
Application of Allele Specific PCR in Identifying Offspring Genotypes of Bi-Allelic SbeIIb Mutant Lines in Rice


Abstract
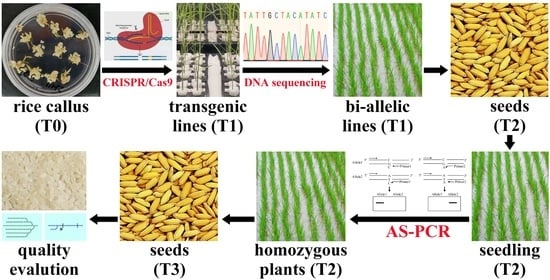
:
1. Introduction
2. Results
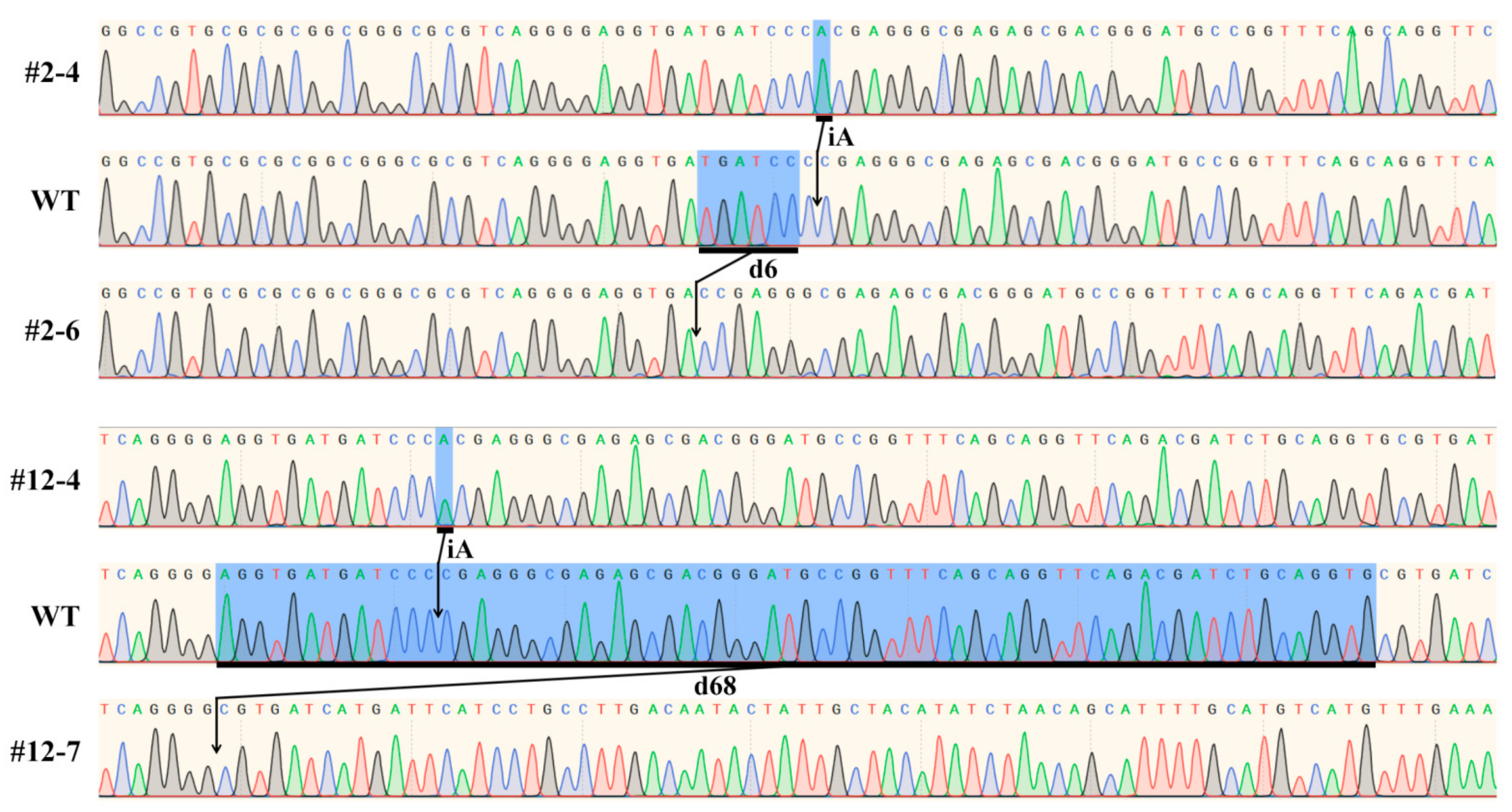
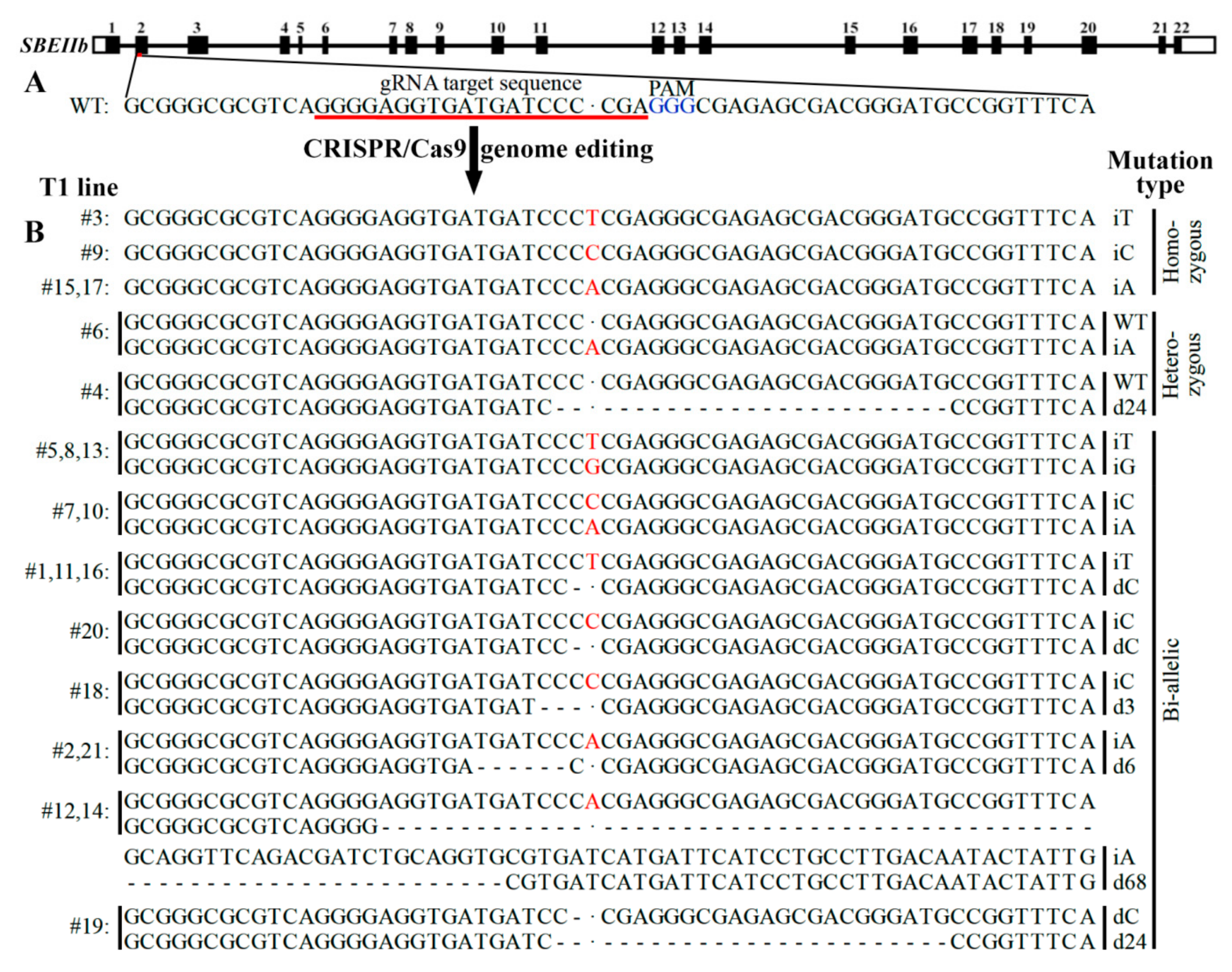
2.1. CRISPR/Cas9 Induced Mutation in SBEIIb Gene and Mutation Types of T1 Lines
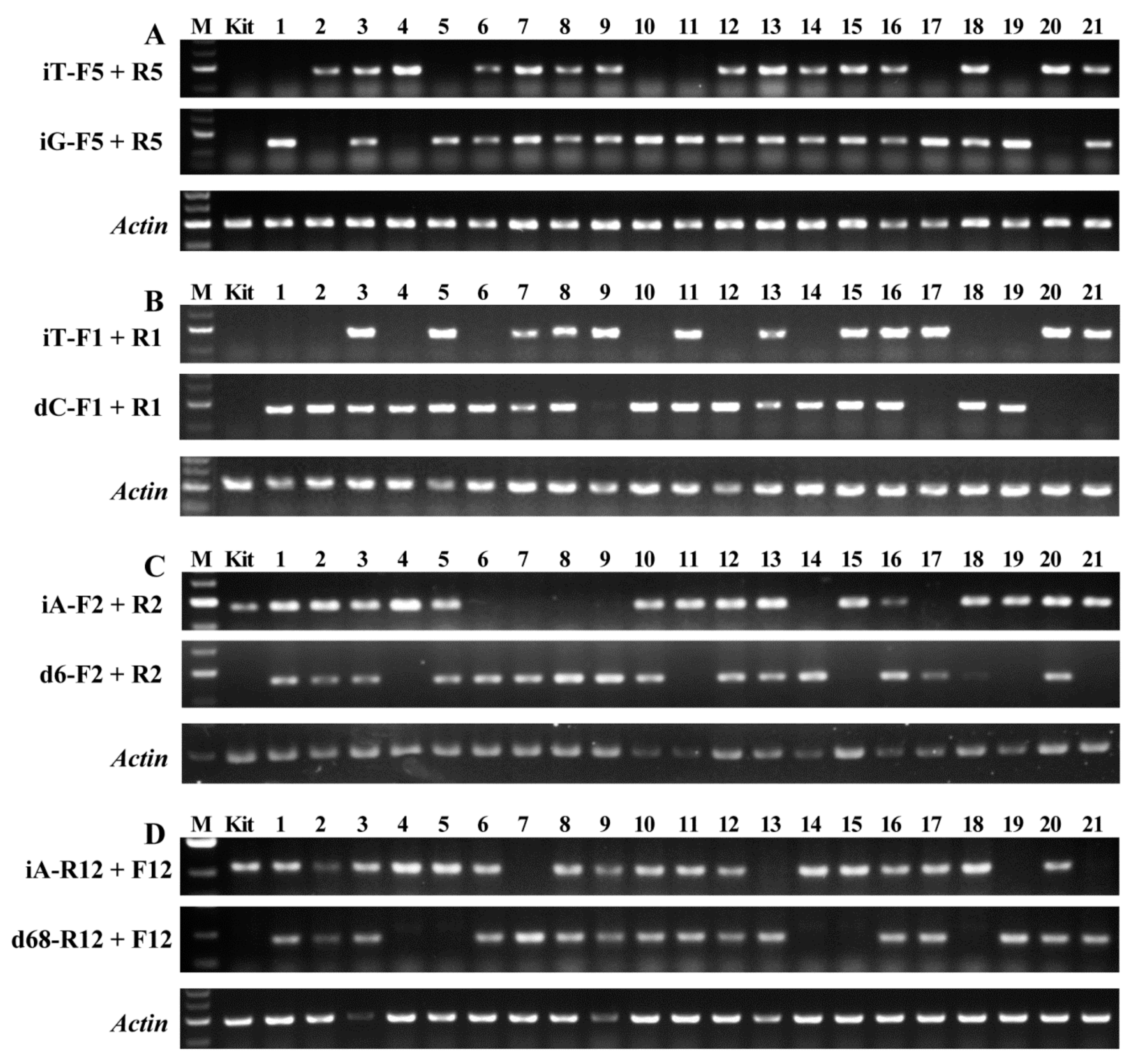
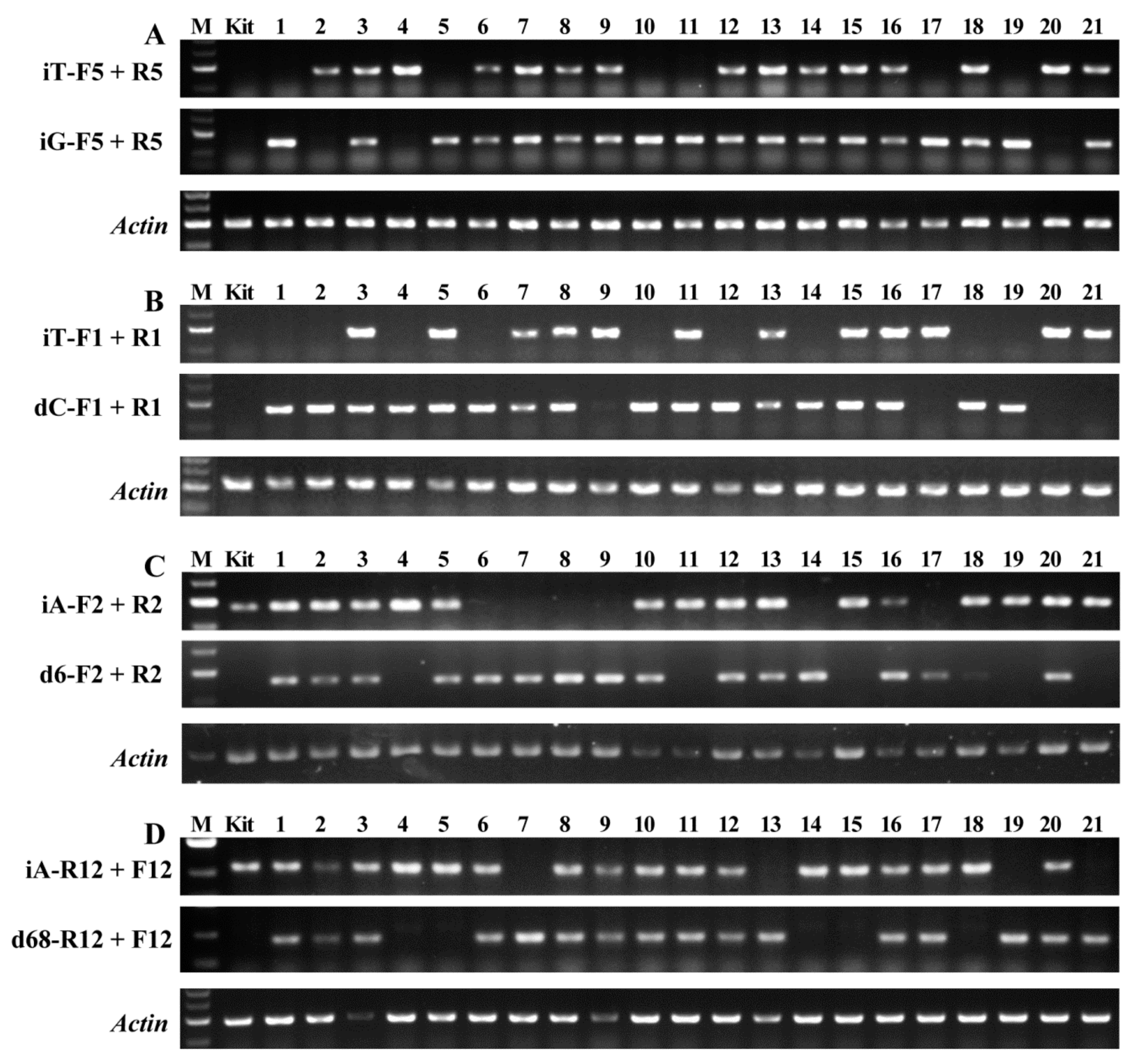
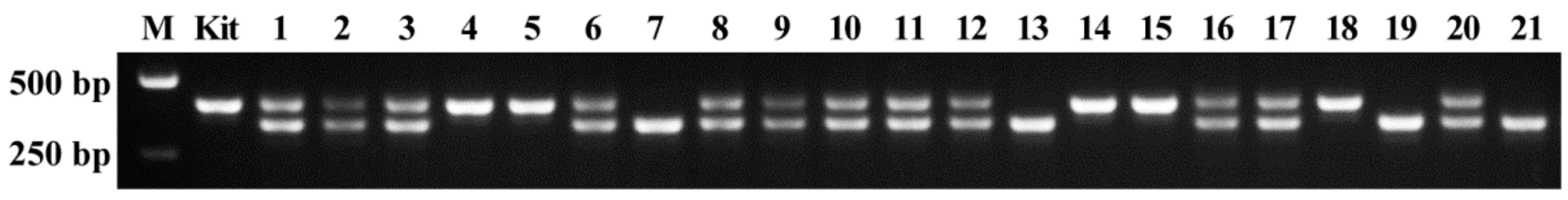
2.2. Detecting Genotypes of Offspring Segregation Plants from Bi-Allelic Mutant Lines
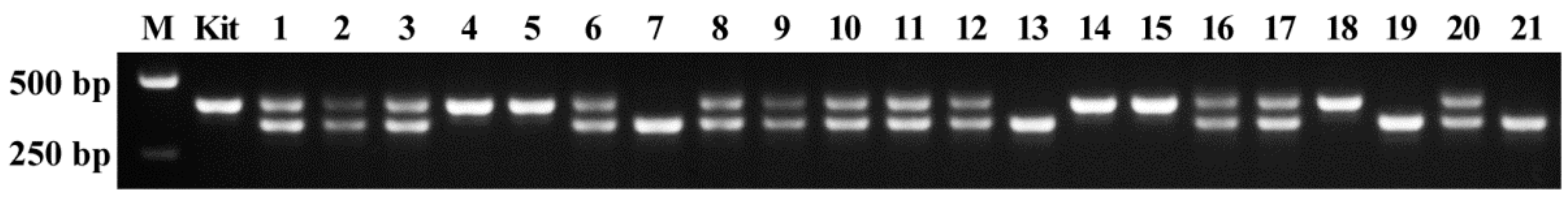
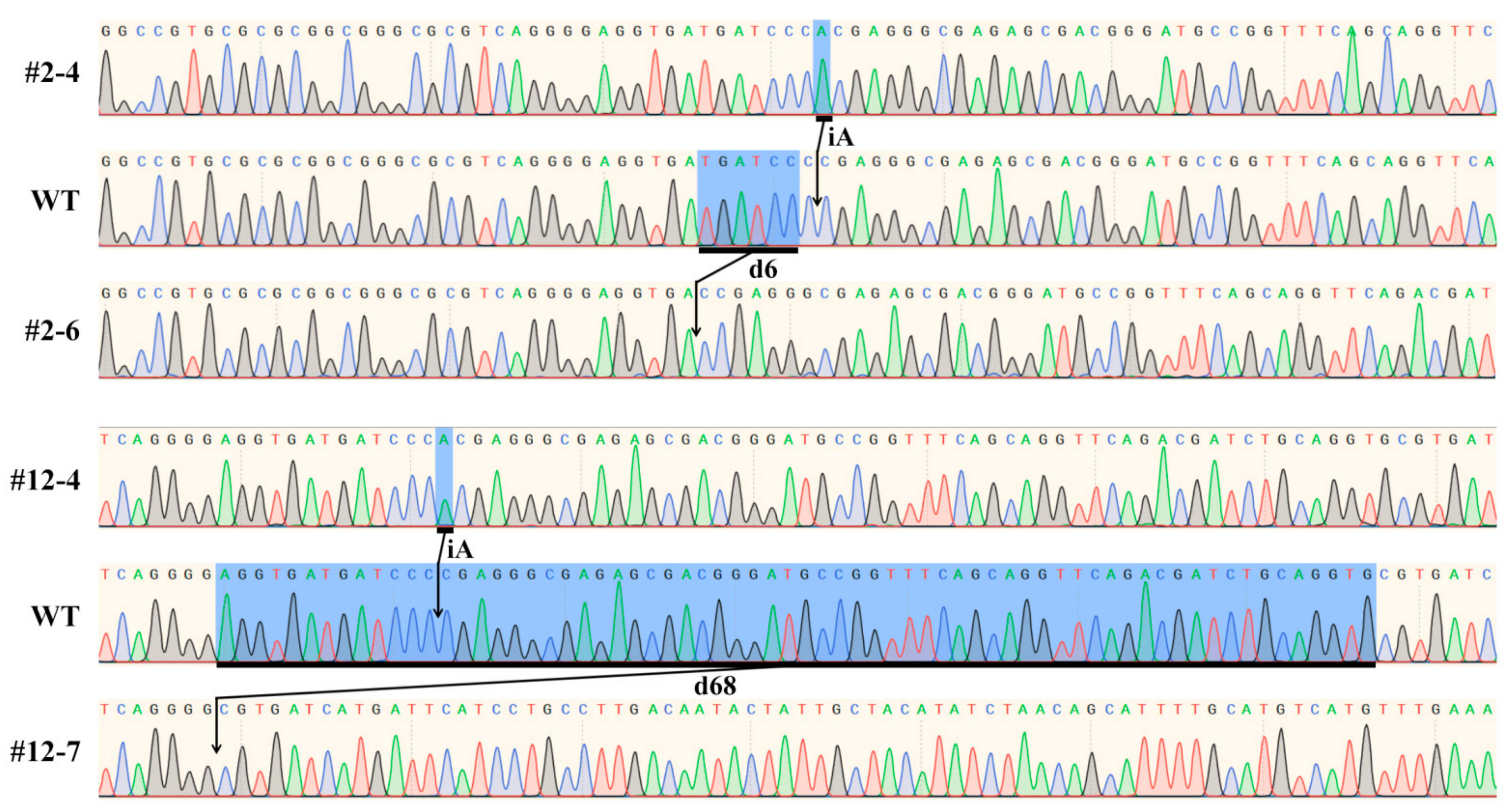
2.3. Accuracy of Genotype Detected by AS-PCR
3. Discussion
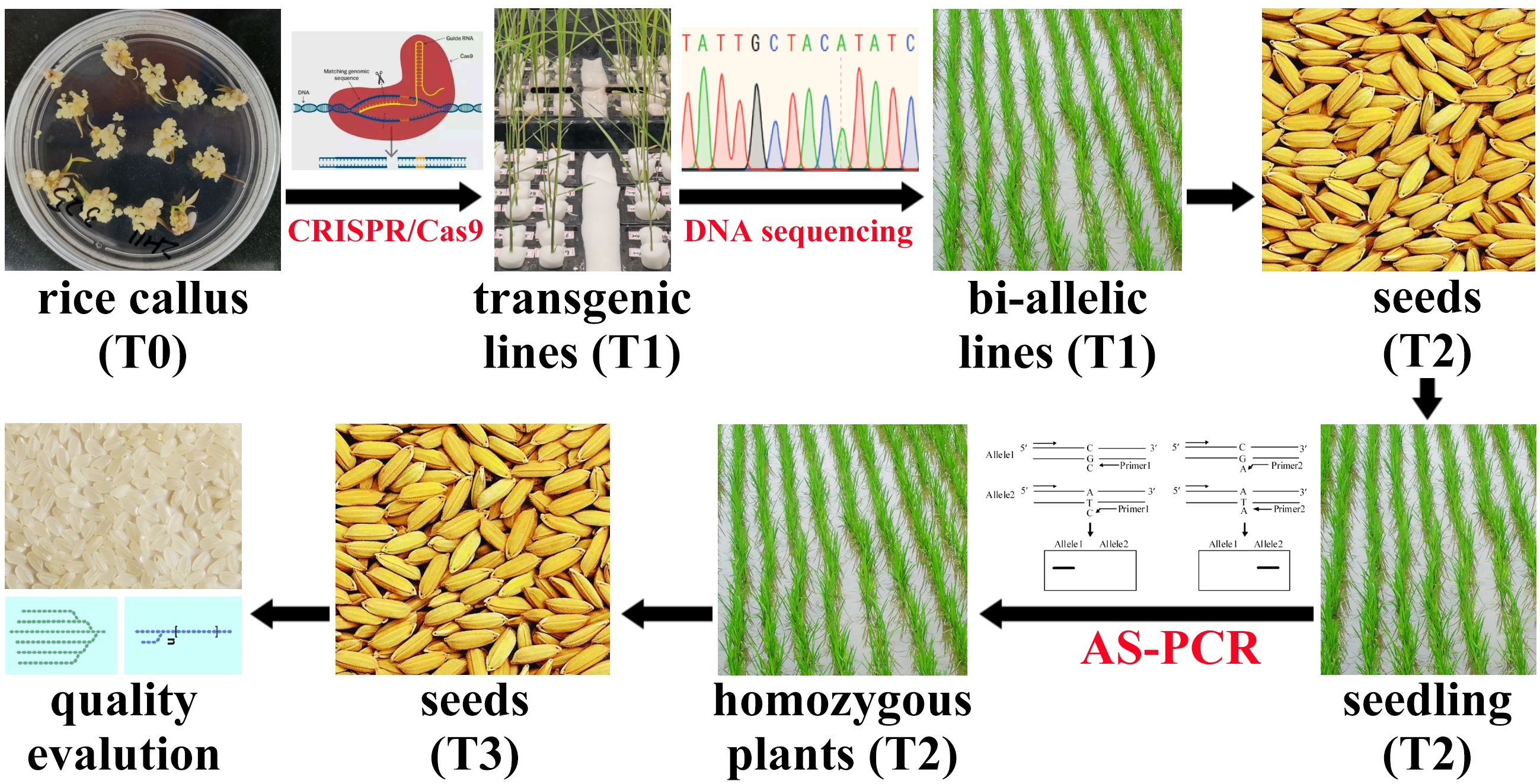
4. Materials and Methods
4.1. Plant Materials
4.2. Leaf DNA Extraction
4.3. Routine PCR and DNA Sequencing
4.4. Allele-Specific Primer Design
4.5. AS-PCR and Genotype Detection
4.6. Detection of Genotype with Large Fragment Deletion
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Aklilu, E. Review on forward and reverse genetics in plant breeding. All Life 2021, 14, 127–135. [Google Scholar] [CrossRef]
- Bahuguna, R.N.; Gupta, P.; Bagri, J.; Singh, D.; Dewi, A.K.; Tao, L.; Islam, M.; Sarsu, F.; Singla-Pareek, S.L.; Pareek, A. Forward and reverse genetics approaches for combined stress tolerance in rice. Ind. J. Plant Physiol. 2018, 23, 630–646. [Google Scholar] [CrossRef]
- Kumar, A.; Jain, S. Forward genetics using radiation hybrids (deletion mutants) in plants. Ind. J. Plant Physiol. 2018, 23, 622–629. [Google Scholar] [CrossRef]
- Forster, B.P.; Shu, Q.Y. Plant Mutagenesis in Crop Improvement: Basic Terms and Applications. In Plant Mutation Breeding and Biotechnology; Shu, Q.Y., Forster, B.P., Nakagawa, H., Eds.; CABI eBook: Wallingford, UK, 2012; pp. 9–20. [Google Scholar] [CrossRef]
- Sahu, P.K.; Sao, R.; Mondal, S.; Vishwakarma, G.; Gupta, S.K.; Kumar, V.; Singh, S.; Sharma, D.; Das, B.K. Next generation sequencing based forward genetic approaches for identification and mapping of causal mutations in crop plants: A comprehensive review. Plants 2020, 9, 1355. [Google Scholar] [CrossRef]
- Karlson, C.K.S.; Mohd-Noor, S.N.; Nolte, N.; Tan, B.C. CRISPR/dCas9-based systems: Mechanisms and applications in plant sciences. Plants 2021, 10, 2055. [Google Scholar] [CrossRef]
- Liu, G.; Lin, Q.; Jin, S.; Gao, C. The CRISPR-Cas toolbox and gene editing technologies. Mol. Cell 2022, 82, 333–347. [Google Scholar] [CrossRef]
- Rasheed, A.; Gill, R.A.; Hassan, M.U.; Mahmood, A.; Qari, S.; Zaman, Q.U.; Ilyas, M.; Aamer, M.; Batool, M.; Li, H.; et al. A critical review: Recent advancements in the use of CRISPR/Cas9 technology to enhance crops and alleviate global food crises. Curr. Issues Mol. Biol. 2021, 43, 1950–1976. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.; Magar, N.D.; Barbadikar, K.M. Current technological interventions and applications of CRISPR/Cas for crop improvement. Mol. Biol. Rep. 2022. [Google Scholar] [CrossRef] [PubMed]
- Singha, D.L.; Das, D.; Sarki, Y.N.; Chowdhury, N.; Sharma, M.; Maharana, J.; Chikkaputtaiah, C. Harnessing tissue-specific genome editing in plants through CRISPR/Cas system: Current state and future prospects. Planta 2022, 255, 28. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Teng, K.C.; Nawaz, G.; Feng, X.; Usman, B.; Wang, X.; Luo, L.; Zhao, N.; Liu, Y.G.; Li, R.B. Generation of semi-dwarf rice (Oryza sativa L.) lines by CRISPR/Cas9-directed mutagenesis of OsGA20ox2 and proteomic analysis of unveiled changes caused by mutations. 3 Biotech 2019, 9, 387. [Google Scholar] [CrossRef]
- Nawaz, G.; Han, Y.; Usman, B.; Liu, F.; Qin, B.X.; Li, R.B. Knockout of OsPRP1, a gene encoding proline-rich protein, confers enhanced cold sensitivity in rice (Oryza sativa L.) at the seedling stage. 3 Biotech 2019, 9, 254. [Google Scholar] [CrossRef]
- Sun, Y.; Jiao, G.; Liu, Z.; Zhang, X.; Li, J.; Guo, X.; Du, W.; Du, J.; Francis, F.; Zhao, Y.; et al. Generation of high-amylose rice through CRISPR/Cas9-mediated targeted mutagenesis of starch branching enzymes. Front. Plant Sci. 2017, 8, 298. [Google Scholar] [CrossRef]
- Usman, B.; Nawaz, G.; Zhao, N.; Liao, S.Y.; Liu, Y.G.; Li, R.B. Precise editing of the OsPYL9 gene by RNA-guided Cas9 nuclease confers enhanced drought tolerance and grain yield in rice (Oryza sativa L.) by regulating circadian rhythm and abiotic stress responsive proteins. Int. J. Mol. Sci. 2020, 21, 7854. [Google Scholar] [CrossRef]
- Wang, F.Q.; Xu, Y.; Li, W.Q.; Chen, Z.H.; Wang, J.; Fan, F.J.; Tao, Y.J.; Jiang, Y.J.; Zhu, Q.H.; Yang, J. Creating a novel herbicide-tolerance OsALS allele using CRISPR/Cas9-mediated gene editing. The Crop J. 2021, 9, 305–312. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Q.; Wang, Y.; Huang, J.; Luo, N.J.; Wei, S.B.; Jin, J. Analysing the rice young panicle transcriptome reveals the gene regulatory network controlled by TRIANGULAR HULL1. Rice 2019, 12, 6. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.Y.; Niu, B.X.; Long, Y.M.; Li, G.S.; Tang, J.T.; Zhang, Y.L.; Ren, D.; Liu, Y.G.; Chen, L.T. Suppression or knockout of SaF/SaM overcomes the Sa-mediated hybrid male sterility in rice. J. Integr. Plant Biol. 2017, 59, 669–679. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Huang, S.M.; Sun, M.Y.; Liu, S.Y.; Liu, Y.M.; Wang, W.X.; Zhang, X.R.; Wang, H.Z.; Hua, W. An improved allele-specific PCR primer design method for SNP marker analysis and its application. Plant Methods 2012, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Soleimani, V.D.; Baum, B.R.; Johnson, D.A. Efficient validation of single nucleotide polymorphisms in plants by allele-specific PCR, with an example from barley. Plant Mol. Biol. Rep. 2003, 21, 281–288. [Google Scholar] [CrossRef]
- Yang, J.; Wang, J.; Fan, F.J.; Zhu, J.Y.; Chen, T.; Wang, C.L.; Zheng, T.Q.; Zhang, J.; Zhong, W.G.; Xu, J.L. Development of AS-PCR marker based on a key mutation confirmed by resequencing of Wx-mp in milky princess and its application in japonica soft rice (Oryza sativa L.) breeding. Plant Breed. 2013, 132, 595–603. [Google Scholar] [CrossRef]
- Chen, H.; Shan, J.; Yang, K.; Wang, Y.Y.; Lu, C.M. Abundant variation of Waxy gene in Yunnan rice landraces and molecular characterization of a novel Wxzm allele. Crop Sci. 2014, 54, 2152–2159. [Google Scholar] [CrossRef]
- Chen, Z.; Lu, Y.; Feng, L.; Hao, W.; Li, C.; Yang, Y.; Fan, X.; Li, Q.; Zhang, C.; Liu, Q. Genetic dissection and functional differentiation of ALKa and ALKb, two natural alleles of the ALK/SSIIa gene, responding to low gelatinization temperature in rice. Rice 2020, 13, 39. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Little, S. Amplification-refractory mutation system (ARMS) analysis of point mutations. Curr. Protoc. Hum. Genet. 1995, 7, 9.8.1–9.8.12. [Google Scholar] [CrossRef]
- Cha, R.S.; Zarbl, H.; Keohavong, P.; Thilly, W.G. Mismatch amplification mutation assay (MAMA): Application to the c-H-ras gene. Genome Res. 1992, 2, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Newton, C.R.; Graham, A.; Heptinstall, L.E.; Powell, S.J.; Summers, C.; Kalsheker, N.; Smith, J.C.; Markham, A.F. Analysis of any point mutation in DNA. The amplification refractory mutation system (ARMS). Nucleic Acids Res. 1989, 17, 2503–2516. [Google Scholar] [CrossRef]
- Ye, S.; Dhillon, S.; Ke, X.; Collins, A.R.; Day, I.N.M. An efficient procedure for genotyping single nucleotide polymorphisms. Nucleic Acids Res. 2001, 29, e88. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.M.; Arnheim, N.; Goodman, M.F. Extension of base mispairs by Taq DNA polymerase: Implications for single nucleotide discrimination in PCR. Nucleic Acids Res. 1992, 20, 4567–4573. [Google Scholar] [CrossRef] [Green Version]
- Kwok, S.; Kellogg, D.E.; McKinney, N.; Spasic, D.; Goda, L.; Levenson, C.; Sninsky, J.J. Effects of primer-template mismatches on the polymerase chain reaction: Human immunodeficiency virus type 1 model studies. Nucleic Acids Res. 1990, 18, 999–1005. [Google Scholar] [CrossRef]
- Hayashi, K.; Hashimoto, N.; Daigen, M.; Ashikawa, I. Development of PCR-based SNP markers for rice blast resistance genes at the Piz locus. Theor. Appl. Genet. 2004, 108, 1212–1220. [Google Scholar] [CrossRef]
- Liu, Q.; Thorland, E.C.; Heit, J.A.; Sommer, S.S. Overlapping PCR for bidirectional PCR amplification of specific alleles: A rapid one-tube method for simultaneously differentiating homozygotes and heterozygotes. Genome Methods 1997, 7, 389–398. [Google Scholar] [CrossRef] [Green Version]
- Nakata, M.; Miyashita, T.; Kimura, R.; Nakata, Y.; Takagi, H.; Kuroda, M.; Yamaguchi, T.; Umemoto, T.; Yamakawa, H. MutMapPlus identified novel mutant alleles of a rice starch branching enzyme IIb gene for fine-tuning of cooked rice texture. Plant Biotechnol. J. 2018, 16, 111–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, K.B.; Yang, S.J.; Jo, Y.S.; Kang, S.S.; Nam, M. Development of Kompetitive Allele Specific PCR markers for identification of persimmon varieties using genotyping-by-sequencing. Electron. J. Biotechnol. 2021, 49, 72–81. [Google Scholar] [CrossRef]
- Li, G.; Jain, R.; Chern, M.; Pham, N.T.; Martin, J.A.; Wei, T.; Schackwitz, W.S.; Lipzen, A.M.; Duong, P.Q.; Jones, K.C.; et al. The sequences of 1504 mutants in the model rice variety Kitaake facilitate rapid functional genomic studies. Plant Cell 2017, 29, 1218–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rice Materials | Primer | Tm (°C) | Amplified Fragment Length (bp) |
|---|---|---|---|
| All lines | F a: 5′-CGTGAGGAGGGTTTAGGTGGAA-3′ | 59.5 | 680 |
| R a: 5′-TCCCGTCACAAACAGAAATCAA-3′ | 53.9 | ||
| #5 (iT/iG) | iT-F5 b: 5′-CAGGGGAGGTGATGATCTCT-3′ | 57.5 | 424 |
| iG-F5 b: 5′-CAGGGGAGGTGATGATCTCG-3′ | 59.5 | 424 | |
| R5: 5′-ACAACTGAACGGAATGAGCG-3′ | 55.4 | ||
| #1 (iT/dC) | iT-F1 b: 5′-CAGGGGAGGTGATGATCTCT-3′ | 57.5 | 424 |
| dC-F1 b: 5′-CAGGGGAGGTGATGATCTCG-3′ | 59.5 | 422 | |
| R1: 5′-ACAACTGAACGGAATGAGCG-3′ | 55.4 | ||
| #2 (iA/d6) | iA-F2 b: 5′-CGCGTCAGGGGAGGTTAT-3′ | 57.2 | 429 |
| d6-F2 b: 5′-CGCGTCAGGGGAGGTTAC-3′ | 59.5 | 422 | |
| R2: 5′-ACAACTGAACGGAATGAGCG-3′ | 55.4 | ||
| #12 (iA/d68) | iA-R12 b: 5′-AGGATGAATCATGATCACCCA-3′ | 53.7 | 292 |
| d68-R12 b: 5′-AGGATGAATCATGATCACCCC-3′ | 55.6 | 223 | |
| F12: 5′-GCCACCTTGTTGTTCTCGTC-3′ | 57.5 | ||
| #12 (iA/d68) | F c: 5′-GCCACCTTGTTGTTCTCGTC-3′ | 57.5 | 413/344 |
| R c: 5′-AGACGTGGACTGCGTGAAAT-3′ | 55.4 |
| PCR Reaction System | #5 | #1 | #2 | #12 |
|---|---|---|---|---|
| Reaction mixture (10 μL) | ||||
| 2 × Taq Master Mix (Quick Load) a | 2.5 μL | 2.5 μL | 2.5 μL | 2.5 μL |
| Forward primer (10 μM) | 0.5 μL | 0.3 μL | 0.3 μL | 0.5 μL |
| Reverse primer (10 μM) | 0.5 μL | 0.3 μL | 0.3 μL | 0.5 μL |
| Template DNA (150 ng/μL) | 1.5 μL | 1.0 μL | 1.0 μL | 1.5 μL |
| Sterilized deionized water | 5 μL | 5.9 μL | 5.9 μL | 5 μL |
| PCR program | ||||
| Stage 1: Initial denaturation (5 min) | 95 °C | 95 °C | 95 °C | 95 °C |
| Stage 2: 28 reaction cycles: Denaturation (30 s) | 95 °C | 95 °C | 95 °C | 95 °C |
| Annealing (30 s) | 58 °C | 58 °C | 57 °C | 53 °C |
| Extension (28 s) | 72 °C | 72 °C | 72 °C | 72 °C |
| Stage 3: Final extension (7 min) | 72 °C | 72 °C | 72 °C | 72 °C |
| Stage 4: Storage (forever) | 16 °C | 16 °C | 16 °C | 16 °C |
| T1 Line | Genotype | Segregation of T2 Plants | |||
|---|---|---|---|---|---|
| Total Number | Targeted Mutation Number | Expected Segregation Ratio | χ2 | ||
| #5 | iT/iG | 33 | 6 iT, 18 iT/iG, 9 iG | 1:2:1 | 0.818 NS |
| #1 | iT/dC | 53 | 14 iT, 22 iT/dC, 17 dC | 1:2:1 | 1.867 NS |
| #2 | iA/d6 | 29 | 7 iA, 15 iA/d6, 7 d6 | 1:2:1 | 0.035 NS |
| #12 | iA/d68 | 30 | 8 iA, 14 iA/d68, 8 d68 | 1:2:1 | 0.133 NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Ren, Y.; Xu, X.; Wang, H.; Wei, C. Application of Allele Specific PCR in Identifying Offspring Genotypes of Bi-Allelic SbeIIb Mutant Lines in Rice. Plants 2022, 11, 524. https://doi.org/10.3390/plants11040524
Jiang Y, Ren Y, Xu X, Wang H, Wei C. Application of Allele Specific PCR in Identifying Offspring Genotypes of Bi-Allelic SbeIIb Mutant Lines in Rice. Plants. 2022; 11(4):524. https://doi.org/10.3390/plants11040524
Chicago/Turabian StyleJiang, Yongqi, Yinhui Ren, Xin Xu, Hao Wang, and Cunxu Wei. 2022. "Application of Allele Specific PCR in Identifying Offspring Genotypes of Bi-Allelic SbeIIb Mutant Lines in Rice" Plants 11, no. 4: 524. https://doi.org/10.3390/plants11040524
APA StyleJiang, Y., Ren, Y., Xu, X., Wang, H., & Wei, C. (2022). Application of Allele Specific PCR in Identifying Offspring Genotypes of Bi-Allelic SbeIIb Mutant Lines in Rice. Plants, 11(4), 524. https://doi.org/10.3390/plants11040524