Abstract
The flexibility of LED technology, in terms of energy efficiency, robustness, compactness, long lifetime, and low heat emission, as well as its applications as a sole source or supplemental lighting system, offers interesting potential, giving the ornamental industry an edge over traditional production practices. Light is a fundamental environmental factor that provides energy for plants through photosynthesis, but it also acts as a signal and coordinates multifaceted plant-growth and development processes. With manipulations of light quality affecting specific plant traits such as flowering, plant architecture, and pigmentation, the focus has been placed on the ability to precisely manage the light growing environment, proving to be an effective tool to produce tailored plants according to market request. Applying lighting technology grants growers several productive advantages, such as planned production (early flowering, continuous production, and predictable yield), improved plant habitus (rooting and height), regulated leaf and flower color, and overall improved quality attributes of commodities. Potential LED benefits to the floriculture industry are not limited to the aesthetic and economic value of the product obtained; LED technology also represents a solid, sustainable option for reducing agrochemical (plant-growth regulators and pesticides) and energy inputs (power energy).
1. Introduction
The ornamental industry produces plants for their aesthetic value from a very wide range of species, including floriculture crops, ornamental grasses, turf grasses, and ornamental trees and shrubs. The economic impact of floriculture has remarkable importance. The worldwide flower and ornamental plant market amounted to as much as USD 52,384.85 million in 2022 and is expected to grow with a CAGR of 7.28% from 2023 to 2028 in the forecast period [1]. Europe is the largest ornamental market, with about 47% market share, followed by Asia–Pacific (20%) and the United States (13.5%) [1]. Despite the economic magnitude of the ornamental market, as with many nonessential commodities and industries, the COVID-19 pandemic and subsequent public shutdown events substantially damaged its production chain across the globe [2]. In addition, the ornamental sector deals with multiple challenges, including market globalization, uncertainty related to climate change, land-use competition, and anthropogenic pressures [3,4]. In this scenario, it is necessary to find more sustainable solutions for agricultural production sectors that allow for increases in the quality and yield of products while reducing production costs, environmental pressures, and natural resource depletion. For the optimization of ornamental production, in terms of both economics and sustainability, one opportunity/possibility is the application of artificial light in controlled environment systems (greenhouses, soilless systems, and indoor farming), which, overall, allows an accurate handling of environmental parameters by using a technology-driven approach [5].
Light is the essential environmental factor coordinating plant growth, development, and function since it represents the driving force for photosynthetic CO2 assimilation. It is also the signal that triggers multiple response pathways that are involved in many developmental aspects of growth, collectively recognized as photomorphogenesis [6]. In the last few decades, the use of artificial lighting for plant cultivation has become an interesting choice, either as a supplementary source when solar radiation is scarce, or as a sole light source, providing energy for photosynthesis, modulating crop morphogenesis, and regulating the flowering process [7,8,9]. By adjusting/modulating the light components/properties, such as quantity (intensity), duration (photoperiod), and quality (spectral composition), it is possible to attain important ornamental production targets to induce flowering, control leaf shape and plant architecture, extend the production season, fine-tune leaf and flower color, improve longevity, and enhance resilience to pathogens [6].
Until recently, the viable and widely used options for artificial lighting systems were high-intensity discharge (high-pressure sodium, HPS; metal halide, MZ) and fluorescent lamps due to their relatively high fluence and economical affordability [10]. However, these conventional lighting systems show some disadvantages, including the generation of excessive heat, high energy needs, and inability to modulate the light spectrum, generally emitting light over a limited broad spectrum (orange-red region, 550–650 nm, with less in the blue region, 400–500 nm) [11,12]. The recently emerged light-emitting diode (LED) technology has great potential for protected ornamental production [13]. LEDs offer several unique advantages over traditional lighting systems since they are the most energy-efficient and environmentally friendly lighting technologies currently available [14]. LEDs provide higher energy efficiency, which allows for reductions in electricity costs; together with their performance characteristics/features such as robustness, compactness, durability, and long lifetime, LEDs represent a cost-effective option that is largely appreciated in commercial settings [13,15]. Low heat emission allows the light source to be placed close to the canopy, ensuring a uniform spectral distribution while preventing tissue damage from photostress [16]. Additionally, owing to the advantages of high-light-intensity selection and spectral modulation, LEDs, remarkably, meet the specific requirements of leaf optical properties, encompassing dynamic photosynthetic activity and biochemistry processes to control the growth and development of plants [17].
This review provides an overview of the use of LED lighting technology for growing/producing ornamental crops (Figure 1).

Figure 1.
Spectral wavelengths affecting decorative traits of ornamental plants. (A) Blue, far-red, and red lights are typically effective in promoting flowering in long-day (LD) plants, whereas growth extension in short-day (SD) plants is promoted by using supplemental illumination at the end of the day with far-red light. (B) Blue radiation in a red background limits extension growth and promotes branching. (C) Stem elongation is regulated by controlling the shade avoidance phenomenon, using a lower R/Fr ratio or higher red percentage in a blue-light environment. (D) The vase life of cut flowers is improved when stored in a cold room and exposed to a sole source of blue light.
The novelty of this review is to elucidate how LED illumination can be exploited to promote/prompt innovation in the ornamental market, while providing guidelines/recommendations for growers to improve the quality and yield of their production systems/practices. Therefore, the modulation of ornamental plant attributes by LEDs is discussed, including flowering regulation, plant architecture, postharvest/postproduction longevity, flower and leaf color, and pathogen and disease control (Table 1).

Table 1.
Main effects of LEDs on different ornamental plants.
2. Flowering Regulation
Flower induction and initiation are complex processes driven by environmental and intrinsic factors that influence the transition from the vegetative phase to reproductive competence [43,44]. The integration of endogenous signals in response to external cues is strictly mediated by a complex network of genetic pathways to ensure the progeny’s success [45,46,47,48,49].
Various species of plants, including many ornamental crops, synchronize their growth and development by sensing changes in the light environment, such as photoperiod, light intensity, spectral composition, and direction [45]. In terms of photoperiodic requirements, most ornamental plants can be classified as long-day (LD) plants, short-day (SD) plants, and day-neutral plants (ND). Flowering of LD plants is induced when the night length is less than a certain threshold (critical duration). Flowering of SD plants is promoted during long nights (short days), whereas, in ND plants, flowering can occur irrespective of the day’s length [50]. During flowering, coordinated endogenous responses to the relative lengths of the light and the dark periods take place in leaves through a complex gene regulatory network involved in light sensing, which is driven by photoreceptor action [51,52]. Molecular evidence has demonstrated that the flowering transition occurs via upregulation of FLOWERING LOCUS T (FT), also known as florigen, and repression of antiflorigenic FT (AFT)/TERMINAL FLOWER 1 (TFL1) [53]. The molecular mechanism of the inductive photoperiod is conserved in both LD and SD plants [54]. In addition to the photoperiod, the spectral composition, hormone pathways, and temperature play significant roles in the control of flowering for both LD and SD crops. In some ornamental crops, low-temperature exposure is required to regulate the transition from vegetative to reproductive growth (vernalization) [55]. In many species, the relationship between the photoperiod and the temperature has been shown to regulate the flowering transition, thus representing the main integrated approach for harvesting schedules and utilizing greenhouse space. This allows the planning of a controlled growing environment, as well as producing continuously predictable yields over predetermined time periods. Typically, a low light intensity is provided at night to boost the flowering of LD plants and reduce their crop production cycle, while preventing flowering in SD plants and promoting their vegetative growth [54]. Chrysanthemums are the second most important ornamental crop and are grown as cut or potted flowers. To ensure year-round availability for the market demand for short-day plants such as chrysanthemums, as well as to assure the programmed flowering on predetermined market dates, artificial lighting is provided as a day-length extension to promote vegetative growth or as a night break to prevent premature flowering [56,57].
In addition to the photoperiod, the spectral composition influences the flowering process in short-day and long-day plants [58]. The specific light quality drives the flower transition, which in turn leads to transcriptional regulation of the genes that encode activators of flowering, i.e., the photoreceptors. Several photoreceptors are involved in the perception and absorption of different wavelengths: phytochromes that preferentially absorb in the red (660 nm)/far-red (730 nm) spectral regions; cryptochromes that preferentially absorb in the blue/UV-A wavelengths; and phototropins (PHOT), ZTL/FKF1/LKP2, and UVR8 that mostly absorb UV-B light [59]. Blue light and far-red light are typically effective in promoting flowering in LD plants (Figure 1 and Figure 2). The efficacy of LEDs compared to conventional lamps (HPS) was evaluated by comparing their regulatory role in the flowering of photoperiodic plants. In this respect, LEDs provide comparable effectiveness to conventional light sources, while featuring a lower total operating cost [60]. In petunias (Petunia hybrida E. Vilm.) and snapdragons (Antirrhinum majus L.), long hours of illumination using a high daily light integral (DLI) and a red/white/far-red lamp significantly encouraged flower formation and development [61]. Similarly, in day-neutral Cyclamen persicum Mill., the combined use of high light intensity with blue and red wavelengths was useful to promote flowering and subsequent development [25]. On the other hand, when LD crops (i.e., snapdragon, tussock bellflower, tickseed, and petunia) were grown under a far-red light-deficient environment, a delay in flower initiation and development was observed [62,63,64]. SD plants cultivated in greenhouses are negatively affected by the lower DLI and shorter photoperiod occurring over the winter season. However, supplemental illumination that prolongs the day length by using far-red light has been reported as a cost-effective strategy that favors growth extension, e.g., as a tool to improve the plant habitus.
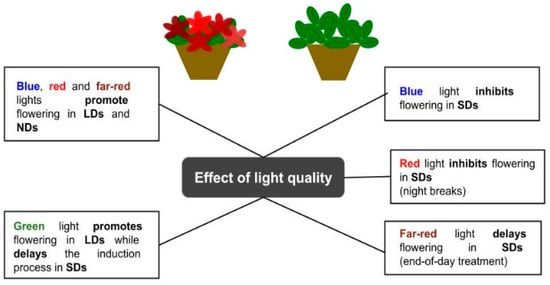
Figure 2.
Schematic representation of the effect of light quality on the flowering process of ornamental plants.
When using end-of-day far-red treatments in poinsettias [65], chrysanthemums, and garden strawberries [66], flowering initiation occurred later in development, while the plants showed longer stems and longer internodes. The delayed flowering status may be attributed to the altered phytochrome level in a far-red light environment at the end of the day [66]. A similar inhibitory effect was reported in chrysanthemums, where short days of solar light, followed by a 4 h extension with blue or red light, were not enough to affect the flowering initiation [67]. When the natural day is short, the use of red light to interrupt the night is a typical practice to inhibit flowering in SD plants due to the photochemical interconversion of phytochrome Pr to the Pfr form during the night [68]. Moreover, by applying far-red light, the flowering inhibition mediated by the phytochrome photoequilibrium was reversed [69]. Furthermore, blue light, as well as its light signaling initiated by the cryptochrome, has a significant impact on flowering and can be used to control the process. At a higher intensity (20 µmol∙m−2∙s−1 or higher), blue light has a flower-promoting effect on LD plants, while showing an inhibitory action on SD plants when applied as a night-break or day-length extension regime [70,71]. The promotion effect of blue light on LD flowering was observed as earlier flowering, a greater flowering index, and more visible flower buds and opened flowers, seemingly associated with lower phytochrome activity, also known as a phytochrome photostationary state [35].
The effect of green light on photoperiodic flowering has been reported in a few studies. Similar to blue-light flowering responses, short-day plants grown in a green light environment showed a delay/inhibition of flowering, depending on the species, as well as the duration and intensity of exposure [21,70,72] (Figure 3). Moreover, Meng and Runkle [32] showed that fluxes of green radiation may function as a long-day signal. Adopting a spectrum with moderate intensities of green light for several hours was effective in saturating the flowering responses of petunias, snapdragons, and ageratum floriculture crops. On the other hand, in chrysanthemum and marigold SD plants, the delivery of green light exerted a delayed flowering effect, suggesting a role for this wavelength in the control of flower induction for photoperiodic plants [32].
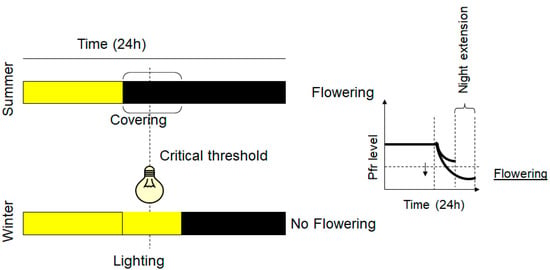
Figure 3.
SD plants and flowering control. Yellow color represents daylight or lighting and black color the night or dark induced by covering the plants. LED light with emission at 730 nm converts the Pfr into Pr and inhibits the flowering of SD plants, overcoming the critical time threshold. On a biochemical level, right graph, nighttime reduces the Pfr level, and the induction of SD plant flowering occurs when the Pfr level declines below a critical concentration threshold. Night extension by covering the plants induces flowering and it is a strategy used in summer for inducing flowering of SD plants.
Application of a UV spectrum can either promote or delay flowering; the responses depend on the species, region of the UV spectrum, and fluence rate. With regard to this last aspect, high UV radiation has been shown to dramatically impact flowering quality (Figure 4). For example, the flowering time and the number of flowers produced in Phacelia campanularia A. Gray and Salvia splendens Sellow ex Nees plants exposed to high UV dosages were significantly hampered [73,74], whereas the flower transitions of Limnanthes alba Hartw. ex Benth. plants were inhibited [73]. In contrast, UV-C radiation improved flowering and even increased the flower number in wild pansy and freesia ornamental plants [74,75].
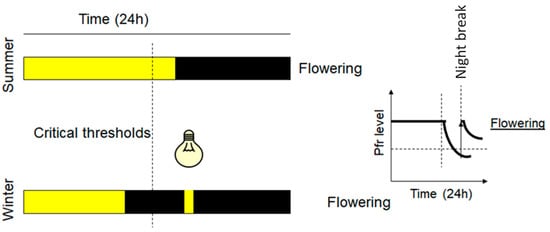
Figure 4.
LD plants and flowering control. Yellow color represents daylight or lighting and black color the night or dark induced by covering the plants LED light with emission at 730 nm converts the Pfr into Pr and induces the flowering of LD plants, overcoming the critical time threshold. On a biochemical level, right graph, short (flash) lighting induces night break and increases the Pfr level, and the induction of LD plant flowering occurs when the Pfr level exceeds a critical concentration threshold.
3. Plant Architecture
LED technology, through the ability to select specific wavelengths, offers the possibility to develop tailored light recipes for the manipulation of plant architecture. Plant quality (distribution of energy across different wavelengths) is often a mix of specific plant traits, such as branching, compactness, rooting, and leaf expansion, which are strongly influenced by the spectral composition of LED light [11]. The reasonable choice for commercial plant production using LED systems is the combination of red and blue wavelengths, since the absorption spectra of photosynthetic pigments mainly focus on blue (400–500 nm) and red (600–700 nm) light [76], and several regulatory mechanisms can be exploited.
Young ornamental plant production is an integral part of the floriculture industry. The bulk of production occurs in winter or early spring to meet the spring and summer sale demand. Unfortunately, this is also when the outdoor photosynthetic daily light integral (DLI) is seasonally low, and is even lower in greenhouses. One of the most cost-effective applications of LED lighting is bedding plant production, which allows the obtainment of more uniform, compact, and high-quality annual young ornamental plants with marketable characteristics, as well as the ability to withstand transplanting shock. Controlling the growth of these commodities is a vital aspect of the ornamental industry since this allows the improvement of both their visual quality and their physiological status. Several studies have used red and blue light to assess their effects on the morphology and anatomy of plants. In general, red and blue LED lights affect physiological and morphological traits, such as stomatal openings, plant height, chlorophyll biosynthesis, stem elongation, branching, leaf expansion, and reproduction [11]. Both supplemental and sole sources of LED lighting, with blue radiation in a red background, limit the extension growth and leaf expansion compared to growth under ambient light supplemented with an HPS lamp or cool white fluorescence, providing an effective nonchemical method to control the height of several species of bedding plants [12,30,77]. Furthermore, the productivity and quality of cuttings can be modulated/influenced by LED treatment. Adventitious rooting is a critical process in the vegetative propagation of ornamental plants, and LED lighting positively affects the growth, survival, and rooting of cuttings. Compared to the application of red or blue light alone, the combined use of red and blue light (R:B ratio of 1:1) in a multilayer sole-source light propagation system reduced stem elongation and improved root biomass in herbaceous perennial cuttings, while avoiding damage during shipping and transplanting [40]. Cuttings are susceptible to fast drying, and the control of transpiration can be achieved using a well-balanced light spectrum, since the stomatal opening response is predominantly initiated by blue light but enhanced under a strong red-light background [78,79]. In Impatiens ×hybrida hort., for instance, environments with a high percentage of red light but a lower percentage of blue light have been shown to increase the number of trichomes, anatomical structures linked to the prevention of water loss by transpiration [29]. Additionally, this light recipe led to a greater plug compactness and survival of cuttings, achieving a tradeoff between the risk of dehydration and quality of cuttings. In other species, such as Chrysanthemum ×morifolium (Ramat.) Hemsl., Lavandula angustifolia Mill., and Rhododendron simsii Planch. hybrids, treatment with red light only (100) was highly efficient in enhancing rooting performance [20].
The cultivation of cut flowers in a protected environment enables year-round production in northern latitudes, which are characterized by unfavorable conditions. In fact, using LED lighting solutions as a supplemental source in greenhouses overcomes the concrete risk of not reaching the minimum lighting requirements for crops [80]. The shoot architecture, particularly stem elongation, can be regulated by controlling the shade avoidance phenomenon related to the excessive growth of plants when subjected to the shade of other plants or when growing in high-density conditions, whereby the availability of photosynthetically active radiation (PAR) is reduced, along with the ratio of red-to-far-red (R/FR) light [81]. Along with physiological changes, a low phytochrome stationary state enhances internode and petiole elongation, axillary bud outgrowth, and hyponasty [81]. Grading standards for cut chrysanthemum flowers on the world market require an elongated and unbranched plant shape and large-sized flowers. Treatment of rooted chrysanthemum cuttings with a combination of blue and far-red light showed a higher internode length compared to sole red light; in decapitated cuttings, the apical bud concomitantly reached a high length with inhibited growth of underlying buds [82]. In lilium, grown as a cut flower, different ratios of red to blue light have been shown to influence different characteristics; when exposed to the highest red percentage (R:B ratio of 80:20), the height of the stems was greatly enhanced [33]. Furthermore, upon increasing the percentage of blue light, several morphological traits were modulated, such as reduced time to harvest (R:B ratio of 20:80), strong inhibition of stem elongation (R:B ratio of 40:60), and slightly improved vase life (R:B ratio of 60:40) [33]. Similarly, potted miniature rose ‘Aga’ plants, grown under a supplemental wide spectrum of red, blue, white, and far-red LEDs, exhibited significantly greater height and shoot length than control plants [37]. Exposure to blue and blue/red light positively affected the photosynthetic performance of Cordyline australis (G. Forst.) Endl., Ficus benjamina L., and Sinningia speciosa Hiern potted foliage plants, while also showing greater stomatal conductance and density, as well as an increase in leaf thickness [23]. Generally, the commercial standard for potted plants requires a compact shape; thus, the light spectrum tends to be manipulated while avoiding light sources with a low R:FR ratio since its effects on overall growth reduces the decorative value of ornamental potted plants. In petunias, despite the promoting effect of a lower ratio of red-to-far-red light on flowering, the overall quality was adversely affected, showing excessive stem elongation, weak stems, and poor branching [83]. However, by adding a moderate green wavelength to a red, far-red, and white background, petunia plants were shorter and developed more branches [60]. Green light has been shown to participate in regulating growth and development through its ability to much more effectively penetrate the lower canopy, thereby optimizing the photosynthetic machinery [84]. The use of pure green light in a recent study promoted elongation via brassinosteroids, triggering the activation of the BRI1-EMS-SUPPRESSOR 1 (BES1) transcription factor and the target genes in its downstream signaling pathway [19]. This effect of green light on stem elongation has been reported, for example, in Zingiber officinale Roscoe, where even the use of supplemental, green-enriched light enabled the plants to strongly improve their photosynthetic performance [85].
4. Postharvest/Postproduction Longevity
The quality of ornamental plants depends not only on their external attributes, such as shape, size, color, and flower and leaf turnover, but also on the ability to preserve their characteristics [3]. In fact, as a fresh commodity, they are still metabolically active and extremely perishable after harvest/production and are highly vulnerable to large postharvest losses. Thus, the longevity of ornamental plants is the main goal for their commercial success, but suboptimal postharvest conditions that often occur during storage and transportation negatively impact the overall quality and accelerate degenerative processes. Regardless of the product type (i.e., cut flowers, potted foliage, and flowering plants), the main postharvest disorders that compromise the decorative value are leaf yellowing, flower and bud senescence, and abscission [3,86]. To avoid a negative impact on the marketability of these commodities and the resulting reduction in profit for both producers and sellers, several postharvest handling approaches have been developed over the years. Technological interventions have mainly focused on the structural optimization of the postharvest chain (transport and storage), as well as the development of novel packaging and precondition techniques to delay the senescence process [86]. Several commercial chemicals are available for cut flowers and potted plants, including ethylene inhibitors, antibacterial agents, synthetic growth regulators, and sugars. Ethylene is widely known as an aging hormone, as it promotes and accelerates senescence-related processes. To prevent leaf and flower abscission, the application of gaseous 1-methylcyclopropene (1-MCP) was effective in blocking ethylene perception and prolonging the longevity (maintaining the freshness) of many ethylene-sensitive potted plants and cut flowers [87]. As biosynthesis inhibitors, amino vinylglycine (AVG) and amino-oxyacetic acid (AOA) avoid endogenous ethylene production by interfering with the key ethylene biosynthetic enzyme 1-aminocyclopropane-1-carboxylic acid (ACC) synthase (ACS, E.C. 4.4.1.14), without impeding the deterioration process trigged by exogenous ethylene [87]. Furthermore, the synthetic compound thidiazuron, with a strong cytokinin-like activity, was successfully used in cut foliage and flowers, as well as in potted plants, to improve postharvest performance (i.e., delaying leaf yellowing and increasing flower longevity) [3,87]. In addition to the crucial role of chemical preservatives, control of the storage and transportation environment assumes great relevance within the ornamental industry [88]. In fact, higher temperatures during storage and transportation are linked to enhanced respiration rates and ethylene production, physiological processes that highly accelerate plant senescence, thereby negatively influencing the overall quality of ornamental products. For this reason, the logistic chain process from the producers to the storage and transportation facilities, and then to the end-user/consumer, is committed to maintaining a cold temperature. However, among the various handling processes of postharvest chains, ornamental products are exposed to fluctuating temperatures, and keeping an optimally low temperature requires high equipment and operating costs. The development of novel systems focused on the postharvest storage environment by manipulating the light spectrum might represent an alternative strategy to support the quality of ornamental products [88]. Recently, the processes underlying the vase life response under different light environments were explored in several ornamental plants. In anthuriums, a tropical cold-sensitive species, prolonged exposure to a low storage temperature compromised the longevity of its cut stems [18]. Applying a sole source of blue light under cold storage led to the highest rate of water loss and electrolyte leakage from the spathes, whereas red light lowered the ROS accumulation in the cells [18]. These metabolic adjustments also led to a significant reduction in vase life in cold storage under blue light, suggesting that oxidative stress and membrane integrity negatively affected the cold tolerance and quality of anthurium spathe. In contrast, cut carnation flowers exposed to blue light showed a prolonged vase life and a markedly delayed senescence [26]. Simultaneously, increases in photosynthetic performance, transpiration rate, sugar content, and water uptake were recorded. These results were in agreement with previous studies on the positive role of blue light in stomatal movements, suggesting a better water transport efficiency and a preserved photosynthetic ability of leaves [89,90,91]. Moreover, the higher antioxidant capacity reported could guarantee a higher membrane integrity and ROS detoxification, consequently preserving/maintaining photosynthetic ability [26] and confirming that the evaluation of antioxidant status in ornamental products represents a key qualitative index to monitor postharvest [26]. The effect of blue light was also investigated in relation to the expression pattern of genes involved in ABA homeostasis, ethylene biosynthesis, and signaling in carnation cut flowers [26]. Both hormones are well known to play regulatory roles during flower senescence in ethylene-sensitive and nonsensitive species [92,93,94,95]. Blue light exposure during storage reduced the expression of the ACS1 and ACO1 genes involved in ethylene biosynthesis, whereas red light led to an increase in their levels [92]. Their relative transcript abundance correlated with their longevity; blue light-exposed cut flowers showed a superior/improved vase life to red light-exposed flowers. Moreover, the activation of transcriptional pathways related to ABA biosynthesis and its transport by blue light appeared to significantly enhance the vase life of cut carnation flowers. Red light has been shown to influence water balance and flower opening in cut roses [88]. In particular, the petal fresh weight was significantly higher under red light compared to the other treatments (blue and white light), and the cut flowers showed a longer vase life. Thus, exposure to a specific light wavelength, particularly red light, might be an effective tool to control flower opening and longevity. LED lighting was found to efficiently maintain plants in an indoor environment distinguished by a low light intensity. Supplemental LED lighting was positioned upward around the base, providing the whole range of radiation (300–800 nm), delaying the senescence of lower leaves, and encouraging flower opening in potted rose plants compared to the downward setting, suggesting a suitable arrangement for indoor plant management [39].
5. Flower and Leaf Color
Another important quality index for the ornamental plant industry, in addition to plant architecture and longevity, is the color of the leaves and flowers, which guides consumers toward their preferences and, therefore, their purchases. The major classes of plant pigments that determine foliage and flower color are chlorophylls, carotenoids, anthocyanins flavonoids, and betalains [96]. Environmental conditions (i.e., temperature, light intensity, and light spectrum) and genetic determinants that mainly drive the development and regulation of pigmentation patterns. Regarding light intensity, plants fall into three categories: high, medium, and low light requirements. The flowers of tuberose and boronia plants, when grown under full sunlight conditions, develop an intense reddish-purple pigmentation compared to those obtained under a shaded environment [97,98]. Similarly, faded flowers were observed when peony flowers were developed in a partially shaded environment due to the downregulation of phenylalanine ammonialyase (PAL E.C. 4.3.1.24) and chalcone synthase (CHS E.C. 2.3.1.74) genes, which are key regulatory steps in the anthocyanin biosynthesis pathway [99]. Furthermore, light has been reported to modulate the accumulation of pigments in various plant organs. For example, in Hibiscus syriacus L. flowers, red light exposure influenced the development of a strong red color in their petals [100]. During winter, the aesthetic quality of some potted foliage plants is negatively affected by the low light intensity since the full biosynthesis of pigments is impeded. The use of LED supplemental lighting enriched with red and blue wavelengths has been reported to increase the accumulation of anthocyanins and carotenoids, leading to a vivid foliage color and an overall improvement in plant’s decorative value [28]. In geraniums and purple fountain grass plants, the supplementation of red and blue LED light at the end of production significantly enhanced red color saturation, expressed as the chroma index, thus increasing aesthetic appeal, quality, and market value [40].
6. Pathogens and Disease Control
Light not only regulates the primary/basic metabolic functions and development (flowering, stem elongation, and morphology) of plants, but can also play a key role in the regulatory network involved in secondary metabolite biosynthesis and accumulation through modulation of the photosensory signaling pathway, orchestrated by photoreceptors [101]. The functional and biological roles of secondary metabolites vary, including defense against phytophages, intra- and/or interspecies communication, protection against harmful solar radiation, and signals for pollination or seed dispersion [102]. Manipulating the light spectrum constitutes an interesting elicitation strategy that actively and suitably interferes with biosynthetic routes, enhancing the concentration of key phytochemicals that can be exploited to improve the growth performance and the final quality of ornamental products (e.g., increasing plant fitness) [11,24,41]. Moreover, light quality, in addition to causing differential metabolic rearrangements, can directly or indirectly impact pathogens and pests, as well as their natural antagonists. Limited exposure to a UV-B light fluorescent lamp is often used to reduce disease incidence in crops that are cultivated in controlled environments (such as growth chambers and greenhouses). In high-density greenhouse-grown roses, low doses of UV-B radiation applied for 6 h completely reduced powdery mildew infection by increasing secondary bioactive compounds [103]. The use of red LED light was instead reported to reduce the number of conidia, suggesting its interesting potential for controlling powdery mildew disease in roses [38]. Gray mold caused by the fungus Botrytis cinerea is one of the most common and destructive plant pathogens, affecting several horticultural crops. Blue light and UV radiation have been shown to suppress the pathogenic development of B. cinerea on harvested vegetables, highlighting their potential application for a wide range of gray-mold-sensitive ornamental crops [88,104]. To limit the damage to flowering plants caused by the behavior of nocturnal moths, LED lights were used as a cheaper pull-and-push strategy against the insects compared to incandescent lamps [105]. Recently, the biological control of thrips and pests has gained popularity as a novel greenhouse system. In Europe, the arthropod Orius laevigatus is available on the market for thrips control in chrysanthemum crop production. O. laevigatus has a relatively quick population buildup, and thus is suitable for the short cropping cycle of chrysanthemums, as long as unfavorable environmental conditions are monitored [106]. In chrysanthemum production, after the introduction of a biocontrol agent for efficient population density establishment, different spectral wavelengths were evaluated in terms of egg-laying activity. The use of red, blue, and green light in equal proportions positively affected the number of eggs laid by O. laevigatus, whereas red light led to the lowest number of eggs [106].
7. Conclusions
The careful selection of components of the light spectrum by using LED lighting technology can significantly improve the quality-related properties/characteristics of ornamental products by influencing several physiological and metabolic processes, such as flowering, branching, rooting, pigment biosynthesis, and vase life. The effects of this technology can vary depending on ornamental species, exposure time, and applied wavelengths; thus, the identification of specific/optimal light formulas is fundamental to achieve the best results. The manipulation of flowering can help reduce costs and production time while obtaining a predictable yield, shaping/modeling the plant habitus and emphasizing attractive features. Moreover, artificial lighting offers a potential alternative to growth retardants used in chemical pinching, as well as an interesting tool for the control of some plant pathogens in greenhouses or growth chambers. The use of LED light in controlled environments can lead to the production of ornamental products with superior characteristics, representing a new frontier of applied sciences with studies focused on species/cultivar-specific light requirements. Furthermore, its application can help in reducing the use of agricultural inputs such as energy and soil in a sustainable manner.
Author Contributions
Conceptualization, A.F. and D.R.; methodology, A.T., S.T., D.R. and A.F.; formal analysis, A.T., S.T., D.R. and A.F.; writing—original draft preparation, A.T., S.T., D.R. and A.F.; writing—review and editing, A.T., S.T., D.R. and A.F.; supervision, D.R. and A.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Acknowledgments
The research was carried out within the PSR Sicilia 2014–2022, Misura 16—Cooperazione—Sottomisura 16.2. “Sostegno a progetti pilota e allo sviluppo di nuovi prodotti, pratiche, processi e tecnologie”. Progetto pilota per lo sviluppo sostenibile di nuovi processi produttivi, nel comparto florovivaistico mediterraneo, ad elevata specializzazione tecnologica, attraverso l’impiego di lampade innovative, ad alto rendimento energetico (LightFlower).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Global Flower and Ornamental Plants Industry Research Report 2023: Competitive Landscape, Market Size, Regional Status and Prospect; Market Reports World, Absolute Reports; Market Reports World: Pune, India, 2023; p. 107.
- Bulgari, R.; Petrini, A.; Cocetta, G.; Nicoletto, C.; Ertani, A.; Sambo, P.; Ferrante, A.; Nicola, S. The impact of COVID-19 on horticulture: Critical issues and opportunities derived from an unexpected occurrence. Horticulturae 2021, 7, 124. [Google Scholar] [CrossRef]
- Ferrante, A.; Trivellini, A.; Scuderi, D.; Romano, D.; Vernieri, P. Post-production physiology and handling of ornamental potted plants. Postharvest Biol. Technol. 2015, 100, 99–108. [Google Scholar] [CrossRef]
- Hill, J.; Von Maltitz, G.; Sommer, S.; Reynolds, J.; Hutchinson, C.; Cherlet, M. World Atlas of Desertification; Publications Office of the European Union: Luxembourg, 2018. [Google Scholar]
- Van Delden, S.H.; SharathKumar, M.; Butturini, M.; Graamans, L.J.A.; Heuvelink, E.; Kacira, M.; Kaiser, E.; Klamer, R.S.; Klerkx, L.; Kootstra, G.; et al. Current status and future challenges in implementing and upscaling vertical farming systems. Nat. Food 2021, 2, 944–956. [Google Scholar] [CrossRef]
- Xu, Y. Nature and source of light for plant factory. In Plant Factory Using Artificial Light; Anpo, M., Fukuda, H., Wada, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 47–69. [Google Scholar]
- Marondedze, C.; Liu, X.; Huang, S.; Wong, C.; Zhou, X.; Pan, X.; An, H.; Xu, N.; Tian, X.; Wong, A. Towards a tailored indoor horticulture: A functional genomics guided phenotypic approach. Hortic. Res. 2018, 5, 68. [Google Scholar] [CrossRef]
- Paucek, I.; Appolloni, E.; Pennisi, G.; Quaini, S.; Gianquinto, G.; Orsini, F. LED Lighting systems for horticulture: Business growth and global distribution. Sustainability 2020, 12, 7516. [Google Scholar] [CrossRef]
- Massa, G.D.; Kim, H.H.; Wheeler, R.M.; Mitchell, C.A.; Cary, A. Plant productivity in response to LED lighting. HortScience 2008, 43, 1951–1956. [Google Scholar] [CrossRef]
- Zheng, L.; He, H.; Song, W. Application of light-emitting diodes and the effect of light quality on horticultural crops: A review. HortScience 2019, 54, 1656–1661. [Google Scholar] [CrossRef]
- Paradiso, R.; Proietti, S. Light-quality manipulation to control plant growth and photomorphogenesis in greenhouse horticulture: The state of the art and the opportunities of modern LED systems. J. Plant Growth Regul. 2021, 41, 742–780. [Google Scholar] [CrossRef]
- Islam, M.A.; Kuwar, G.; Clarke, J.L.; Blystad, D.-R.; Gislerød, H.R.; Olsen, J.E.; Torre, S. Artificial light from light emitting diodes (LEDs) with a high portion of blue light results in shorter poinsettias compared to high pressure sodium (HPS) lamps. Sci. Hortic. 2012, 147, 136–143. [Google Scholar] [CrossRef]
- Singh, D.; Basu, C.; Meinhardt-Wollweber, M.; Roth, B. LEDs for energy efficient greenhouse lighting. Renew. Sustain. Energy Rev. 2015, 49, 139–147. [Google Scholar] [CrossRef]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Bula, R.J.; Morrow, R.C.; Tibbitts, T.W.; Barta, D.J.; Ignatius, R.W.; Martin, T.S. Light-emitting diodes as a radiation source for plants. HortScience 1991, 26, 203–205. [Google Scholar] [CrossRef]
- Morrow, R.C. LED lighting in horticulture. HortScience 2008, 43, 1947–1950. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Liakopoulos, G.; Bresta, P.; Nikolopoulos, D. The optical properties of leaf structural elements and their contribution to photosynthetic performance and photoprotection. Plants 2021, 10, 1455. [Google Scholar] [CrossRef]
- Aliniaeifard, S.; Falahi, Z.; Dianati Daylami, S.; Li, T.; Woltering, E. Postharvest spectral light composition affects chilling injury in anthurium cut flowers. Front. Plant Sci. 2020, 11, 846. [Google Scholar] [CrossRef]
- Hao, Y.; Zeng, Z.; Zhang, X.; Xie, D.; Li, X.; Ma, L.; Liu, M.; Liu, H. Green means go: Green light promotes hypocotyl elongation via brassinosteroid signaling. Plant Cell 2023, koad022. [Google Scholar] [CrossRef]
- Christiaens, A.; Van Labeke, M.C.; Gobin, B.; Van Huylenbroeck, J. Rooting of ornamental cuttings affected by spectral light quality. Acta Hortic. 2015, 1104, 219–224. [Google Scholar] [CrossRef]
- Jeong, S.W.; Park, S.; Jin, J.S. Influences of four different light-emitting diode lights on flowering and polyphenol variations in the leaves of Chrysanthemum (Chrysanthemum morifolium). J. Agric. Food Chem. 2012, 60, 9793–9800. [Google Scholar] [CrossRef]
- Christiaens, A.; Gobin, B.; Van Huylenbroeck, J.; Van Labeke, M.C. Adventitious rooting of Chrysanthemum is stimulated by a low red: Far-red ratio. J. Plant Physiol. 2019, 236, 117–123. [Google Scholar] [CrossRef]
- Zheng, L.; Van Labeke, M.C. Long-term effects of red- and blue-light emitting diodes on leaf anatomy and photosynthetic efficiency of three ornamental pot plants. Front. Plant Sci. 2017, 8, 917. [Google Scholar] [CrossRef]
- Orlando, M.; Trivellini, A.; Puccinelli, M.; Ferrante, A.; Incrocci, L.; Mensuali, A. Increasing the functional quality of Crocus sativus L. by-product (tepals) by controlling spectral composition. Hortic. Environ. Biotechnol. 2022, 63, 363–373. [Google Scholar] [CrossRef]
- Heo, J.W.; Lee, C.W.; Murthy, H.N.; Paek, K.Y. Influence of light quality and photoperiod on flowering of Cyclamen persicum Mill. cv. ‘Dixie White’. Plant Growth Regul. 2003, 40, 7–10. [Google Scholar] [CrossRef]
- Aalifar, M.; Aliniaeifard, S.; Arab, M.; Zare Mehrjerdi, M.; Dianati Daylami, S.; Serek, M.; Woltering, E.; Li, T. Blue light improves vase life of carnation cut flowers through its effect on the antioxidant defense system. Front. Plant Sci. 2020, 11, 511. [Google Scholar] [CrossRef] [PubMed]
- Aalifar, M.; Aliniaeifard, S.; Arab, M.; Mehrjerdi, M.; Serek, M. Blue light postpones senescence of carnation flowers through regulation of ethylene and abscisic acid pathway-related genes. Plant Physiol. Biochem. 2020, 151, 103–112. [Google Scholar] [CrossRef]
- De Keyser, E.; Dhooghe, E.; Christiaens, A.; Van Labeke, M.-C.; Van Huylenbroeck, J. LED light quality intensifies leaf pigmentation in ornamental pot plants. Sci. Hortic. 2019, 253, 270–275. [Google Scholar] [CrossRef]
- Kobori, M.M.R.G.; Mello, S.D.C.; Freitas, I.S.D.; Silveira, F.F.; Alves, M.C.; Azevedo, R.A. Supplemental light with different blue and red ratios in the physiology, yield and quality of Impatiens. Sci. Hortic. 2022, 306, 111424. [Google Scholar] [CrossRef]
- Wollaeger, H.M.; Runkle, E.S. Growth of impatiens, petunia, salvia, and tomato seedlings under blue, green, and red light-emitting diodes. HortScience 2014, 49, 734–740. [Google Scholar] [CrossRef]
- Wojciechowska, R.; Hanus-Fajerska, E.; Kamińska, I.; Koźmińska, A.; Długosz-Grochowska, O.; Kapczyńska, A. High ratio of red-to-blue LED light improves the quality of Lachenalia ‘Rupert’ inflorescence. Folia Hortic. 2019, 31, 93–100. [Google Scholar] [CrossRef]
- Meng, Q.; Runkle, E.S. Regulation of flowering by green light depends on its photon flux density and involves cryptochromes. Phys. Plant. 2019, 166, 762–771. [Google Scholar] [CrossRef]
- Flores-Perez, S.; Castillo-Gonzalez, A.M.; Valdez-Aguilar, L.A.; Garcia-Avita, E. Use of different proportions of red and blue LEDs to improve the growth of Lilium spp. Rev. Mex. Cienc. Agric. 2021, 12, 835–847. [Google Scholar] [CrossRef]
- Park, Y.; Runkle, E.S. Far-red radiation and photosynthetic photon flux density independently regulate seedling growth but interactively regulate flowering. Environ. Exp. Bot. 2018, 155, 206–216. [Google Scholar] [CrossRef]
- Kong, Y.; Schiestel, K.; Zheng, Y. Blue light associated with low phytochrome activity can promote flowering: A comparison with red light in four bedding plant species. Acta Hortic. 2020, 1296, 433–440. [Google Scholar] [CrossRef]
- Horibe, T.; Horie, K.; Kawai, M.; Kurachi, Y.; Watanabe, Y.; Makita, M. Effect of light environment on flower opening and water balance in cut rose. Environ. Control Biol. 2020, 58, 15–20. [Google Scholar] [CrossRef]
- Matysiak, B. The effect of supplementary LED lighting on the morphological and physiological traits of Miniature Rosa × hybrida ‘Aga’ and the development of Powdery Mildew (Podosphaera pannosa) under greenhouse conditions. Plants 2021, 10, 417. [Google Scholar] [CrossRef]
- Suthaparan, A.; Torre, S.; Stensvand, A.; Herrero, M.L.; Pettersen, R.I.; Gadoury, D.M.; Gislerod, H.R. Specific light-emitting diodes can suppress sporulation of Podosphaera pannosa on greenhouse roses. Plant Dis. 2010, 94, 1105–1110. [Google Scholar] [CrossRef]
- Yamori, N.; Matsushima, Y.; Yamori, W. Upward LED lighting from the base suppresses senescence of lower leaves and promotes flowering in indoor rose management. HortScience 2021, 56, 716–721. [Google Scholar] [CrossRef]
- Owen, W.G.; Lopez, R.G. Comparison of sole-source and supplemental lighting on callus formation and initial rhizogenesis of Gaura and Salvia cuttings. HortScience 2019, 54, 684–691. [Google Scholar] [CrossRef]
- Orlando, M.; Trivellini, A.; Incrocci, L.; Ferrante, A.; Mensuali, A. The inclusion of green light in a red and blue light background impact the growth and functional quality of vegetable and flower microgreen species. Horticulturae 2022, 8, 217. [Google Scholar] [CrossRef]
- García-Caparrós, P.; Martínez-Ramírez, G.; Almansa, E.M.; Javier Barbero, F.; Chica, R.M.; Teresa Lao, M. Growth, photosynthesis, and physiological responses of ornamental plants to complementation with monochromic or mixed red-blue LEDs for use in indoor environments. Agronomy 2020, 10, 284. [Google Scholar] [CrossRef]
- Srikanth, A.; Schmid, M. Regulation of flowering time: All roads lead to Rome. Cell. Mol. Life Sci. 2011, 68, 2013–2037. [Google Scholar] [CrossRef]
- Cho, L.H.; Yoon, J.; An, G. The control of flowering time by environmental factors. Plant J. 2017, 90, 708–719. [Google Scholar] [CrossRef] [PubMed]
- Thomas, B.; Vince-Prue, D. Daylength Perception in Short-Day Plants, 2nd ed.; Academic Press: London, UK, 1996; pp. 1–427. [Google Scholar]
- Sung, S.; Amasino, R.M. Remembering winter: Toward a molecular understanding of vernalization. Annu. Rev. Plant Biol. 2005, 56, 491–508. [Google Scholar] [CrossRef] [PubMed]
- Lang, A. The effect of gibberellin upon flower formation. Proc. Natl. Acad. Sci. USA 1957, 43, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Simpson, G.C. The autonomous pathway: Epigenetic and post-transcriptional gene regulation in the control of Arabidopsis flowering time. Curr. Opin. Plant Biol. 2004, 7, 570–574. [Google Scholar] [CrossRef]
- Wang, J.W. Regulation of flowering time by the miR156-mediated age pathway. J. Exp. Bot. 2014, 65, 4723–4730. [Google Scholar] [CrossRef]
- Erwin, J. Factors affecting flowering in ornamental plants. In Flower Breeding and Genetics; Anderson, N.O., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 7–48. [Google Scholar]
- Weller, J.L.; Kendrik, R.E. Photomorphogenesis and photoperiodism in plants. In Photobiology: The Science of Light and Life; Bjorn, L.O., Ed.; Springer: New York, NY, USA, 2015; pp. 299–321. [Google Scholar]
- Proietti, S.; Scariot, V.; De Pascale, S.; Paradiso, R. Flowering mechanisms and environmental stimuli for flower transition: Bases for production scheduling in greenhouse floriculture. Plants 2022, 11, 432. [Google Scholar] [CrossRef]
- Higuchi, Y. Florigen and anti-florigen: Flowering regulation in horticultural crops. Breed. Sci. 2018, 68, 109–118. [Google Scholar] [CrossRef]
- Dixon, L.E.; Karsai, I.; Kiss, T.; Adamski, N.M.; Liu, Z.; Ding, Y.; Allard, V.; Boden, S.A.; Griffiths, S. VERNALIZATION1 controls developmental responses of winter wheat under high ambient temperatures. Development 2019, 146, dev172684. [Google Scholar] [CrossRef]
- Meng, Q.; Runkle, E.S. Moderate-intensity blue radiation can regulate flowering, but not extension growth, of several photoperiodic ornamental crops. Environ. Exp. Bot. 2017, 134, 12–20. [Google Scholar] [CrossRef]
- Park, Y.G.; Jeong, B.R. Both the quality and positioning of the night interruption light are important for flowering and plant extension growth. J. Plant Growth Regul. 2020, 39, 583–593. [Google Scholar] [CrossRef]
- Higuchi, Y.; Sumitomo, K.; Oda, A.; Shimizu, H.; Hisamatsu, T. Day light quality affects the night-break response in the short-day plant chrysanthemum, suggesting differential phytochrome-mediated regulation of flowering. J. Plant Physiol. 2012, 169, 1789–1796. [Google Scholar] [CrossRef]
- Cerdán, P.D.; Chory, J. Regulation of flowering time by light quality. Nature 2003, 423, 881–885. [Google Scholar] [CrossRef]
- Casal, J.J. Phytochromes, cryptochromes, phototropin: Photoreceptor interactions in plants. Photochem. Photobiol. 2000, 71, 1–11. [Google Scholar] [CrossRef]
- Meng, Q.; Runkle, E.S. Controlling flowering of photoperiodic ornamental crops with light-emitting diode lamps: A coordinated grower trial. HortTechnology 2014, 1, 702–711. [Google Scholar] [CrossRef]
- Garrett-Owen, W.; Meng, Q.; Lopez, R.G. Promotion of flowering from far-red radiation depends on the photosynthetic daily light integral. HortScience 2018, 53, 465–471. [Google Scholar] [CrossRef]
- Van Haeringen, C.J.; West, J.S.; Davis, F.J.; Gilbert, A.; Hadley, P.; Pearson, S.; Wheldon, A.E.; Henbest, R.G.C. The development of solid spectral filters for the regulation of plant growth. Photochem. Photobiol. 1998, 67, 407–413. [Google Scholar] [CrossRef]
- Runkle, E.S.; Heins, R.D. Specific functions of red, far red, and blue light in flowering and stem extension of long-day plants. J. Am. Soc. Hortic. Sci. 2001, 126, 275–282. [Google Scholar] [CrossRef]
- Kurilčik, A.; Miklušytė-Čanova, R.; Dapkūnienė, S.; Žilinskaitė, S.; Kurilčik, G.; Tamulaitis, G.; Duchovskis, P.; Žukauskas, A. In vitro culture of Chrysanthemum plantlets using light-emitting diodes. Open Life Sci. 2008, 3, 161–167. [Google Scholar] [CrossRef]
- Zhang, M.; Runkle, E.S. Regulating flowering and extension growth of poinsettia using red and far-red light-emitting diodes for end-of-day lighting. HortScience 2019, 54, 323–327. [Google Scholar] [CrossRef]
- Hisamatsu, T.; Sumitomo, K.; Shimizu, H. End-of-day far-red treatment enhances responsiveness to gibberellins and promotes stem extension in chrysanthemum. J. Hortic. Sci. Biotechnol. 2008, 83, 695–700. [Google Scholar] [CrossRef]
- SharathKumar, M.; Heuvelink, E.; Marcelis, L.F.M.; van Ieperen, W. Floral induction in the short-day plant chrysanthemum under blue and red extended long-days. Front. Plant Sci. 2021, 11, 610041. [Google Scholar] [CrossRef] [PubMed]
- Craig, D.S.; Runkle, E.S. A moderate to high red to far-red light ratio from light-emitting diodes controls flowering of short-day plants. J. Am. Soc. Hortic. Sci. 2013, 138, 167–172. [Google Scholar] [CrossRef]
- Park, Y.; Runkle, E.S. Blue radiation attenuates the effects of the red to far-red ratio on extension growth but not on flowering. Environ. Exp. Bot. 2019, 168, 103871. [Google Scholar] [CrossRef]
- Hamamoto, H.; Shimaji, H.; Higashide, T. Budding and bolting responses of horticultural plants to night-break treatments with LEDs of various colors. J. Agric. Meterol. 2003, 59, 103–110. [Google Scholar] [CrossRef]
- Yang, J.; Song, J.; Jeong, B.R. Low-intensity blue light supplemented during photoperiod in controlled environment induces flowering and antioxidant production in kalanchoe. Antioxidants 2022, 11, 811. [Google Scholar] [CrossRef]
- Hamamoto, H.; Yamazaki, K. Reproductive response of okra and native rosella to long-day treatment with red, blue, and green light-emitting diode lights. HortScience 2009, 5, 1494–1497. [Google Scholar] [CrossRef]
- Sampson, B.J.; Cane, J.H. Impact of enhanced ultraviolet-B radiation on flower, pollen, and nectar production. Am. J. Bot. 1999, 86, 108–114. [Google Scholar] [CrossRef]
- Bridgen, M.P. Using ultraviolet-C (UV-C) irradiation on greenhouse ornamental plants for growth regulation. Acta Hortic. 2016, 1134, 49–56. [Google Scholar] [CrossRef]
- Darras, A.I.; Vlachodimitropoulou, A.; Dimitriadis, C. Regulation of corm sprouting, growth and flowering of pot Freesia hybrida L. plants by cold and UV-C irradiation forcing. Sci. Hortic. 2019, 252, 110–112. [Google Scholar] [CrossRef]
- Li, Y.; Xin, G.F.; Liu, C.; Shi, Q.H.; Yang, F.J.; Wei, M. Effects of red and blue light on leaf anatomy, CO2 assimilation and the photosynthetic electron transport capacity of sweet pepper (Capsicum annuum L.) seedlings. BMC Plant Biol. 2020, 20, 318. [Google Scholar] [CrossRef]
- Randall, W.C.; Lopez, R.G. Comparisons of bedding plant seedlings grown under sole source light-emitting diodes (LEDs) and greenhouse supplemental lighting from LEDs and high-pressure sodium lamps. HortScience 2015, 50, 705–713. [Google Scholar] [CrossRef]
- Mao, J.; Zhang, Y.; Sang, Y.; Li, Q.; Yang, H. From the cover: A role for Arabidopsis cryptochromes and COP1 in the regulation of stomatal opening. Proc. Natl. Acad. Sci. USA 2005, 102, 12270–12275. [Google Scholar] [CrossRef]
- Yang, J.; Li, C.; Kong, D.; Guo, F.; Wei, H. Light-mediated signaling and metabolic changes coordinate stomatal opening and closure. Front. Plant Sci. 2020, 11, 601478. [Google Scholar] [CrossRef]
- Llewellyn, D.; Schiestel, K.; Zheng, Y. Light-emitting diodes can replace high-pressure sodium lighting for cut gerbera production. HortScience 2019, 54, 95–99. [Google Scholar] [CrossRef]
- Ruberti, I.; Sessa, G.; Ciolfi, A.; Possenti, M.; Carabelli, M.; Morelli, G. Plant adaptation to dynamically changing environment: The shade avoidance response. Biotechnol. Adv. 2012, 30, 1047–1058. [Google Scholar] [CrossRef]
- Dierck, R.; Dhooghe, E.; Van Huylenbroeck, J.; Van Der Straeten, D.; De Keyser, E. Light quality regulates plant architecture in different genotypes of Chrysanthemum morifolium Ramat. Sci. Hortic. 2017, 218, 177–186. [Google Scholar] [CrossRef]
- Park, I.S.; Cho, K.J.; Kim, J.; Cho, J.Y.; Lim, T.J.; Oh, W. Growth and flowering responses of petunia to various artificial light sources with different light qualities. Korean J. Hortic. Sci. Technol. 2016, 34, 55–66. [Google Scholar] [CrossRef]
- Klein, R.M. Effects of green light on biological systems. Biol. Rev. 1992, 67, 199–284. [Google Scholar] [CrossRef]
- Cao, B.; Lv, X.; Chen, Z.; Xu, K. Supplementing green light under strong sunlight improves growth and functional ingredients of ginger (Zingiber officinale Rosc.) in summer. Ind. Crop Prod. 2021, 167, 113527. [Google Scholar] [CrossRef]
- Scariot, V.; Paradiso, R.; Rogers, H.; De Pascale, S. Ethylene control in cut flowers: Classical and innovative approaches. Postharvest Biol. Technol. 2014, 97, 83–92. [Google Scholar] [CrossRef]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [PubMed]
- Horibe, T. Use of light stimuli as a postharvest technology for cut flowers. Front. Plant Sci. 2020, 11, 573490. [Google Scholar] [CrossRef] [PubMed]
- Noichinda, S.; Bodhipadma, K.; Mahamontri, C.; Narongruk, T.; Ketsa, S. Light during storage prevents loss of ascorbic acid, and increases glucose and fructose levels in Chinese kale (Brassica oleracea var. alboglabra). Postharvest Biol. Technol. 2007, 44, 312–315. [Google Scholar] [CrossRef]
- Elibox, W.; Umaharan, P. Cultivar differences in the deterioration of vase-life in cut-flowers of Anthurium andraeanum is determined by mechanisms that regulate water uptake. Sci. Hortic. 2010, 124, 102–108. [Google Scholar] [CrossRef]
- Lü, P.; Cao, J.; He, S.; Liu, J.; Li, H.; Cheng, G.; Ding, Y.; Joyce, D.C. Nano-silver pulse treatments improve water relations of cut rose cv. Movie Star flowers. Postharvest Biol. Technol. 2010, 57, 196–202. [Google Scholar] [CrossRef]
- Hunter, D.A.; Ferrante, A.; Vernieri, P.; Reid, M.S. Role of abscisic acid in perianth senescence of daffodil (Narcissus pseudonarcissus ‘Dutch Master’). Physiol. Plant. 2004, 121, 313–321. [Google Scholar] [CrossRef]
- Hunter, D.A.; Yi, M.F.; Xu, X.; Reid, M.S. Role of ethylene in perianth senescence of daffodil (Narcissus pseudonarcissus L. ‘Dutch Master’). Postharvest Biol. Technol. 2004, 32, 269–280. [Google Scholar] [CrossRef]
- Trivellini, A.; Ferrante, A.; Vernieri, P.; Serra, G. Effects of abscisic acid on ethylene biosynthesis and perception in Hibiscus rosa-sinensis L. flower development. J. Exp. Bot. 2011, 62, 5437–5452. [Google Scholar] [CrossRef]
- Trivellini, A.; Ferrante, A.; Vernieri, P.; Serra, G. Effects of promoters and inhibitors of ABA and ethylene on flower senescence of Hibiscus rosa-sinensis L. J. Plant Growth Regul. 2011, 30, 175–184. [Google Scholar] [CrossRef]
- Zhao, D.; Tao, J. Recent advances on the development and regulation of flower color in ornamental plants. Front. Plant. Sci. 2015, 6, 1–13. [Google Scholar] [CrossRef]
- Lee, K.M.; Jeong, T.Y.; Song, J.Y.; Jeong, B.R. Flower color change of Boronia heterophylla as affected by light intensity and preservative chemicals. Korean J. Hortic. Sci. Technol. 2007, 25, 458–462. [Google Scholar]
- Huang, K.L.; Miyajima, I.; Okubo, H. Effects of temperature and shade treatment on flower colors and characteristics in newly established reddish-purple tuberose (Polianthes). J. Fac. Agric. Kyushu Univ. 2000, 45, 57–63. [Google Scholar] [CrossRef]
- Zhao, D.Q.; Hao, Z.J.; Tao, J. Effects of shade on plant growth and flower quality in herbaceous peony (Paeonia lactiflora Pall.). Plant Physiol. Biochem. 2012, 61, 187–196. [Google Scholar] [CrossRef]
- Young, H.K.; Hwan, C.J.; Sun, K.K.; Yang, H.K. Effects of light quality on growth and flowering of Hibiscus syriacus L. J. Korean Soc. Hortic. Sci. 1997, 38, 272–277. [Google Scholar]
- Thoma, F.; Somborn-Schulz, A.; Schlehuber, D.; Keuter, V.; Deerberg, G. Effects of light on secondary metabolites in selected leafy greens: A review. Front. Plant Sci. 2020, 11, 497. [Google Scholar] [CrossRef]
- Erb, M.; Kliebenstein, D.J. Plant secondary metabolites as defenses, regulators, and primary metabolites: The blurred functional trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Kobayashi, M.; Kanto, T.; Fujikawa, T.; Yamada, M.; Ishiwata, M.; Satou, M.; Hisamatsu, T. Supplemental UV radiation controls rose powdery mildew disease under greenhouse conditions. Environ. Control Biol. 2013, 51, 157–163. [Google Scholar] [CrossRef]
- Meyer, P.; Van de Poel, B.; De Coninck, B. UV-B light and its application potential to reduce disease and pest incidence in crops. Hortic. Res. 2021, 8, 194. [Google Scholar] [CrossRef]
- Yoon, J.B.; Nomura, M.; Ishikura, S. Analysis of the flight activity of the cotton bollworm Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae) under yellow LED lighting. Jpn. J. Appl. Entomol. Zool. 2012, 56, 103–110. [Google Scholar] [CrossRef]
- Dielemana, J.A.; Kruidhof, H.M.; Weerheim, K. LED lighting strategies in cut flowers: Balancing plant physiology and biological control of pests. Acta Hortic. 2020, 1296, 591–604. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).