improvement of crops and industrial plants. Plants 2019, 8, 290. [Google Scholar] [CrossRef]
- Selote, D.; Shine, M.B.; Robin, G.P.; Kachroo, A. Soybean NDR1-like proteins bind pathogen effectors and regulate resistance signaling. New Phytol. 2014, 202, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shine, M.B.; Gao, Q.-M.; Navarre, D.; Jiang, W.; Liu, C.; Chen, Q.; Hu, G.; Kachroo, A. Enhanced disease susceptibility1 mediates pathogen resistance and virulence function of a bacterial effector in soybean. Plant Physiol. 2014, 165, 1269–1284. [Google Scholar] [CrossRef]
- Pagán, I.; García-Arenal, F. Tolerance of plants to pathogens: A unifying view. Annu. Rev. Phytopathol. 2020, 58, 77–96. [Google Scholar] [CrossRef]
- Kover, P.X.; Schaal, B.A. Genetic variation for disease resistance and tolerance among Arabidopsis thaliana accessions. Proc. Nat. Acad. Sci. USA 2002, 99, 11270–11274. [Google Scholar] [CrossRef]
- Velásquez, A.C.; Oney, M.; Huot, B.; Xu, S.; He, S.Y. Diverse mechanisms of resistance to Pseudomonas syringae in a thousand natural accessions of Arabidopsis thaliana. New Phytol. 2017, 214, 1673–1687. [Google Scholar] [CrossRef] [PubMed]
- Goss, E.M.; Bergelson, J. Fitness consequences of infection of Arabidopsis thaliana with its natural bacterial pathogen Pseudomonas viridiflava. Oecologia 2007, 152, 71–81. [Google Scholar] [CrossRef]
- Duque-Jaramillo, A.; Ulmer, N.; Alseekh, S.; Bezrukov, I.; Fernie, A.R.; Skirycz, A.; Karasov, T.L.; Weigel, D. The genetic and physiological basis of Arabidopsis thaliana tolerance to Pseudomonas viridiflava. New Phytol. 2023, 240, 1961–1975. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, J.; Somerville, S.C.; Hammerschmidt, R. Identification of a gene in Arabidopsis thaliana that controls resistance to Xanthomonas campestris pv. campestris. Physiol. Mol. Plant Pathol. 1991, 38, 57–65. [Google Scholar] [CrossRef]
- Buell, C.R.; Somerville, S.C. Use of Arabidopsis recombinant inbred lines reveals a monogenic and a novel digenic resistance mechanism to Xanthomonas campestris pv campestris. Plant J. 1997, 12, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Kralik, P.; Ricchi, M. A basic guide to real time PCR in microbial diagnostics: Definitions, parameters, and everything. Front. Microbiol. 2017, 8, 108. [Google Scholar] [CrossRef] [PubMed]
- Bock, C.H.; Chiang, K.-S.; Del Ponte, E.M. Plant disease severity estimated visually: A century of research, best practices, and opportunities for improving methods and practices to maximize accuracy. Trop. Plant Pathol. 2022, 47, 25–42. [Google Scholar] [CrossRef]
- Brouwer, M.; Lievens, B.; Van Hemelrijck, W.; Van den Ackerveken, G.; Cammue, B.P.A.; Thomma, B.P.H.J. Quantification of disease progression of several microbial pathogens on Arabidopsis thaliana using real-time fluorescence PCR. FEMS Microbiol. Lett. 2003, 228, 241–248. [Google Scholar] [CrossRef]
- Goudarzi, R.; Mortazavi, M.M. Loop-mediated isothermal amplification: A rapid molecular technique for early diagnosis of Pseudomonas syringae pv. syringae of stone fruits. J. Genet. Eng. Biotechnol. 2020, 18, 55. [Google Scholar] [CrossRef]
- Guilbaud, C.; Morris, C.E.; Barakat, M.; Ortet, P.; Berge, O. Isolation and identification of Pseudomonas syringae facilitated by a PCR targeting the whole P. syringae group. FEMS Microbiol. Ecol. 2016, 92, fiv146. [Google Scholar] [CrossRef]
- Yeh, K.B.; Wood, H.; Scullion, M.; Russell, J.A.; Parker, K.; Gnade, B.T.; Jones, A.R.; Whittier, C.; Mereish, K. Molecular detection of biological agents in the field: Then and now. mSphere 2019, 4, e00695-19. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, K.N.; Kim, K.-H.; Takemoto, J.Y. PCR Detection of cyclic lipodepsinonapeptide-producing Pseudomonas syringae pv. syringae and similarity of strains. Appl. Environ. Microbiol. 1998, 64, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.; Lovelace, A.H.; Kvitko, B.H. Validation of RT-qPCR approaches to monitor Pseudomonas syringae gene expression during infection and exposure to pattern-triggered immunity. Mol. Plant-Microbe Interact. 2018, 31, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.-X.; Zhang, F.; Culma, C.M.; Lin, S.; Niu, Y.; Zhang, T.; Yang, Q.; Smith, M.; Hu, J. Construction of high-density linkage maps and identification of quantitative trait loci associated with Verticillium wilt resistance in autotetraploid alfalfa (Medicago sativa L.). Plant Dis. 2020, 104, 1439–1444. [Google Scholar] [CrossRef] [PubMed]
- Cullis, B.R.; Smith, A.B.; Coombes, N.E. On the design of early generation variety trials with correlated data. J. Agric. Biol. Environ. Stat. 2006, 11, 381–393. [Google Scholar] [CrossRef]
- Shen, C.; Du, H.; Chen, Z.; Lu, H.; Zhu, F.; Chen, H.; Meng, X.; Liu, Q.; Liu, P.; Zheng, L.; et al. The chromosome-level genome sequence of the autotetraploid alfalfa and resequencing of core germplasms provide genomic resources for alfalfa research. Mol. Plant 2020, 13, 1250–1261. [Google Scholar] [CrossRef]
- Bonhomme, M.; Fariello, M.I.; Navier, H.; Hajri, A.; Badis, Y.; Miteul, H.; Samac, D.A.; Dumas, B.; Baranger, A.; Jacquet, C.; et al. A local score approach improves GWAS resolution and detects minor QTL: Application to Medicago truncatula quantitative disease resistance to multiple Aphanomyces euteiches isolates. Heredity 2019, 123, 517–531. [Google Scholar] [CrossRef]
- Guo, L.; Zhao, L.; Lu, X.; Chen, X.; Wang, S.; Wang, J.; Wang, D.; Yin, Z.; Chen, C.; Fan, Y.; et al. Characters and structures of the nucleobase–ascorbate transporters (NAT) family genes in Gossypium hirsutum and their roles in responding to salt and drought stresses. J. Cott. Res. 2022, 5, 10. [Google Scholar] [CrossRef]
- Han, G.; Qiao, Z.; Li, Y.; Yang, Z.; Wang, C.; Zhang, Y.; Liu, L.; Wang, B. RING zinc finger proteins in plant abiotic stress tolerance. Front. Plant Sci. 2022, 13, 877011. [Google Scholar] [CrossRef]
- Deng, X.; Xu, X.; Liu, Y.; Zhang, Y.; Yang, L.; Zhang, S.; Xu, J. Induction of γ-aminobutyric acid plays a positive role to Arabidopsis resistance against Pseudomonas syringae. J. Int. Plant Biol. 2020, 62, 1797–1812. [Google Scholar] [CrossRef]
- Bart, R.S.; Chern, M.; Vega-Sánchez, M.E.; Canlas, P.; Ronald, P.C. Rice Snl6, a cinnamoyl-CoA reductase-like gene family member, is required for NH1-mediated immunity to Xanthomonas oryzae pv. oryzae. PLoS Genet. 2010, 6, e1001123. [Google Scholar] [CrossRef] [PubMed]
- Tenhaken, R.; Thulke, O. Cloning of an enzyme that synthesizes a key nucleotide-sugar precursor of hemicellulose biosynthesis from soybean: UDP-glucose dehydrogenase. Plant Physiol. 1996, 112, 1127–1134. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.A.; Gidley, M.J.; Fincher, G.B. Heterogeneity in the chemistry, structure and function of plant cell walls. Nat. Chem. Biol. 2010, 6, 724–732. [Google Scholar] [CrossRef] [PubMed]
- De Strooper, B.; Saftig, P.; Craessaerts, K.; Vanderstchele, H.; Guhde, G.; Annaert, W.; Von Figura, K.; Van Leuven, F. Deficiency of presenilin-1 inhibits the normal cleavage of amyloid precursor protein. Nature 1998, 391, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, S.; Qian, Q.; Zeng, D.; Zhang, M.; Guo, L.; Liu, X.; Zhang, B.; Deng, L.; Liu, X.; et al. BC10, a DUF266-containing and golgi-located type ii membrane protein, is required for cell-wall biosynthesis in rice (Oryza sativa L.). Plant J. 2009, 57, 446–462. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.C. Structure and function of the receptor-like protein kinases of higher plants. Plant Mol. Biol. 1994, 26, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.-F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Microbiol. 2018, 16, 316–328. [Google Scholar] [CrossRef]
- Bender, C.L.; Alarcón-Chaidez, F.; Gross, D.C. Pseudomonas syringae phytotoxins: Mode of action, regulation, and biosynthesis by peptide and polyketide synthetases. Microbiol. Mol. Biol. Rev. 1999, 63, 266–292. [Google Scholar] [CrossRef]
- Moore, R.A.; Starratt, A.N.; Ma, S.-W.; Morris, V.L.; Cuppels, D.A. Identification of a chromosomal region required for biosynthesis of the phytotoxin coronatine by Pseudomonas syringae pv. tomato. Can. J. Microbiol. 1989, 35, 910–917. [Google Scholar] [CrossRef]
- Uppalapati, S.R.; Ayoubi, P.; Weng, H.; Palmer, D.A.; Mitchell, R.E.; Jones, W.; Bender, C.L. The phytotoxin coronatine and methyl jasmonate impact multiple phytohormone pathways in tomato. Plant J. Cell Mol. Biol. 2005, 42, 201–217. [Google Scholar] [CrossRef]
- Lipps, S.M.; Samac, D.A.; Ishii, S. Genome sequence resource for strains of Pseudomonas syringae phylogroup 2b and Pseudomonas viridiflava phylogroup 7a causing bacterial stem blight of alfalfa. Phytopathology 2022, 112, 2028–2031. [Google Scholar] [CrossRef] [PubMed]
- Baltenneck, J.; Reverchon, S.; Hommais, F. Quorum sensing regulation in phytopathogenic bacteria. Microorganisms 2021, 9, 239. [Google Scholar] [CrossRef] [PubMed]
- Koh, C.-L.; Sam, C.-K.; Yin, W.-F.; Tan, L.Y.; Krishnan, T.; Chong, Y.M.; Chan, K.G. Plant-derived natural products as sources of anti-quorum sensing compounds. Sensors 2013, 13, 6217–6228. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Teplitski, M.; Robinson, J.B.; Bauer, W.D. Production of substances by Medicago truncatula that affect bacterial quorum sensing. Mol. Plant-Microbe Interact. 2003, 16, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Miles, C.; Wayne, M. Quantitative trait locus (QTL) analysis. Nat. Educ. 2008, 1, 208. [Google Scholar]
- Shelp, B.J.; Bown, A.W.; McLean, M.D. Metabolism and functions of gamma-aminobutyric acid. Trends Plant Sci. 1999, 4, 446–452. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Dou, N.; Zhang, H.; Wu, C. The versatile GABA in plants. Plant Signal. Behav. 2021, 16, 1862565. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, B.M.; Neale, H.C.; Geilfus, C.M.; Jackson, R.W.; Arnold, D.L.; Preston, G.M. Early changes in apoplast composition associated with defence and disease in interactions between Phaseolus vulgaris and the halo blight pathogen Pseudomonas syringae pv. phaseolicola. Plant Cell Environ. 2016, 39, 2172–2184. [Google Scholar] [CrossRef]
- Ward, J.L.; Forcat, S.; Beckmann, M.; Bennett, M.; Miller, S.J.; Baker, J.M.; Hawkins, N.D.; Vermeer, C.P.; Lu, C.; Lin, W.; et al. The metabolic transition during disease following infection of Arabidopsis thaliana by Pseudomonas syringae pv. tomato. Plant J. 2010, 63, 443–457. [Google Scholar] [CrossRef]
- Van der Biezen, E.A.; Jones, J.D.G. The NB-ARC Domain: A novel signalling motif shared by plant resistance gene products and regulators of cell death in animals. Curr. Biol. 1998, 8, R226–R228. [Google Scholar] [CrossRef]
- Van Ooijen, G.; Mayr, G.; Kasiem, M.M.A.; Albrecht, M.; Cornelissen, B.J.C.; Takken, F.L.W. Structure-function analysis of the NB-ARC domain of plant disease resistance proteins. J. Exp. Bot. 2008, 59, 1383–1397. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Yao, L.; Wan, R.; Li, Z.; Liu, C.; Wang, X. Ectopic expression in Arabidopsis thaliana of an NB-ARC encoding putative disease resistance gene from wild Chinese Vitis pseudoreticulata enhances resistance to phytopathogenic fungi and bacteria. Front. Plant Sci. 2015, 6, 1087. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.A.; Shadle, G.L.; Zhou, R.; Nakashima, J.; Chen, F.; Dixon, R.A. Improving saccharification efficiency of alfalfa stems through modification of the terminal stages of monolignol biosynthesis. BioEnergy Res. 2008, 1, 180–192. [Google Scholar] [CrossRef]
- Moya, Y.S. Development of Genetic Mapping and DNA Markers for Tolerance to Bacterial Stem Blight Caused by Pseudomonas syringae pv. syringae in Alfalfa. Master’s Thesis, University of Minnesota, St. Paul, MN, USA, 2023. [Google Scholar]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple Genotyping-by-Sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Duitama, J.; Quintero, J.C.; Cruz, D.F.; Quintero, C.; Hubmann, G.; Foulquié-Moreno, M.R.; Verstrepen, K.J.; Thevelein, J.M.; Tohme, J. An integrated framework for discovery and genotyping of genomic variants from high-throughput sequencing experiments. Nucl. Acids Res. 2014, 42, e44. [Google Scholar] [CrossRef]
- Broad Institute Picard Toolkit. Available online: http://broadinstitute.github.io/picard/ (accessed on 1 February 2022).
- Rosyara, U.R.; De Jong, W.S.; Douches, D.S.; Endelman, J.B. Software for genome-wide association studies in autopolyploids and its application to potato. Plant Genome 2016, 9, 1–10. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. Roy. Stat. Soc. B Met. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Medina, C.A.; Samac, D.A.; Yu, L.-X. Pan-transcriptome identifying master genes and regulation network in response to drought and salt stresses in alfalfa (Medicago sativa L.). Sci. Rep. 2021, 11, 17203. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | SNP | −log p-Value | R2 | Effect | Uniprot ID | Gene | Annotation | Role | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Chr1_6983943 | C/A | 3.47 | 0.056 | −0.257 | G7J119 | − | NB-ARC domain disease resistance protein | Response to biotic stress | [39] |
| Chr1_95741849 | G/A | 3.4 | 0.051 | 0.251 | UPI000B7B775A | NAT4 | Nucleobase-ascorbate transporter 4 | Response to abiotic stress | [40] |
| Chr2_16508382 | T/C | 3.73 | 0.028 | 0.219 | W4ZQ03 | − | Bifunctional inhibitor/plant lipid transfer protein/seed storage helical domain-containing protein | − | |
| Chr3_75660736 | C/G | 4.15 | 0.033 | 0.404 | UPI0011DFD893 | − | RING-type E3 ubiquitin transferase | Plant growth and response to abiotic stress | [41] |
| Chr4_77596753 | T/A | 3.36 | 0.037 | −0.266 | UPI001016EB81 | GAD | Glutamate decarboxylase | Response to biotic stress | [42] |
| Chr5_19814514 | G/A | 3.45 | 0.031 | 0.214 | A0A445LXJ0 | Snl6 | Cinnamoyl-CoA reductase-like SNL6 | Response to biotic stress | [43] |
| Chr5_34806446 | A/G | 3.31 | 0.008 | 0.220 | A0A061EAW6 | UGD | UDP-glucose 6-dehydrogenase | Cell wall biosynthesis | [44] |
| Chr7_6703934 | T/C | 3.38 | 0.076 | 0.218 | A0A059BED4 | PE | Pectinesterase | Cell wall stability | [45] |
| Chr7_75396634 | A/T | 3.87 | 0.029 | 0.343 | A0A445CS76 | PSEN | Presenilin | Protein cleavage | [46] |
| Chr8_78651242 | C/T | 3.41 | 0.032 | −0.195 | A0A371G107 | GT-BC10 | Glycosyltransferase BC10 | Biosynthesis of polysaccharides and glycoproteins | [47] |
| Chr8_80754378 | T/G | 3.32 | 0.032 | −0.314 | A0A445DK30 | − | Protein kinase domain-containing protein | Plant signaling and response to abiotic stress | [48] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moya, Y.S.; Medina, C.; Herrera, B.; Chamba, F.; Yu, L.-X.; Xu, Z.; Samac, D.A. Genetic Mapping of Tolerance to Bacterial Stem Blight Caused by Pseudomonas syringae pv. syringae in Alfalfa (Medicago sativa L.). Plants 2024, 13, 110. https://doi.org/10.3390/plants13010110
Moya YS, Medina C, Herrera B, Chamba F, Yu L-X, Xu Z, Samac DA. Genetic Mapping of Tolerance to Bacterial Stem Blight Caused by Pseudomonas syringae pv. syringae in Alfalfa (Medicago sativa L.). Plants. 2024; 13(1):110. https://doi.org/10.3390/plants13010110
Chicago/Turabian StyleMoya, Yeidymar Sierra, Cesar Medina, Bianca Herrera, Fabian Chamba, Long-Xi Yu, Zhanyou Xu, and Deborah A. Samac. 2024. "Genetic Mapping of Tolerance to Bacterial Stem Blight Caused by Pseudomonas syringae pv. syringae in Alfalfa (Medicago sativa L.)" Plants 13, no. 1: 110. https://doi.org/10.3390/plants13010110
APA StyleMoya, Y. S., Medina, C., Herrera, B., Chamba, F., Yu, L.-X., Xu, Z., & Samac, D. A. (2024). Genetic Mapping of Tolerance to Bacterial Stem Blight Caused by Pseudomonas syringae pv. syringae in Alfalfa (Medicago sativa L.). Plants, 13(1), 110. https://doi.org/10.3390/plants13010110