
Analysis of the Structural Organization and Expression of the Vrn-D1 Gene Controlling Growth Habit (Spring vs. Winter) in Aegilops tauschii Coss.


Abstract
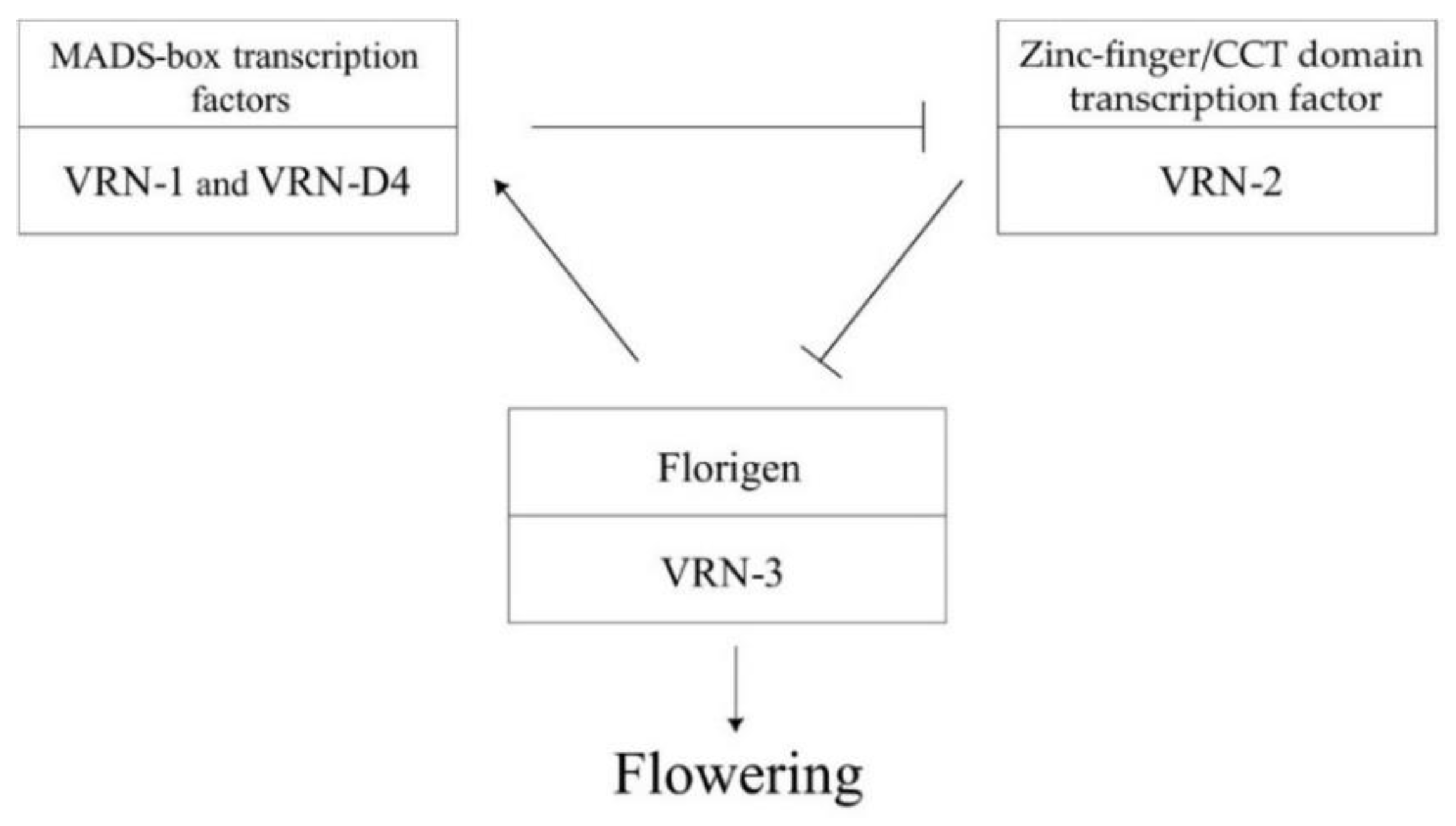
1. Introduction
2. Results
2.1. Growth Habit of the Ae. tauschii Accessions
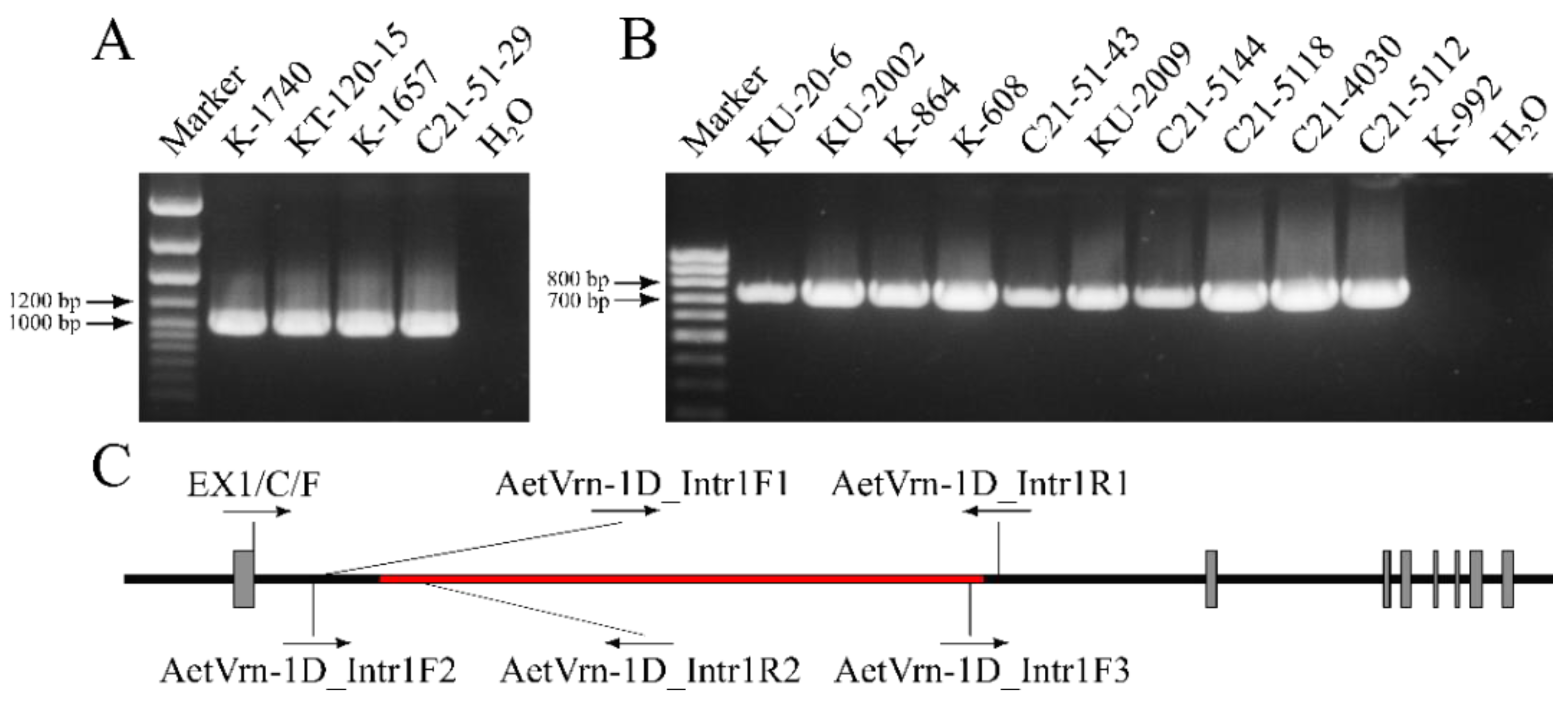
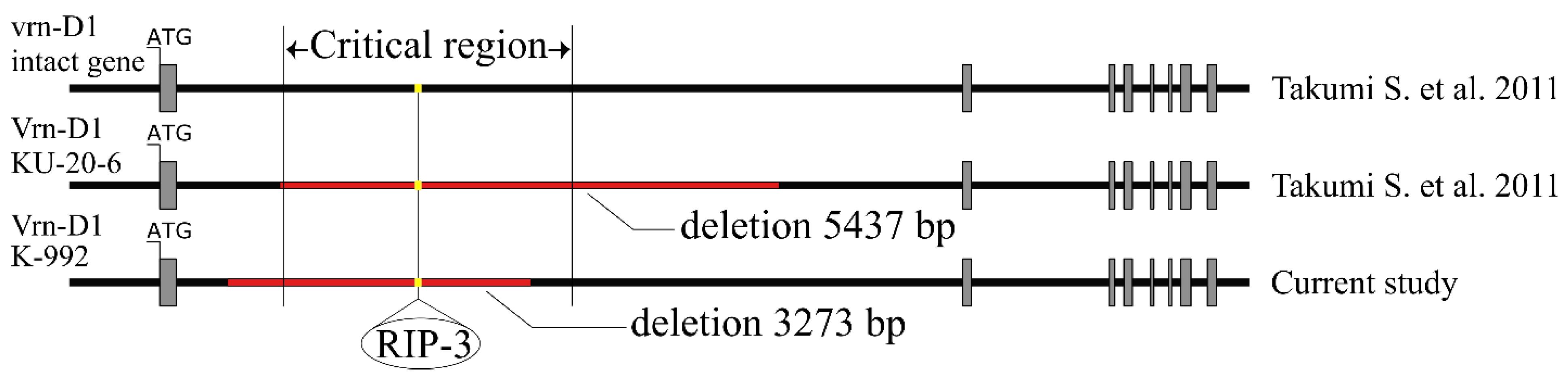
2.2. Determination of the Structure of the Vrn Genes
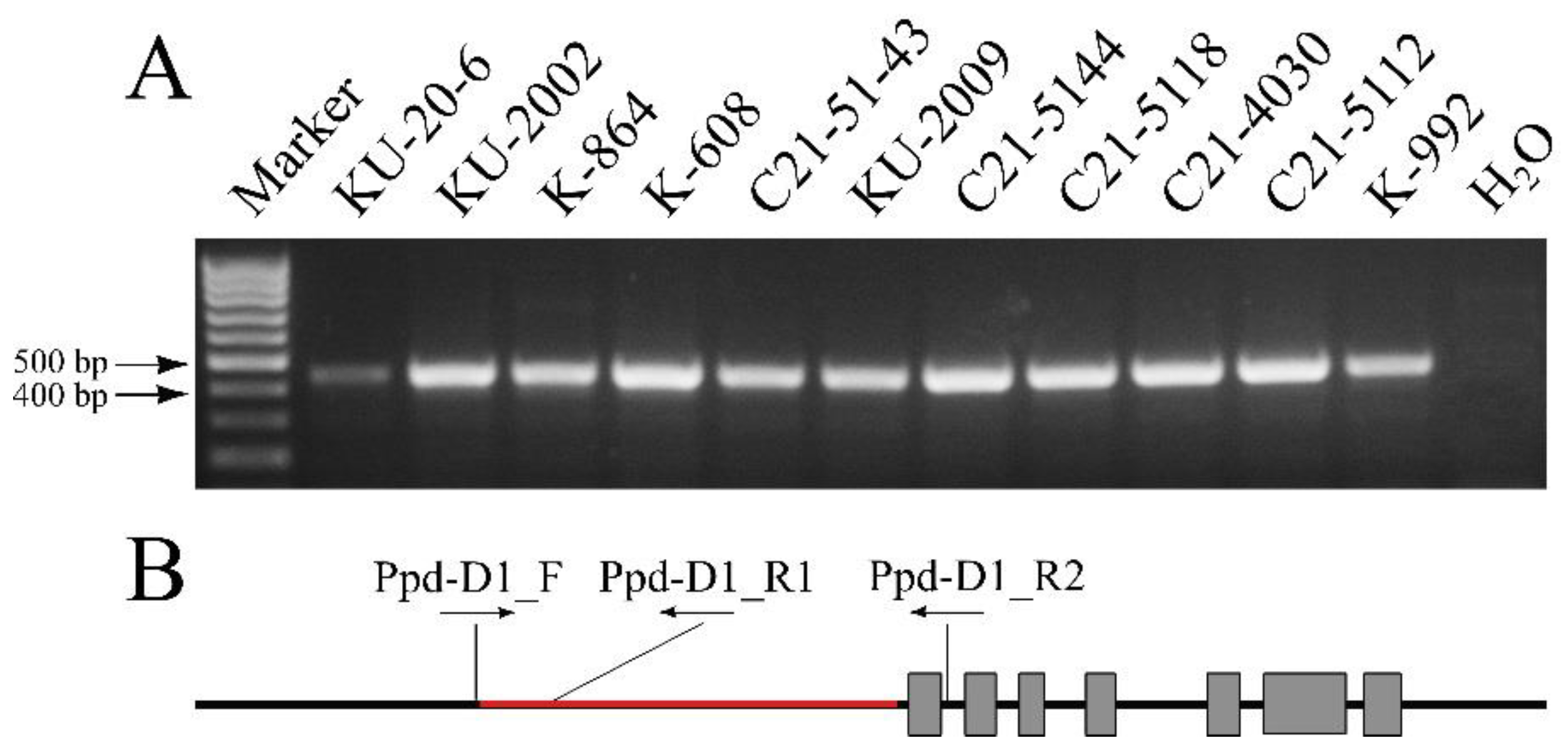
2.3. Determination of the Structure of the Ppd Genes
2.4. Analysis of the Nucleotide Sequence of the Intron of the Vrn-D1 Gene
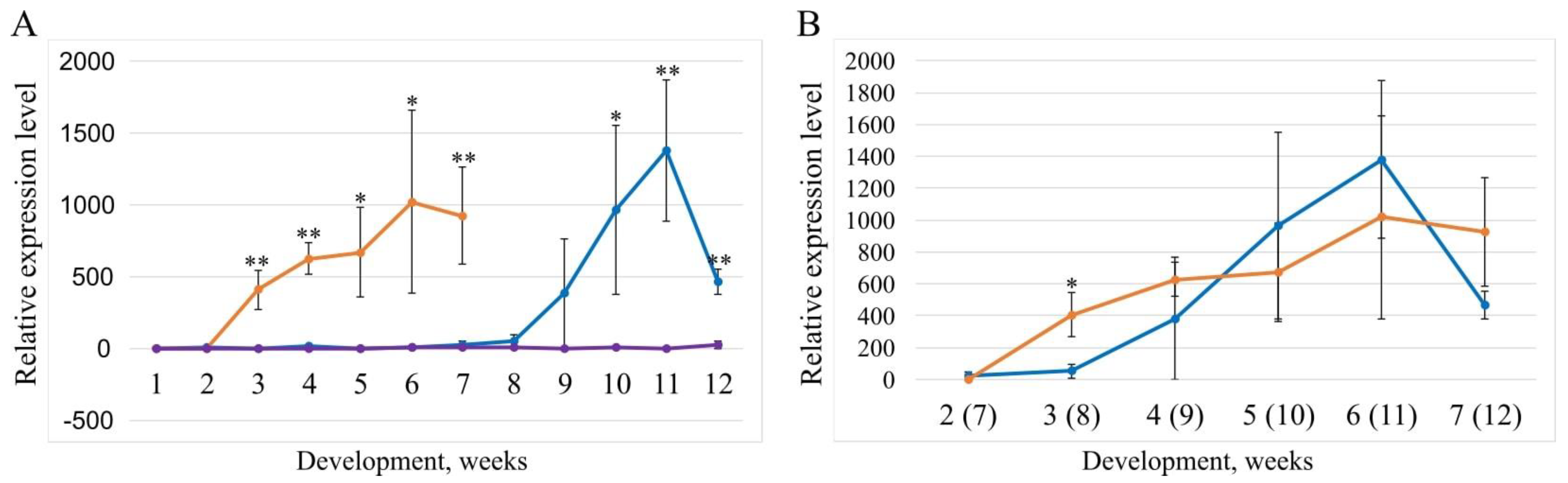
2.5. Study of the Relative Expression Levels of Different Variants of the Vrn-D1 Gene in Ae. tauschii
3. Discussion
4. Materials and Methods
4.1. Plant Material, Growth Conditions, Assessment of Vernalization Requirements and Determination of Heading Time
4.2. Isolation of Total DNA, PCR Amplification and Determination of Nucleotide Sequences of the Vrn-D1, ZCCT-D1, ZCCT-D2 and Ppd-D1 Genes
4.3. Analysis of the Expression Level of the Vrn-D1 Gene, Isolation of Total RNA, cDNA Synthesis, Real-Time PCR Amplification and Evaluation of the Expression Level
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sleper, D.A.; Poehlman, J.M. Breeding Field Crops, 5th ed.; Blackwell Pub.: Ames, IA, USA, 2006; ISBN 978-0-8138-2428-4. [Google Scholar]
- Würschum, T.; Langer, S.M.; Longin, C.F.H.; Tucker, M.R.; Leiser, W.L. A Three-component system incorporating Ppd-D1, copy number variation at Ppd-B1, and numerous small-effect quantitative trait loci facilitates adaptation of heading time in winter wheat cultivars of worldwide origin: Heading time control in wheat. Plant Cell Environ. 2018, 41, 1407–1416. [Google Scholar] [CrossRef]
- Distelfeld, A.; Li, C.; Dubcovsky, J. Regulation of flowering in temperate cereals. Curr. Opin. Plant Biol. 2009, 12, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Kamran, A.; Iqbal, M.; Spaner, D. Flowering time in wheat (Triticum aestivum L.): A key factor for global adaptability. Euphytica 2014, 197, 1–26. [Google Scholar] [CrossRef]
- Cockram, J.; Jones, H.; Leigh, F.J.; O’Sullivan, D.; Powell, W.; Laurie, D.A.; Greenland, A.J. Control of flowering time in temperate cereals: Genes, domestication, and sustainable productivity. J. Exp. Bot. 2007, 58, 1231–1244. [Google Scholar] [CrossRef]
- Flood, R.G.; Halloran, G.M. Genetics and physiology of vernalization response in wheat. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 1986; Volume 39, pp. 87–125. ISBN 978-0-12-000739-4. [Google Scholar]
- Yan, L.; Loukoianov, A.; Tranquilli, G.; Helguera, M.; Fahima, T.; Dubcovsky, J. Positional cloning of the wheat vernalization gene VRN1. Proc. Natl. Acad. Sci. USA 2003, 100, 6263–6268. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Helguera, M.; Kato, K.; Fukuyama, S.; Sherman, J.; Dubcovsky, J. Allelic variation at the VRN-1 promoter region in polyploid wheat. Theor. Appl. Genet. 2004, 109, 1677–1686. [Google Scholar] [CrossRef]
- Yan, L.; Loukoianov, A.; Blechl, A.; Tranquilli, G.; Ramakrishna, W.; SanMiguel, P.; Bennetzen, J.L.; Echenique, V.; Dubcovsky, J. The wheat VRN2 gene is a flowering repressor down-regulated by vernalization. Science 2004, 303, 1640–1644. [Google Scholar] [CrossRef]
- Yan, L.; Fu, D.; Li, C.; Blechl, A.; Tranquilli, G.; Bonafede, M.; Sanchez, A.; Valarik, M.; Yasuda, S.; Dubcovsky, J. The wheat and barley vernalization gene VRN3 is an orthologue of FT. Proc. Natl. Acad. Sci. USA 2006, 103, 19581–19586. [Google Scholar] [CrossRef]
- Fu, D.; Szűcs, P.; Yan, L.; Helguera, M.; Skinner, J.S.; von Zitzewitz, J.; Hayes, P.M.; Dubcovsky, J. Large deletions within the first intron in VRN-1 are associated with spring growth habit in barley and wheat. Mol. Genet. Genom. 2005, 273, 54–65. [Google Scholar] [CrossRef]
- Kippes, N.; Zhu, J.; Chen, A.; Vanzetti, L.; Lukaszewski, A.; Nishida, H.; Kato, K.; Dvorak, J.; Dubcovsky, J. Fine mapping and epistatic interactions of the vernalization gene VRN-D4 in hexaploid wheat. Mol. Genet. Genom. 2014, 289, 47–62. [Google Scholar] [CrossRef]
- Kippes, N.; Debernardi, J.M.; Vasquez-Gross, H.A.; Akpinar, B.A.; Budak, H.; Kato, K.; Chao, S.; Akhunov, E.; Dubcovsky, J. Identification of the VERNALIZATION 4 gene reveals the origin of spring growth habit in ancient wheats from South Asia. Proc. Natl. Acad. Sci. USA 2015, 112, E5401–E5410. [Google Scholar] [CrossRef]
- Trevaskis, B.; Bagnall, D.J.; Ellis, M.H.; Peacock, W.J.; Dennis, E.S. MADS box genes control vernalization-induced flowering in cereals. Proc. Natl. Acad. Sci. USA 2003, 100, 13099–13104. [Google Scholar] [CrossRef] [PubMed]
- Danyluk, J.; Kane, N.A.; Breton, G.; Limin, A.E.; Fowler, D.B.; Sarhan, F. TaVRT-1, a putative transcription factor associated with vegetative to reproductive transition in cereals. Plant Physiol. 2003, 132, 1849–1860. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Dubcovsky, J. Wheat TILLING mutants show that the vernalization gene VRN1 down-regulates the flowering repressor VRN2 in leaves but is not essential for flowering. PLoS Genet. 2012, 8, e1003134. [Google Scholar] [CrossRef]
- Xiao, J.; Xu, S.; Li, C.; Xu, Y.; Xing, L.; Niu, Y.; Huan, Q.; Tang, Y.; Zhao, C.; Wagner, D.; et al. O-GlcNAc-mediated interaction between VER2 and TaGRP2 elicits TaVRN1 MRNA accumulation during vernalization in winter wheat. Nat. Commun. 2014, 5, 4572. [Google Scholar] [CrossRef] [PubMed]
- Dubcovsky, J.; Loukoianov, A.; Fu, D.; Valarik, M.; Sanchez, A.; Yan, L. Effect of photoperiod on the regulation of wheat vernalization genes VRN1 and VRN2. Plant Mol. Biol. 2006, 60, 469–480. [Google Scholar] [CrossRef]
- Muterko, A.; Kalendar, R.; Salina, E. Novel alleles of the VERNALIZATION1 genes in wheat are associated with modulation of DNA curvature and flexibility in the promoter region. BMC Plant Biol. 2016, 16, 9. [Google Scholar] [CrossRef]
- Shcherban, A.B.; Khlestkina, E.K.; Efremova, T.T.; Salina, E.A. The effect of two differentially expressed wheat VRN-B1 alleles on the heading time is associated with structural variation in the first intron. Genetica 2013, 141, 133–141. [Google Scholar] [CrossRef]
- Shcherban, A.B.; Efremova, T.T.; Salina, E.A. Identification of a new Vrn-B1 allele using two near-isogenic wheat lines with difference in heading time. Mol. Breed. 2012, 29, 675–685. [Google Scholar] [CrossRef]
- Milec, Z.; Sumíková, T.; Tomková, L.; Pánková, K. Distribution of different Vrn-B1 alleles in hexaploid spring wheat germplasm. Euphytica 2013, 192, 371–378. [Google Scholar] [CrossRef]
- Santra, D.K.; Santra, M.; Allan, R.E.; Campbell, K.G.; Kidwell, K.K. Genetic and molecular characterization of vernalization genes Vrn-A1, Vrn-B1, and Vrn-D1 in spring wheat germplasm from the pacific northwest region of the U.S.A. Plant Breed. 2009, 128, 576–584. [Google Scholar] [CrossRef]
- Muterko, A.; Balashova, I.; Cockram, J.; Kalendar, R.; Sivolap, Y. The new wheat vernalization response allele Vrn-D1s is caused by DNA transposon insertion in the first intron. Plant Mol. Biol. Rep. 2015, 33, 294–303. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, M.; Wang, S.; Chen, F.; Cui, D. Allelic variation at the vernalization and photoperiod sensitivity loci in chinese winter wheat cultivars (Triticum aestivum L.). Front. Plant Sci. 2015, 6, 470. [Google Scholar] [CrossRef] [PubMed]
- Golovnina, K.A.; Kondratenko, E.; Blinov, A.G.; Goncharov, N.P. Molecular characterization of vernalization loci VRN1 in wild and cultivated wheats. BMC Plant Biol. 2010, 10, 168. [Google Scholar] [CrossRef]
- Hemming, M.N.; Peacock, W.J.; Dennis, E.S.; Trevaskis, B. Low-temperature and daylength cues are integrated to regulate FLOWERING LOCUS T in barley. Plant Physiol. 2008, 147, 355–366. [Google Scholar] [CrossRef]
- Li, C.; Dubcovsky, J. Wheat FT protein regulates VRN1 transcription through interactions with FDL2. Plant J. 2008, 55, 543–554. [Google Scholar] [CrossRef]
- Li, C.; Lin, H.; Dubcovsky, J. Factorial combinations of protein interactions generate a multiplicity of florigen activation complexes in wheat and barley. Plant J. 2015, 84, 70–82. [Google Scholar] [CrossRef]
- Kippes, N.; Chen, A.; Zhang, X.; Lukaszewski, A.J.; Dubcovsky, J. Development and characterization of a spring hexaploid wheat line with no functional VRN2 genes. Theor. Appl. Genet. 2016, 129, 1417–1428. [Google Scholar] [CrossRef]
- Distelfeld, A.; Tranquilli, G.; Li, C.; Yan, L.; Dubcovsky, J. Genetic and molecular characterization of the VRN2 loci in tetraploid wheat. Plant Physiol. 2009, 149, 245–257. [Google Scholar] [CrossRef]
- Chen, F.; Gao, M.; Zhang, J.; Zuo, A.; Shang, X.; Cui, D. Molecular characterization of vernalization and response genes in bread wheat from the Yellow and Huai Valley of China. BMC Plant Biol. 2013, 13, 199. [Google Scholar] [CrossRef]
- Fosket, D.E. Plant Growth and Development: A Molecular Approach; Academic Press: San Diego, CA, USA, 1994; ISBN 978-0-12-262430-8. [Google Scholar]
- Worland, T. Genetic basis of worldwide wheat varietal improvement. In The World Wheat Book: A History of Wheat Breeding; Lavoisier Publishing: Paris, France, 2001; pp. 59–100. [Google Scholar]
- Díaz, A.; Zikhali, M.; Turner, A.S.; Isaac, P.; Laurie, D.A. Copy Number Variation Affecting the Photoperiod-B1 and Vernalization-A1 Genes Is Associated with Altered Flowering Time in Wheat (Triticum aestivum). PLoS ONE 2012, 7, e33234. [Google Scholar] [CrossRef] [PubMed]
- Shaw, L.M.; Turner, A.S.; Laurie, D.A. The impact of photoperiod insensitive Ppd-1a mutations on the photoperiod pathway across the three genomes of hexaploid wheat (Triticum aestivum): Photoperiod-insensitive Ppd-1 mutations in wheat. Plant J. 2012, 71, 71–84. [Google Scholar] [CrossRef] [PubMed]
- Beales, J.; Turner, A.; Griffiths, S.; Snape, J.W.; Laurie, D.A. A Pseudo-Response Regulator is misexpressed in the photoperiod insensitive Ppd-D1a mutant of wheat (Triticum aestivum L.). Theor. Appl. Genet. 2007, 115, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, E.P.; Turner, A.S.; Laurie, D.A. Photoperiod insensitive Ppd-A1a mutations in Tetraploid Wheat (Triticum durum Desf.). Theor. Appl. Genet. 2009, 118, 285–294. [Google Scholar] [CrossRef]
- Turner, A.; Beales, J.; Faure, S.; Dunford, R.P.; Laurie, D.A. The Pseudo-Response Regulator Ppd-H1 Provides Adaptation to Photoperiod in Barley. Science 2005, 310, 1031–1034. [Google Scholar] [CrossRef]
- Cockram, J.; Mackay, I.J.; O’Sullivan, D.M. The Role of Double-Stranded Break Repair in the Creation of Phenotypic Diversity at Cereal VRN1 Loci. Genetics 2007, 177, 2535–2539. [Google Scholar] [CrossRef] [PubMed]
- Nakamichi, N.; Kita, M.; Niinuma, K.; Ito, S.; Yamashino, T.; Mizoguchi, T.; Mizuno, T. Arabidopsis Clock-Associated Pseudo-Response Regulators PRR9, PRR7 and PRR5 Coordinately and Positively Regulate Flowering Time Through the Canonical CONSTANS-Dependent Photoperiodic Pathway. Plant Cell Physiol. 2007, 48, 822–832. [Google Scholar] [CrossRef]
- Imaizumi, T.; Schultz, T.F.; Harmon, F.G.; Ho, L.A.; Kay, S.A. FKF1 F-Box Protein Mediates Cyclic Degradation of a Repressor of CONSTANS in Arabidopsis. Science 2005, 309, 293–297. [Google Scholar] [CrossRef]
- Nishida, H.; Yoshida, T.; Kawakami, K.; Fujita, M.; Long, B.; Akashi, Y.; Laurie, D.A.; Kato, K. Structural variation in the 5′ upstream region of photoperiod-insensitive alleles Ppd-A1a and Ppd-B1a identified in hexaploid wheat (Triticum aestivum L.), and their effect on heading time. Mol. Breed. 2013, 31, 27–37. [Google Scholar] [CrossRef]
- Campoli, C.; Drosse, B.; Searle, I.; Coupland, G.; von Korff, M. Functional characterisation of HvCO1, the barley (Hordeum vulgare) flowering time ortholog of CONSTANS: Functional characterisation of HvCO1 in barley. Plant J. 2012, 69, 868–880. [Google Scholar] [CrossRef]
- Kato, H.; Taketa, S.; Ban, T.; Iriki, K.; Murai, K. The influence of a spring habit gene, Vrn-D1, on heading time in wheat. Plant Breed. 2001, 120, 115–120. [Google Scholar] [CrossRef]
- Kato, K.; Wada, T. Genetic analysis and selection experiment for narrow-sense earliness in wheat by using segregating hybrid progenies. Breed. Sci. 1999, 49, 233–238. [Google Scholar] [CrossRef]
- Slafer, G.A.; Rawson, H.M. Responses to photoperiod change with phenophase and temperature during wheat development. Field Crops Res. 1996, 46, 1–13. [Google Scholar] [CrossRef]
- Efremova, T.T.; Chumanova, E.V.; Trubacheeva, N.V.; Arbuzova, V.S.; Belan, I.A.; Pershina, L.A. Prevalence of VRN1 locus alleles among spring common wheat cultivars cultivated in western siberia. Russ. J. Genet. 2016, 52, 146–153. [Google Scholar] [CrossRef]
- Potokina, E.K.; Koshkin, V.A.; Alekseeva, E.A.; Matvienko, I.I.; Filobok, V.A.; Bespalova, L.A. The combination of the Ppd and Vrn gene alleles determines the heading date in common wheat varieties. Russ. J. Genet. Appl. Res. 2012, 2, 311–318. [Google Scholar] [CrossRef]
- Kihara, H. Discovery of the DD-Analyser, one of the ancestors of Triticum vulgare. Agric. Hort. 1944, 19, 13–14. [Google Scholar]
- Mcfadden, E.S.; Sears, E.R. The origin of Triticum spelta and its free-threshing hexaploid relatives. J. Hered. 1946, 37, 81–89. [Google Scholar] [CrossRef]
- Nakai, Y. Isozyme cariations in Aegilops and Triticum. IV. The origin of the common wheats revealed from the study on esterase isozymes in synthesized hexaploid wheats. Jpn. J. Genet. 1979, 54, 175–189. [Google Scholar] [CrossRef]
- Nishikawa, K.; Furuta, Y.; Wada, T. Genetic studies of α-amylase isozymes in wheat. III. Intraspecific Variation in Aegilops squarrosa and Birthplace of Hexaploid Wheat. Jpn. J. Genet. 1980, 55, 325–336. [Google Scholar] [CrossRef]
- Jaaska, V. Electrophoretic survey of seedling esterases in wheats in relation to their phylogeny. Theor. Appl. Genet. 1980, 56, 273–284. [Google Scholar] [CrossRef]
- Lagudah, E.S.; Appels, R.; Brown, A.H.D.; McNeil, D. The molecular–genetic analysis of Triticum tauschii, the D-genome donor to hexaploid wheat. Genome 1991, 34, 375–386. [Google Scholar] [CrossRef]
- Lubbers, E.L.; Gill, K.S.; Cox, T.S.; Gill, B.S. Variation of molecular markers among geographically diverse accessions of Triticum tauschii. Genome 1991, 34, 354–361. [Google Scholar] [CrossRef]
- Dvorak, J.; Luo, M.-C.; Yang, Z.-L.; Zhang, H.-B. The structure of the Aegilops tauschii genepool and the evolution of hexaploid wheat. Theor. Appl. Genet. 1998, 97, 657–670. [Google Scholar] [CrossRef]
- Goncharov, N.P. Genetic resources of wheat related species: The Vrn genes controlling growth habit (Spring vs. Winter). Euphytica 1998, 100, 371–376. [Google Scholar] [CrossRef]
- Goncharov, N.P. Comparative Genetics of Wheats and Their Related Species; Novosibirsk Academic Publishing House “GEO”: Novosibirsk, Russia, 2012; ISBN 978-5-904682-90-3. [Google Scholar]
- Iwaki, K.; Nakagawa, K.; Kuno, H.; Kato, K. Ecogeographical differentiation in east Asian wheat, revealed from the geographical variation of growth habit and Vrn genotype. Euphytica 2000, 111, 137–143. [Google Scholar] [CrossRef]
- Dudnikov, A.J. Allozymes and growth habit of Aegilops tauschii: Genetic control and linkage patterns. Euphytica 2003, 129, 89–97. [Google Scholar] [CrossRef]
- Takumi, S.; Koyama, K.; Fujiwara, K.; Kobayashi, F. Identification of a large deletion in the first intron of the Vrn-D1 locus, associated with loss of vernalization requirement in wild wheat progenitor Aegilops tauschii Coss. Genes Genet. Syst. 2011, 86, 183–195. [Google Scholar] [CrossRef][Green Version]
- Dubcovsky, J.; Chen, C.; Yan, L. Molecular characterization of the allelic variation at the VRN-H2 vernalization locus in barley. Mol. Breed. 2005, 15, 395–407. [Google Scholar] [CrossRef]
- Huang, L.; Wang, Q.; Zhang, L.-Q.; Yuan, Z.-W.; Wang, J.-R.; Zhang, H.-G.; Zheng, Y.-L.; Liu, D.-C. Haplotype variations of gene Ppd-D1 in Aegilops tauschii and their implications on wheat origin. Genet. Resour. Crop Evol. 2012, 59, 1027–1032. [Google Scholar] [CrossRef]
- Guo, Z.; Song, Y.; Zhou, R.; Ren, Z.; Jia, J. Discovery, evaluation and distribution of haplotypes of the wheat Ppd-D1 gene. New Phytol. 2010, 185, 841–851. [Google Scholar] [CrossRef]
- Jones, H.; Gosman, N.; Horsnell, R.; Rose, G.A.; Everest, L.A.; Bentley, A.R.; Tha, S.; Uauy, C.; Kowalski, A.; Novoselovic, D.; et al. Strategy for exploiting exotic germplasm using genetic, morphological, and environmental diversity: The Aegilops tauschii Coss. example. Theor. Appl. Genet. 2013, 126, 1793–1808. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.; Zhang, L.; Ning, S.; Zheng, Y.-L.; Liu, D. Evaluation of Aegilops tauschii for heading date and its gene location in a re-synthesized hexaploid wheat. Agric. Sci. China 2009, 8, 1–7. [Google Scholar] [CrossRef]
- Cockram, J.; Chiapparino, E.; Taylor, S.A.; Stamati, K.; Donini, P.; Laurie, D.A.; O’Sullivan, D.M. Haplotype analysis of vernalization loci in European barley germplasm reveals novel VRN-H1 alleles and a predominant winter VRN-H1/VRN-H2 multi-locus haplotype. Theor. Appl. Genet. 2007, 115, 993–1001. [Google Scholar] [CrossRef] [PubMed]
- von Zitzewitz, J.; Szűcs, P.; Dubcovsky, J.; Yan, L.; Francia, E.; Pecchioni, N.; Casas, A.; Chen, T.H.H.; Hayes, P.M.; Skinner, J.S. Molecular and structural characterization of barley vernalization genes. Plant Mol. Biol. 2005, 59, 449–467. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yu, M.; Fang, T.; Cao, S.; Carver, B.F.; Yan, L. Vernalization requirement duration in winter wheat is controlled by TaVRN-A1 at the protein level. Plant J. 2013, 76, 742–753. [Google Scholar] [CrossRef]
- Goncharov, N.P.; Chikida, N.N. Genetics of growth habit in Aegilops squarrosa L. Genetika 1995, 31, 396–399. [Google Scholar]
- Abbas, A.; Yu, H.; Li, X.; Cui, H.; Chen, J.; Huang, P. Selection and Validation of Reference Genes for RT-QPCR Analysis in Aegilops tauschii (Coss.) under Different Abiotic Stresses. Int. J. Mol. Sci. 2021, 22, 11017. [Google Scholar] [CrossRef]
- Loukoianov, A.; Yan, L.; Blechl, A.; Sanchez, A.; Dubcovsky, J. Regulation of VRN-1 Vernalization Genes in Normal and Transgenic Polyploid Wheat. Plant Physiol. 2005, 138, 2364–2373. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Accession Location | Growth Habit | Days to Heading, Mean ± Standard Deviation | d ≥ LSD0.05 |
|---|---|---|---|---|
| KU-20-6 (Control) | Pakistan | S | 65 ± 5.2 | |
| KU-2002 | Pakistan | S | 67.5 ± 4.7 | |
| K-864 | unknown | S | 73.1 ± 2 | |
| K-608 | Georgia | S | 81.4 ± 7.6 | * |
| C21-51-43 | Pakistan | S | 84.8 ± 11 | * |
| KU-2009 | Pakistan | S | 72.8 ± 7.6 | |
| C21-5144 | Pakistan | S | 70.8 ± 5.8 | |
| C21-5118 | unknown | S | 78.8 ± 3.7 | * |
| C21-4030 | unknown | S | 77.1 ± 5.95 | * |
| C21-5112 | unknown | S | 82 ± 9.9 | * |
| K-992 | Afghanistan | S | 57 ± 5.9 | |
| LSD0.05 | 9.87 | |||
| K-1740 (Control) | Afghanistan | W | 224.8 ± 4.1 | |
| KT-120-15 | China | W | 211.5 ± 7.2 | |
| K-1657 | Palestine | W | 221.3 ± 5.3 | |
| C21-51-29 | Iran | W | 226.7 ± 7.1 | |
| LSD0.05 | 6.73 |
| Accession Number | Vrn-D1 First Intron | ZCCT-D1 First Intron | ZCCT-D2 Second Exon | Ppd-D1 Promoter |
|---|---|---|---|---|
| KU-20-6 | 5437 bp deletion | intact | intact | intact |
| KU-2002 | 5437 bp deletion | intact | intact | intact |
| K-864 | 5437 bp deletion | intact | intact | intact |
| K-608 | 5437 bp deletion | intact | intact | intact |
| C21-51-43 | 5437 bp deletion | intact | intact | intact |
| KU-2009 | 5437 bp deletion | intact | intact | intact |
| C21-5144 | 5437 bp deletion | intact | intact | intact |
| C21-5118 | 5437 bp deletion | intact | intact | intact |
| C21-4030 | 5437 bp deletion | intact | intact | intact |
| C21-5112 | 5437 bp deletion | intact | intact | intact |
| K-992 | 3273 bp deletion | intact | intact | intact |
| K-1740 | intact | nd | nd | nd |
| KT-120-15 | intact | nd | nd | nd |
| K-1657 | intact | nd | nd | nd |
| C21-51-29 | intact | nd | nd | nd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chepurnov, G.Y.; Ovchinnikova, E.S.; Blinov, A.G.; Chikida, N.N.; Belousova, M.K.; Goncharov, N.P. Analysis of the Structural Organization and Expression of the Vrn-D1 Gene Controlling Growth Habit (Spring vs. Winter) in Aegilops tauschii Coss. Plants 2023, 12, 3596. https://doi.org/10.3390/plants12203596
Chepurnov GY, Ovchinnikova ES, Blinov AG, Chikida NN, Belousova MK, Goncharov NP. Analysis of the Structural Organization and Expression of the Vrn-D1 Gene Controlling Growth Habit (Spring vs. Winter) in Aegilops tauschii Coss. Plants. 2023; 12(20):3596. https://doi.org/10.3390/plants12203596
Chicago/Turabian StyleChepurnov, Grigory Yurievich, Ekaterina Sergeevna Ovchinnikova, Alexander Genadevich Blinov, Nadezhda Nikolaevna Chikida, Mariya Khasbulatovna Belousova, and Nikolay Petrovich Goncharov. 2023. "Analysis of the Structural Organization and Expression of the Vrn-D1 Gene Controlling Growth Habit (Spring vs. Winter) in Aegilops tauschii Coss." Plants 12, no. 20: 3596. https://doi.org/10.3390/plants12203596
APA StyleChepurnov, G. Y., Ovchinnikova, E. S., Blinov, A. G., Chikida, N. N., Belousova, M. K., & Goncharov, N. P. (2023). Analysis of the Structural Organization and Expression of the Vrn-D1 Gene Controlling Growth Habit (Spring vs. Winter) in Aegilops tauschii Coss. Plants, 12(20), 3596. https://doi.org/10.3390/plants12203596