

Compost and Phosphorus/Potassium-Solubilizing Fungus Effectively Boosted Quinoa’s Physio-Biochemical Traits, Nutrient Acquisition, Soil Microbial Community, and Yield and Quality in Normal and Calcareous Soils

 , , , ,
, , , ,
Abstract
1. Introduction
2. Results
2.1. Photosynthetic Pigments
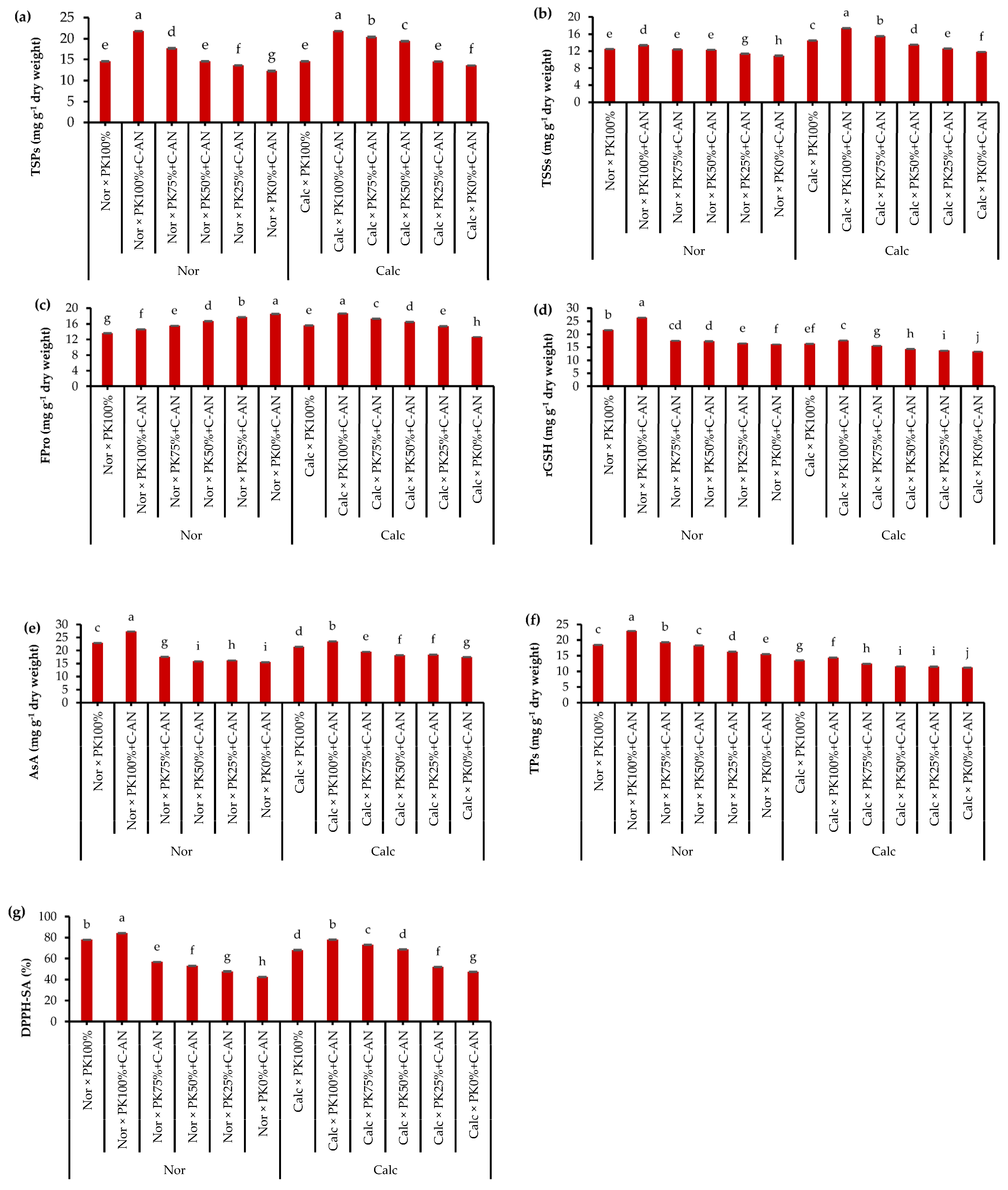
2.2. Total Soluble Proteins, Osmoregulatory Compounds, and Nonenzyme Antioxidants
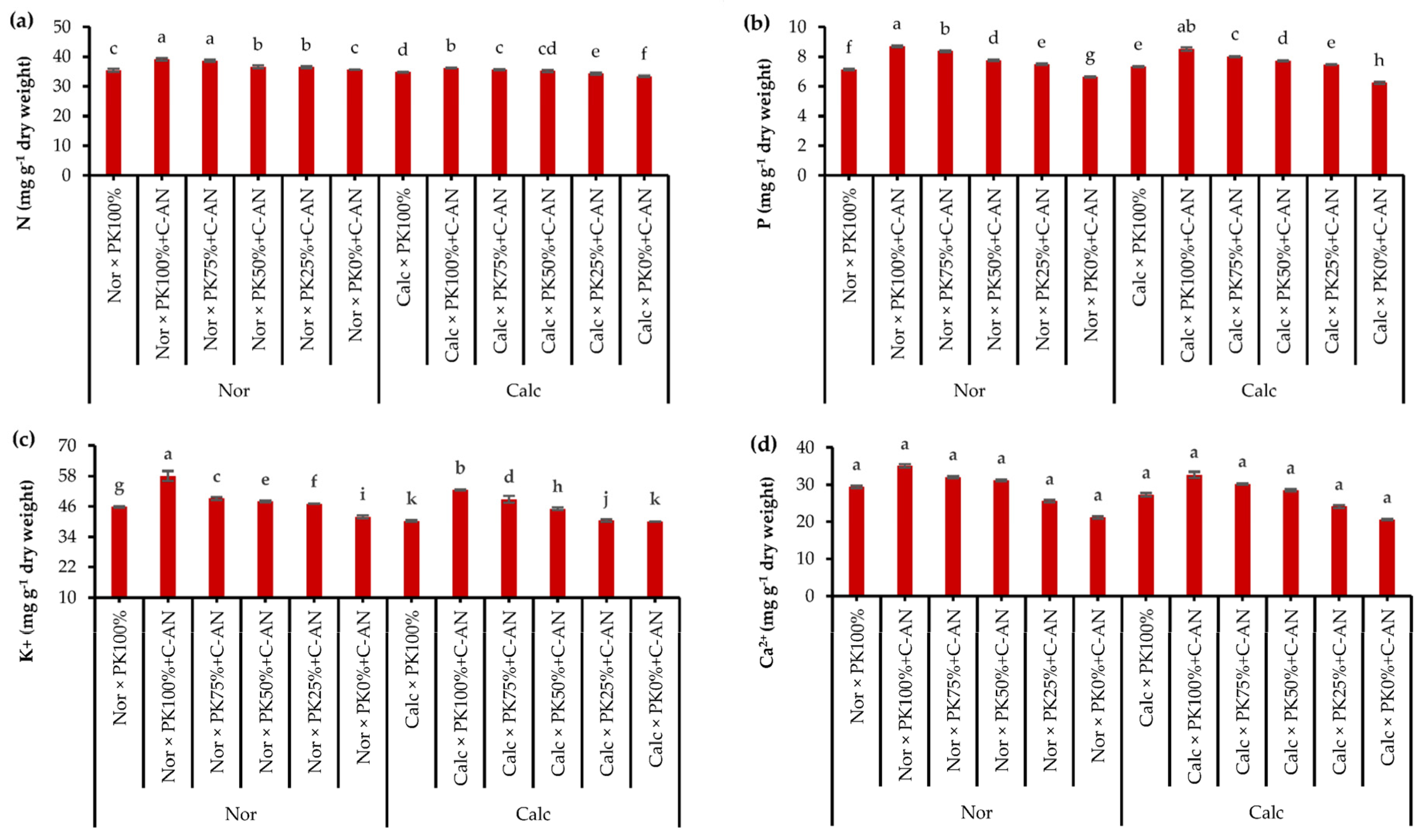
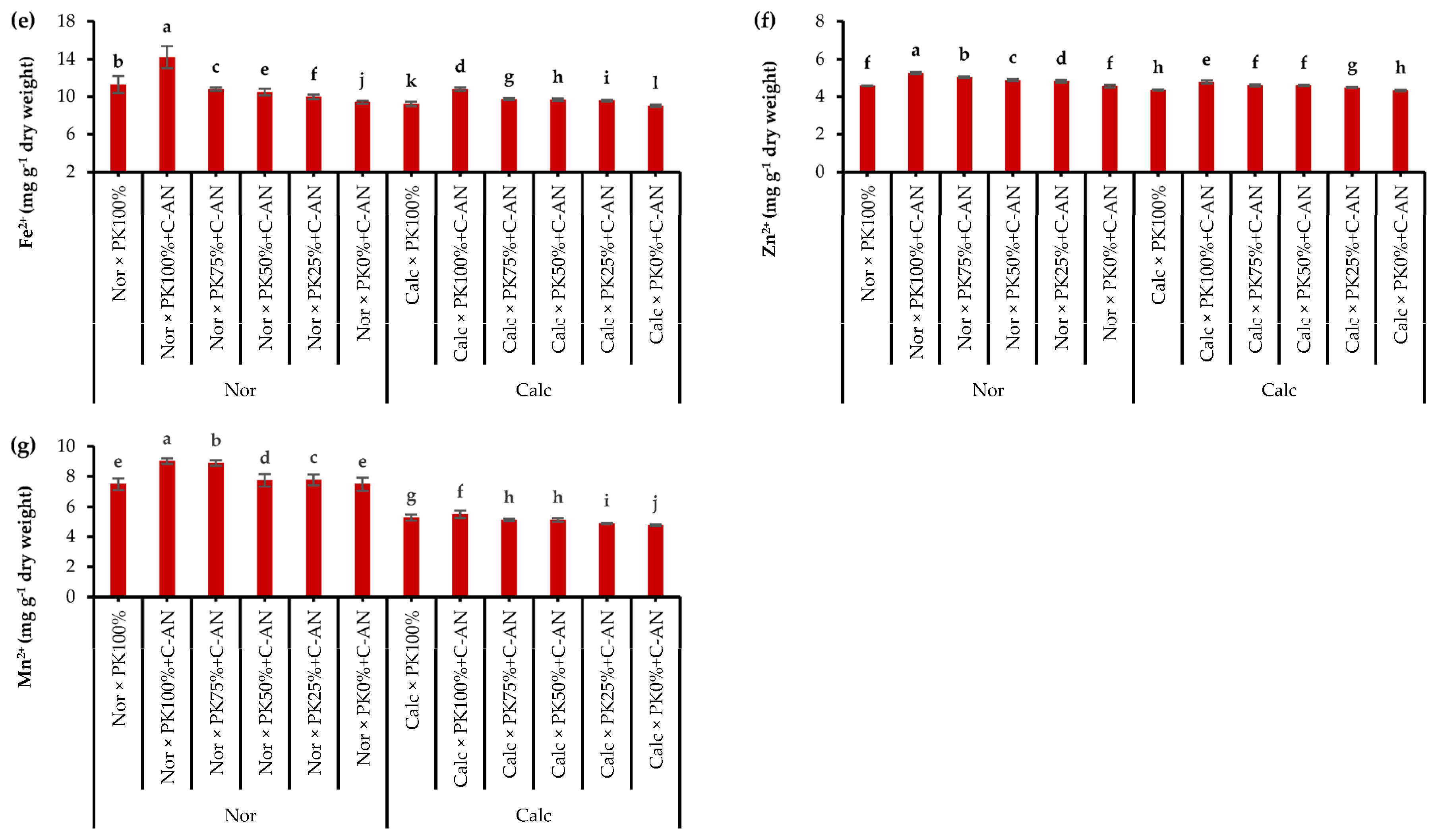
2.3. Quinoa Leaf Nutrient Contents
2.4. Seed Quality
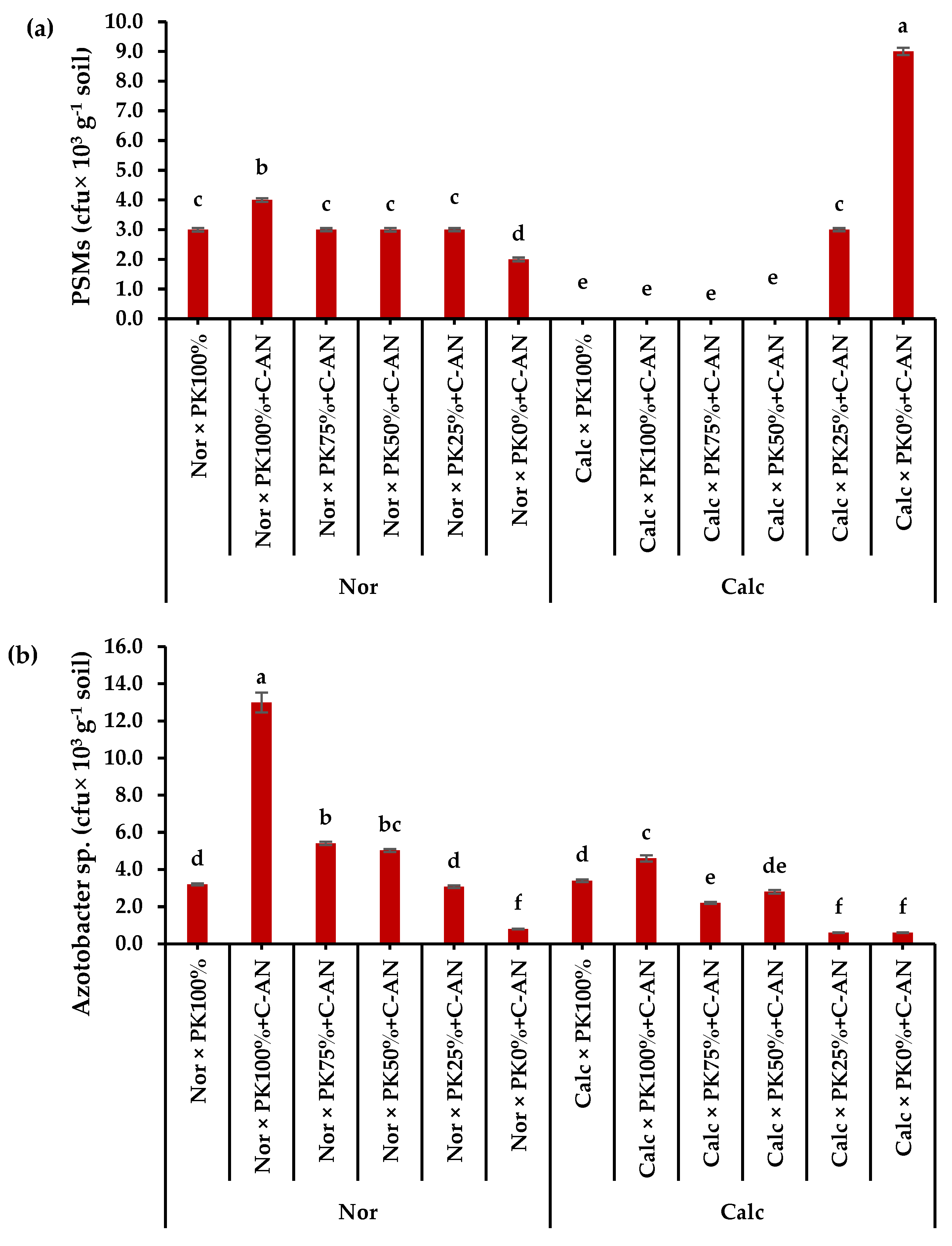
2.5. Microbial Community
2.6. Quinoa’s Growth, Yield and Yield-Related Attributes, and Harvest Index
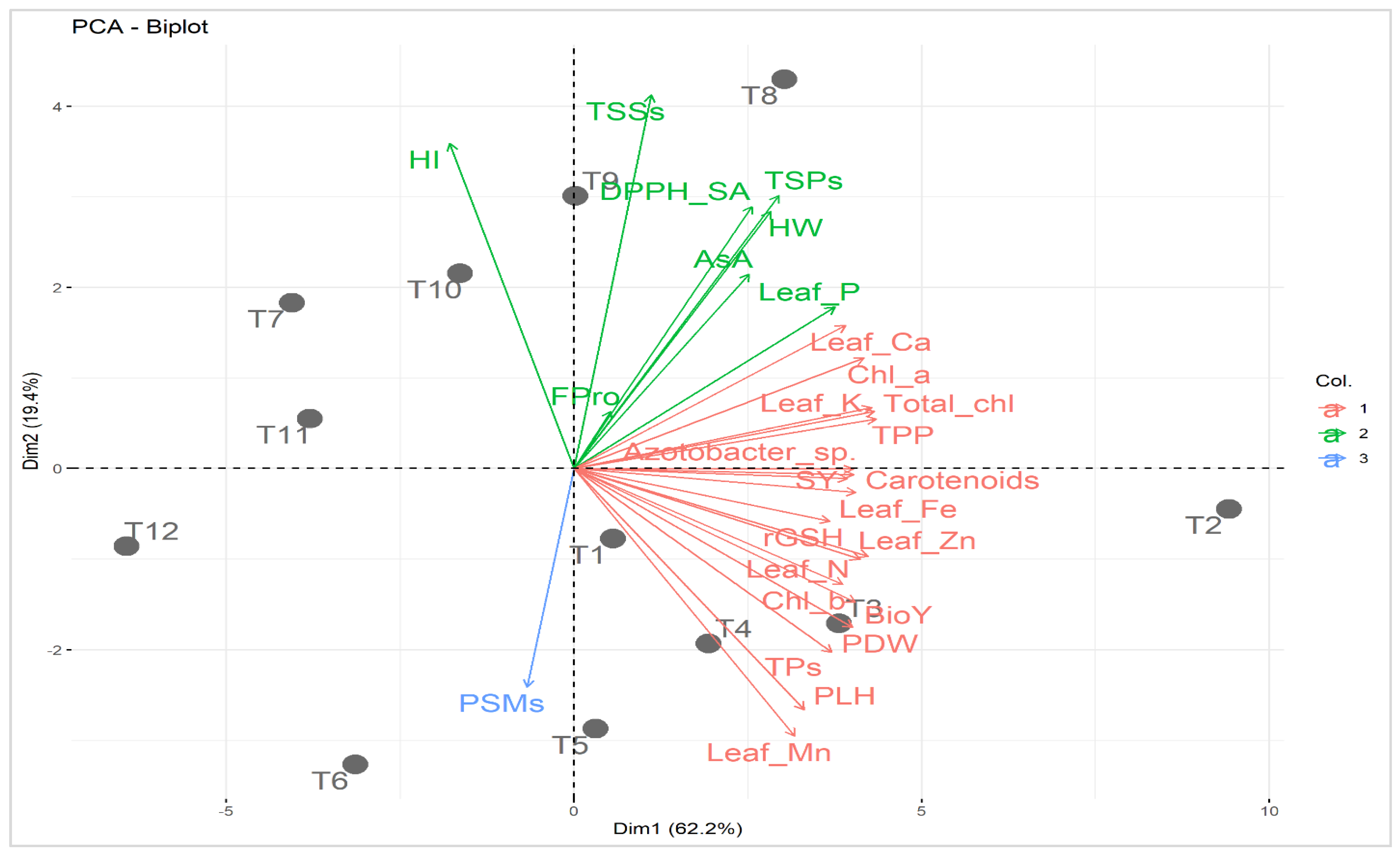
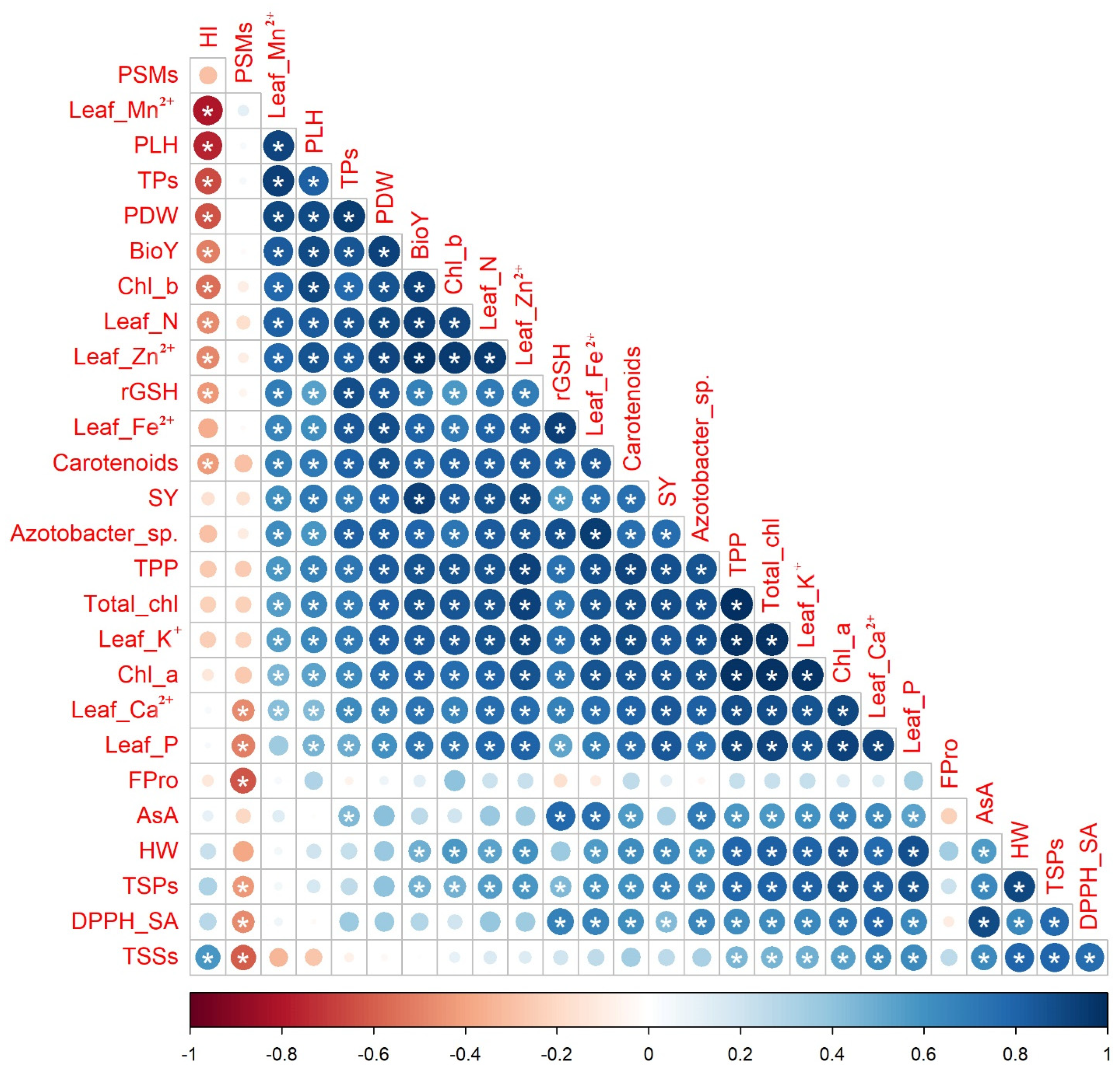
2.7. Relationship among Applied Treatments and Studied Attributes
3. Discussion
4. Materials and Methods
4.1. Experimental Location and Climatic Conditions
4.2. Fungi Isolation and Assessment of Its Phosphate and Potassium Solubilizing Ability
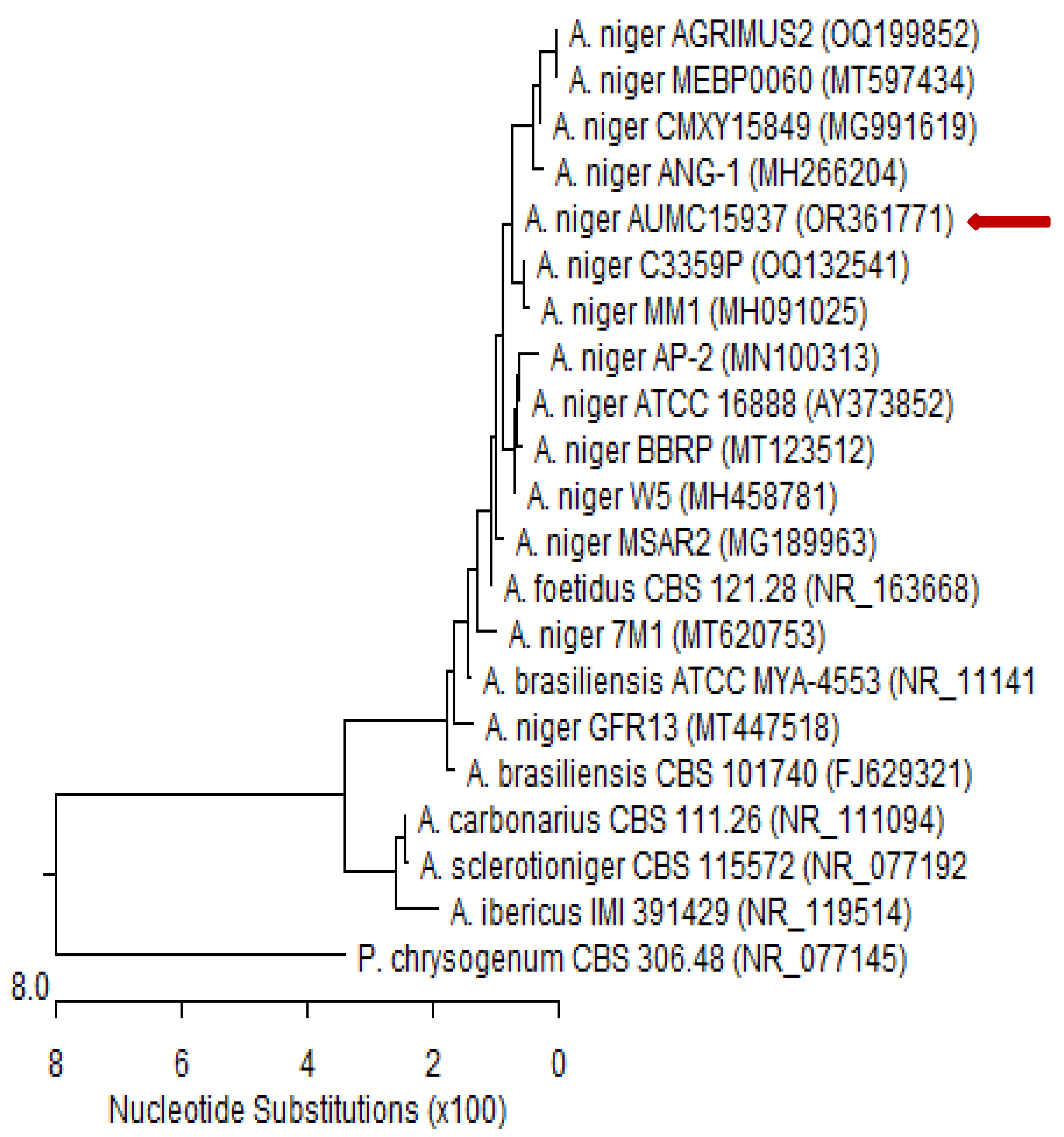
4.3. Molecular Identification of the Tested Fungal Isolate
4.4. Preparation of Aspergillus niger Inoculum
4.5. Compost Preparation
4.6. Treatments, Experimental Design, and Agronomic Management
4.7. Sampling and Measurements
4.7.1. Photosynthetic Pigment Assay
4.7.2. Osmoprotectants, Nonenzymatic Antioxidants, and 2,2-diphenyl-1-picrylhydrazyl-Scavenging Activity (DPPH-SA)
4.7.3. Determination of Macro- and Micronutrient Contents in Quinoa’s Leaf and Seed
4.7.4. Quinoa Seed’s Proximate Chemical Composition
4.7.5. Quinoa Seed’s Phytochemical and Antioxidant Activity
4.7.6. Soil Microbial Evaluation
4.7.7. Quinoa’s Growth, Yield and Yield-Related Attributes, and Harvest Index
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wassif, M.M.; Wassif, O.M. Sustainable soil management to mitigate soil erosion hazards in Egypt. In Management and Development of Agricultural and Natural Resources in Egypt’s Desert; Springer: Cham, Switzerland, 2021; pp. 163–211. [Google Scholar]
- Alghamdi, S.A.; Alharby, H.F.; Abdelfattah, M.A.; Mohamed, I.A.; Hakeem, K.R.; Rady, M.M.; Shaaban, A. Spirulina platensis-inoculated humified compost boosts rhizosphere soil hydro-physico-chemical properties and Atriplex nummularia forage yield and quality in an arid saline calcareous soil. J. Soil Sci. Plant Nutr. 2023, 23, 2215–2236. [Google Scholar] [CrossRef]
- Beheshti, M.; Alikhani, H.A.; Pourbabaee, A.A.; Etesami, H.; Rahmani, H.A.; Noroozi, M. Enriching periphyton with phosphate-solubilizing microorganisms improves the growth and concentration of phosphorus and micronutrients of rice plant in calcareous paddy soil. Rhizosphere 2022, 24, 100590. [Google Scholar] [CrossRef]
- Mirzaee, S.; Ghorbani-Dashtaki, S. Deriving and evaluating hydraulics and detachment models of rill erosion for some calcareous soils. Catena 2018, 164, 107–115. [Google Scholar] [CrossRef]
- Souei, A.; Zouaghi, T. Using statistical models and GIS to delimit the groundwater recharge potential areas and to estimate the infiltration rate: A case study of Nadhour-Sisseb-El Alem Basin, Tunisia. J. Arid Land 2021, 13, 1122–1141. [Google Scholar] [CrossRef]
- Barka, H.A.F.; Benzaghta, M.A.; Kasheem, A.M. Effect of different organic matters on chemical properties of calcareous soil. Sirte Univ. Sci. J. 2018, 8, 101–110. [Google Scholar]
- Taalab, A.S.; Ageeb, G.W.; Siam, H.S.; Mahmoud, S.A. Some characteristics of calcareous soils. A review. Middle East J. 2019, 8, 96–105. [Google Scholar]
- FAO. FAO Soils Portal: Management of Calcareous Soils. 2016. Available online: http://www.fao.org/soils-portal/soil-management/managementof-some-problem-soils/calcareous-soils/ar/ (accessed on 1 April 2023).
- Mekdad, A.A.A.; El-Sherif, A.M.A.; Rady, M.M.; Shaaban, A. Culture management and application of humic acid in favor of Helianthus annuus L. oil yield and nutritional homeostasis in a dry environment. J. Soil Sci. Plant Nutr. 2022, 22, 71–86. [Google Scholar] [CrossRef]
- Mahidi, S.S.; Hassan, G.I.; Hussain, A.; Rasool, F. Phosphorus availability issue-Its fixation and role of phosphate solubilizing bacteria in phosphate solubilization-Case study. Res. J. Agric. Sci. 2011, 2, 174–179. [Google Scholar]
- Shaaban, A.; El-Mageed, T.A.A.; El-Momen, W.R.A.; Saudy, H.S.; Al-Elwany, O.A.A.I. The integrated application of phosphorous and zinc affects the physiological status, yield and quality of canola grown in phosphorus-suffered deficiency saline soil. In Gesunde Pflanzen; Springer: Berlin/Heidelberg, Germany, 2023. [Google Scholar] [CrossRef]
- Zakirullah, M.; Khalil, S. Timing and rate of phosphorus application influence maize phenology, yield and profitability in Northwest Pakistan. Int. J. Plant Prod. 2012, 4, 281–292. [Google Scholar]
- Naeem, A.; Akhtar, M.; Ahmad, W. Optimizing available phosphorus in calcareous soils fertilized with diammonium phosphate and phosphoric acid using Freundlich adsorption isotherm. Sci. World J. 2013, 2013, 680257. [Google Scholar] [CrossRef]
- Rawat, P.; Das, S.; Shankhdhar, D.; Shankhdhar, S.C. Phosphate-solubilizing microorganisms: Mechanism and their role in phosphate solubilization and uptake. J. Soil Sci. Plant Nutr. 2021, 21, 49–68. [Google Scholar] [CrossRef]
- Hussain, S.; Sharif, M.; Ahmad, W. Selection of efficient phosphorus solubilizing bacteria strains and mycorrhizea for enhanced cereal growth, root microbe status and N and P uptake in alkaline calcareous soil. Soil Sci. Plant Nutr. 2021, 67, 259–268. [Google Scholar] [CrossRef]
- Wahid, F.; Fahad, S.; Danish, S.; Adnan, M.; Yue, Z.; Saud, S.; Siddiqui, M.H.; Brtnicky, M.; Hammerschmiedt, T.; Datta, R. Sustainable management with mycorrhizae and phosphate solubilizing bacteria for enhanced phosphorus uptake in calcareous soils. Agriculture 2020, 10, 334. [Google Scholar] [CrossRef]
- Li, Z.; Bai, T.; Dai, L.; Wang, F.; Tao, J.; Meng, S.; Hu, Y.; Wang, S.; Hu, S. A study of organic acid production in contrasts between two phosphate solubilizing fungi: Penicillium oxalicum and Aspergillus niger. Sci. Rep. 2016, 6, 25313. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Xia, J.; Zhou, N.; Xu, M.; Li, X.; Zhang, L.; Du, S.; Gao, H. The utilization of phosphogypsum as a sustainable phosphate-based fertilizer by Aspergillus niger. Agronomy 2022, 12, 646. [Google Scholar] [CrossRef]
- Naeem, U.; Afzaal, M.; Qazi, A.; Yasar, A.; Mahfooz, Y.; Naz, A.U.; Awan, H. Investigating the effect of Aspergillus niger inoculated press mud (biofertilizer) on the potential of enhancing maize (Zea mays L.) yield, potassium use efficiency and potassium agronomic efficiency. Cereal Res. Commun. 2022, 50, 157–170. [Google Scholar] [CrossRef]
- Abd El-Mageed, T.A.; Mekdad, A.A.A.; Rady, M.O.A.; Abdelbaky, A.S.; Saudy, H.S.; Shaaban, A. Physio-biochemical and agronomic changes of two sugar beet cultivars grown in saline soil as influenced by potassium fertilizer. J. Soil Sci. Plant Nutr. 2022, 22, 3636–3654. [Google Scholar] [CrossRef]
- Attia, M.M. Status of potassium in some calcareous soils of Egypt and factors affecting its forms. Ann. Agric. Sci. Moshtohor. 2019, 57, 177–184. [Google Scholar] [CrossRef][Green Version]
- Basak, B.B.; Maity, A.; Ray, P.; Biswas, D.R.; Roy, S. Potassium supply in agriculture through biological potassium fertilizer: A promising and sustainable option for developing countries. Arch. Agron. Soil Sci. 2022, 68, 101–114. [Google Scholar] [CrossRef]
- Sattar, A.; Naveed, M.; Ali, M.; Zahir, Z.A.; Nadeem, S.M.; Yaseen, M.; Meena, V.S.; Farooq, M.; Singh, R.; Rahman, M.; et al. Perspectives of potassium solubilizing microbes in sustainable food production system: A review. Appl. Soil Ecol. 2019, 133, 146–159. [Google Scholar] [CrossRef]
- Kour, D.; Rana, K.L.; Yadav, A.N.; Yadav, N.; Kumar, M.; Kumar, V.; Vyas, P.; Dhaliwal, H.S.; Saxena, A.K. Microbial biofertilizers: Bioresources and eco-friendly technologies for agricultural and environmental sustainability. Biocatal. Agric. Biotechnol. 2020, 23, 101487. [Google Scholar] [CrossRef]
- Lian, B.; Wang, B.; Pan, M.; Liu, C.Q.; Teng, H.H. Microbial release of potassium from K-bearing minerals by thermophilic fungus Aspergillus fumigatus. Geochim. Cosmochim. Acta 2008, 72, 87–98. [Google Scholar] [CrossRef]
- Meena, V.S.; Maurya, B.R.; Verma, J.P. Does a rhizospheric microorganism enhance K+ availability in agricultural soils? Microbiol. Res. 2014, 169, 337–347. [Google Scholar] [CrossRef]
- Pinzari, F.; Cuadros, J.; Jungblut, A.D.; Najorka, J.; Humphreys-Williams, E. Fungal strategies of potassium extraction from silicates of different resistance as manifested in differential weathering and gene expression. Geochim. Cosmochim. Acta 2022, 316, 168–200. [Google Scholar] [CrossRef]
- Zhang, C.; Kong, F. Isolation and identification of potassium-solubilizing bacteria from tobacco rhizospheric soil and their effect on tobacco plants. Appl. Soil Ecol. 2014, 82, 18–25. [Google Scholar] [CrossRef]
- Qiang, X.; Ding, J.; Lin, W.; Li, Q.; Xu, C.; Zheng, Q.; Li, Y. Alleviation of the detrimental effect of water deficit on wheat (Triticum aestivum L.) growth by an indole acetic acid-producing endophytic fungus. Plant Soil 2019, 439, 373–391. [Google Scholar] [CrossRef]
- Sun, J.; Xu, G.; Shao, H.; Xu, S. Potential retention and release capacity of phosphorus in the newly formed wetland soils from the Yellow River Delta, China. Clean–Soil Air Water 2012, 40, 1131–1136. [Google Scholar] [CrossRef]
- Wang, L.; Liu, L.; Zheng, B. Eutrophication development and its key regulating factors in a water-supply reservoir in North China. J. Environ. Sci. 2013, 25, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Mundim, G.d.S.M.; Maciel, G.M.; Mendes, G.d.O. Aspergillus niger as a biological input for improving vegetable seedling production. Microorganisms 2022, 10, 674. [Google Scholar] [CrossRef]
- Cihangir, N. Stimulation of the gibberellic acid synthesis by Aspergillus niger in submerged culture using a precursor. World J. Microbiol. Biotechnol. 2002, 18, 727–729. [Google Scholar] [CrossRef]
- Hung, R.; Lee Rutgers, S. Applications of Aspergillus in Plant Growth Promotion, New and Future Developments in Microbial Biotechnology and Bioengineering: Aspergillus System Properties and Applications; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Ni, H.; Wu, Y.; Zong, R.; Ren, S.; Pan, D.; Yu, L.; Li, J.; Qu, Z.; Wang, Q.; Zhao, G.; et al. Combination of Aspergillus niger MJ1 with Pseudomonas stutzeri DSM4166 or mutant Pseudomonas fluorescens CHA0-nif improved crop quality, soil properties, and microbial communities in barrier soil. Front. Microbiol. 2023, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Min, T.; Ru, S.; Li, J. Response of cotton root growth and rhizosphere soil bacterial communities to the application of acid compost tea in calcareous soil. Appl. Soil Ecol. 2022, 177, 104523. [Google Scholar] [CrossRef]
- Aboukila, E.F.; Nassar, I.N.; Rashad, M.; Hafez, M.; Norton, J.B. Reclamation of calcareous soil and improvement of squash growth using brewer’s spent grain and compost. J. Saudi Soc. Agric. Sci. 2018, 17, 390–397. [Google Scholar]
- Luo, T.; Zhu, Y.; Lu, W.; Chen, L.; Min, T.; Li, J.; Wei, C. Acidic compost tea enhances phosphorus availability and cotton yield in calcareous soils by decreasing soil pH. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2021, 71, 657–666. [Google Scholar] [CrossRef]
- Estrada-Bonilla, G.A.; Durrer, A.; Cardoso, E.J. Use of compost and phosphate-solubilizing bacteria affect sugarcane mineral nutrition, phosphorus availability, and the soil bacterial community. Appl. Soil Ecol. 2021, 157, 103760. [Google Scholar] [CrossRef]
- Sánchez, Ó.J.; Ospina, D.A.; Montoya, S. Compost supplementation with nutrients and microorganisms in composting process. Waste Manag. 2017, 69, 136–153. [Google Scholar] [CrossRef] [PubMed]
- Kiani-Pouya, A.; Roessner, U.; Jayasinghe, N.S.; Lutz, A.; Rupasinghe, T.; Bazihizina, N.; Bohm, J.; Alharbi, S.; Hedrich, R.; Shabala, S. Epidermal bladder cells confer salinity stress tolerance in the halophyte quinoa and Atriplex species. Plant Cell Environ. 2017, 40, 1900–1915. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, S.H.; Solgi, S.; Sepaskhah, A.R. Quinoa: A super or pseudo-super crop? Evidences from evapotranspiration, root growth, crop coefficients, and water productivity in a hot and semi-arid area under three planting densities. Agric. Water Manag. 2019, 225, 105784. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Tagliazucchi, D.; Babini, E.; Rutella, G.S.; Saa, D.L.T.; Gianotti, A. Bioactive peptides from vegetable food matrices: Research trends and novel biotechnologies for synthesis and recovery. J. Funct. Foods 2016, 27, 549–569. [Google Scholar] [CrossRef]
- Adel, H. Towards expanding quinoa cultivation in Egypt: The effect of compost and vermicompost on quinoa pests, natural enemies and yield under field conditions. Agric. Sci. 2020, 11, 191–209. [Google Scholar] [CrossRef]
- Bazile, D.; Bertero, H.D.; Nieto, C. State of the Art Report on Quinoa Around the World in 2013; FAO&CIRAD: Roma, Italy, 2015. [Google Scholar]
- Hinojosa, L.; Gonz’alez, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Rady, M.O.A.; Marey, R.A.; Abd El-Mageed, T.A. Exogenously applied proline enhances growth and productivity of drought stressed onion by improving photosynthetic efficiency, water use efficiency and up-regulating osmoprotectants. Sci. Hortic. 2020, 272, 109580. [Google Scholar] [CrossRef]
- Abd El-Mageed, T.A.; Abdelkhalik, A.; Abd El-Mageed, S.A.; Semida, W.M. Co-composted poultry litter biochar enhanced soil quality and eggplant productivity under different irrigation regimes. J. Soil Sci. Plant Nutr. 2021, 21, 1917–1933. [Google Scholar] [CrossRef]
- Shaaban, A.; Al-Elwany, O.A.A.I.; Abdou, N.M.; Hemida, K.A.; El-Sherif, A.M.A.; Abdel-Razek, M.A.; Semida, W.M.; Mohamed, G.F.; Abd El-Mageed, T.A. Filter mud enhanced yield and soil properties of water-stressed Lupinus termis L. in saline calcareous soil. J. Soil Sci. Plant Nutr. 2022, 22, 1572–1588. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Hossain, M.S.; Al Mahmud, J.; Hossen, M.S.; Masud, A.A.C.; Moumita; Fujita, M. Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agronomy 2018, 8, 31. [Google Scholar] [CrossRef]
- Malhotra, H.; Vandana; Sharma, S.; Pandey, R. Phosphorus nutrition: Plant growth in response to deficiency and excess. In Plant Nutrients and Abiotic Stress Tolerance; Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K., Hawrylak-Nowak, B., Eds.; Springer: Singapore, 2018; pp. 171–190. [Google Scholar]
- Meng, X.; Chen, W.W.; Wang, Y.Y.; Huang, Z.R.; Ye, X.; Chen, L.S.; Yang, L.T. Effects of phosphorus deficiency on the absorption of mineral nutrients, photosynthetic system performance and antioxidant metabolism in Citrus grandis. PLoS ONE 2021, 16, e0246944. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shu, A.; Song, W.; Shi, W.; Li, M.; Zhang, W.; Li, Z.; Liu, G.; Yuan, F.; Zhang, S.; et al. Long-term organic fertilizer substitution increases rice yield by improving soil properties and regulating soil bacteria. Geoderma 2021, 404, 115287. [Google Scholar] [CrossRef]
- Semida, W.M.; Abd El-Mageed, T.A.; Howladar, S.M. A novel organo-mineral fertilizer can alleviate negative effects of salinity stress for eggplant production on reclaimed saline calcareous soil. Acta Hort. 2014, 1034, 493–500. [Google Scholar] [CrossRef]
- Alghamdi, S.A.; Al-Ghamdi, F.A.; El-Zohri, M.; Al-Ghamdi, A.A.M. Modifying of Calcareous Soil with Some Acidifying Materials and Its Effect on Helianthus Annuus (L.) Growth. Saudi J. Biol. Sci. 2023, 30, 103568. [Google Scholar] [CrossRef]
- Kranz, C.N.; McLaughlin, R.A.; Johnson, A.; Miller, G.; Heitman, J.L. The effects of compost incorporation on soil physical properties in urban soils—A concise Review. J. Environ. Manag. 2020, 261, 110209. [Google Scholar] [CrossRef]
- Wu, Q.F.; Hu, H.B.; He, L.M. Effect of aspergillus niger strain xf-1 on soil nutrients and growth of Amorpha fruticosa. Appl. Ecol. Environ. Res. 2020, 18, 5079–5092. [Google Scholar] [CrossRef]
- Qaswar, M.; Chai, R.; Ahmed, W.; Jing, H.; Han, T.; Liu, K.; Ye, X.; Xu, Y.; Anthonio, C.K.; Zhang, H. Partial substitution of chemical fertilizers with organic amendments increased rice yield by changing phosphorus fractions and improving phosphatase activities in fluvo-aquic soil. J. Soils Sediments 2020, 20, 1285–1296. [Google Scholar] [CrossRef]
- Abdelkhalik, A.; Abd El-mageed, T.A.; Mohamed, I.A.A.; Semida, W.M.; Al-elwany, O.A.A.I.; Ibrahim, I.M.; Hemida, K.A.; El-saadony, M.T.; Abuqamar, S.F.; El-tarabily, K.A. Soil application of effective microorganisms and nitrogen alleviates salt stress in hot pepper (Capsicum annum L.) plants. Front. Plant Sci. 2023, 13, 1079260. [Google Scholar] [CrossRef]
- Khan, K.S.; Ali, M.M.; Naveed, M.; Ishaq, M.; Rehmani, A.; Waleed, M.; Ali, H.M.; Abdelsalam, N.R.; Ghareeb, R.Y.; Feng, G. Co-application of organic amendments and inorganic P increase maize growth and soil carbon, phosphorus availability in calcareous soil. Front. Environ. Sci. 2022, 10, 949371. [Google Scholar] [CrossRef]
- Abdelkhalik, A.; Pascual, B.; Nájera, I.; Baixauli, C.; Pascual-Seva, N. Deficit irrigation as a sustainable practice in improving irrigation water use efficiency in cauliflower under Mediterranean conditions. Agronomy 2019, 9, 732. [Google Scholar] [CrossRef]
- Rady, M.M.; Mossa, A.T.H.; Youssof, A.M.; Osman, A.S.; Ahmed, S.M.; Mohamed, I.A. Exploring the reinforcing effect of nano-potassium on the antioxidant defense system reflecting the increased yield and quality of salt-stressed squash plants. Sci. Hortic. 2023, 308, 111609. [Google Scholar] [CrossRef]
- Semida, W.M.; Abd El-mageed, T.A.; Abdelkhalik, A.; Hemida, K.A.; Abdurrahman, H.A.; Howladar, S.M.; Leilah, A.A.A.; Rady, M.O.A. Selenium modulates antioxidant activity, osmoprotectants, and photosynthetic efficiency of onion under saline soil conditions. Agronomy 2021, 11, 855. [Google Scholar] [CrossRef]
- Zulfiqar, F.; Akram, N.A.; Ashraf, M. Osmoprotection in plants under abiotic stresses: New insights into a classical phenomenon. Planta 2020, 251, 3. [Google Scholar] [CrossRef] [PubMed]
- Abd El-mageed, T.A.; Gyushi, M.A.H.; Hemida, K.A.; El-Saadony, M.T.; Abd El-Mageed, S.A.; Abdalla, H.; AbuQamar, S.F.; El-Tarabily, K.A.; Abdelkhalik, A. Coapplication of effective microorganisms and nanomagnesium boosts the defenses against salt stress in Ipomoea batatas. Front. Plant Sci. 2022, 13, 883274. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ispizua, E.; Calatayud, Á.; Marsal, J.I.; Basile, F.; Cannata, C.; Abdelkhalik, A.; Soler, S.; Valcárcel, J.V.; Martínez-Cuenca, M.-R. Postharvest changes in the nutritional properties of commercial and traditional lettuce varieties in relation with overall visual quality. Agronomy 2022, 12, 403. [Google Scholar] [CrossRef]
- Martínez-Ispizua, E.; Calatayud, Á.; Marsal, J.I.; Cannata, C.; Basile, F.; Abdelkhalik, A.; Soler, S.; Valcárcel, J.V.; Martínez-Cuenca, M.-R. The nutritional quality potential of microgreens, baby leaves, and adult lettuce: An underexploited nutraceutical source. Foods 2022, 11, 423. [Google Scholar] [CrossRef]
- Zhang, Q.; Dai, W. Plant response to salinity stress. In Stress Physiology of Woody Plants; Dai, W., Ed.; CRC Press: Boca Raton, FL, USA, 2019; pp. 155–173. [Google Scholar]
- Baliyan, S.; Mukherjee, R.; Priyadarshini, A.; Vibhuti, A.; Gupta, A.; Pandey, R.P.; Chang, C. Determination of antioxidants by DPPH radical scavenging activity and quantitative phytochemical analysis of Ficus religiosa. Molecules 2022, 27, 1326. [Google Scholar] [CrossRef]
- Sánchez-Navarro, V.; Zornoza, R.; Faz, Á.; Fernández, J.A. Cowpea crop response to mineral and organic fertilization in SE Spain. Processes 2021, 9, 822. [Google Scholar] [CrossRef]
- Ye, L.; Zhao, X.; Bao, E.; Li, J.; Zou, Z.; Cao, K. Bio-organic fertilizer with reduced rates of chemical fertilization improves soil fertility and enhances tomato yield and quality. Sci. Rep. 2020, 10, 177. [Google Scholar] [CrossRef] [PubMed]
- Adnan, M.; Fahad, S.; Khan, I.A.; Saeed, M.; Ihsan, M.Z.; Saud, S.; Riaz, M.; Wang, D.; Wu, C. Integration of Poultry Manure and Phosphate Solubilizing Bacteria Improved Availability of Ca Bound P in Calcareous Soils. 3 Biotech 2019, 9, 368. [Google Scholar] [CrossRef]
- Mohamed, A.S.; Mohamed, M.H.M.; Halawa, S.S.; Saleh, S.A. Partial exchange of mineral N fertilizer for common bean plants by organic N fertilizer in the presence of salicylic acid as foliar application. In Gesunde Pflanzen; Springer: Berlin/Heidelberg, Germany, 2023. [Google Scholar] [CrossRef]
- Aykul, S.; Martinez-Hackert, E. Determination of half-maximal inhibitory concentration using biosensor-based protein interaction analysis. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef]
- Van Loon, W.A.; Linssen, J.P.; Legger, A.; Voragen, A.G. Anti-radical power gives insight into early lipid oxidation events during frying. J. Sci. Food Agric. 2006, 86, 1446–1451. [Google Scholar] [CrossRef]
- Youssef, M.A.; Farag, M.I.H. Co-application of organic manure and bio-fertilizer to improve soil fertility and production of quinoa and proceeding Jew’s Mallow crops. J. Soil Sci. Plant Nutr. 2021, 21, 2472–2488. [Google Scholar] [CrossRef]
- Yang, Y.; Syed, S.; Mao, S.; Li, Q.; Ge, F.; Lian, B.; Lu, C. Bioorganic–mineral fertilizer can remediate chemical fertilizer-oversupplied soil: Purslane planting as an example. J. Soil Sci. Plant Nutr. 2020, 20, 892–900. [Google Scholar] [CrossRef]
- Chen, S.; Gao, J.; Chen, H.; Zhang, Z.; Huang, J.; Lv, L.; Tan, J. The role of long-term mineral and manure fertilization on P species accumulation and phosphate-solubilizing microorganisms in paddy red soils. Soil 2023, 9, 101–116. [Google Scholar] [CrossRef]
- Adnan, M.; Fahad, S.; Zamin, M.; Shah, S.; Mian, I.A.; Danish, S.; Zafar-Ul-hye, M.; Battaglia, M.L.; Naz, R.M.M.; Saeed, B.; et al. Coupling Phosphate-Solubilizing Bacteria with Phosphorus Supplements Improve Maize Phosphorus Acquisition and Growth under Lime Induced Salinity Stress. Plants 2020, 9, 900. [Google Scholar] [CrossRef] [PubMed]
- Ponce, V.M.; Pandey, R.P.; Ercan, S. Characterization of drought across the climate spectrum. J. Hydrol. Eng. ASCE 2000, 5, 222–224. [Google Scholar] [CrossRef]
- Klute, A.; Dirksen, C. Hydraulic conductivity and diffusivity: Laboratory methods, in methods of soil analysis: Part 1-physical and mineralogical methods (Soil Science Society of America). Am. Soc. Agron. 1986, 9, 687–734. [Google Scholar]
- Page, A.I.; Miller, R.H.; Keeny, D.R. Methods of soil analysis, in Part II. In Chemical and Microbiological Methods, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 225–246. [Google Scholar]
- Pikovskaya, R. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Hu, X.; Chen, J.; Guo, J. Two phosphate and potassium-solubilizing bacteria isolated from Tianmu Mountain, Zhejiang, China. World J. Microbiol. Biotechnol. 2006, 22, 983–990. [Google Scholar] [CrossRef]
- Premono, M.E.; Moawad, A.; Vlek, P. Effect of phosphate-solubilizing Pseudomonas putida on the growth of maize and its survival in the rhizosphere. Indones. J. Crop Sci. 1996, 11, 13–23. [Google Scholar]
- Elias, F.; Woyessa, D.; Muleta, D. Phosphate solubilization potential of rhizosphere fungi isolated from plants in jimma zone, Southwest Ethiopia. Int. J. Microbiol. 2016, 2016, 5472601. [Google Scholar] [CrossRef]
- Prajapati, K.; Modi, H.A. The importance of potassium in plant growth-a review. Indian J. Plant Sci. 2012, 1, 177–186. [Google Scholar]
- Pitt, J.I.; Hocking, A.D. The ecology of fungal food spoilage. In Fungi and Food Spoilage; Springer: New York, NY, USA, 2009; pp. 3–9. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS Primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, S.; Lv, Y.; Zhai, H.; Cai, J.; Hu, Y. Linalool, the main volatile constituent from Zanthoxylum schinifolium pericarp, prevents growth of Aspergillus flavus in post-harvest grains. Food Control 2022, 137, 108967. [Google Scholar] [CrossRef]
- Idrovo-Novillo, J.; Gavilanes-Terán, I.; Angeles Bustamante, M.; Paredes, C. Composting as a method to recycle renewable plant resources back to the ornamental plant industry: Agronomic and economic assessment of composts. Process Saf. Environ. Prot. 2018, 116, 388–395. [Google Scholar] [CrossRef]
- Sosa-Zuniga, V.; Brito, V.; Fuentes, F.; Steinfort, U. Phenological growth stages of quinoa (Chenopodium quinoa) based on the BBCH scale. Ann. Appl. Biol. 2017, 171, 117–124. [Google Scholar] [CrossRef]
- Meier, U.; Bleiholder, H.; Buhr, L.; Feller, C.; Hack, H.; Heß, M.; Lancashire, P.D.; Schnock, U.; Stauß, R.; Van Den Boom, T. The BBCH system to coding the phenological growth stages of plants–history and publications. J. Kulturpflanzen 2009, 61, 41–52. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterisation by UV-VIS. In Current Protocols in Food Analytical Chemistry; Wiley: Madison, WI, USA, 2001; Volume 1, pp. F4.3.1–F4.3.8. [Google Scholar]
- Shibaeva, T.G.; Mamaev, A.V.; Sherudilo, E.G. Evaluation of a SPAD-502 plus chlorophyll meter to estimate chlorophyll content in leaves with interveinal chlorosis. Russ. J. Plant Physiol. 2020, 67, 690–696. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Fales, F.W. The assimilation and degradation of carbohydrates by yeast cells. J. Biol. Chem. 1951, 193, 113–124. [Google Scholar] [CrossRef]
- Schlegel, H.Q. Die verwertung organischer säuren durch Chlorella im licht. Planta 1956, 47, 510–526. [Google Scholar] [CrossRef]
- Ábrahám, E.; Hourton-Cabassa, C.; Erdei, L.; Szabados, L. Methods for determination of proline in plants. Methods Mol. Biol. 2010, 639, 317–331. [Google Scholar]
- Anderson, M.E. Determination of glutathione and glutathione disulfide in biological samples. Meth. Enzymol. 1985, 113, 548–555. [Google Scholar]
- Jagota, S.; Dani, H. A new colorimetric technique for the estimation of vitamin C using Folin phenol reagent. Anal. Biochem. 1982, 127, 178–182. [Google Scholar] [CrossRef]
- Sauvesty, A.; Page, F.; Huot, J. A simple method for extracting plant phenolic compounds. Can. J. For. Res. 1992, 22, 654–659. [Google Scholar] [CrossRef]
- Abe, N.; Murata, T.; Hirota, A. Novel 1,1-diphenyl-2-picryhy-drazyl-radical scavengers, bisorbicillin and demethyltrichodimerol, from a fungus. Biosci. Biotechnol. Biochem. 1998, 62, 661–666. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official method of analysis. In Association of Analytical Chemists, 19th ed.; AOAC: Washington, DC, USA, 2012; pp. 121–130. [Google Scholar]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall of India Pvt. Ltd.: New Delhi, India, 1967; pp. 144–197. [Google Scholar]
- Johnson, C.M.; Ulrich, A. Analytical methods for use in plant analysis. Calif. Agric. Exp. Stat Bull. 1959, 767, 25–78. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005; p. 1141. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar]
- Liyana-Pathirana, C.; Shahidi, F. Optimization of extraction of phenolic compounds from wheat using response surface methodology. Food Chem. 2005, 93, 47–56. [Google Scholar] [CrossRef]
- Nostro, A.; Guerrini, A.; Marino, A.; Tacchini, M.; Di Giulio, M.; Grandini, A.; Akin, M.; Cellini, L.; Bisignano, G.; Saraçoğlu, H.T. In vitro activity of plant extracts against biofilm-producing food-related bacteria. Int. J. Food Microbiol. 2016, 238, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, M.S. Properties of sufficiency and statistical tests. Proc. R. Soc. Ser. A 1937, 160, 268–282. [Google Scholar]
- Casella, G. Statistical Design, 1st ed.; Springer: Gainesville, FL, USA, 2008; pp. 32611–33545. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ST | PK+C-AN | Chl a | Chl b | Total Chl | Carotenoids | TPP |
|---|---|---|---|---|---|---|
| (mg cm−2) | ||||||
| Nor | PK100% | 27.0 ± 0.4 cd | 5.39 ± 0.27 a | 32.4 ± 0.4 e | 9.03 ± 0.16 ab | 41.4 ± 0.5 de |
| PK100%+C-AN | 34.3 ± 0.4 a | 7.64 ± 0.29 a | 41.9 ± 0.5 a | 9.48 ± 0.28 a | 51.4 ± 0.7 a | |
| PK75%+C-AN | 28.7 ± 0.6 c | 7.37 ± 0.16 a | 36.0 ± 0.7 bc | 8.57 ± 0.08 bc | 44.6 ± 0.7 bc | |
| PK50%+C-AN | 27.6 ± 0.5 c | 7.15 ± 0.19 a | 34.8 ± 0.6 cd | 8.41 ± 0.15 c | 43.2 ± 0.7 cd | |
| PK25%+C-AN | 27.0 ± 0.5 cd | 6.93 ± 0.11 a | 33.9 ± 0.6 de | 8.51 ± 0.17 c | 42.4 ± 0.5 d | |
| PK0%+C-AN | 22.5 ± 0.3 f | 6.01 ± 0.60 a | 28.5 ± 0.4 fg | 7.92 ± 0.13 d | 36.4 ± 0.3 fg | |
| Calc | PK100% | 23.6 ± 0.7 ef | 4.61 ± 0.36 a | 28.2 ± 0.4 g | 7.13 ± 0.38 fg | 35.3 ± 0.8 gh |
| PK100%+C-AN | 30.6 ± 1.2 b | 6.80 ± 0.31 a | 37.4 ± 1.0 b | 9.01 ± 0.17 ab | 46.5 ± 1.1 b | |
| PK75%+C-AN | 28.7 ± 0.8 c | 5.74 ± 0.21 a | 34.4 ± 0.8 cd | 8.44 ± 0.14 c | 42.9 ± 0.8 cd | |
| PK50%+C-AN | 27.0 ± 0.8 cd | 5.25 ± 0.13 a | 32.2 ± 0.8 e | 7.64 ± 0.11 de | 39.9 ± 0.8 e | |
| PK25%+C-AN | 25.2 ± 0.5 de | 4.96 ± 0.11 a | 30.2 ± 0.5 f | 7.36 ± 0.16 ef | 37.6 ± 0.4 f | |
| PK0%+C-AN | 23.0 ± 0.3 f | 4.56 ± 0.14 a | 27.5 ± 0.4 g | 6.67 ± 0.17 g | 34.2 ± 0.3 h | |
| p-value for ST × PK+C-AN | 0.007 ** | 0.073 ns | 0.047 * | <0.001 ** | 0.016 * | |
| ST | PK+C-AN | Crude Protein | Ash | Crude Lipid | Crude Fiber | Carbohydrates |
|---|---|---|---|---|---|---|
| (%) | ||||||
| Nor | PK100% | 14.7 ± 0.10 f | 2.61 ± 0.03 e | 6.35 ± 0.15 a–c | 3.57 ± 0.04 f | 64.5 ± 0.44 f |
| PK100%+C-AN | 16.1 ± 0.12 a | 3.20 ± 0.07 c | 6.48 ± 0.15 ab | 3.61 ± 0.04 e | 62.4 ± 0.52 i | |
| PK75%+C-AN | 15.2 ± 0.11 d | 2.91 ± 0.06 d | 6.55 ± 0.15 ab | 3.70 ± 0.04 d | 63.9 ± 0.46 g | |
| PK50%+C-AN | 15.4 ± 0.11 c | 2.97 ± 0.04 d | 6.45 ± 0.22 ab | 3.82 ± 0.04 c | 63.1 ± 0.47 h | |
| PK25%+C-AN | 15.6 ± 0.11 b | 2.41 ± 0.03 f | 6.59 ± 0.25 a | 4.00 ± 0.04 b | 63.3 ± 0.47 h | |
| PK0%+C-AN | 14.9 ± 0.10 e | 1.48 ± 0.10 h | 6.13 ± 0.15 c–e | 4.13 ± 0.05 a | 65.2 ± 0.43 e | |
| Calc | PK100% | 11.9 ± 0.08 k | 2.00 ± 0.03 g | 5.90 ± 0.14 e | 3.24 ± 0.04 l | 71.1 ± 0.35 a |
| PK100%+C-AN | 12.4 ± 0.13 j | 3.67 ± 0.04 a | 6.03 ± 0.14 de | 3.27 ± 0.04 k | 68.5 ± 0.40 b | |
| PK75%+C-AN | 13.3 ± 0.11 i | 3.34 ± 0.04 b | 6.03 ± 0.14 de | 3.36 ± 0.04 j | 67.7 ± 0.39 cd | |
| PK50%+C-AN | 13.4 ± 0.09 i | 2.94 ± 0.03 d | 5.87 ± 0.15 e | 3.40 ± 0.04 i | 68.1 ± 0.37 bc | |
| PK25%+C-AN | 13.6 ± 0.10 h | 2.60 ± 0.06 e | 6.15 ± 0.15 c–e | 3.74 ± 0.04 h | 68.3 ± 0.37 b | |
| PK0%+C-AN | 14.0 ± 0.10 g | 2.57 ± 0.05 e | 6.28 ± 0.15 b–d | 3.52 ± 0.04 g | 67.6 ± 0.39 d | |
| p-value for ST × PK+C-AN | <0.001 ** | <0.001 ** | 0.003 ** | <0.001 ** | <0.001 ** | |
| ST | PK+C-AN | TPC | TFC | IC50 (mg mL−1) | ARP |
|---|---|---|---|---|---|
| (mg 100−1 g Dry Seed) | |||||
| Nor | PK100% | 66.0 ± 0.53 f | 26.2 ± 0.38 f | 0.43 ± 0.00 e | 2.08 ± 0.02 g |
| PK100%+C-AN | 84.0 ± 0.56 a | 44.5 ± 0.41 a | 0.54 ± 0.01 a | 2.78 ± 0.05 c | |
| PK75%+C-AN | 74.9 ± 0.50 c | 39.6 ± 0.41 b | 0.50 ± 0.01 b | 2.40 ± 0.04 d | |
| PK50%+C-AN | 75.4 ± 0.52 c | 33.7 ± 0.41 d | 0.47 ± 0.01 c | 2.42 ± 0.03 d | |
| PK25%+C-AN | 69.9 ± 0.52 d | 30.4 ± 0.32 e | 0.43 ± 0.01 e | 2.21 ± 0.03 ef | |
| PK0%+C-AN | 53.6 ± 0.36 i | 30.0 ± 0.28 e | 0.37 ± 0.01 f | 1.91 ± 0.03 h | |
| Calc | PK100% | 57.1 ± 0.43 h | 23.6 ± 0.55 g | 0.35 ± 0.00 g | 2.28 ± 0.02 e |
| PK100%+C-AN | 77.7 ± 0.52 b | 43.6 ± 0.44 a | 0.48 ± 0.01 c | 3.21 ± 0.03 a | |
| PK75%+C-AN | 68.3 ± 1.24 e | 36.0 ± 1.12 c | 0.45 ± 0.00 d | 2.94 ± 0.03 b | |
| PK50%+C-AN | 69.2 ± 0.72 de | 30.0 ± 0.27 e | 0.43 ± 0.00 e | 2.75 ± 0.03 c | |
| PK25%+C-AN | 63.6 ± 0.43 g | 25.1 ± 0.30 f | 0.38 ± 0.00 f | 2.42 ± 0.03 d | |
| PK0%+C-AN | 51.5 ± 0.35 j | 21.9 ± 0.27 h | 0.32 ± 0.00 h | 2.16 ± 0.02 f | |
| p-value for ST × PK+C-AN | <0.001 ** | <0.001 ** | 0.005 ** | <0.001 ** | |
| ST | PK+C-AN | P | K+ | Ca2+ | Mg2+ | Na+ | Fe2+ | Zn2+ |
|---|---|---|---|---|---|---|---|---|
| (g 100−1 g Dry Seed) | ||||||||
| Nor | PK100% | 0.35 ± 0.00 ef | 2.52 ± 0.02 g | 1.63 ± 0.02 a | 0.84 ± 0.05 e | 0.30 ± 0.01 c | 0.92 ± 0.02 h | 0.46 ± 0.01 e |
| PK100%+C-AN | 0.48 ± 0.01 a | 3.19 ± 0.11 a | 1.95 ± 0.04 a | 1.42 ± 0.12 a | 0.37 ± 0.02 a | 1.15 ± 0.08 a | 0.50 ± 0.01 a | |
| PK75%+C-AN | 0.44 ± 0.01 b | 2.70 ± 0.03 c | 1.74 ± 0.02 a | 1.08 ± 0.12 b | 0.34 ± 0.01 b | 1.01 ± 0.02 c | 0.49 ± 0.01 b | |
| PK50%+C-AN | 0.42 ± 0.00 c | 2.64 ± 0.02 e | 1.69 ± 0.02 a | 0.87 ± 0.07 c | 0.30 ± 0.00 c | 0.95 ± 0.00 f | 0.47 ± 0.01 c | |
| PK25%+C-AN | 0.33 ± 0.00 f | 2.59 ± 0.00 f | 1.49 ± 0.03 a | 0.84 ± 0.07 de | 0.29 ± 0.00 d | 0.89 ± 0.01 i | 0.47 ± 0.01 c | |
| PK0%+C-AN | 0.27 ± 0.01 h | 2.30 ± 0.04 i | 1.23 ± 0.02 a | 0.63 ± 0.01 k | 0.25 ± 0.01 g | 0.86 ± 0.01 j | 0.43 ± 0.00 g | |
| Calc | PK100% | 0.36 ± 0.01 e | 2.21 ± 0.02 k | 1.42 ± 0.02 a | 0.68 ± 0.04 i | 0.21 ± 0.02 j | 0.82 ± 0.01 k | 0.45 ± 0.00 f |
| PK100%+C-AN | 0.47 ± 0.00 a | 2.89 ± 0.01 b | 1.72 ± 0.03 a | 0.85 ± 0.04 d | 0.28 ± 0.01 e | 1.08 ± 0.04 b | 0.47 ± 0.00 c | |
| PK75%+C-AN | 0.42 ± 0.01 c | 2.69 ± 0.08 d | 1.55 ± 0.02 a | 0.80 ± 0.05 f | 0.26 ± 0.00 f | 0.98 ± 0.03 d | 0.46 ± 0.00 d | |
| PK50%+C-AN | 0.40 ± 0.01 d | 2.47 ± 0.03 h | 1.52 ± 0.02 a | 0.75 ± 0.02 g | 0.24 ± 0.01 h | 0.96 ± 0.01 e | 0.45 ± 0.00 ef | |
| PK25%+C-AN | 0.30 ± 0.01 g | 2.22 ± 0.03 j | 1.38 ± 0.02 a | 0.70 ± 0.02 h | 0.22 ± 0.01 i | 0.94 ± 0.01 g | 0.46 ± 0.01 e | |
| PK0%+C-AN | 0.23 ± 0.01 i | 2.20 ± 0.00 k | 1.03 ± 0.02 a | 0.66 ± 0.02 j | 0.16 ± 0.03 k | 0.82 ± 0.00 k | 0.44 ± 0.01 g | |
| p-value for ST × PK+C-AN | <0.001 ** | <0.001 ** | 0.220 ns | <0.001 ** | <0.001 ** | <0.001 ** | <0.001 ** | |
| ST | PK+C-AN | Plant Height (cm) | Plant Dry Weight (g) | HW (kg hL−1) | Seed Yield | Biological Yield | HI (%) |
|---|---|---|---|---|---|---|---|
| (t ha−1) | |||||||
| Nor | PK100% | 96.8 ± 1.3 d | 26.5 ± 0.2 c | 67.6 ± 0.4 fg | 2.24 ± 0.04 e | 4.95 ± 0.24 d | 45.7 ± 1.9 a |
| PK100%+C-AN | 112.5 ± 1.3 a | 39.3 ± 0.4 a | 70.1 ± 0.5 b | 3.27 ± 0.05 a | 7.70 ± 0.20 a | 42.6 ± 1.8 a | |
| PK75%+C-AN | 109.8 ± 1.0 b | 29.9 ± 0.8 b | 69.3 ± 0.5 b–d | 3.20 ± 0.04 a | 6.96 ± 0.26 b | 46.1 ± 1.4 a | |
| PK50% + C-AN | 108.8 ± 0.7 b | 26.5 ± 0.2 c | 68.5 ± 0.5 d–f | 3.01 ± 0.06 b | 6.64 ± 0.43 b | 46.3 ± 3.2 a | |
| PK25%+C-AN | 107.3 ± 0.8 bc | 24.2 ± 0.4 d | 68.2 ± 0.4 e–g | 2.01 ± 0.09 f | 5.42 ± 0.25 c | 37.4 ± 1.8 a | |
| PK0%+C-AN | 104.8 ± 1.0 c | 23.7 ± 0.5 d | 66.5 ± 0.5 h | 1.76 ± 0.05 g | 4.21 ± 0.27 ef | 42.5 ± 1.9 a | |
| Calc | PK100% | 80.3 ± 1.0 g | 13.0 ± 0.4 i | 67.6 ± 0.6f g | 1.57 ± 0.04 h | 3.01 ± 0.04 g | 52.3 ± 1.5 a |
| PK100%+C-AN | 93.0 ± 1.4 e | 21.2 ± 0.5 e | 72.4 ± 1.03 a | 2.50 ± 0.07 c | 4.79 ± 0.04 d | 52.2 ± 1.0 a | |
| PK75%+C-AN | 91.8 ± 1.3 e | 19.0 ± 0.2 f | 69.4 ± 0.1 bc | 2.43 ± 0.07 cd | 4.56 ± 0.10 de | 53.3 ± 0.8 a | |
| PK50%+C-AN | 90.8 ± 1.5 ef | 17.2 ± 0.4 g | 69.0 ± 0.5 c–e | 2.27 ± 0.07 de | 4.25 ± 0.11 ef | 53.7 ± 1.8 a | |
| PK25%+C-AN | 88.3 ± 1.8 f | 16.2 ± 0.3 h | 68.4 ± 0.4 d–f | 2.12 ± 0.15 ef | 4.02 ± 0.19 f | 52.4 ± 1.3 a | |
| PK0%+C-AN | 82.2 ± 1.0 g | 12.4 ± 0.2 i | 67.4 ± 0.2 g | 1.63 ± 0.09 gh | 3.18 ± 0.10 g | 51.3 ± 2.8 a | |
| p-value for ST × PK+C-AN | 0.034 * | <0.001 ** | 0.002 ** | <0.001 ** | <0.001 ** | 0.077 ns | |
| Property | Unite | Soil Type | |
|---|---|---|---|
| Normal | Calcareous | ||
| Sand | (%) | 79.90 ± 0.72 | 79.00 ± 0.91 |
| Silt | 9.60 ± 0.51 | 10.10 ± 0.75 | |
| Clay | 10.50 ± 0.46 | 10.90 ± 0.42 | |
| Texture class | S.L | S.L | |
| Dry bulk density | (g cm−3) | 1.65 ± 0.06 | 1.59 ± 0.04 |
| pH (in 1:2.5 soil: water (w:v) suspension) | 7.52 ± 0.03 | 7.69 ± 0.05 | |
| ECe (in soil past (1:2.5, w:v) extract) | (dS m−1) | 3.56 ± 0.05 | 3.19 ± 0.05 |
| Cation exchange capacity | (meq/100 g soil) | 9.35 ± 0.32 | 9.89 ± 0.45 |
| CaCO3 | (%) | 6.58 ± 0.07 | 15.54 ± 0.09 |
| Organic carbon | 47.09 ± 0.95 | 43.60 ± 0.81 | |
| Organic matter | 0.81 ± 0.02 | 0.75 ± 0.03 | |
| ** Exchangeable cations | |||
| Ca2+ | (meq L−1) | 12.89 ± 0.24 | 17.66 ± 0.33 |
| Mg2+ | 9.54 ± 0.36 | 10.59 ± 0.18 | |
| Na+ | 11.25 ± 0.18 | 12.75 ± 0.26 | |
| K+ | 0.78 ± 0.01 | 1.00 ± 0.02 | |
| ** Exchangeable anions | |||
| Cl− | (meq L−1) | 8.17 ± 0.62 | 14.21 ± 0.56 |
| SO42− | 25.07 ± 0.74 | 21.54 ± 0.66 | |
| HCO3− | 1.22 ± 0.03 | 6.25 ± 0.08 | |
| CO32− | - | - | |
| Available soil nutrients | |||
| N | (%) | 0.08 ± 0.01 | 0.04 ± 0.01 |
| P | (mg kg−1 soil) | 7.54 ± 0.10 | 5.44 ± 0.12 |
| K+ | 50.00 ± 1.4 | 42.08 ± 1.2 | |
| Fe2+ | 2.20 ± 0.05 | 2.00 ± 0.06 | |
| Mg2+ | 4.10 ± 0.22 | 3.90 ± 0.18 | |
| Zn2+ | 0.79 ± 0.03 | 0.70 ± 0.05 | |
| Cu2+ | 0.51 ± 0.05 | 0.44 ± 0.06 | |
| Total microbial count | |||
| PSMs | (cfu × 103 g−1 soil) | 0.00 | 0.00 |
| Azotobacter sp. | 2.00 ± 0.46 | 0.00 | |
| Parameter | Unite | Mean Value ± SE |
|---|---|---|
| Bulk density | (g cm−3) | 0.75 ± 0.09 |
| Moisture content | (%) | 38.40 ± 0.93 |
| Water holding pores | 25.50 ± 0.65 | |
| pH (in 1:2.5 compost: water (w:v) suspension) | 7.64 ± 0.05 | |
| ECe | (dS m−1) | 2.10 ± 0.05 |
| CaCO3 | (%) | 1.65 ± 0.03 |
| Organic carbon | 27.06 ± 0.65 | |
| Organic matter | 46.55 ± 0.70 | |
| Carbon/nitrogen ratio | 20.81 ± 0.35 | |
| Total macro-and micronutrients | ||
| N | (%) | 1.30 ± 0.03 |
| P | 0.95 ± 0.02 | |
| K+ | 0.84 ± 0.01 | |
| Fe+2 | (mg kg−1) | 0.979 ± 18 |
| Mg2+ | 0.469 ± 26 | |
| Zn2+ | 0.659 ± 33 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youssef, S.M.; Shaaban, A.; Abdelkhalik, A.; Abd El Tawwab, A.R.; Abd Al Halim, L.R.; Rabee, L.A.; Alwutayd, K.M.; Ahmed, R.M.M.; Alwutayd, R.; Hemida, K.A. Compost and Phosphorus/Potassium-Solubilizing Fungus Effectively Boosted Quinoa’s Physio-Biochemical Traits, Nutrient Acquisition, Soil Microbial Community, and Yield and Quality in Normal and Calcareous Soils. Plants 2023, 12, 3071. https://doi.org/10.3390/plants12173071
Youssef SM, Shaaban A, Abdelkhalik A, Abd El Tawwab AR, Abd Al Halim LR, Rabee LA, Alwutayd KM, Ahmed RMM, Alwutayd R, Hemida KA. Compost and Phosphorus/Potassium-Solubilizing Fungus Effectively Boosted Quinoa’s Physio-Biochemical Traits, Nutrient Acquisition, Soil Microbial Community, and Yield and Quality in Normal and Calcareous Soils. Plants. 2023; 12(17):3071. https://doi.org/10.3390/plants12173071
Chicago/Turabian StyleYoussef, Samah M., Ahmed Shaaban, Abdelsattar Abdelkhalik, Ahmed R. Abd El Tawwab, Laila R. Abd Al Halim, Laila A. Rabee, Khairiah Mubarak Alwutayd, Reda M. M. Ahmed, Rahaf Alwutayd, and Khaulood A. Hemida. 2023. "Compost and Phosphorus/Potassium-Solubilizing Fungus Effectively Boosted Quinoa’s Physio-Biochemical Traits, Nutrient Acquisition, Soil Microbial Community, and Yield and Quality in Normal and Calcareous Soils" Plants 12, no. 17: 3071. https://doi.org/10.3390/plants12173071
APA StyleYoussef, S. M., Shaaban, A., Abdelkhalik, A., Abd El Tawwab, A. R., Abd Al Halim, L. R., Rabee, L. A., Alwutayd, K. M., Ahmed, R. M. M., Alwutayd, R., & Hemida, K. A. (2023). Compost and Phosphorus/Potassium-Solubilizing Fungus Effectively Boosted Quinoa’s Physio-Biochemical Traits, Nutrient Acquisition, Soil Microbial Community, and Yield and Quality in Normal and Calcareous Soils. Plants, 12(17), 3071. https://doi.org/10.3390/plants12173071