
The LHT Gene Family in Rice: Molecular Characterization, Transport Functions and Expression Analysis
Abstract
:1. Introduction
2. Results
2.1. Identification of the LHT Gene Family in Rice
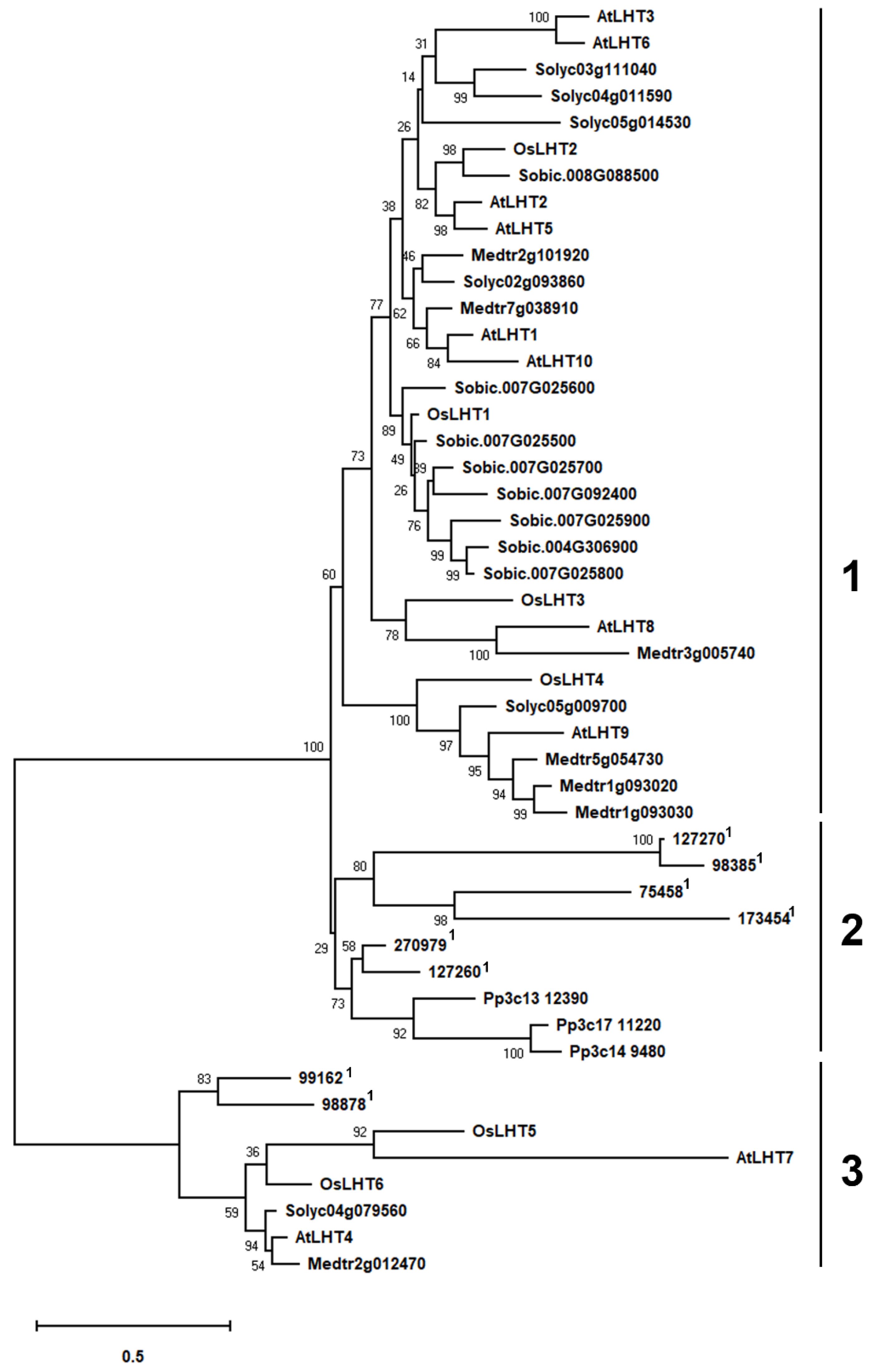
2.2. Phylogenetic Analysis of the OsLHT Gene Family
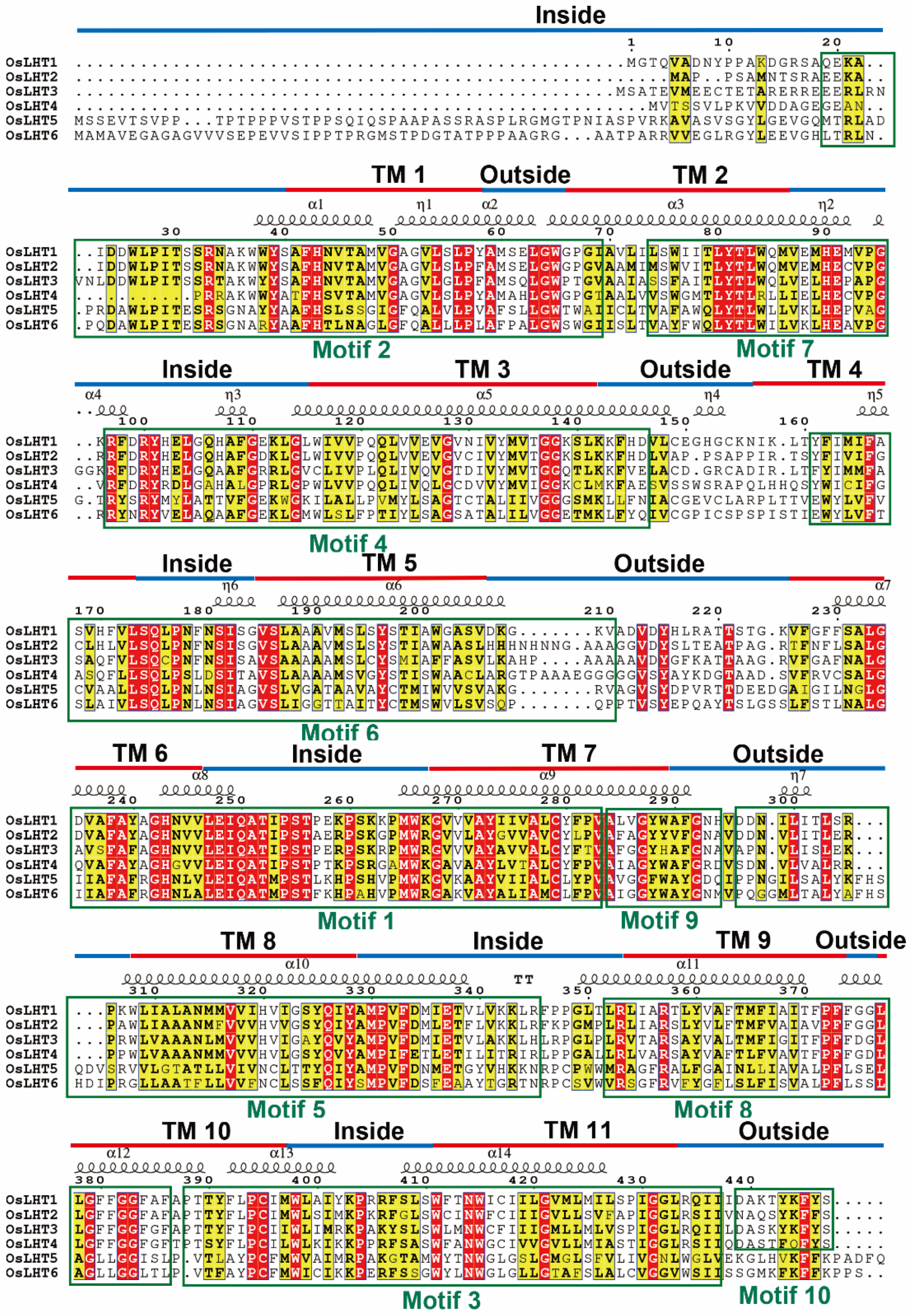
2.3. OsLHT Gene Structures and Protein Structures
2.4. The Subcellular Localization of OsLHTs
2.5. The Transport Functions of OsLHTs
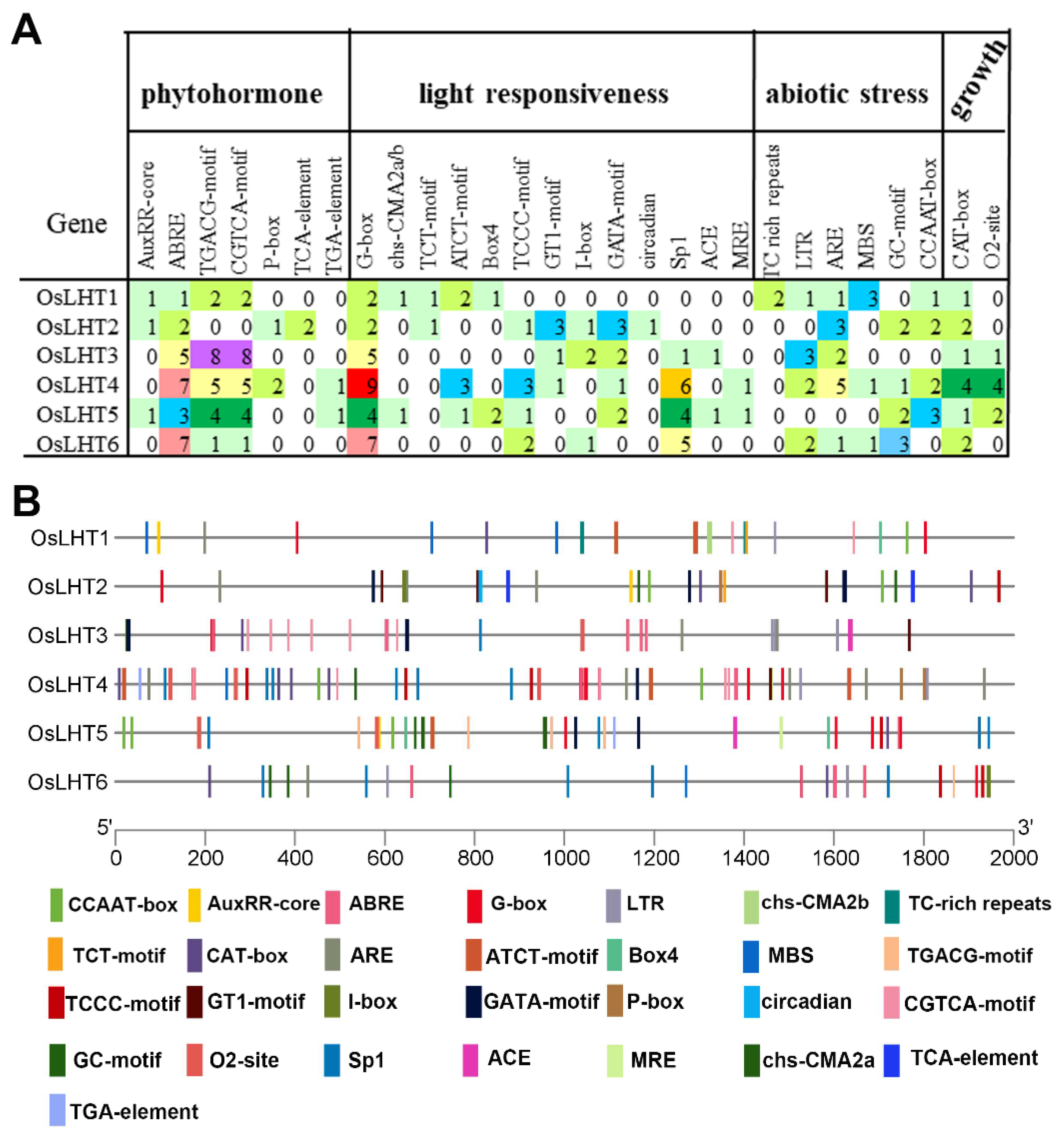
2.6. Analysis of Promoter Cis-Elements in the OsLHT Gene Family
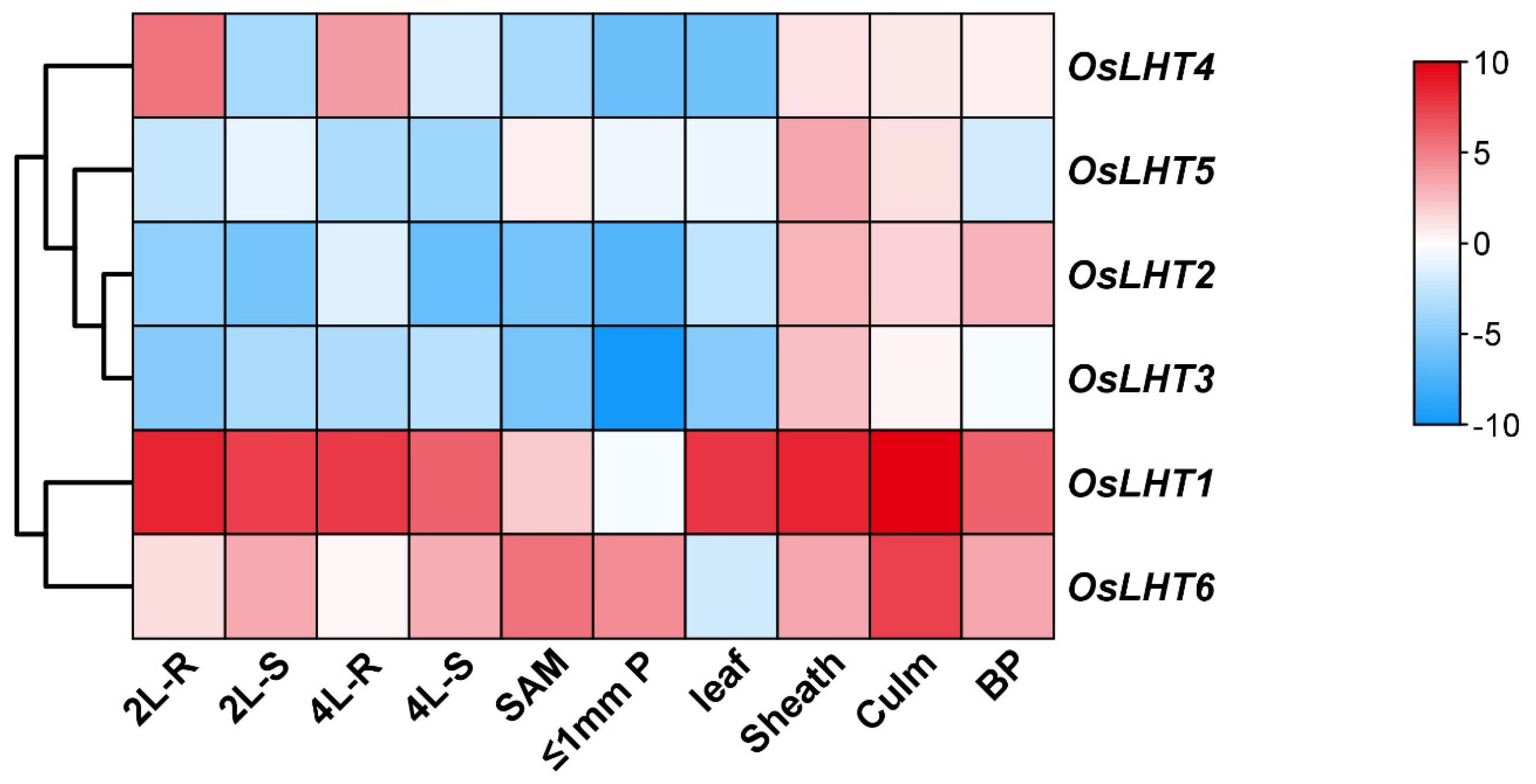
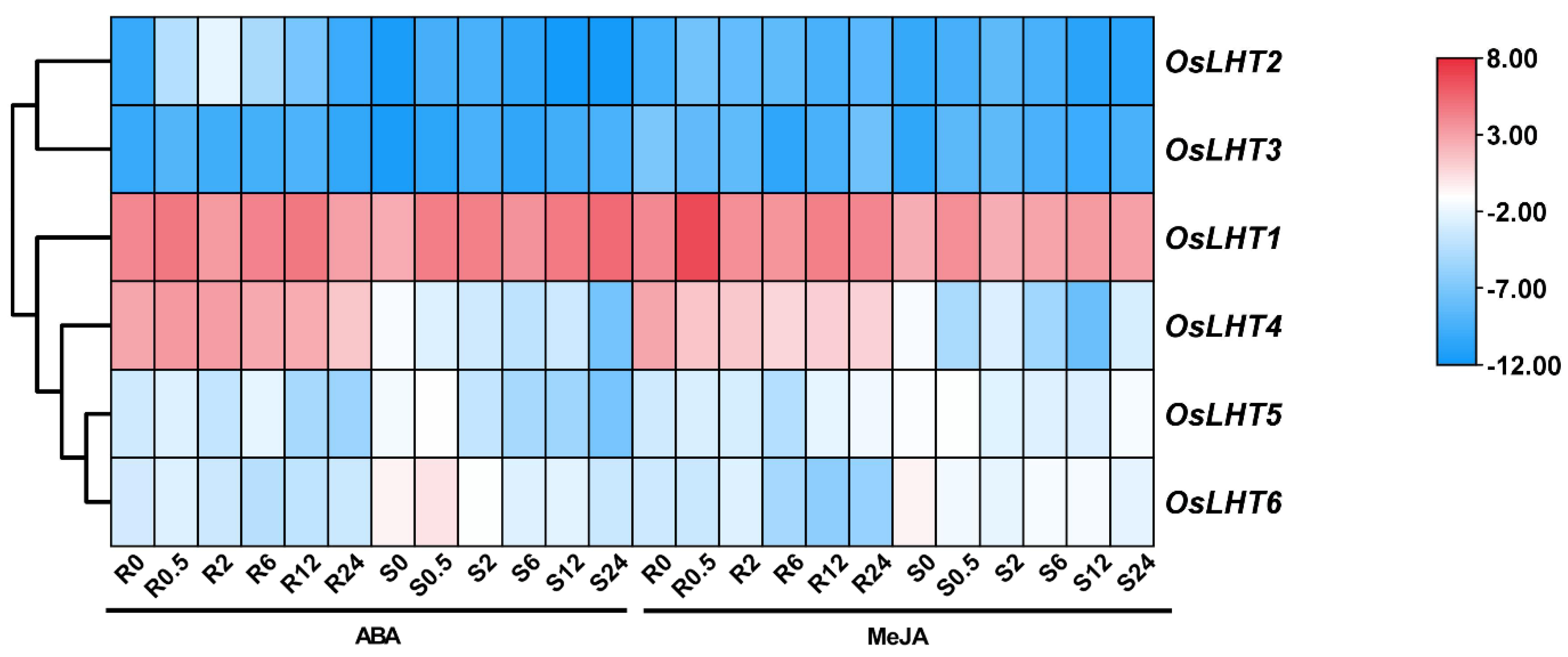
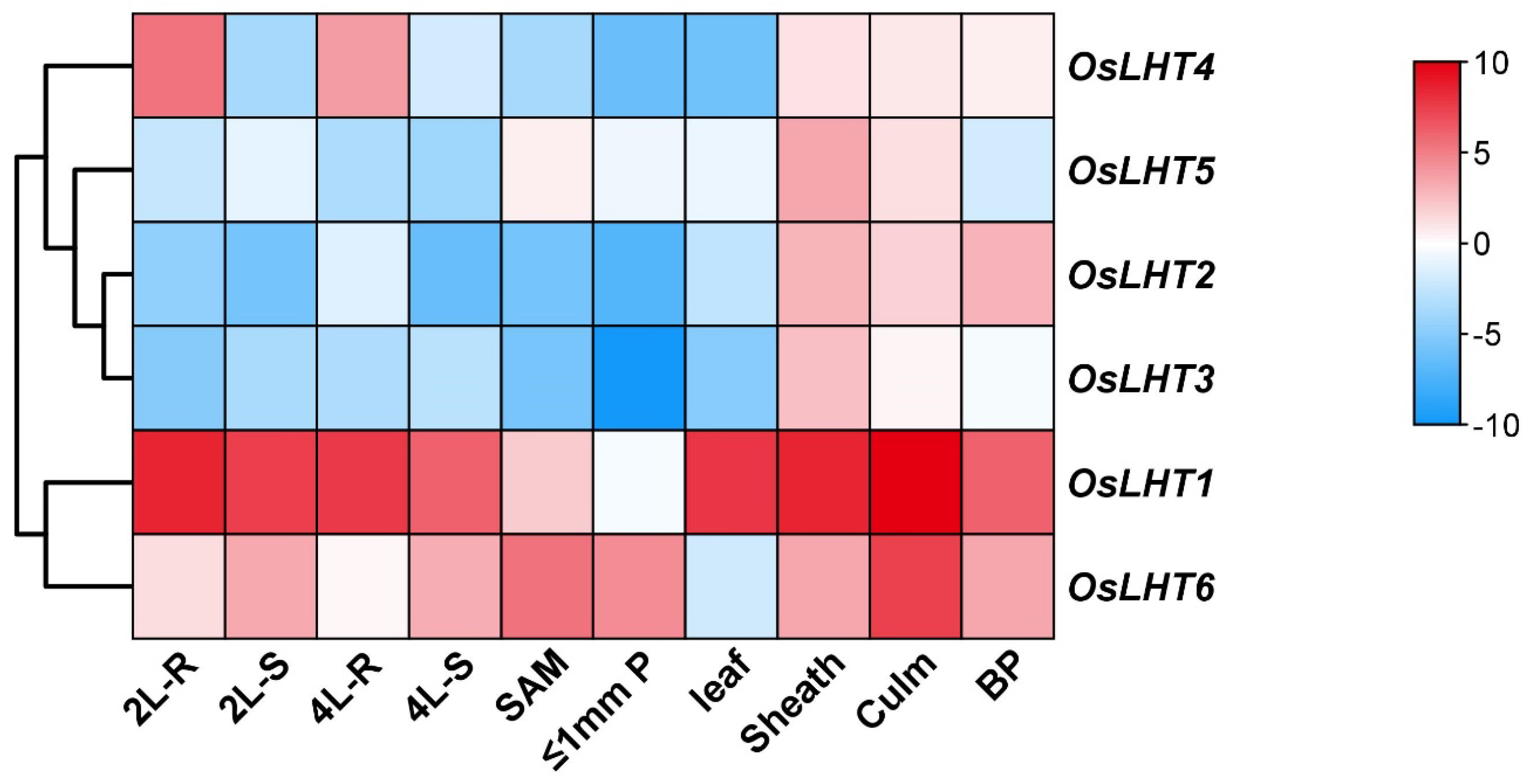
2.7. Expression Profiling of OsLHT Genes
3. Discussion
4. Materials and Methods
4.1. Identification of LHT Genes in Rice
4.2. Chromosomal Localization and Gene Duplication
4.3. Phylogenetic Tree Analysis
4.4. Gene Structures, Conserved Motifs and Three-Dimensional Modeling
4.5. Functional Characterization of OsLHT Using the 22Δ10a Yeast Strain
4.6. Plant Materials and Growth Conditions
4.7. RNA Isolation and Quantitative Real-Time PCR
4.8. Subcellular Localization Assays
4.9. Analysis of Cis-Elements in OsLHT Promoter Regions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nasholm, T.; Kielland, K.; Ganeteg, U. Uptake of organic nitrogen by plants. New Phytol. 2009, 182, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Frommer, W.B.; Hummel, S.; Riesmeier, J.W. Expression cloning in yeast of a cDNA encoding a broad specificity amino acid permease from Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1993, 90, 5944–5948. [Google Scholar] [CrossRef] [PubMed]
- Tegeder, M. Transporters for amino acids in plant cells: Some functions and many unknowns. Curr. Opin. Plant Biol. 2012, 15, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Rentsch, D.; Schmidt, S.; Tegeder, M. Transporters for uptake and allocation of organic nitrogen compounds in plants. FEBS Lett. 2007, 581, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.M.; Ma, H.L.; Yu, L.; Wang, X.; Zhao, J. Genome-wide survey and expression analysis of amino acid transporter gene family in rice (Oryza sativa L.). PLoS ONE 2012, 7, e49210. [Google Scholar] [CrossRef]
- Cheng, L.; Yuan, H.Y.; Ren, R.; Zhao, S.Q.; Han, Y.P.; Zhou, Q.Y.; Ke, D.X.; Wang, Y.X.; Wang, L. Genome-wide identification, classification, and expression analysis of amino acid transporter gene family in Glycine Max. Front. Plant Sci. 2016, 7, 515. [Google Scholar] [CrossRef]
- Wu, M.; Wu, S.N.; Chen, Z.; Dong, Q.; Yan, H.W.; Xiang, Y. Genome-wide survey and expression analysis of the amino acid transporter gene family in poplar. Tree Genet. Genomes 2015, 11, 83. [Google Scholar] [CrossRef]
- Ma, H.L.; Cao, X.L.; Shi, S.D.; Li, S.L.; Gao, J.P.; Ma, Y.L.; Zhao, Q.; Chen, Q. Genome-wide survey and expression analysis of the amino acid transporter superfamily in potato (Solanum tuberosum L.). Plant Physiol. Biochem. 2016, 107, 164–177. [Google Scholar] [CrossRef]
- Wan, Y.F.; King, R.; Mitchell, R.A.C.; Hassani-Pak, K.; Hawkesford, M.J. Spatiotemporal expression patterns of wheat amino acid transporters reveal their putative roles in nitrogen transport and responses to abiotic stress. Sci. Rep. 2017, 7, 5461. [Google Scholar] [CrossRef]
- Yang, G.Z.; Wei, Q.X.; Huang, H.; Xia, J.X. Amino Acid Transporters in Plant Cells: A Brief Review. Plants 2020, 9, 967. [Google Scholar] [CrossRef]
- Dinkeloo, K.; Boyd, S.; Pilot, G. Update on amino acid transporter functions and on possible amino acid sensing mechanisms in plants. Semin. Cell Dev. Biol. 2018, 74, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef] [PubMed]
- Denance, N.; Szurek, B.; Noel, L.D. Emerging functions of nodulin-like proteins in non-nodulating plant species. Plant Cell Physiol. 2014, 55, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Ladwig, F.; Stahl, M.; Ludewig, U.; Hirner, A.A.; Hammes, U.Z.; Stadler, R.; Harter, K.; Koch, W. Siliques are Red1 from Arabidopsis acts as a bidirectional amino acid transporter that is crucial for the amino acid homeostasis of siliques. Plant Physiol. 2012, 158, 1643–1655. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.; Lee, Y.H.; Tegeder, M. Distinct expression of members of the LHT amino acid transporter family in flowers indicates specific roles in plant reproduction. Sex Plant Reprod. 2008, 21, 143–152. [Google Scholar] [CrossRef]
- Hirner, A.; Ladwig, F.; Stransky, H.; Okumoto, S.; Keinath, M.; Harms, A.; Frommer, W.B.; Koch, W. Arabidopsis LHT1 is a high-affinity transporter for cellular amino acid uptake in both root epidermis and leaf mesophyll. Plant Cell 2006, 18, 1931–1946. [Google Scholar] [CrossRef]
- Liu, G.S.; Ji, Y.Y.; Bhuiyan, N.H.; Pilot, G.; Selvaraj, G.; Zou, J.T.; Wei, Y.D. Amino acid homeostasis modulates salicylic acid-associated redox status and defense responses in Arabidopsis. Plant Cell 2010, 22, 3845–3863. [Google Scholar] [CrossRef]
- Wang, X.H.; Yang, G.Z.; Shi, M.X.; Hao, D.L.; Wei, Q.X.; Wang, Z.G.; Fu, S.; Su, Y.H.; Xia, J.X. Disruption of an amino acid transporter LHT1 leads to growth inhibition and low yields in rice. BMC Plant Biol. 2019, 19, 268. [Google Scholar] [CrossRef]
- Guo, N.; Gu, M.J.; Hu, J.Q.; Qu, H.Y.; Xu, G.H. Rice OsLHT1 functions in leaf-to-panicle nitrogen allocation for grain yield and quality. Front. Plant Sci. 2020, 11, 1150. [Google Scholar] [CrossRef]
- Guo, N.; Hu, J.Q.; Yan, M.; Qu, H.Y.; Luo, L.; Tegeder, M.; Xu, G.H. Oryza sativa Lysine-Histidine-type Transporter 1 functions in root uptake and root-to-shoot allocation of amino acids in rice. Plant J. 2020, 103, 395–411. [Google Scholar] [CrossRef]
- Shin, K.; Lee, S.; Song, W.Y.; Lee, R.A.; Lee, I.; Ha, K.; Koo, J.C.; Park, S.K.; Nam, H.G.; Lee, Y.; et al. Genetic identification of ACC-RESISTANT2 reveals involvement of LYSINE HISTIDINE TRANSPORTER1 in the uptake of 1-aminocyclopropane-1-carboxylic acid in Arabidopsis thaliana. Plant Cell Physiol. 2015, 56, 572–582. [Google Scholar] [CrossRef]
- Choi, J.; Eom, S.; Shin, K.; Lee, R.A.; Choi, S.; Lee, J.H.; Lee, S.; Soh, M.S. Identification of lysine histidine transporter 2 as an 1-Aminocyclopropane carboxylic acid transporter in Arabidopsis thaliana by transgenic complementation approach. Front. Plant Sci. 2019, 10, 1092. [Google Scholar] [CrossRef]
- Jiang, X.Y.; Xie, Y.; Ren, Z.F.; Ganeteg, U.; Lin, F.; Zhao, C.; Xu, H.H. Design of a new glutamine-fipronil conjugate with alpha-amino acid function and Its uptake by A-thaliana lysine histidine transporter 1 (AtLHT1). J. Agric. Food Chem. 2018, 66, 7597–7605. [Google Scholar] [CrossRef]
- Chen, Y.; Yan, Y.; Ren, Z.F.; Ganeteg, U.; Yao, G.K.; Li, Z.L.; Huang, T.; Li, J.H.; Tian, Y.Q.; Lin, F.; et al. AtLHT1 Transporter Can Facilitate the Uptake and Translocation of a Glycinergic-Chlorantraniliprole Conjugate in Arabidopsis thaliana. J. Agric. Food Chem. 2018, 66, 12527–12535. [Google Scholar] [CrossRef]
- Perchlik, M.; Foster, J.; Tegeder, M. Different and overlapping functions of Arabidopsis LHT6 and AAP1 transporters in root amino acid uptake. J. Exp. Bot. 2014, 65, 5193–5204. [Google Scholar] [CrossRef]
- Besnard, J.; Pratelli, R.; Zhao, C.; Sonawala, U.; Collakova, E.; Pilot, G.; Okumoto, S. UMAMIT14 is an amino acid exporter involved in phloem unloading in Arabidopsis roots. J. Exp. Bot. 2016, 67, 6385–6397. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, X.; Zheng, J.; Men, Y.; Zhang, Y.; Liu, L.; Han, Y.; Hou, S.; Sun, Z. Amino acid transporter (AAT) gene family in Tartary buckwheat (Fagopyrum tataricum L. Gaertn.): Characterization, expression analysis and functional prediction. Int. J. Biol. Macromol. 2022, 217, 330–344. [Google Scholar] [CrossRef]
- Lee, Y.H.; Tegeder, M. Selective expression of a novel high-affinity transport system for acidic and neutral amino acids in the tapetum cells of Arabidopsis flowers. Plant J. 2004, 40, 60–74. [Google Scholar] [CrossRef]
- Tegeder, M.; Rentsch, D. Uptake and partitioning of amino acids and peptides. Mol. Plant 2010, 3, 997–1011. [Google Scholar] [CrossRef]
- Bock, K.W.; Honys, D.; Ward, J.M.; Padmanaban, S.; Nawrocki, E.P.; Hirschi, K.D.; Twell, D.; Sze, H. Integrating membrane transport with male gametophyte development and function through transcriptomics. Plant Physiol. 2006, 140, 1151–1168. [Google Scholar] [CrossRef]
- Chen, L.; Ortiz-Lopez, A.; Jung, A.; Bush, D.R. ANT1, an aromatic and neutral amino acid transporter in Arabidopsis. Plant Physiol. 2001, 125, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Schwacke, R.; Grallath, S.; Breitkreuz, K.E.; Stransky, E.; Stransky, H.; Frommer, W.B.; Rentsch, D. LeProT1, a transporter for proline, glycine betaine, and gamma-amino butyric acid in tomato pollen. Plant Cell 1999, 11, 377–392. [Google Scholar] [PubMed]
- Grallath, S.; Weimar, T.; Meyer, A.; Gumy, C.; Suter-Grotemeyer, M.; Neuhaus, J.M.; Rentsch, D. The AtProT family. Compatible solute transporters with similar substrate specificity but differential expression patterns. Plant Physiol. 2005, 137, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Chen, Z.; Luo, X.; Su, J.; Yao, G.; Xu, H.; Lin, F. Overexpression of AtAAP1 increased the uptake of an alanine-chlorantraniliprole conjugate in Arabidopsis thaliana. Environ. Sci. Pollut. Res. Int. 2019, 26, 36680–36687. [Google Scholar] [CrossRef]
- Verslues, P.E.; Sharp, R.E. Proline accumulation in maize (Zea mays L.) primary roots at low water potentials. II. Metabolic source of increased proline deposition in the elongation zone. Plant Physiol. 1999, 119, 1349–1360. [Google Scholar] [CrossRef]
- Omidbakhshfard, M.A.; Proost, S.; Fujikura, U.; Mueller-Roeber, B. Growth-Regulating Factors (GRFs): A small transcription factor family with important functions in plant biology. Mol. Plant 2015, 8, 998–1010. [Google Scholar] [CrossRef]
- Bai, X.T.; Xu, J.M.; Shao, X.M.; Luo, W.C.; Niu, Z.M.; Gao, C.Y.; Wan, D.S. A novel gene coding γ-aminobutyric acid transporter may improve the tolerance of populus euphratica to adverse environments. Front. Plant Sci. 2019, 11, 1083. [Google Scholar] [CrossRef]
- Ramesh, S.A.; Tyerman, S.D.; Gilliham, M.; Xu, B. gamma-Aminobutyric acid (GABA) signalling in plants. Cell Mol. Life Sci. 2017, 74, 1577–1603. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus ID | Gene Name | Chr a | ORF Length (bp) | Number of Amino Acids | Molecular Weight (Mw/ Da) | Theoretical pI | FL-cDNAc | WoLF PSORT |
|---|---|---|---|---|---|---|---|---|
| LOC_Os08g03350 | OsLHT1 | 8 | 1344 | 447 | 49,830.95 | 9.20 | AK102015 | PM |
| LOC_Os12g14100 | OsLHT2 | 12 | 1341 | 446 | 48,941.59 | 8.87 | AK070297 | plas: 9.5, cyto_plas: 5.5, E.R.: 2, golg: 2 |
| LOC_Os05g14820 | OsLHT3 | 5 | 1371 | 456 | 49,939.82 | 9.13 | EU956703 | plas: 12, golg: 2 |
| LOC_Os04g38860 | OsLHT4 | 4 | 1335 | 444 | 47,795.99 | 9.09 | AK243224 | plas: 8, vacu: 3, cyto: 1, E.R.: 1 |
| LOC_Os04g47420 | OsLHT5 | 4 | 1539 | 512 | 55,030.86 | 9.13 | AK060598 | chlo: 12.5, chlo_mito: 7.5 |
| LOC_Os12g30040 | OsLHT6 | 12 | 1527 | 508 | 54,950.29 | 9.06 | AK100852 | plas: 6, chlo: 3, cyto: 2, nucl: 1, mito: 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, T.; Wu, C.; Yang, W.; Lv, T.; Zhou, Y.; Tian, C. The LHT Gene Family in Rice: Molecular Characterization, Transport Functions and Expression Analysis. Plants 2023, 12, 817. https://doi.org/10.3390/plants12040817
Fan T, Wu C, Yang W, Lv T, Zhou Y, Tian C. The LHT Gene Family in Rice: Molecular Characterization, Transport Functions and Expression Analysis. Plants. 2023; 12(4):817. https://doi.org/10.3390/plants12040817
Chicago/Turabian StyleFan, Tian, Chunting Wu, Weiqi Yang, Tianxiao Lv, Yuping Zhou, and Changen Tian. 2023. "The LHT Gene Family in Rice: Molecular Characterization, Transport Functions and Expression Analysis" Plants 12, no. 4: 817. https://doi.org/10.3390/plants12040817
APA StyleFan, T., Wu, C., Yang, W., Lv, T., Zhou, Y., & Tian, C. (2023). The LHT Gene Family in Rice: Molecular Characterization, Transport Functions and Expression Analysis. Plants, 12(4), 817. https://doi.org/10.3390/plants12040817