
Comparison of Magnoliaceae Plastomes: Adding Neotropical Magnolia to the Discussion

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
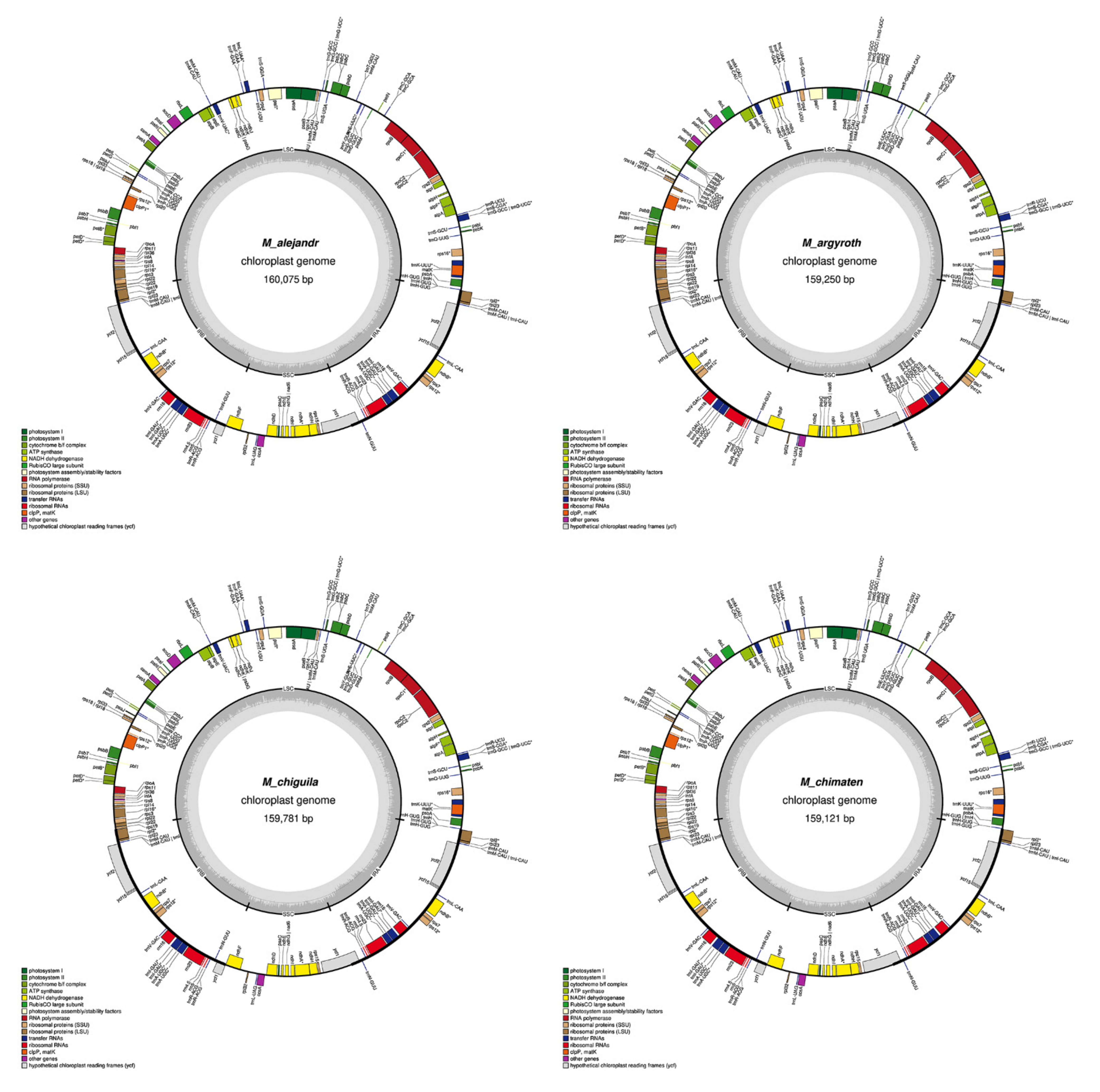
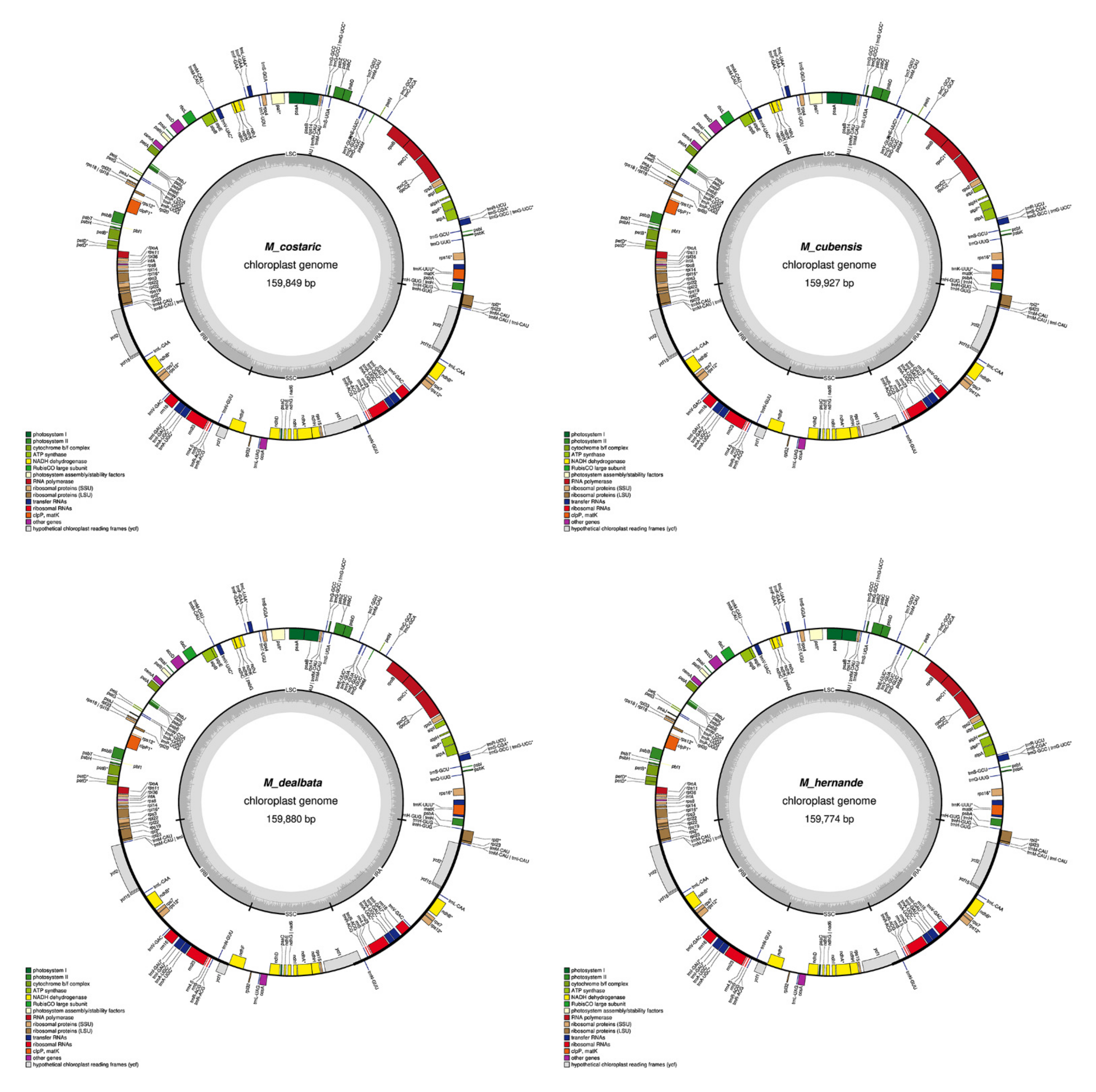
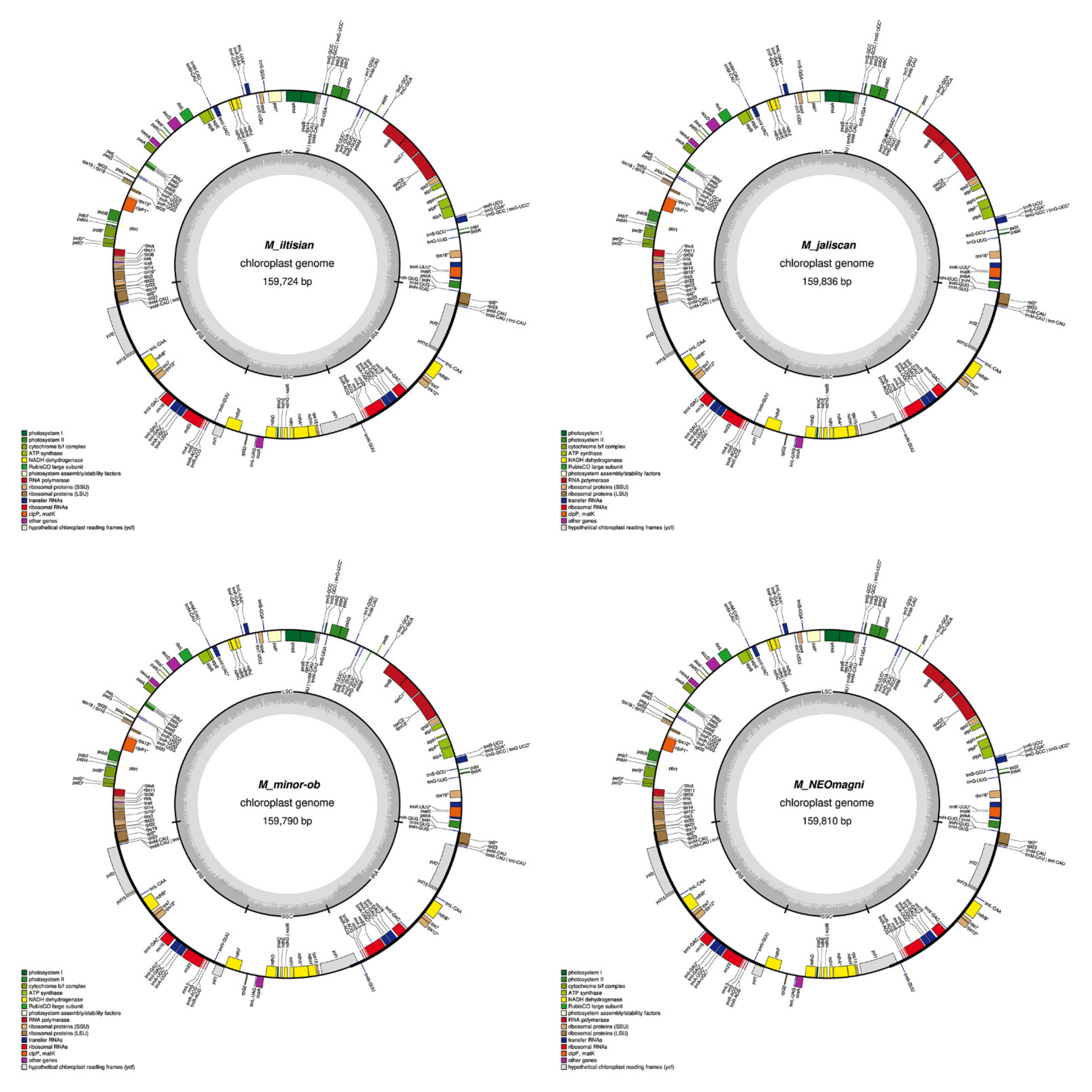
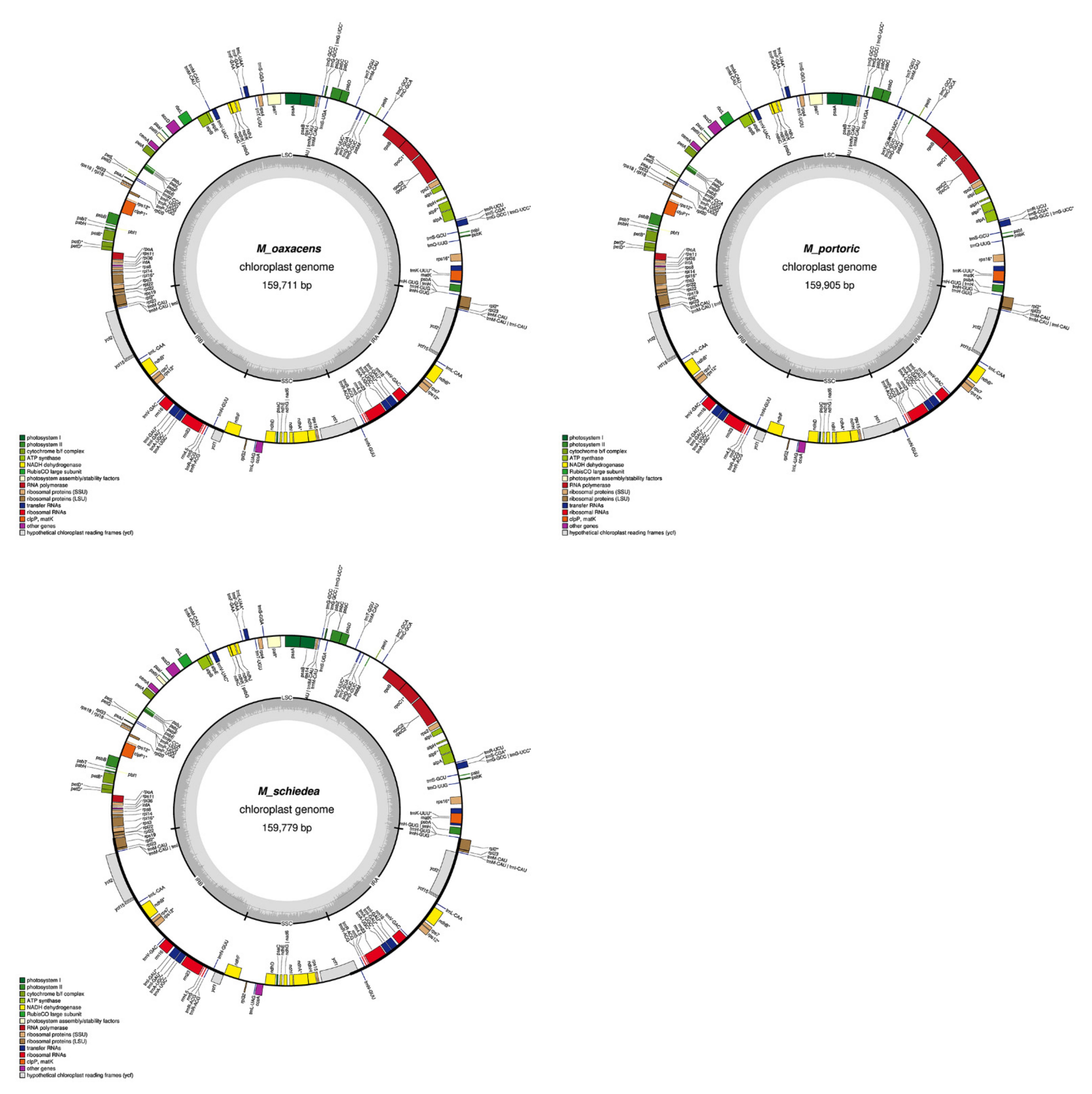
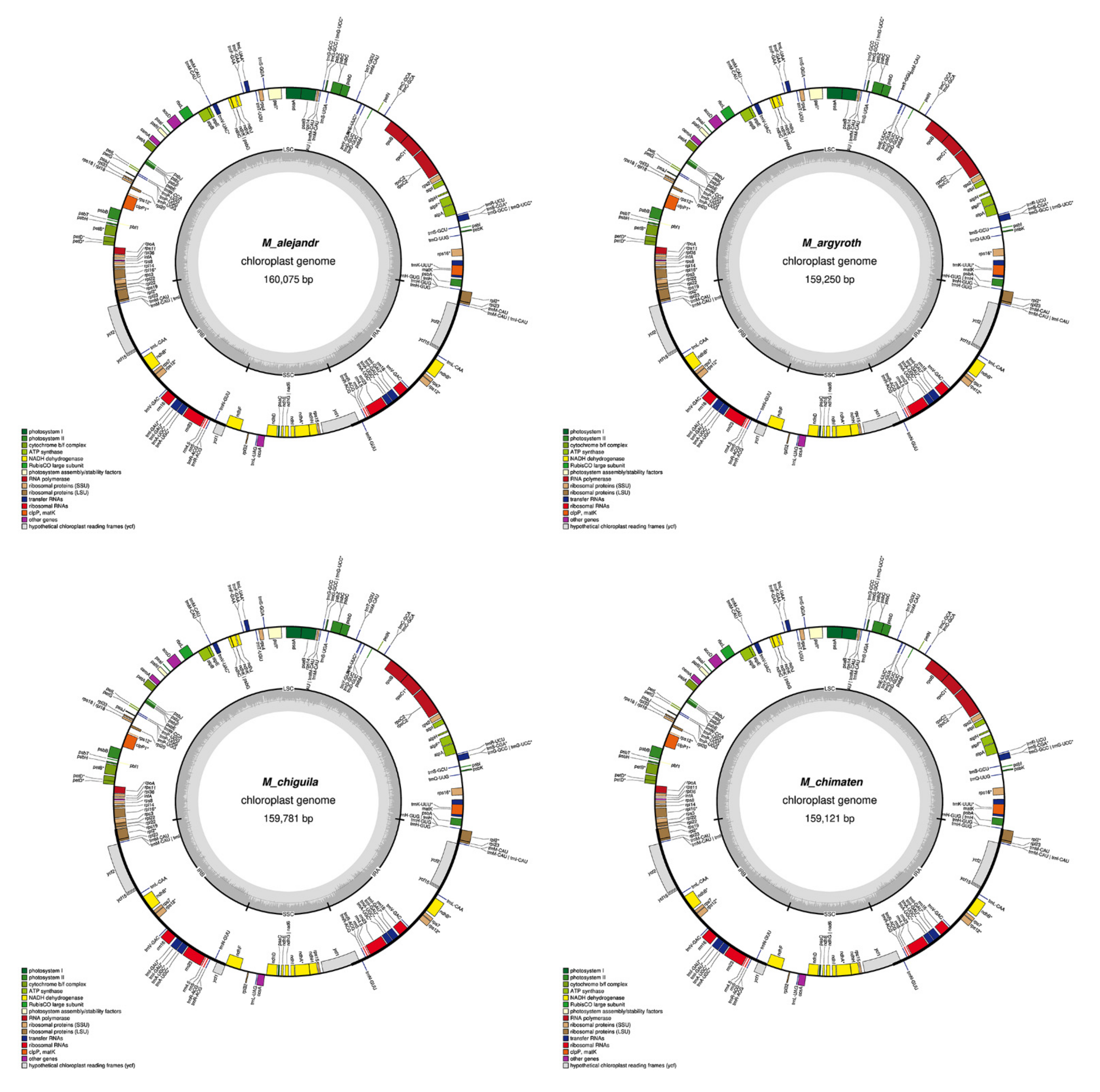
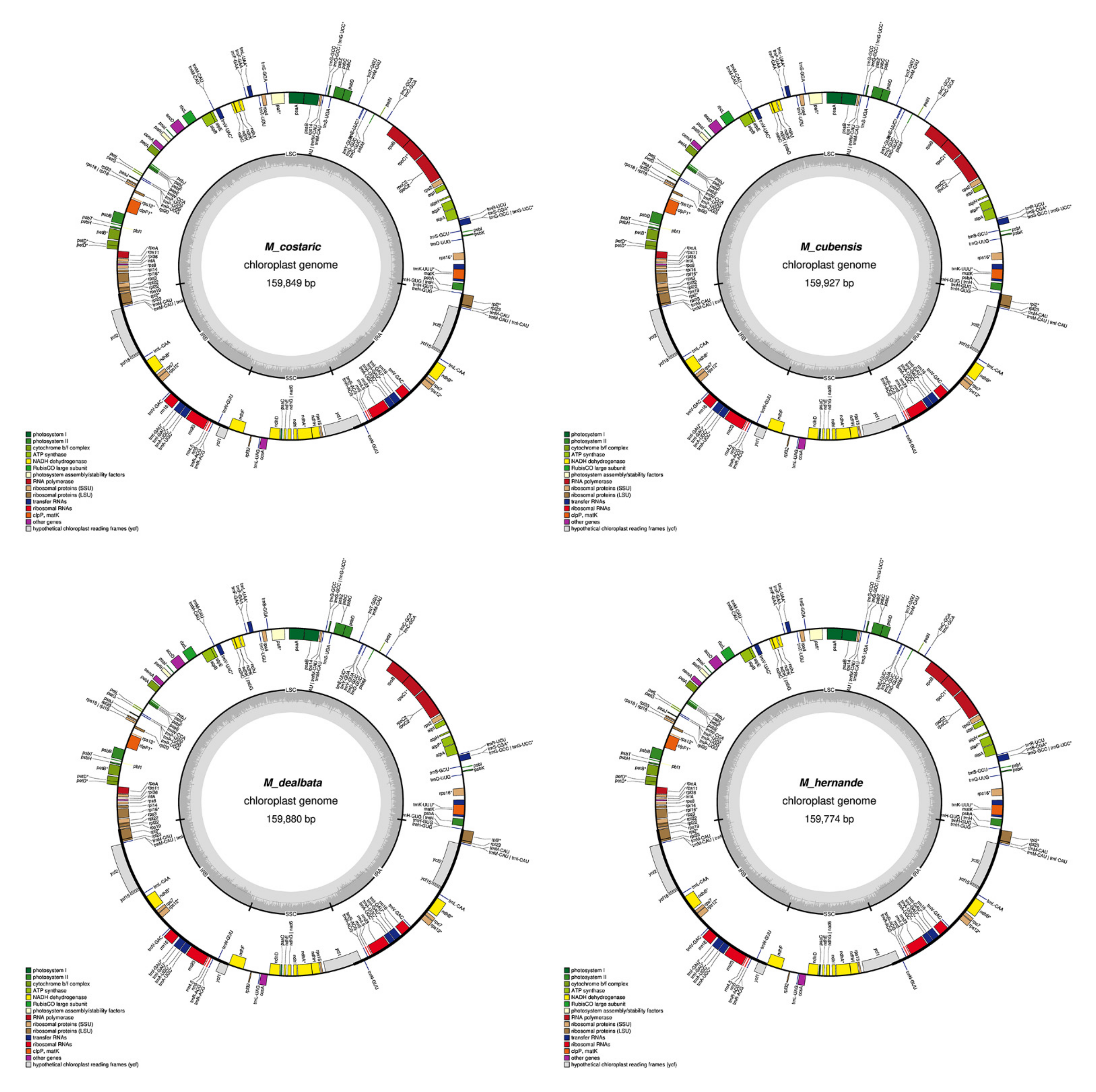
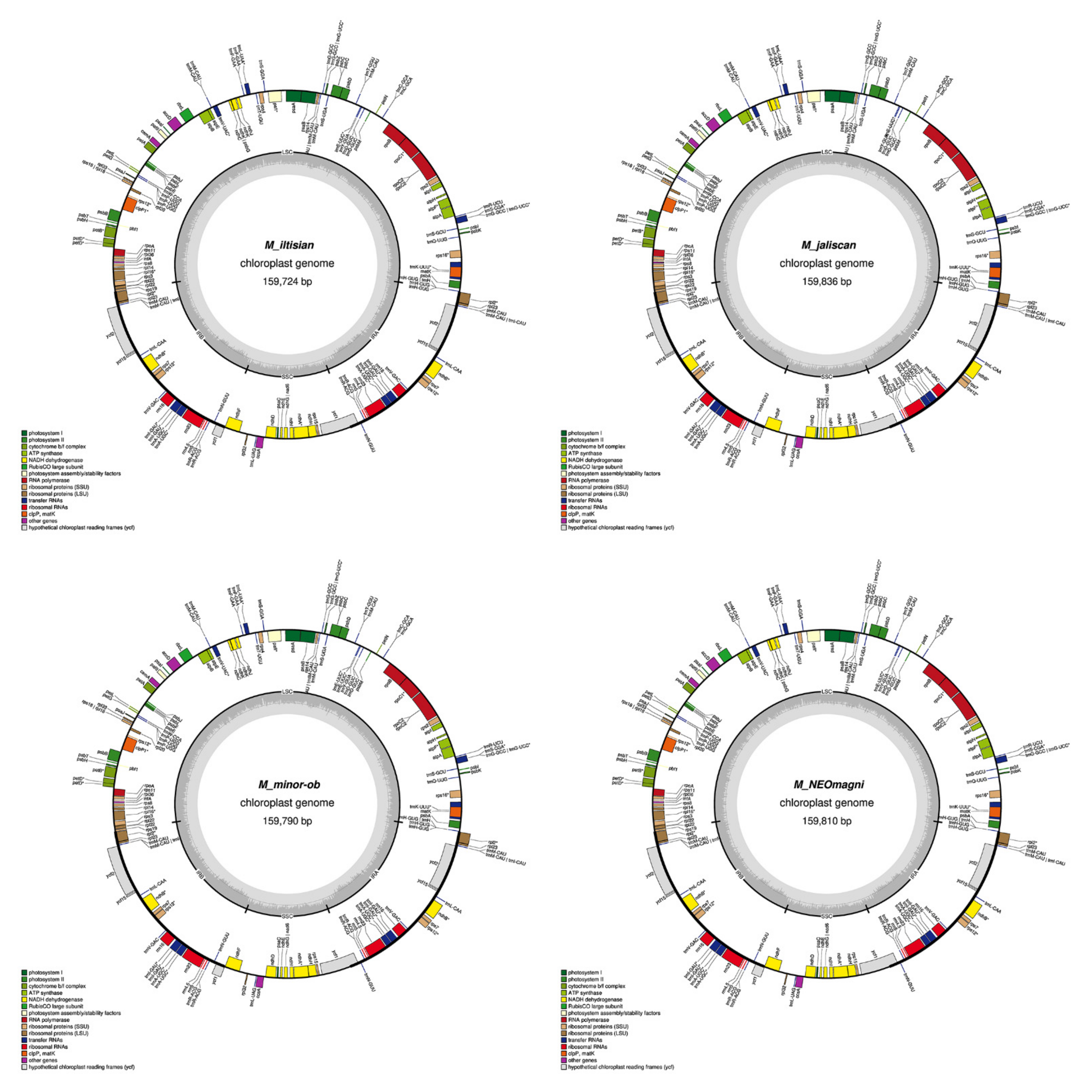
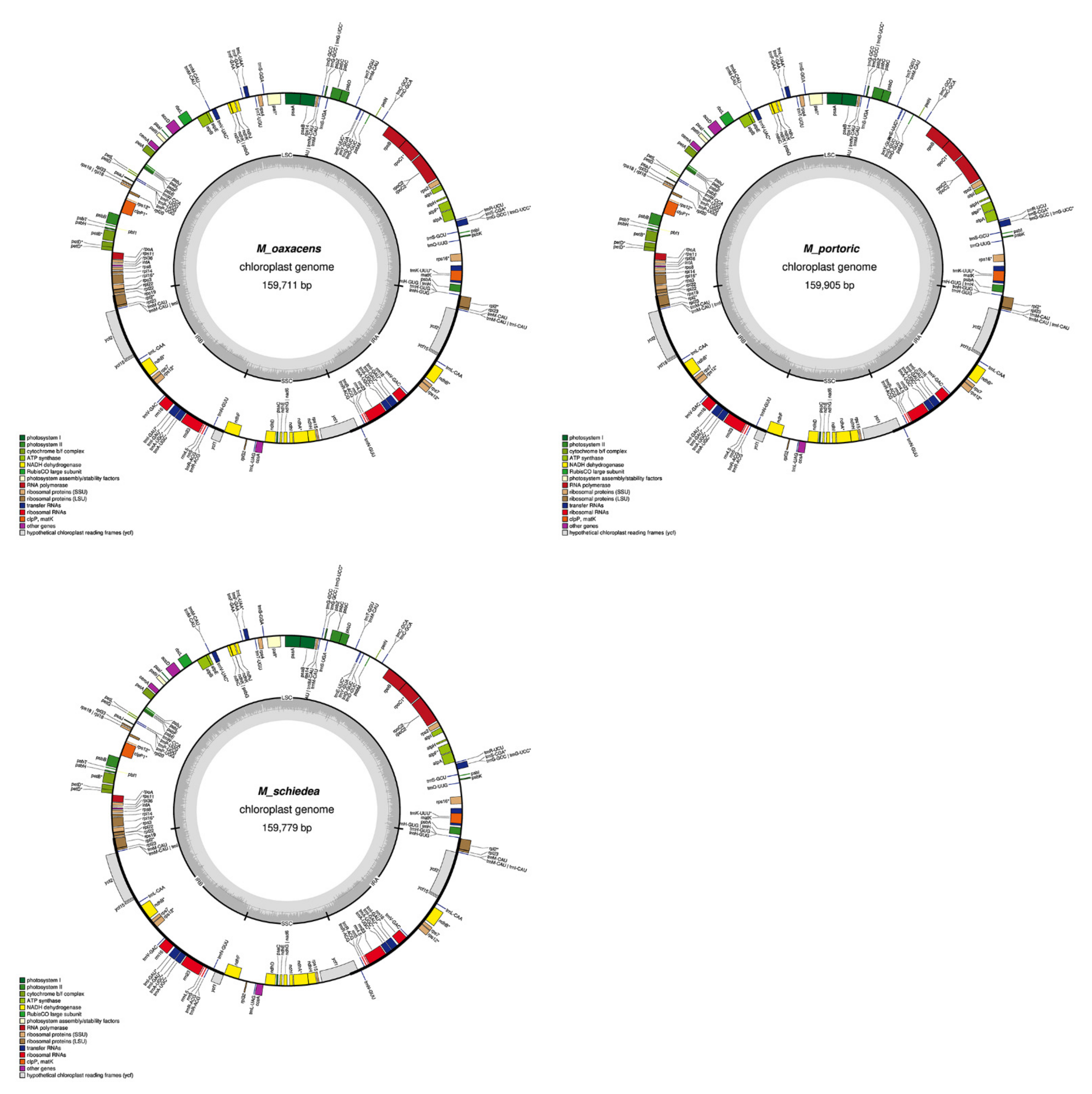
2.1. Chloroplast Assembly and the Annotation of Neotropical Magnolia
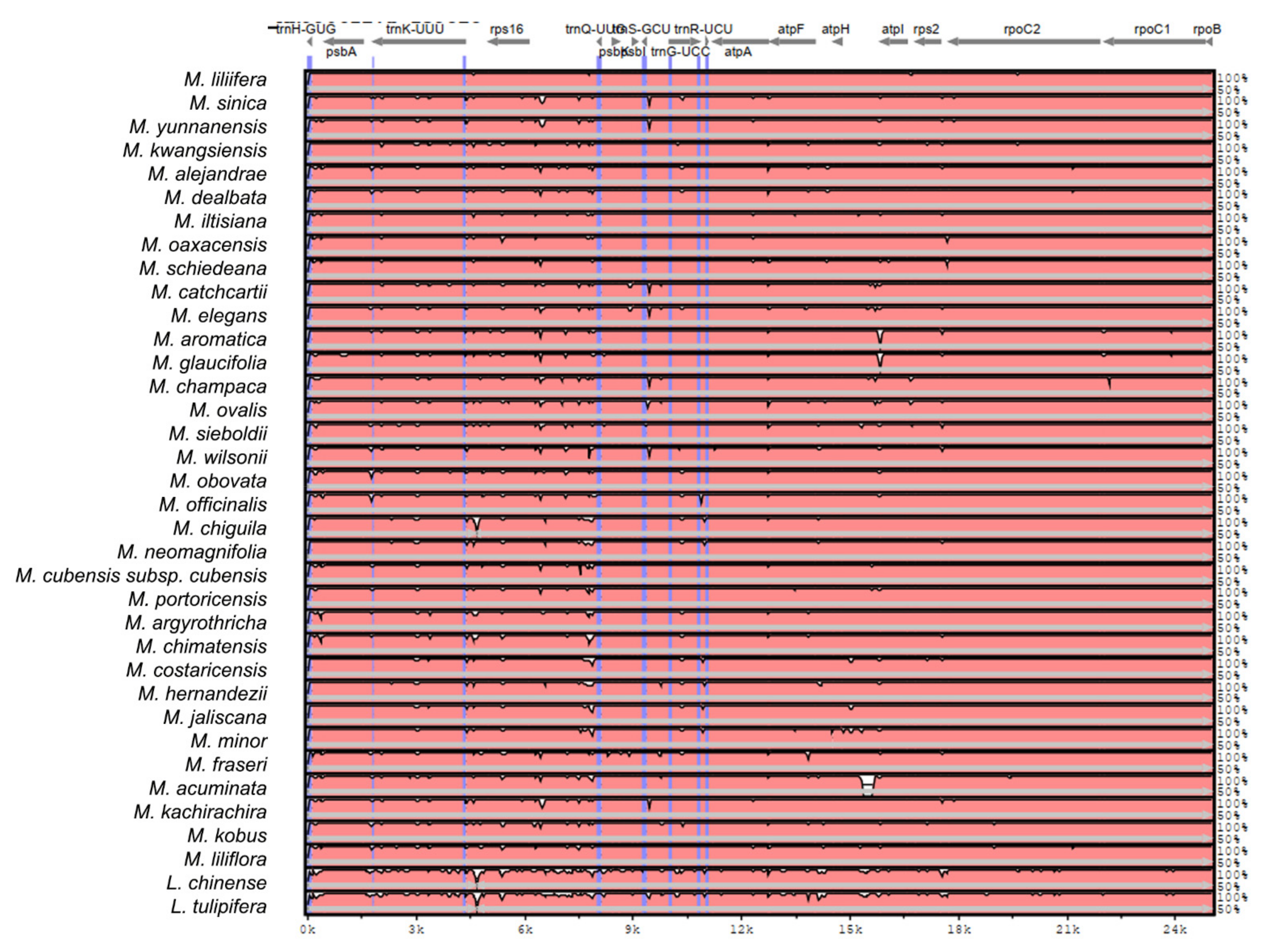
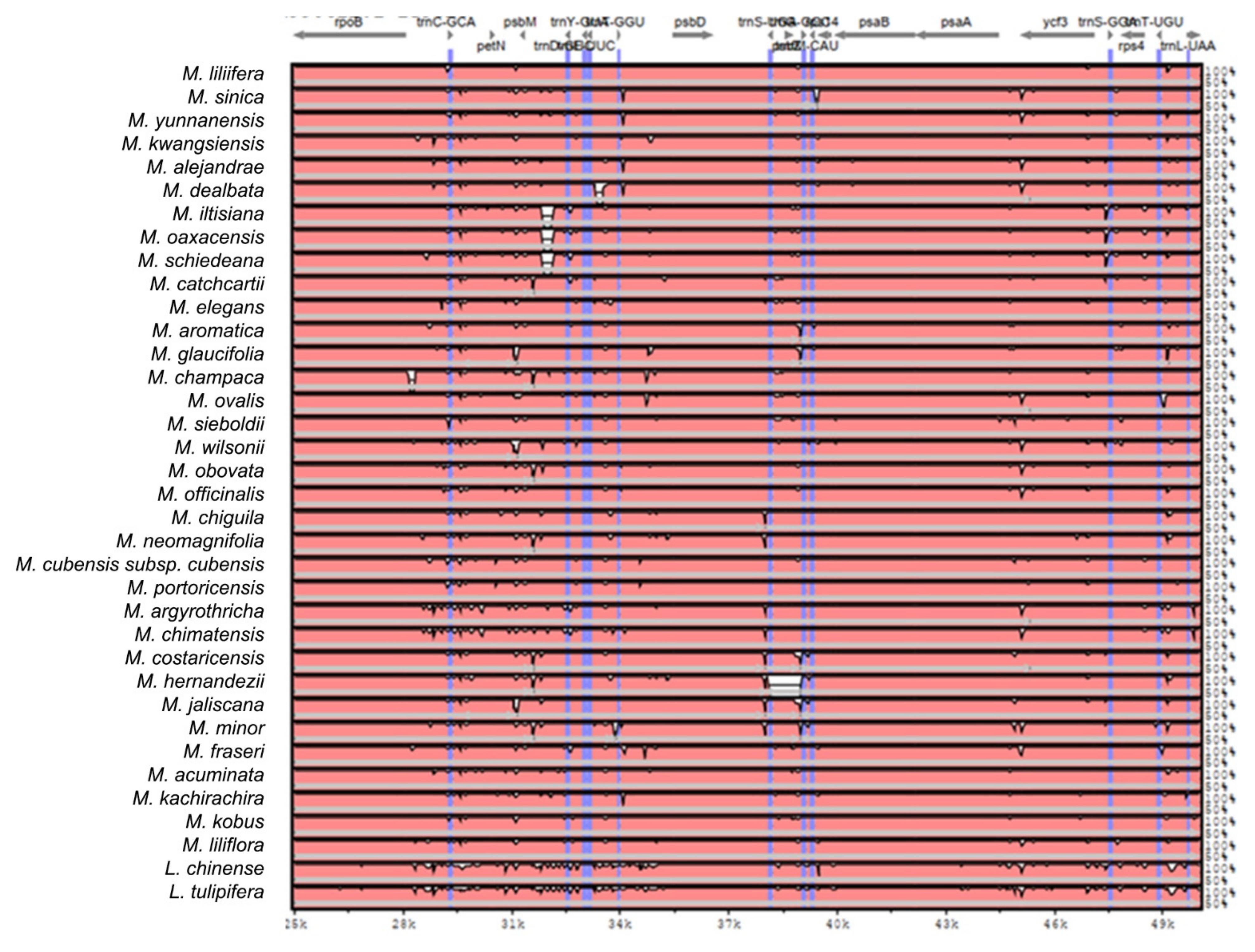
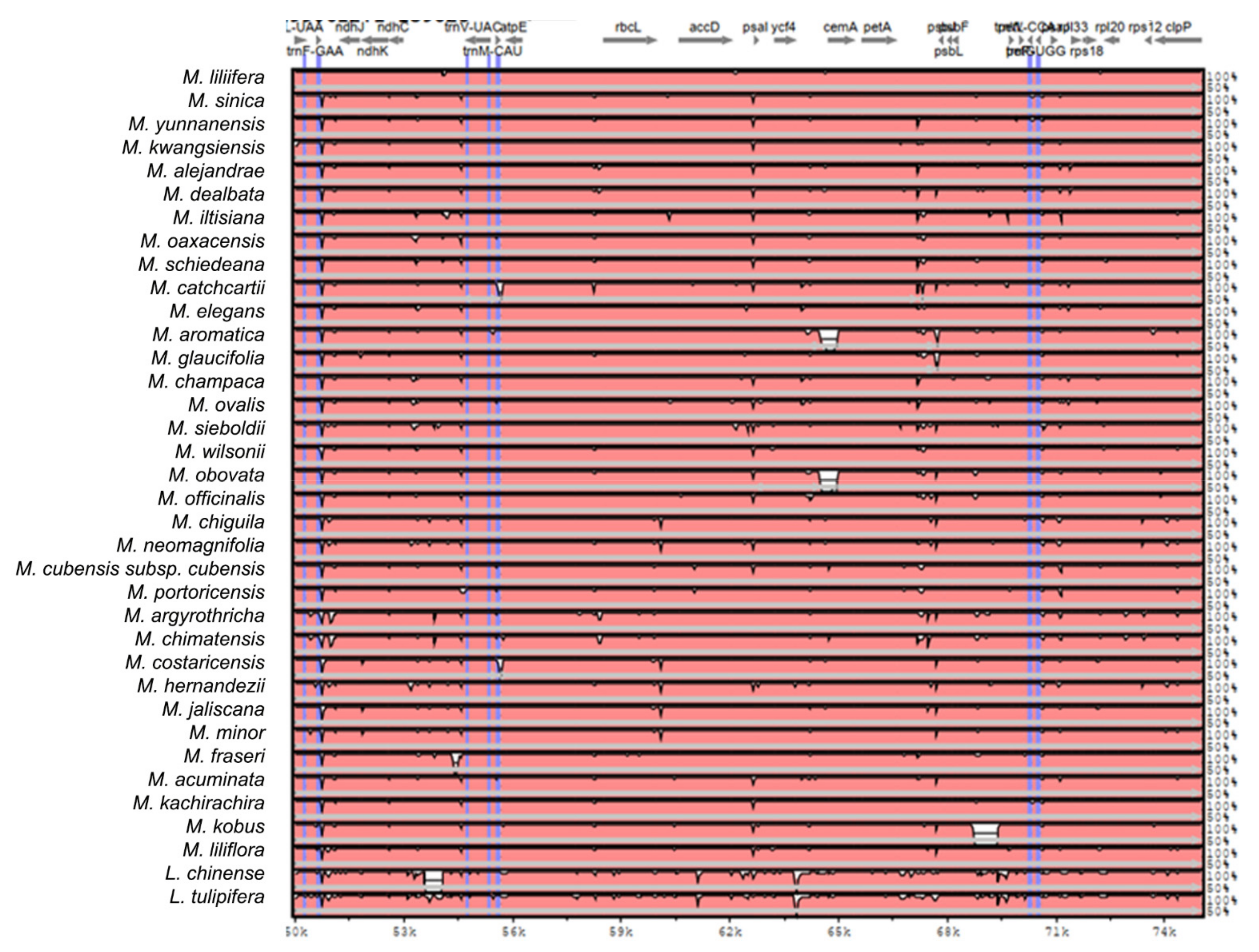
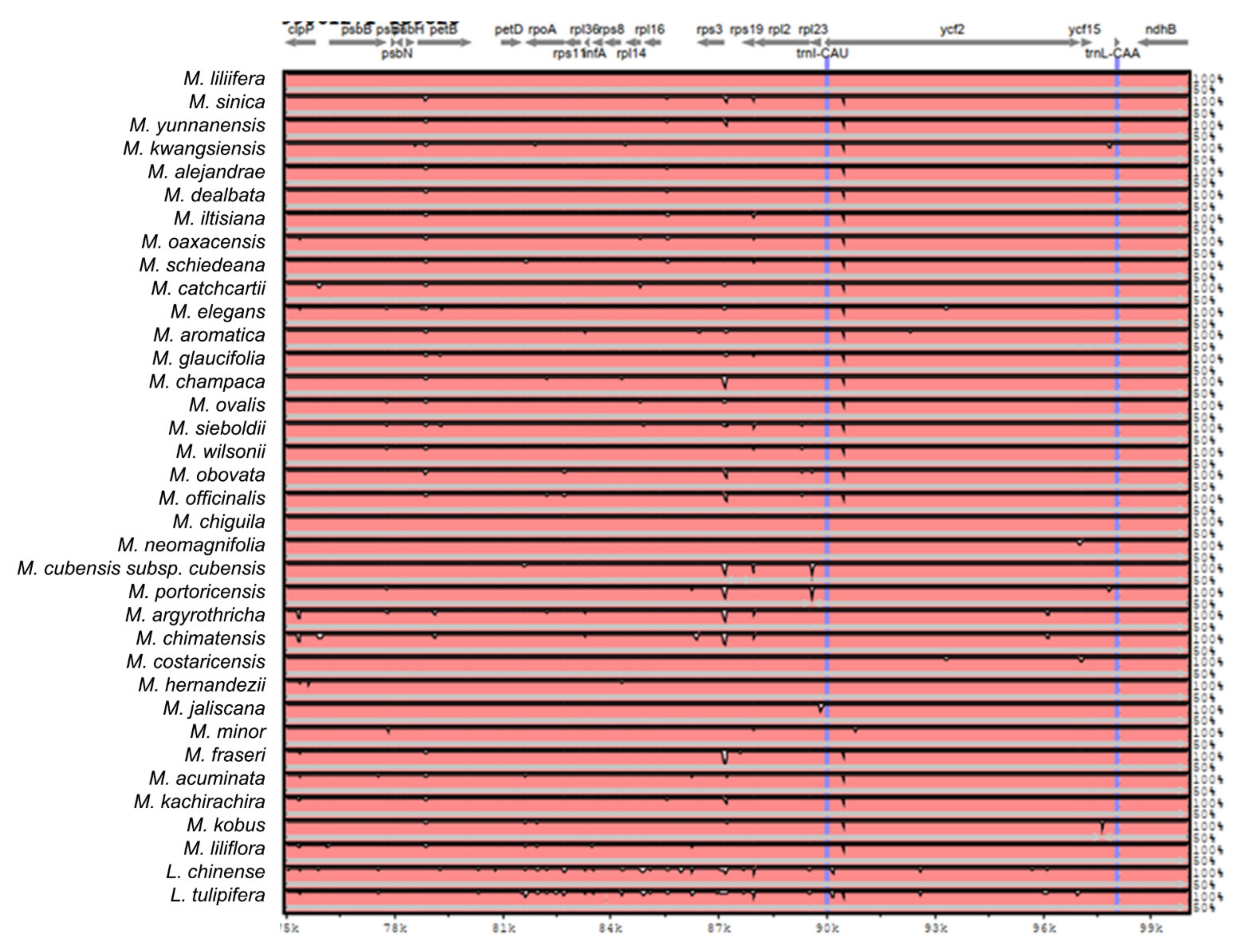
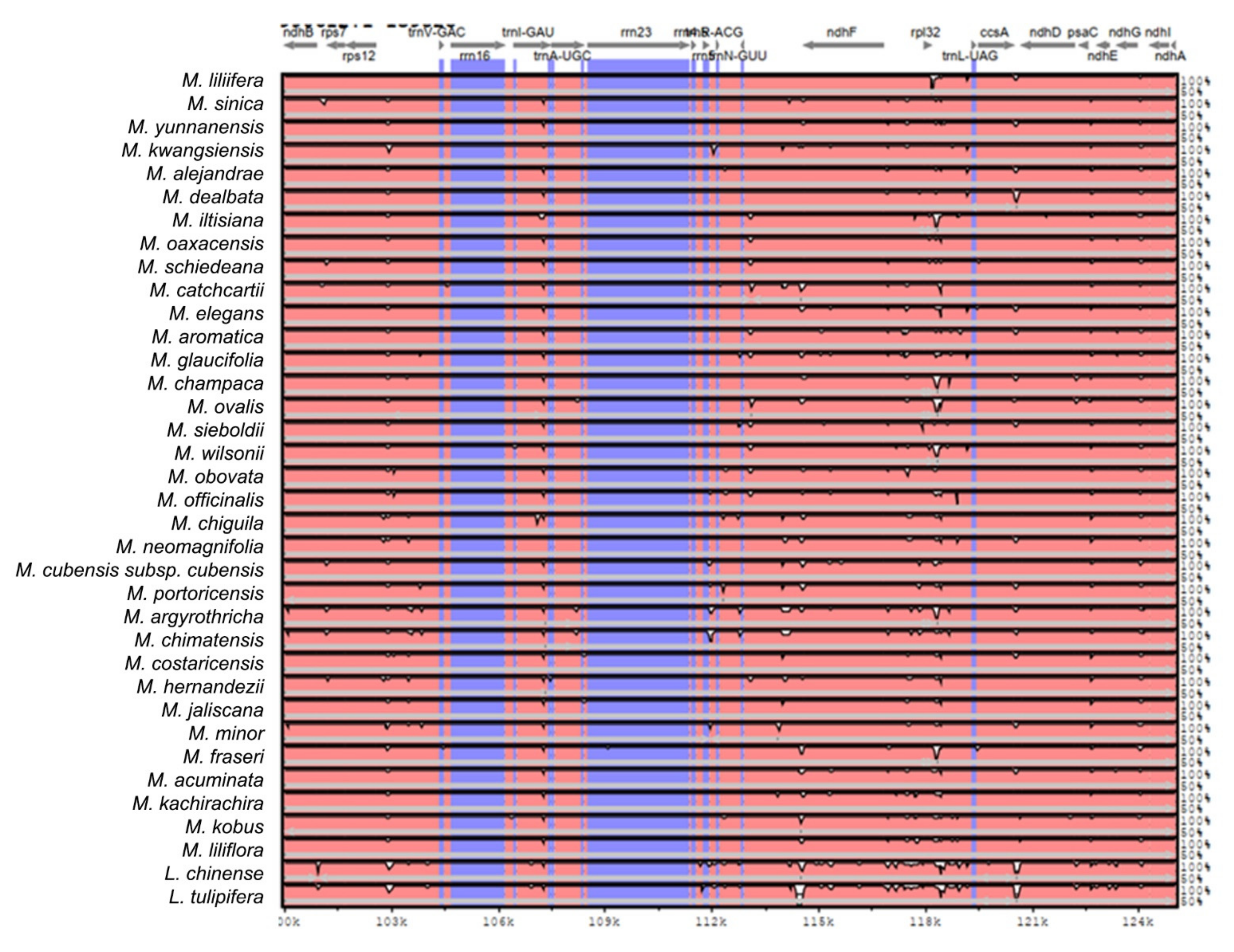
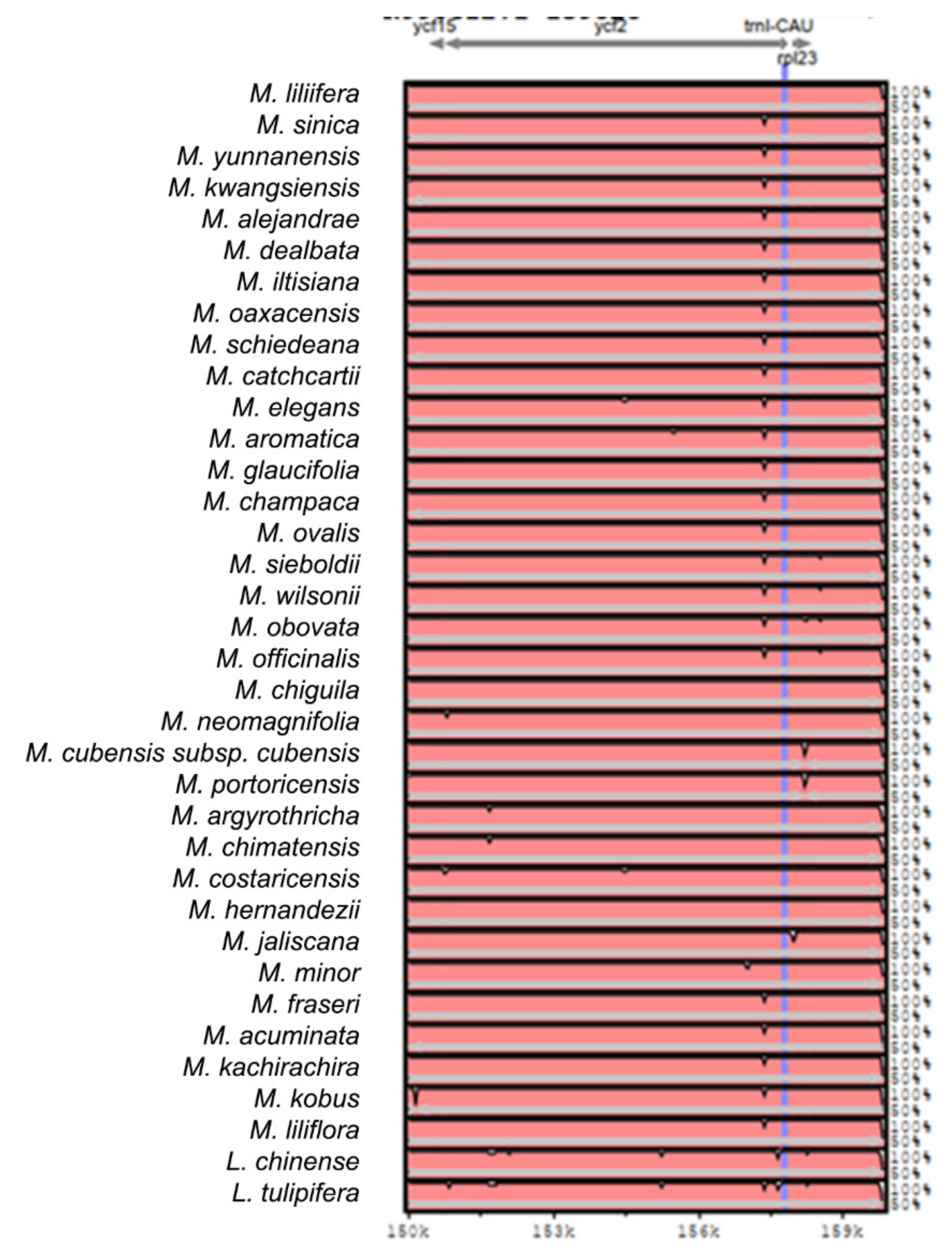
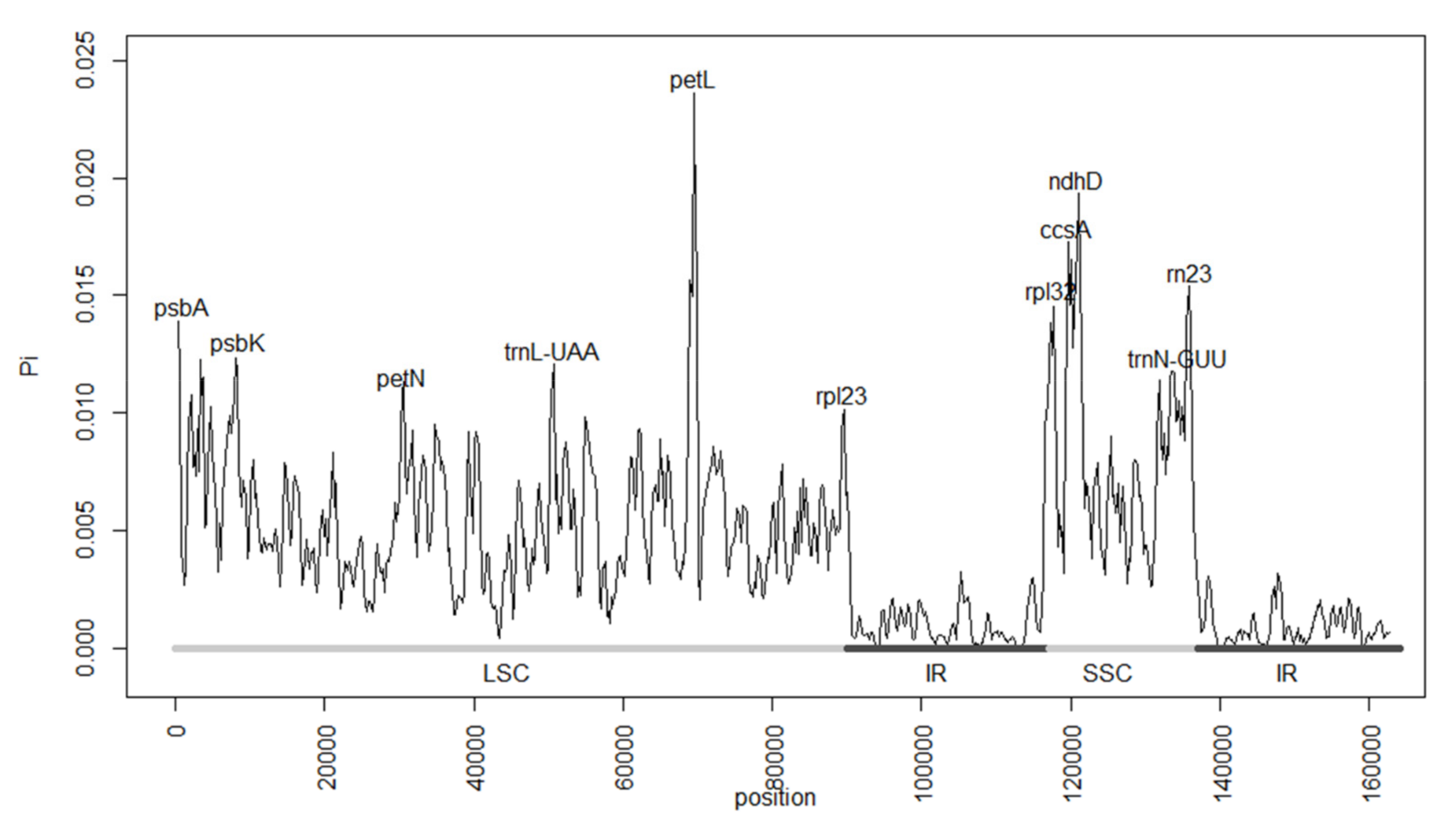
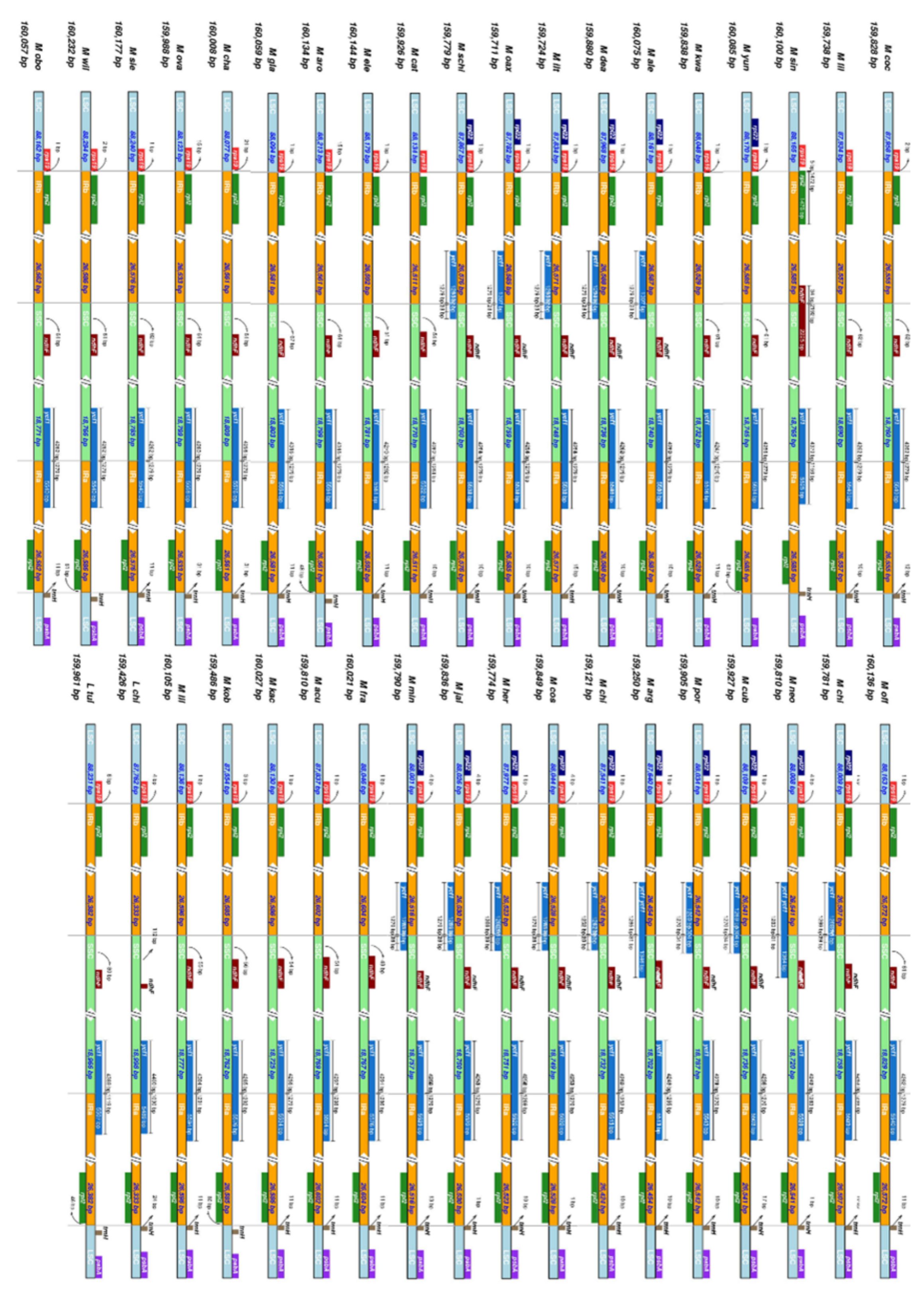
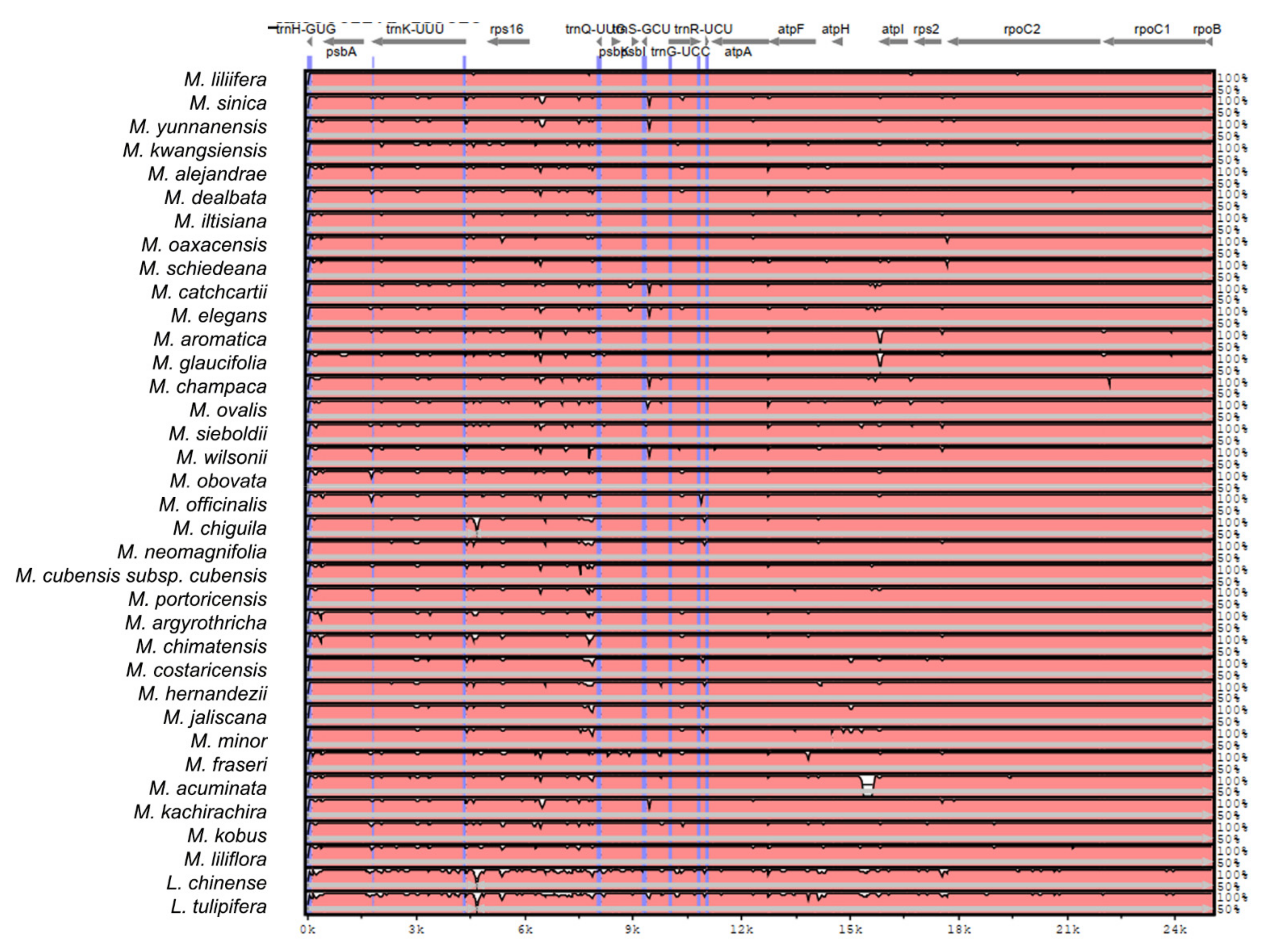
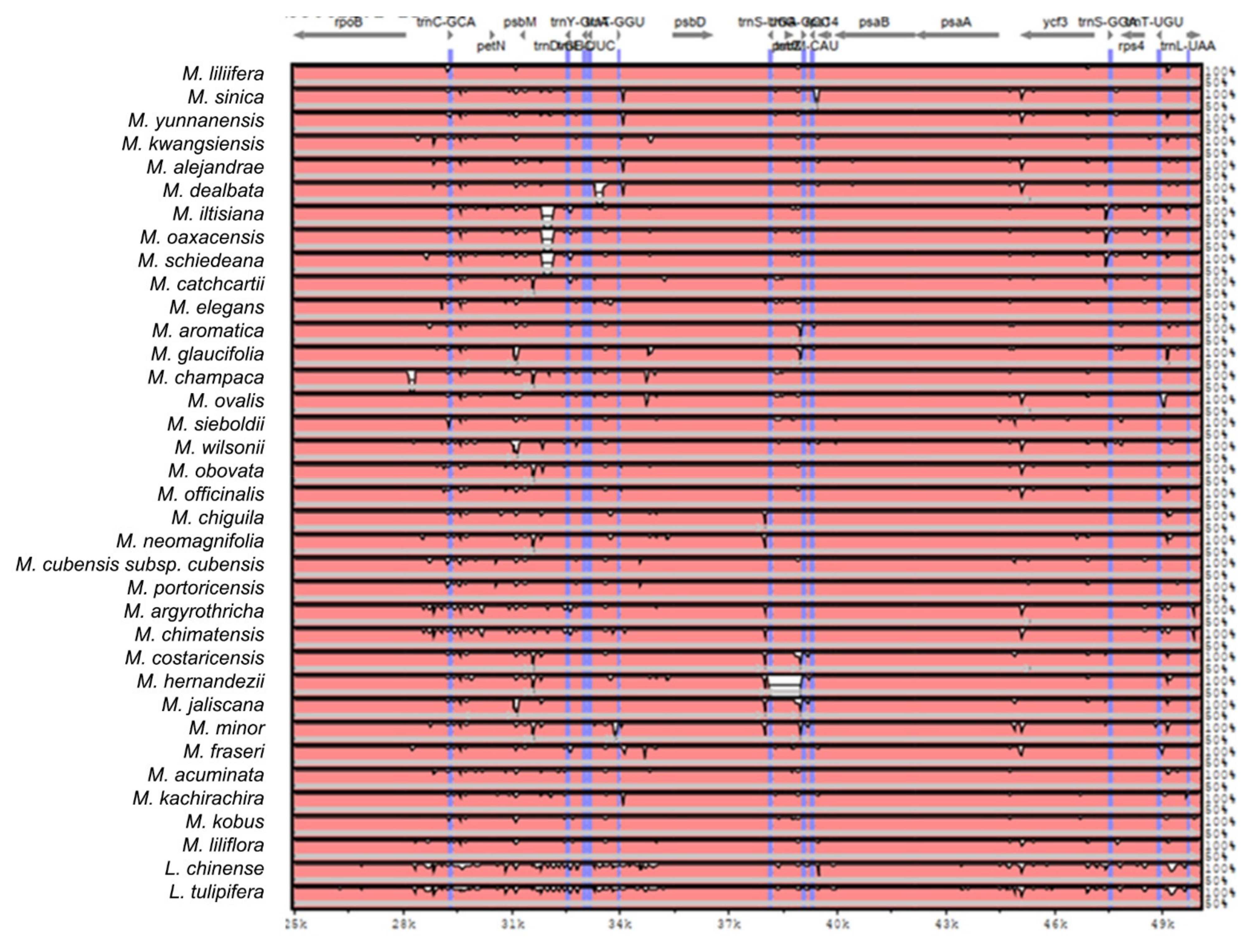
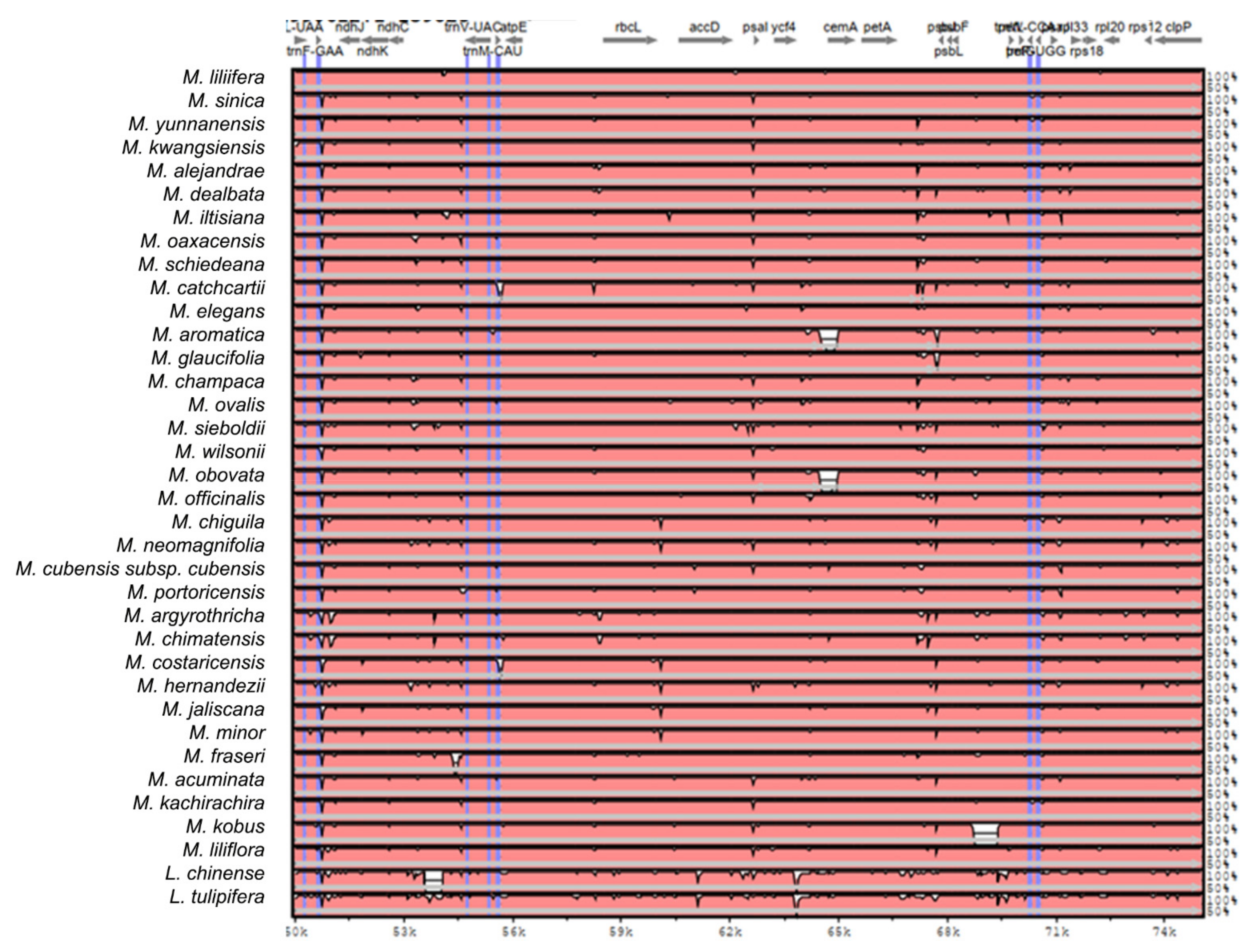
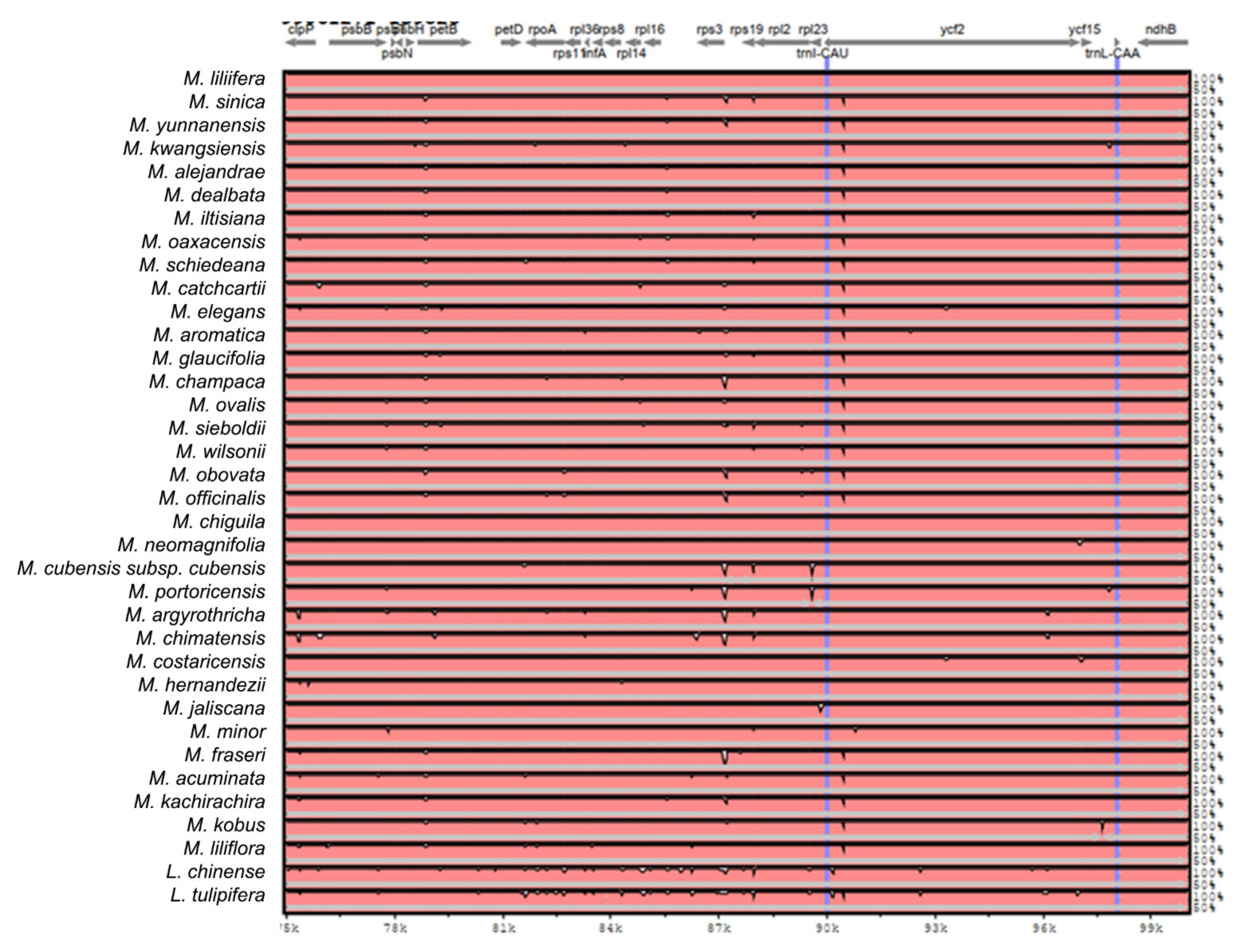
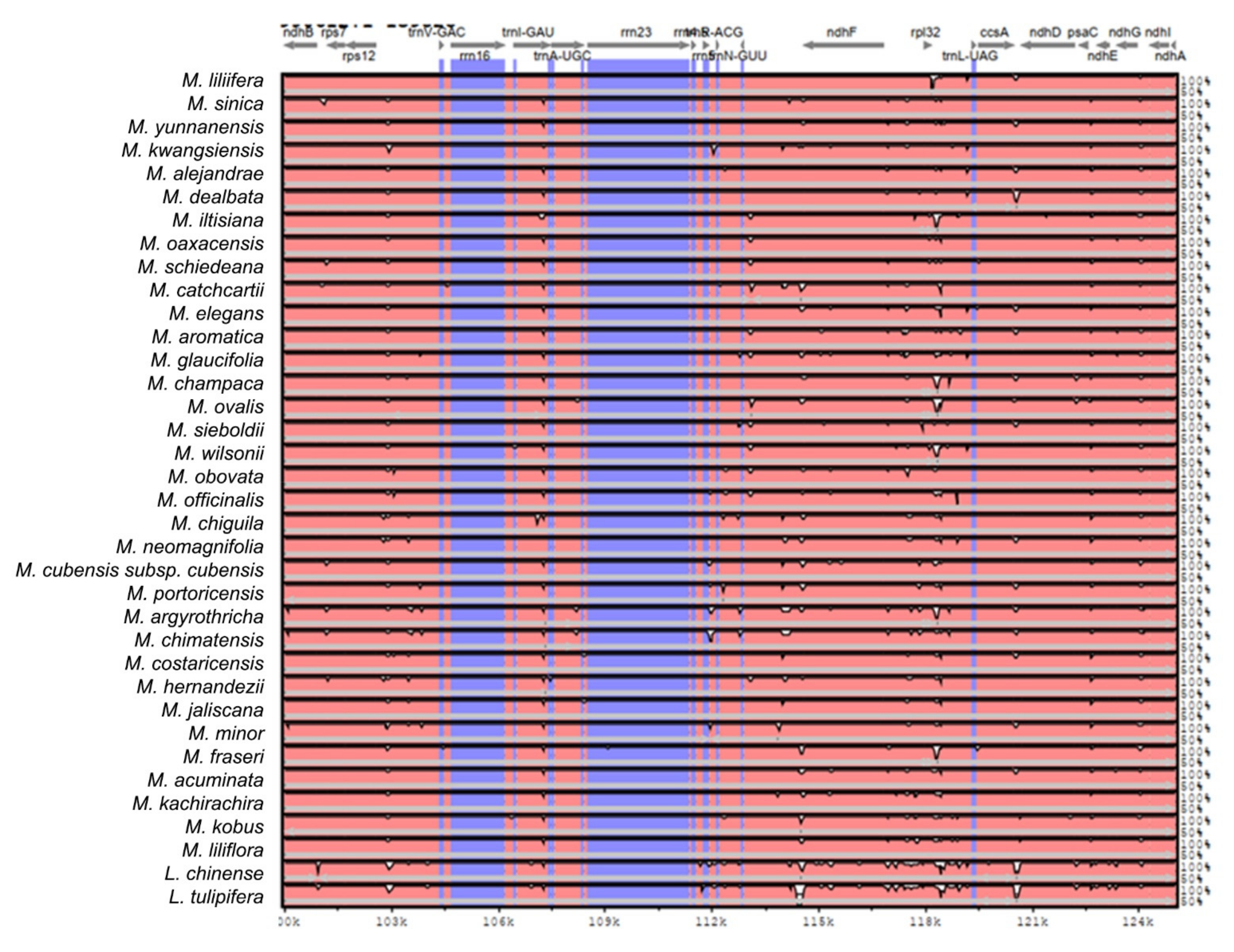
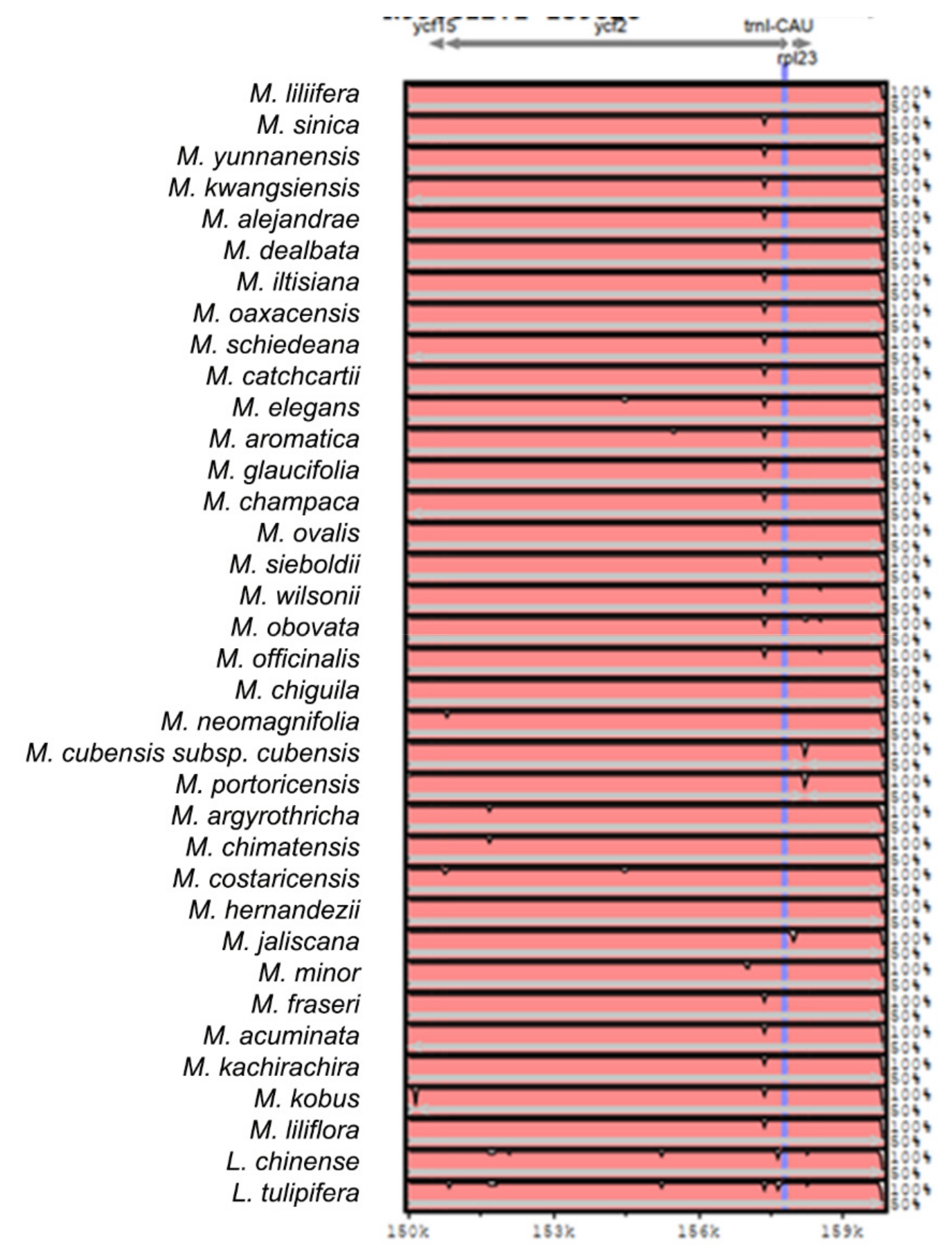
2.2. The Magnoliaceae Plastome
Gene Organization and Nucleotide Diversity of the Magnoliaceae plastomes
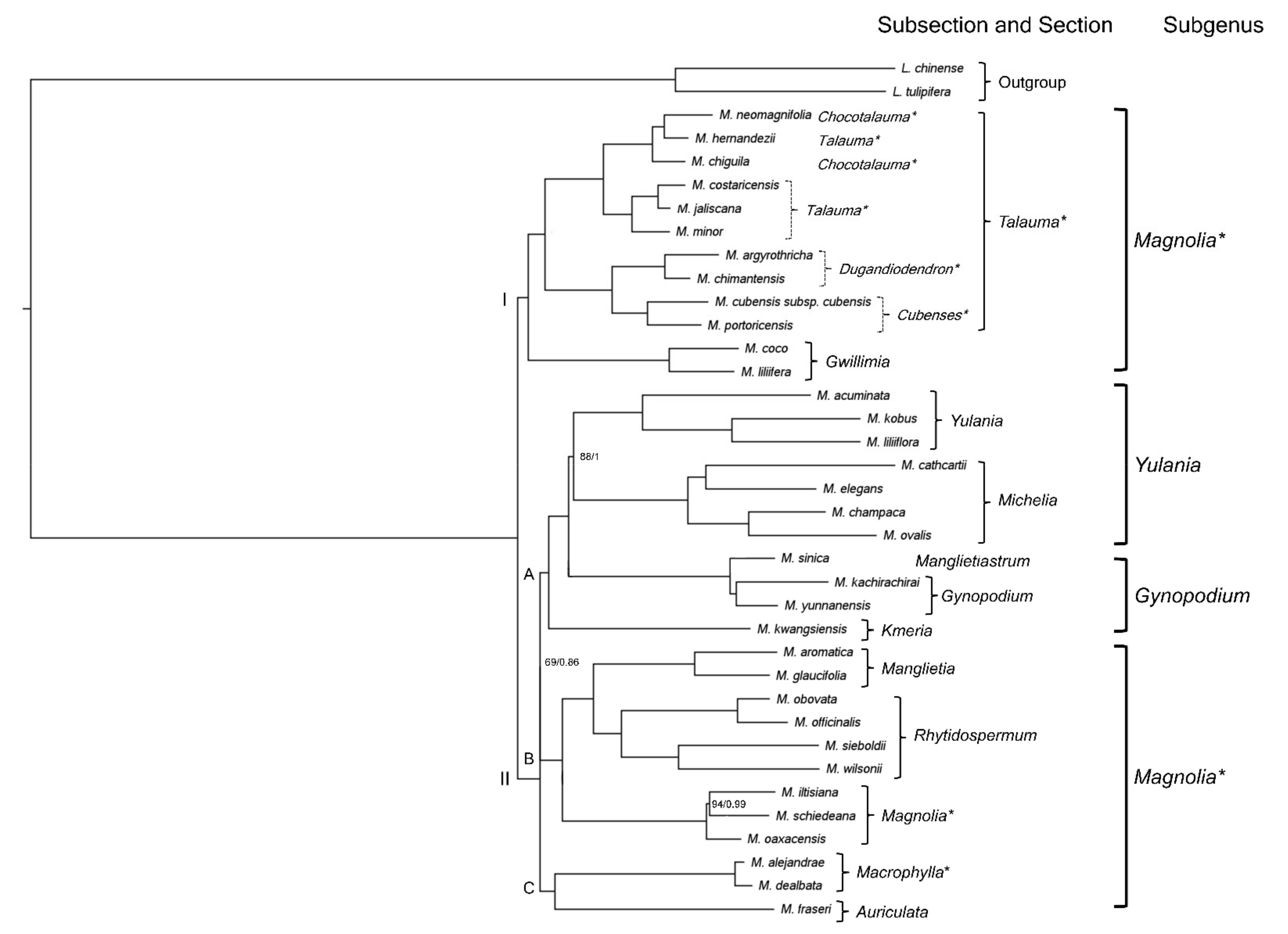
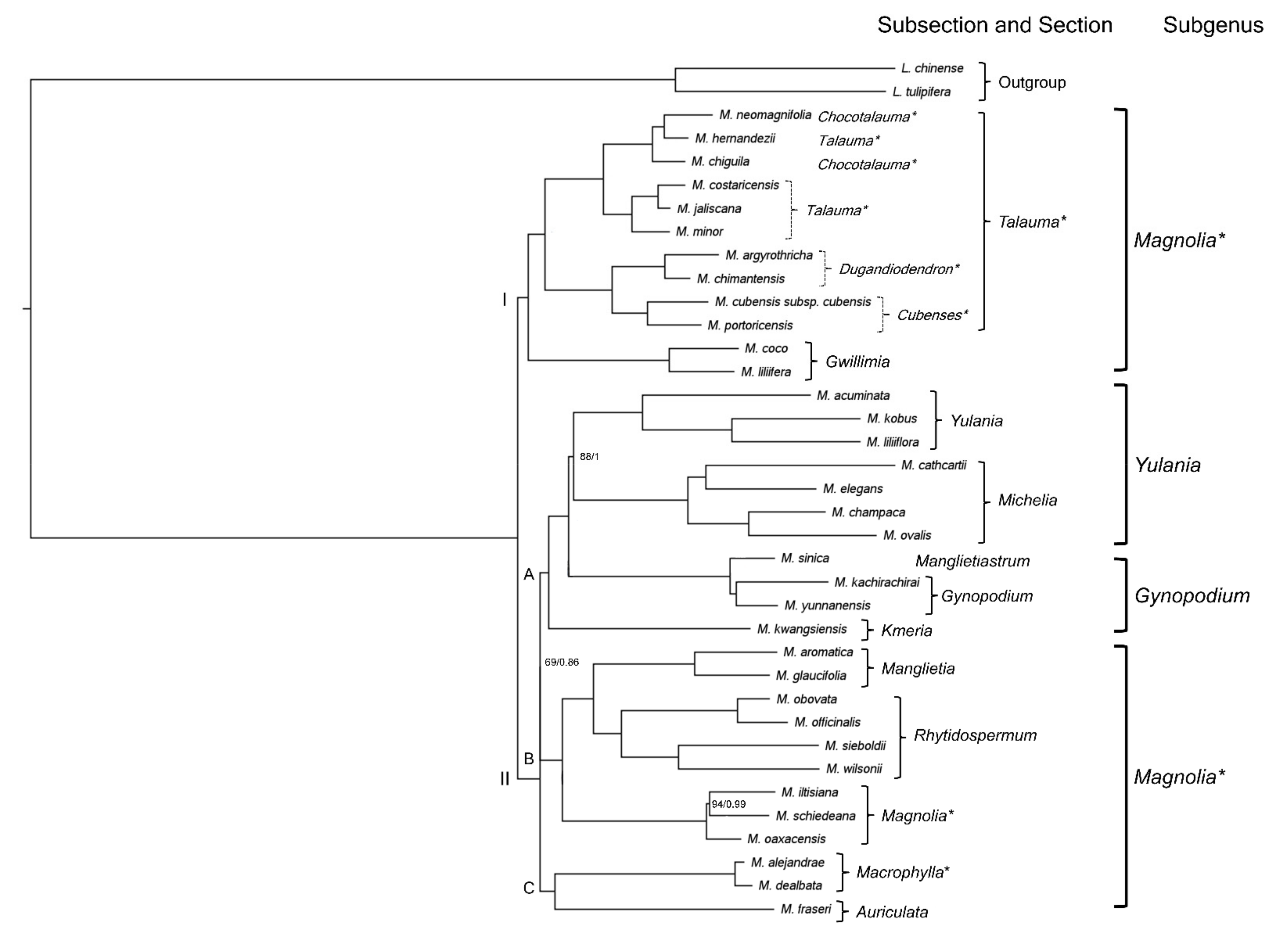
2.3. Neotropical Magnolia Phylogeny
3. Discussion
3.1. Chloroplast Assembly and the Annotation of Neotropical Magnolia
3.2. The Magnoliaceae Plastome
3.3. Neotropical Magnolia Phylogeny
4. Materials and Methods
4.1. Sampling, DNA Extraction, and Sequencing
4.2. Chloroplast Assembly and Annotation
4.3. Genome Comparison
4.4. Phylogenomics of the Magnoliacceae Plastome
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Section (Subsection) | Species | Voucher | Raw Reads | Trimmed Reads | NCBI Reference |
|---|---|---|---|---|---|
| Macrophylla | M. alejandrae García-Mor. and Iamonico | M. Mata 1188b | 1,583,270 | 1,369,706 | TBD |
| M. dealbata Zucc. | M. Mata 0866a | 1,576,433 | 1,435,083 | TBD | |
| Magnolia | M. iltisiana A. Vázquez | S. Cházaro B. and M. Rodríguez 8590 | 2,654,727 | 2,564,393 | TBD |
| M. oaxacensis A. Vázquez | M.S. Samain and E. Martínez 2019-019 | 1,857,563 | 1,792,016 | TBD | |
| M. schiedeana Schltdl. | F. Aldaba 187 | 1,713,367 | 1,672,698 | TBD | |
| Talauma (Chocotalauma) | M. chiguila F. Arroyo, Á.J. Pérez, and A. Vázquez | E. Veltjen and A. Dahua 2018-005 | 1,678,118 | 1,582,603 | TBD |
| M. neomagnifolia I.M.Turner | NA | 1,772,721 | 1,731,278 | TBD | |
| Talauma (Cubenses) | M. cubensis Urb. subsp. cubensis | A. Palmarola et al. HFC-89195 | 638,499 | 586,472 | TBD |
| M. portoricensis Bello | E. Veltjen et al. 2016-033 | 1,613,856 | 1,549,640 | TDB | |
| Talauma (Dugandiodendron) | M. argyrothricha (G. Lozano C.) Govaerts | NA | 2,008,282 | 1,910,887 | TBD |
| M. chimantensis Steyerm. and Maguire | J. Steyermark 1191 | 1,603,135 | 1,514,091 | TBD | |
| Talauma (Talauma) | M. costaricensis A. Vázquez | J.E. Jiménez 4622 | 1,889,242 | 1,806,280 | TBD |
| M. hernandezii (G. Lozano C.) Govaerts | Hernández 1001 | 1,089,114 | 716,041 | TBD | |
| M. jaliscana A. Vázquez and R. Guzmán | J.A. Vázquez García et al. 9335 | 1,800,168 | 1,671,463 | TBD | |
| M. minor (Urb.) Govaerts | B. Falcón HFC88953 | 1,750,979 | 1,611,208 | TBD |
| Algorithm A1. Unix commands used for the assembly of the WGS reads. |
| ### run fastQC mkdir raw_wgs/out_fastQC fastqc raw_wgs/*.fastq.gz --outdir=raw_wgs/out_fastQC ### run multiQC mkdir raw_wgs/out_fastQC/multiQC multiqc raw_wgs/out_fastQC --outdir raw_wgs/out_fastQC/multiqc #################### ##### trimming ##### mkdir trimmed while read p; do echo "#######################################"; echo "$p"; echo "#######################################"; trimmomatic PE -phred33 -trimlog trimmed/$p’.log’ raw_wgs/$p’_R1.fastq.gz’ raw_wgs/$p’_R2.fastq.gz’ trimmed/$p’_R1_paired.fastq’ trimmed/$p’_R1_unpaired.fastq’ trimmed/$p’_R2_paired.fastq’ trimmed/$p’_R2_unpaired.fastq’ ILLUMINACLIP:Trimmomatic-0.39/adapters/TruSeq3-PE-2.fa:2:30:10:1:TRUE SLIDINGWINDOW:5:20 ; done < raw_wgs/samples.txt ################################ ##### Quality check of ##### ##### trimmed reads ##### ### run fastQC mkdir trimmed/out_fastQC fastqc trimmed/*.fastq.gz --outdir=trimmed/out_fastQC ### run multiQC mkdir trimmed/out_fastQC/multiQC multiqc trimmed/out_fastQC --outdir trimmed/out_fastQC/multiqc ################################################### ##### chloroplast assembly using getOrganelle ##### ### run getOrganelle while read f; do get_organelle_from_reads.py --max-reads 3E7 -1 trimmed/$f’_R1_paired.fastq’ -2 trimmed/$f’_R2_paired.fastq.gz’ -o getOrganelle/$f -s targets/*_complete.fasta -R 25 -k 21,45,65,85,105 -F embplant_pt -t 8 ; echo ’###############################################################’; echo sample $f finished; echo ’###############################################################’; done < trimmed/samples.txt |
References
- Cooper, G. Chloroplast and Other Plastids. In The Cell: A Molecular Approach; ASM Press: New York, NY, USA, 2007; p. 820. ISBN 978-0-87893-219-1. [Google Scholar]
- Jensen, P.E.; Leister, D. Chloroplast evolution, structure and functions. Prime Rep. 2014, 6, 40. [Google Scholar] [CrossRef]
- Grivet, D.; Heinze, B.; Vendramin, G.G.; Petit, R.J. Genome walking with consensus primers: Application to the large single copy region of chloroplast DNA. Mol. Ecol. Notes 2001, 1, 345–349. [Google Scholar] [CrossRef]
- Henriquez, C.L.; Abdullah; Ahmed, I.; Carlsen, M.M.; Zuluaga, A.; Croat, T.B.; McKain, M.R. Molecular evolution of chloroplast genomes in Monsteroideae (Araceae). Planta 2020, 251, 72. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Su, T.; An, M.-T.; Hu, G.-X. The Complete Chloroplast Genome of the Vulnerable Oreocharis esquirolii (Gesneriaceae): Structural Features, Comparative and Phylogenetic Analysis. Plants 2020, 9, 1692. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.D. Comparative organization of chloroplast genomes. Annu. Rev. Genet. 1985, 19, 325–354. [Google Scholar] [CrossRef]
- Daniell, H.; Lin, C.-S.; Yu, M.; Chang, W.-J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicke, S.; Schneeweiss, G.M.; dePamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Zheng, Y. Dynamic evolution and phylogenomic analysis of the chloroplast genome in Schisandraceae. Sci. Rep. 2018, 8, 9285. [Google Scholar] [CrossRef]
- Gonçalves, D.J.P.; Jansen, R.K.; Ruhlman, T.A.; Mandel, J.R. Under the rug: Abandoning persistent misconceptions that obfuscate organelle evolution. Mol. Phylogenet. Evol. 2020, 151, 106903. [Google Scholar] [CrossRef]
- Jansen, R.K.; Kaittanis, C.; Saski, C.; Lee, S.-B.; Tomkins, J.; Alverson, A.J.; Daniell, H. Phylogenetic analyses of Vitis (Vitaceae) based on complete chloroplast genome sequences: Effects of taxon sampling and phylogenetic methods on resolving relationships among rosids. BMC Evol. Biol. 2006, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.-Q.; Ge, S. The phylogeny of the BEP clade in grasses revisited: Evidence from the whole-genome sequences of chloroplasts. Mol. Phylogenet. Evol. 2012, 62, 573–578. [Google Scholar] [CrossRef]
- Duan, L.; Harris, A.; Mao, L.-Y.; Zhang, Z.-R.; Arslan, E.; Ertuğrul, K.; Loc, P.K.; Hayashi, H.; Wen, J.; Chen, H.-F. Chloroplast Phylogenomics and Biogeography of Liquorice (Leguminosae: Glycyrrhiza). In Prime Archives in Plant Sciences; Vide Leaf: Hyderabad, India, 2020; ISBN 978-81-944664-9-9. [Google Scholar]
- Li, H.; Liu, B.; Davis, C.C.; Yang, Y. Plastome phylogenomics, systematics, and divergence time estimation of the Beilschmiedia group (Lauraceae). Mol. Phylogenet. Evol. 2020, 10, 6901. [Google Scholar] [CrossRef]
- Su, C.; Duan, L.; Liu, P.; Liu, J.; Chang, Z.; Wen, J. Chloroplast phylogenomics and character evolution of eastern Asian Astragalus (Leguminosae): Tackling the phylogenetic structure of the largest genus of flowering plants in Asia. Mol. Phylogenet. Evol. 2020, 10, 7025. [Google Scholar] [CrossRef]
- Cronn, R.; Liston, A.; Parks, M.; Gernandt, D.S.; Shen, R.; Mockler, T. Multiplex sequencing of plant chloroplast genomes using Solexa sequencing-by-synthesis technology. Nucleic Acids Res. 2008, 36, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Parks, M.; Cronn, R.; Liston, A. Increasing phylogenetic resolution at low taxonomic levels using massively parallel sequencing of chloroplast genomes. BMC Biol. 2009, 7, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mardis, E.R. Next-Generation Sequencing Platforms. Annu. Rev. Anal. Chem. 2013, 6, 287–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrett, C.F.; Bacon, C.D.; Antonelli, A.; Cano, Á.; Hofmann, T. An introduction to plant phylogenomics with a focus on palms. Bot. J. Linn. Soc. 2016, 182, 234–255. [Google Scholar] [CrossRef] [Green Version]
- Bleidorn, C. Phylogenomics: An Introduction; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; ISBN 978-3-319-54062-7. [Google Scholar]
- Bakker, F.T.; Bieker, V.C.; Martin, M.D. Editorial: Herbarium Collection-Based Plant Evolutionary Genetics and Genomics. Front. Ecol. Evol. 2020, 8, 603948. [Google Scholar] [CrossRef]
- Zeng, C.-X.; Hollingsworth, P.M.; Yang, J.; He, Z.-S.; Zhang, Z.-R.; Li, D.-Z.; Yang, J.-B. Genome skimming herbarium specimens for DNA barcoding and phylogenomics. Plant Methods 2018, 14, 43. [Google Scholar] [CrossRef] [Green Version]
- Brewer, G.E.; Clarkson, J.J.; Maurin, O.; Zuntini, A.R.; Barber, V.; Bellot, S.; Biggs, N.; Cowan, R.S.; Davies, N.M.J.; Dodsworth, S.; et al. Factors Affecting Targeted Sequencing of 353 Nuclear Genes From Herbarium Specimens Spanning the Diversity of Angiosperms. Front. Plant Sci. 2019, 10, 1102. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, E.A.; Wen, J. Using nuclear gene data for plant phylogenetics: Progress and prospects II. Next-gen approaches. J. Syst. Evol. 2015, 53, 371–379. [Google Scholar] [CrossRef]
- McKain, M.R.; Johnson, M.G.; Uribe-Convers, S.; Eaton, D.; Yang, Y. Practical considerations for plant phylogenomics. Appl. Plant Sci. 2018, 6, e1038. [Google Scholar] [CrossRef]
- Dodsworth, S.; Pokorny, L.; Johnson, M.G.; Kim, J.T.; Maurin, O.; Wickett, N.J.; Forest, F.; Baker, W.J. Hyb-Seq for Flowering Plant Systematics. Trends Plant Sci. 2019, 24, 887–891. [Google Scholar] [CrossRef]
- Straub, S.C.K.; Parks, M.; Weitemier, K.; Fishbein, M.; Cronn, R.C.; Liston, A. Navigating the tip of the genomic iceberg: Next-generation sequencing for plant systematics. Am. J. Bot. 2012, 99, 349–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.; Tang, Y.; Xu, X. The complete chloroplast genome of Sedum emarginatum (Crassulaceae). Mitochondrial DNA Part B 2020, 5, 3082–3083. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Liu, Y.; Xu, C.; Gao, Y.; Yuan, Q.; Suo, Z.; Zhang, Z.; Sun, J. Chloroplast phylogenomic insights into the evolution of Distylium (Hamamelidaceae). BMC Genom. 2021, 22, 293. [Google Scholar] [CrossRef]
- Li, L.; Hu, Y.; He, M.; Zhang, B.; Wu, W.; Cai, P.; Huo, D.; Hong, Y. Comparative chloroplast genomes: Insights into the evolution of the chloroplast genome of Camellia sinensis and the phylogeny of Camellia. BMC Genom. 2021, 22, 138. [Google Scholar] [CrossRef]
- Azarin, K.; Usatov, A.; Makarenko, M.; Khachumov, V.; Gavrilova, V. Comparative analysis of chloroplast genomes of seven perennial Helianthus species. Gene 2021, 774, 145418. [Google Scholar] [CrossRef]
- Liu, K.; Wang, R.; Guo, X.-X.; Zhang, X.-J.; Qu, X.-J.; Fan, S.-J. Comparative and Phylogenetic Analysis of Complete Chloroplast Genomes in Eragrostideae (Chloridoideae, Poaceae). Plants 2021, 10, 109. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, S.; Wang, J.; Shen, H.; Ai, B.; Gao, W.; Zhang, C.; Fei, Q.; Yuan, D.; Wu, Z.; et al. Chloroplast genomes in Populus (Salicaceae): Comparisons from an intensively sampled genus reveal dynamic patterns of evolution. Sci. Rep. 2021, 11, 9471. [Google Scholar] [CrossRef]
- Chen, F.; Dong, W.; Zhang, J.; Guo, X.; Chen, J.; Wang, Z.; Lin, Z.; Tang, H.; Zhang, L. The Sequenced Angiosperm Genomes and Genome Databases. Front. Plant Sci. 2018, 9, 418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Song, Y.; Li, X.; Chen, J.; Mo, L.; Zhang, X.; Lin, Z.; Zhang, L. Genome sequences of horticultural plants: Past, present, and future. Hortic. Res. 2019, 6, 112. [Google Scholar] [CrossRef] [Green Version]
- Qiao, X.; Li, Q.; Yin, H.; Qi, K.; Li, L.; Wang, R.; Zhang, S.; Paterson, A.H. Gene duplication and evolution in recurring polyploidization–diploidization cycles in plants. Genome Biol. 2019, 20, 38. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.D.; Kang, Y.; Kim, C. Application of Genomic Big Data in Plant Breeding: Past, Present, and Future. Plants 2020, 9, 1454. [Google Scholar] [CrossRef] [PubMed]
- Stevens, P.F. Angiosperm Phylogeny Website. Available online: http://www.mobot.org/MOBOT/research/APweb/ (accessed on 29 October 2021).
- Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2016, 44, D67–D72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, E.; Park, S.; Park, J.; Kim, S. The chloroplast genome sequence of Magnolia kobus DC. (Magnoliaceae). Mitochondrial DNA Part B 2018, 3, 342–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wu, T.; Fu, Y.; Hao, J.; Ma, H.; Zhu, Y.; Sima, Y. Complete chloroplast genome sequence of Michelia champaca var. champaca Linnaeus, an ornamental tree species of Magnoliaceae. Mitochondrial DNA Part B 2020, 5, 2839–2841. [Google Scholar] [CrossRef] [PubMed]
- Sima, Y.; Wu, T.; Fu, Y.; Hao, J.; Chen, S. Complete chloroplast genome sequence of Lirianthe coco (Loureiro) N. H. Xia & C. Y. Wu (Magnoliaceae), a popular ornamental species. Mitochondrial DNA Part B 2020, 5, 2410–2412. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, B.; Nie, Z.; Chen, H.; Chen, F.; Figlar, R.B.; Wen, J. Major clades and a revised classification of Magnolia and Magnoliaceae based on whole plastid genome sequences via genome skimming. J. Syst. Evol. 2020, 58, 673–695. [Google Scholar] [CrossRef]
- Xu, X.; Wu, S.; Xia, J.; Yan, B. The complete chloroplast genome of Magnolia delavayi, a threatened species endemic to Southwest China. Mitochondrial DNA Part B 2020, 5, 2734–2735. [Google Scholar] [CrossRef]
- Zhang, M.; Guo, S.; Yanpeng, Y.; Zhou, L.; Bo, R.; Li, W.; Shi, X.; Feixia, H.; Cheng, P.; Xianmei, Y.; et al. Chloroplast genome and historical biogeography of the three Magnolias. Res. Square 2021, 13, 21. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Azuma, H.; García-Franco, J.G.; Rico-Gray, V.; Thien, L.B. Molecular phylogeny of the Magnoliaceae: The biogeography of tropical and temperate disjunctions. Am. J. Bot. 2001, 88, 2275–2285. [Google Scholar] [CrossRef] [PubMed]
- Dandy, J.E. The Genera of Magnolieae. Bull. Misc. Inform. Kew 1927, 1927, 257. [Google Scholar] [CrossRef]
- Figlar, R.B.; Nooteboom, H.P. Notes on Magnoliaceae IV. Blum.-J. Plant. Tax. Plant. Geog. 2004, 49, 87–100. [Google Scholar] [CrossRef]
- Figlar, R.B. A New Classification for Magnolia. In Rhododendrons with Camellias and Magnolias; The Royal Horticulture Society: London, UK, 2006; pp. 69–82. [Google Scholar]
- Figlar, R.B. A Brief Taxonomic History of Magnolia. Available online: https://www.magnoliasociety.org/ClassificationArticle (accessed on 4 September 2021).
- Magnoliaceae in Flora of China @ Efloras.Org. Available online: http://efloras.org/florataxon.aspx?flora_id=2&taxon_id=10530 (accessed on 29 October 2021).
- Sima, Y.-K.; Lu, S.-G. A New System for the Family Magnoliaceae. In Proceedings of the Second International Symposium on the Family Magnoliaceae, Guangzhou, China, 5–9 May 2009; Huazhong University Science & Technology Press: Wuhan, China, 2012; pp. 55–71. [Google Scholar]
- Nie, Z.-L.; Wen, J.; Azuma, H.; Qiu, Y.-L.; Sun, H.; Meng, Y.; Sun, W.-B.; Zimmer, E.A. Phylogenetic and biogeographic complexity of Magnoliaceae in the Northern Hemisphere inferred from three nuclear data sets. Mol. Phylogenet. Evol. 2008, 48, 1027–1040. [Google Scholar] [CrossRef] [PubMed]
- Veltjen, E.; Testé, E.; Palmarola-Bejarano, A.; Asselman, P.; Hernández-Rodríguez, M.; González-Torres, L.R.; Chatrou, L.W.; Goetghebeur, P.; Larridon, I.; Samain, M.-S. The evolutionary history of the caribbean magnolias (Magnoliaceae): Testing species delimitations and biogeographical hypotheses using molecular data. Mol. Phylogenet. Evol. 2022, in press. [Google Scholar] [CrossRef]
- Linnaeus, C. Species Plantarum; Salvius: Stockholm, Sweden, 1753. [Google Scholar]
- Kim, S.; Park, C.-W.; Kim, Y.-D.; Suh, Y. Phylogenetic relationships in family Magnoliaceae inferred from ndhF sequences. Am. J. Bot. 2001, 88, 717–728. [Google Scholar] [CrossRef]
- Arroyo, F.; Serna-González, M. Two new species of Magnolia (Magnoliaceae) from Peru. Phytotaxa 2020, 471, 54–60. [Google Scholar] [CrossRef]
- De Azevedo, C.O.; Marinho, L.C.; Machado, A.F.P.; Arroyo, F.; Vázquez-García, J.A. Magnolia brasiliensis (Magnoliaceae), a new species and new record for the Northeastern region of Brazil. Brittonia 2018, 70, 306–311. [Google Scholar] [CrossRef]
- Pérez, Á.J.; Arroyo, F.; Neill, D.A.; Vázquez-García, J.A. Magnolia chiguila and M. mashpi (Magnoliaceae): Two new species and a new subsection (Chocotalauma, sect. Talauma) from the Chocó biogeographic region of Colombia and Ecuador. Phytotaxa 2016, 286, 267. [Google Scholar] [CrossRef]
- Serrano, M.J.; Grajeda-Estrada, R.; Villalobos, A.; Álvarez-Ruano, M.R.; Vázquez-García, J.A. Magnolia poqomchi, a new species of subsection Magnolia (Magnoliaceae) from San Cristóbal Verapaz, Alta Verapaz, Guatemala. Phytotaxa 2020, 454, 231–243. [Google Scholar] [CrossRef]
- Vázquez-García, J.A.; Pérez-Farrera, M.Á.; Martínez-Camilo, R.; Muñiz-Castro, M.Á.; Martínez-Meléndez, N. Magnolia lacandonica (subsection Talauma, Magnoliaceae), a new rainforest species from Chiapas, Mexico. Phytotaxa 2013, 79, 30–36. [Google Scholar] [CrossRef]
- Vázquez-García, J.A.; Gómez-Domínguez, H.; López-Cruz, A. Magnolia perezfarrerae, una especie nueva y una clave para las especies mexicanas de Magnolia (sección Talauma, subsección Talauma, Magnoliaceae). Bot. Sci. 2013, 91, 417–425. [Google Scholar] [CrossRef]
- Muñiz-Castro, M.Á.; Castro-Félix, P.; Carranza-Aranda, A.S.; Vázquez-García, J.A.; Santerre, A. Population genetics, species boundaries, and conservation in the Magnolia pacifica species complex along a continentality and moisture gradient in western Mexico. Bot. Sci. 2020, 25, 51. [Google Scholar] [CrossRef]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 29 October 2021).
- Qiu, Y.-L.; Chase, M.W.; Les, D.H.; Parks, C.R. Molecular Phylogenetics of the Magnoliidae: Cladistic Analyses of Nucleotide Sequences of the Plastid Gene rbcL. Ann. Mo. Bot. Gard. 1993, 80, 587–606. [Google Scholar] [CrossRef]
- Qiu, Y.-L.; Parks, C.R.; Chase, M.W. Molecular divergence in the eastern Asia-eastern North America disjunct section Rytidospermum of Magnolia (Magnoliaceae). Am. J. Bot. 1995, 82, 1589–1598. [Google Scholar] [CrossRef]
- Qiu, Y.-L.; Chase, M.W.; Parks, C.R. A chloroplast DNA Phylogenetic study of the eastern Asia-eastern North America disjunct section Rytidospermum of Magnolia (Magnoliaceae). Am. J. Bot. 1995, 82, 1582–1588. [Google Scholar] [CrossRef]
- Kim, S.; Suh, Y. Phylogeny of Magnoliaceae based on ten chloroplast DNA regions. J. Plant Biol. 2013, 56, 290–305. [Google Scholar] [CrossRef]
- Gao, F.; Chen, C.; Arab, D.A.; Du, Z.; He, Y.; Ho, S.Y.W. EasyCodeML: A visual tool for analysis of selection using CodeML. Ecol. Evol. 2019, 9, 3891–3898. [Google Scholar] [CrossRef] [Green Version]
- Freudenthal, J.A.; Pfaff, S.; Terhoeven, N.; Korte, A.; Ankenbrand, M.J.; Förster, F. A systematic comparison of chloroplast genome assembly tools. Genome Biol. 2020, 21, 254. [Google Scholar] [CrossRef]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Shi, L.; Zhu, Y.; Chen, H.; Zhang, J.; Lin, X.; Guan, X. CpGAVAS, an integrated web server for the annotation, visualization, analysis, and GenBank submission of completely sequenced chloroplast genome sequences. BMC Genom. 2012, 13, 715. [Google Scholar] [CrossRef] [Green Version]
- Guyeux, C.; Charr, J.-C.; Tran, H.T.M.; Furtado, A.; Henry, R.J.; Crouzillat, D.; Guyot, R.; Hamon, P. Evaluation of chloroplast genome annotation tools and application to analysis of the evolution of coffee species. PLoS ONE 2019, 14, e0216347. [Google Scholar] [CrossRef] [PubMed]
- Kahraman, K.; Lucas, S.J. Comparison of different annotation tools for characterization of the complete chloroplast genome of Corylus avellana cv Tombul. BMC Genom. 2019, 20, 874. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Shao, J.; Zhang, H.; Jiang, M.; Huang, L.; Zhang, Z.; Yang, D.; He, M.; Ronaghi, M.; Luo, X.; et al. Sequencing and Analysis of Strobilanthes cusia (Nees) Kuntze Chloroplast Genome Revealed the Rare Simultaneous Contraction and Expansion of the Inverted Repeat Region in Angiosperm. Front. Plant Sci. 2018, 9, 324. [Google Scholar] [CrossRef] [Green Version]
- Abdullah; Henriquez, C.L.; Mehmood, F.; Carlsen, M.M.; Islam, M.; Waheed, M.T.; Poczai, P.; Croat, T.B.; Ahmed, I. Complete Chloroplast Genomes of Anthurium huixtlense and Pothos scandens (Pothoideae, Araceae): Unique Inverted Repeat Expansion and Contraction Affect Rate of Evolution. J. Mol. Evol. 2020, 88, 562–574. [Google Scholar] [CrossRef]
- Frailey, D.C.; Chaluvadi, S.R.; Vaughn, J.N.; Coatney, C.G.; Bennetzen, J.L. Gene loss and genome rearrangement in the plastids of five Hemiparasites in the family Orobanchaceae. BMC Plant Biol. 2018, 18, 30. [Google Scholar] [CrossRef] [Green Version]
- Cauz-Santos, L.A.; da Costa, Z.P.; Callot, C.; Cauet, S.; Zucchi, M.I.; Bergès, H.; van den Berg, C.; Vieira, M.L.C. A Repertory of Rearrangements and the Loss of an Inverted Repeat Region in Passiflora Chloroplast Genomes. Genome Biol. Evol. 2020, 12, 1841–1857. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Z.; Yang, J.; Lv, G. Complete chloroplast genome of seven Fritillaria species, variable DNA markers identification and phylogenetic relationships within the genus. PLoS ONE 2018, 13, e0194613. [Google Scholar] [CrossRef] [Green Version]
- Xiao-Ming, Z.; Junrui, W.; Li, F.; Sha, L.; Hongbo, P.; Lan, Q.; Jing, L.; Yan, S.; Weihua, Q.; Lifang, Z.; et al. Inferring the evolutionary mechanism of the chloroplast genome size by comparing whole-chloroplast genome sequences in seed plants. Sci. Rep. 2017, 7, 1555. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-L.; Song, Y.; Ni, J.; Yao, X.; Tan, Y.-H.; Xu, Z.-F. Comparative chloroplast genomics and phylogenetics of nine Lindera species (Lauraceae). Sci. Rep. 2018, 8, 8844. [Google Scholar] [CrossRef] [Green Version]
- Han, E.-K.; Cho, W.-B.; Choi, G.; Lee, J.-H. The complete chloroplast genome of Sarcandra glabra (Chloranthaceae): A perianthless basal angiosperm. Mitochondrial DNA Part B 2018, 3, 661–662. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Gichira, A.W.; Li, Z.; Nzei, J.M.; Guo, Y.; Wang, Q.; Chen, J. Intergeneric Relationships within the Early-Diverging Angiosperm Family Nymphaeaceae Based on Chloroplast Phylogenomics. Int. J. Mol. Sci. 2018, 19, 3780. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, K.H.; Morden, C.W.; Palmer, J.D. Function and evolution of a minimal plastid genome from a nonphotosynthetic parasitic plant. Proc. Natl. Acad. Sci. USA 1992, 89, 10648–10652. [Google Scholar] [CrossRef] [Green Version]
- Greiner, S.; Wang, X.; Rauwolf, U.; Silber, M.V.; Mayer, K.; Meurer, J.; Haberer, G.; Herrmann, R.G. The complete nucleotide sequences of the five genetically distinct plastid genomes of Oenothera, subsection Oenothera: I. Sequence evaluation and plastome evolution. Nucleic Acids Res. 2008, 36, 2366–2378. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-P.; Huang, J.-P.; Wu, C.-S.; Hsu, C.-Y.; Chaw, S.-M. Comparative Chloroplast Genomics Reveals the Evolution of Pinaceae Genera and Subfamilies. Genome Biol. Evol. 2010, 2, 504–517. [Google Scholar] [CrossRef] [PubMed]
- Chris Blazier, J.; Guisinger, M.M.; Jansen, R.K. Recent loss of plastid-encoded ndh genes within Erodium (Geraniaceae). Plant Mol. Biol. 2011, 76, 263–272. [Google Scholar] [CrossRef]
- Zhu, A.; Guo, W.; Gupta, S.; Fan, W.; Mower, J.P. Evolutionary dynamics of the plastid inverted repeat: The effects of expansion, contraction, and loss on substitution rates. New Phytol. 2016, 209, 1747–1756. [Google Scholar] [CrossRef] [Green Version]
- Guisinger, M.M.; Kuehl, J.V.; Boore, J.L.; Jansen, R.K. Extreme Reconfiguration of Plastid Genomes in the Angiosperm Family Geraniaceae: Rearrangements, Repeats, and Codon Usage. Mol. Biol. Evol. 2011, 28, 583–600. [Google Scholar] [CrossRef] [Green Version]
- Abdullah; Mehmood, F.; Rahim, A.; Heidari, P.; Ahmed, I.; Poczai, P. Comparative plastome analysis of Blumea, with implications for genome evolution and phylogeny of Asteroideae. Ecol. Evol. 2021, 3, 7614. [Google Scholar] [CrossRef]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. The chloroplast genome sequence of bittersweet (Solanum dulcamara): Plastid genome structure evolution in Solanaceae. PLoS ONE 2018, 13, e0196069. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; He, J.; Ding, C.; Lyu, R.; Pei, L.; Cheng, J.; Xie, L. Comparative Analysis of Complete Chloroplast Genomes of Anemoclema, Anemone, Pulsatilla, and Hepatica Revealing Structural Variations Among Genera in Tribe Anemoneae (Ranunculaceae). Front. Plant Sci. 2018, 1, 97. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zuo, Y.; Zhu, X.; Liao, S.; Ma, J. Complete Chloroplast Genomes and Comparative Analysis of Sequences Evolution among Seven Aristolochia (Aristolochiaceae) Medicinal Species. Int. J. Mol. Sci. 2019, 20, 1045. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Sun, R.; Liu, H.; Chang, L.; Li, S.; Zhao, M.; Shennan, C.; Lei, J.; Dong, J.; Zhong, C.; et al. Complete chloroplast genome sequencing of ten wild Fragaria species in China provides evidence for phylogenetic evolution of Fragaria. Genomics 2021, 113, 1170–1179. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-F.; Landis, J.B.; Wang, H.-X.; Zhu, Z.-X.; Wang, H.-F. Comparative analysis of chloroplast genome structure and molecular dating in Myrtales. BMC Plant Biol. 2021, 21, 219. [Google Scholar] [CrossRef]
- Tian, X.; Guo, J.; Zhou, X.; Ma, K.; Ma, Y.; Shi, T.; Shi, Y. Comparative and Evolutionary Analyses on the Complete Plastomes of Five Kalanchoe Horticultural Plants. Front. Plant Sci. 2021, 12, 651. [Google Scholar] [CrossRef]
- Raman, G.; Park, K.T.; Kim, J.-H.; Park, S. Characteristics of the completed chloroplast genome sequence of Xanthium spinosum: Comparative analyses, identification of mutational hotspots and phylogenetic implications. BMC Genom. 2020, 21, 855. [Google Scholar] [CrossRef]
- Azuma, H.; Thien, L.B.; Kawano, S. Molecular Phylogeny of Magnolia (Magnoliaceae) Inferred from cpDNA Sequences and Evolutionary Divergence of the Floral Scents. J. Plant. Res. 1999, 112, 291–306. [Google Scholar] [CrossRef]
- Missouri Botanical Garden Tropicos. Available online: https://www.tropicos.org/home (accessed on 10 November 2021).
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Larridon, I.; Walter, H.E.; Guerrero, P.C.; Duarte, M.; Cisternas, M.A.; Hernández, C.P.; Bauters, K.; Asselman, P.; Goetghebeur, P.; Samain, M.-S. An integrative approach to understanding the evolution and diversity of Copiapoa (Cactaceae), a threatened endemic Chilean genus from the Atacama Desert. Am. J. Bot. 2015, 102, 1506–1520. [Google Scholar] [CrossRef] [Green Version]
- Thiers, B. Index Herbariorum: A global directory of public herbaria and associated staff; New York Botanical Garden: New York, NY, USA, 2019. [Google Scholar]
- Reserva Natural “El Refugio”. Available online: http://elrefugionatura.jimdofree.com/about-1/ (accessed on 7 December 2021).
- Andrews, S. FastQC. A Quality Control Tool for High Throughput Sequence Data. 2019. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 20 November 2021).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; dePamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [Green Version]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA Genes in Genomic Sequences. Methods Mol. Biol. 2019, 1962, 1–14. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [Green Version]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) version 1.3.1: Expanded toolkit for the graphical visualization of organellar genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Brudno, M.; Malde, S.; Poliakov, A.; Do, C.B.; Couronne, O.; Dubchak, I.; Batzoglou, S. Glocal alignment: Finding rearrangements during alignment. Bioinformatics 2003, 19, i54–i62. [Google Scholar] [CrossRef] [Green Version]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple Alignment of Conserved Genomic Sequence with Rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. IRscope: An online program to visualize the junction sites of chloroplast genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]



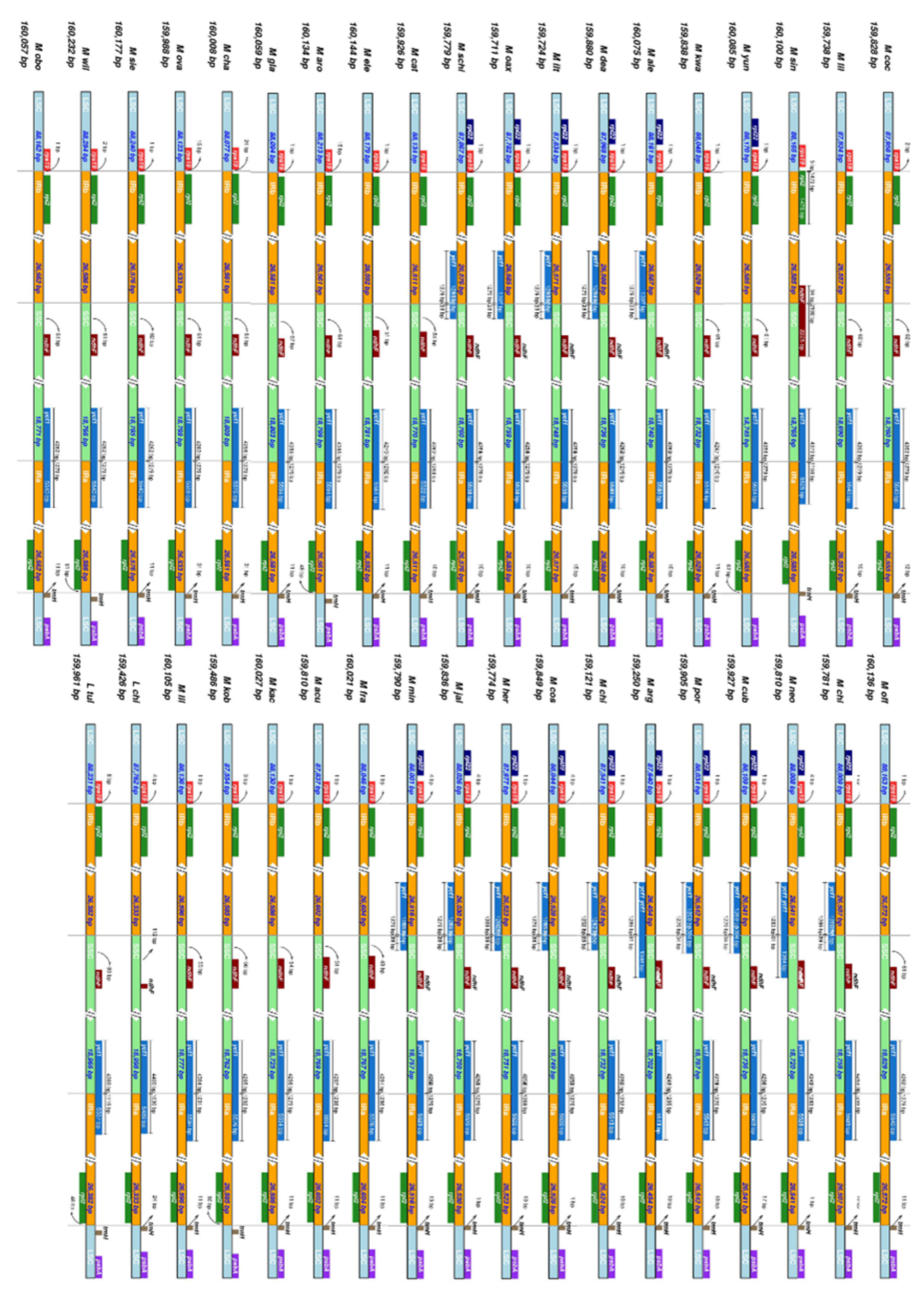
| Genus | Section (Subsection) | Species | Plastome Sequence Length | Assembly Coverage | GC Content | Gene Content | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | LSC | SSC | IR | CDS | tRNAs | rRNAs | |||||
| Liriodendron | NA | L. chinense | 159,426 | 87,762 | 18,998 | 26,333 | - | 0.39 | 92 | 45 | 8 |
| L. tulipifera | 159,961 | 88,231 | 18,966 | 26,382 | - | 0.39 | 92 | 45 | 8 | ||
| Magnolia | Auriculata | M. fraseri | 160,021 | 88,047 | 18,767 | 26,603 | - | 0.39 | 92 | 45 | 8 |
| Gwillimia (Blumiana) | M. liliifera | 159,738 | 87,934 | 18,690 | 26,557 | - | 0.39 | 92 | 45 | 8 | |
| Gwillimia (Gwillimia) | M. coco | 159,828 | 87,958 | 18,760 | 26,555 | - | 0.39 | 92 | 45 | 8 | |
| Gynopodium | M. kachirachirai | 160,027 | 88,130 | 18,725 | 26,586 | - | 0.39 | 92 | 45 | 8 | |
| M. yunnanensis | 160,085 | 88,170 | 18,745 | 26,585 | - | 0.39 | 92 | 45 | 8 | ||
| Kmeria | M. kwangsiensis | 159,838 | 88,048 | 18,732 | 26,529 | - | 0.39 | 92 | 45 | 8 | |
| Macrophylla | M. alejandrae | 160,075 | 88,161 | 18,740 | 26,587 | 65.9 | 0.39 | 92 | 45 | 8 | |
| M. dealbata | 159,880 | 87,968 | 18,736 | 26,588 | 130.2 | 0.39 | 92 | 45 | 8 | ||
| Magnolia | M. iltisiana | 159,724 | 87,834 | 18,748 | 26,571 | 78.4 | 0.39 | 92 | 45 | 8 | |
| M. oaxacensis | 159,711 | 87,782 | 18,759 | 26,585 | 48.5 | 0.39 | 92 | 45 | 8 | ||
| M. schiedeana | 159,779 | 87,867 | 18,760 | 26,576 | 74.1 | 0.39 | 92 | 45 | 8 | ||
| Manglietia | M. aromatica | 160,134 | 88,213 | 18,799 | 26,561 | - | 0.39 | 92 | 45 | 8 | |
| M. glaucifolia1 | 160,059 | 88,094 | 18,803 | 26,581 | - | 0.39 | 92 | 45 | 8 | ||
| Manglietiastrum | M. sinica | 160,100 | 88,294 | 18,765 | 26,585 | - | 0.39 | 92 | 45 | 8 | |
| Michelia (Aromadendron) | M. elegans | 160,144 | 88,179 | 18,781 | 26,592 | - | 0.39 | 92 | 45 | 8 | |
| Michelia (Elmerrillia) | M. ovalis2 | 159,988 | 88,123 | 18,799 | 26,533 | - | 0.39 | 92 | 45 | 8 | |
| Michelia (Maingola) | M. catchcartii | 159,926 | 88,134 | 18,770 | 26,511 | - | 0.39 | 92 | 45 | 8 | |
| Michelia (Michelia) | M. champaca var. champaca | 160,008 | 88,077 | 18,809 | 26,561 | - | 0.39 | 92 | 45 | 8 | |
| Rhytidospermum (Oyama) | M. sieboldii | 160,177 | 88,240 | 18,785 | 26,576 | - | 0.39 | 92 | 45 | 8 | |
| M. wilsonii | 160,232 | 88,294 | 18,766 | 26,586 | - | 0.39 | 92 | 45 | 8 | ||
| Rhytidospermum (Rhytidospermum) | M. obovata | 160,057 | 88,162 | 18,771 | 26,562 | - | 0.39 | 92 | 45 | 8 | |
| M. officinalis | 160,136 | 88,163 | 18,829 | 26,572 | - | 0.39 | 92 | 45 | 8 | ||
| Talauma (Chocotalauma) | M. chiguila | 159,781 | 88,009 | 18,758 | 26,507 | 37.5 | 0.39 | 92 | 45 | 8 | |
| Talauma (Cubenses) | M. cubensis subsp. cubensis | 159,927 | 88,009 | 18,736 | 26,541 | 19.8 | 0.39 | 92 | 45 | 8 | |
| M. portoricensis | 159,905 | 88,034 | 18,787 | 26,542 | 78.4 | 0.39 | 92 | 45 | 8 | ||
| Talauma (Dugandiodendron) | M. argyrothricha | 159,250 | 87,640 | 18,702 | 26,454 | 195.0 | 0.39 | 92 | 45 | 8 | |
| M. chimantensis | 159,121 | 87,541 | 18,732 | 26,424 | 78.0 | 0.39 | 92 | 45 | 8 | ||
| M. neomagnifolia | 159,810 | 88,008 | 18,720 | 26,541 | 73.2 | 0.39 | 92 | 45 | 8 | ||
| Talauma (Talauma) | M. costaricensis | 159,849 | 88,044 | 18,749 | 26,528 | 88.3 | 0.39 | 92 | 45 | 8 | |
| M. hernandezii | 159,774 | 87,977 | 18,751 | 26,523 | 56.5 | 0.39 | 92 | 45 | 8 | ||
| M. jaliscana | 159,836 | 88,026 | 18,750 | 26,530 | 82.0 | 0.39 | 92 | 45 | 8 | ||
| M. minor | 159,790 | 88,001 | 18,757 | 26,516 | 66.4 | 0.39 | 92 | 45 | 8 | ||
| Yulania (Tulipastrum) | M. acuminata | 159,810 | 87,837 | 18,769 | 26,602 | - | 0.39 | 92 | 45 | 8 | |
| Yulania (Yulania) | M. kobus | 159,486 | 87,554 | 18,762 | 26,585 | - | 0.39 | 92 | 45 | 8 | |
| M. liliiflora | 160,105 | 88,136 | 18,777 | 26,596 | - | 0.39 | 92 | 45 | 8 | ||
| Minimum | 159,121 | 87,541 | 18,690 | 26,333 | 19.8 | 0.39 | 92 | 45 | 8 | ||
| Maximum | 160,232 | 88,294 | 18,998 | 26,603 | 195.0 | 0.39 | 92 | 45 | 8 | ||
| Mean | 159,878 | 88,018 | 18,772 | 26,544 | 78.2 | 0.39 | 92 | 45 | 8 | ||
| Category | Gene Group | Gene Names |
|---|---|---|
| Photosynthesis | ATP synthase | atpA, atpB, atpE, atpF, atpH, and atpI |
| Cytochrome complex | petA, petB, petD, petG, petL, and petN | |
| NADH dehydrogenase | ndhA, ndhB, ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, and ndhK | |
| Photosystem I | pafI, pafII, pbf1, psaA, psaB, psaC, psaI, and psaJ | |
| Photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbG, psbH, psbI, psbJ, psbK, psbL, psbM, psbT, and psbZ | |
| Rubisco large subunit | rbcL | |
| rRNAs | rrn4.5, rrn5, rrn16, rrn23 | |
| Self-replication | Ribosomal proteins (LSU) | rpl2, rpl14, rpl16, rpl18, rpl20, rpl22, rpl23, rpl32, rpl33, and rpl36 |
| Ribosomal proteins (SSU) | rps2, rps3, rps4, rps7, rps8, rps11, rps12, rps14, rps15, rps16, rps18, and rps19 | |
| RNA polymerase | rpoA, rpoB, rpoC1, and rpoC2 | |
| tRNAs | trnA-UGC, trnC-ACA, trnC-GCA, trnD-GUC, trnE-UUC, trnF-GAA, trnfM-CAU, trnG-GCC, trnG-UCC, trnH LSC, trnH-GUG, trnI-CAU, trnI-GAU, trnK-UUU, trnL-CAA, trnL-UAA, trnL-UAG, trnM-CAU, trnN-GUU, trnP-GGG, trnP-UGG, trnQ-UUG, trnR-ACG, trnR-UCU, trnS-CGA, trnS-GCU, trnS-GGA, trnS-UGA, trnT-GGU, trnT-UGU, trnV-GAC, trnV-UAC, trnW-CCA, and trnY-GUA | |
| Other | Conserved open reading frames | ycf1, ycf2, and ycf15 |
| Cytochrome c synthesis | ccsA | |
| Membrane protein | cemA | |
| Protease | clpP1 | |
| RNA processing | matK | |
| Subunit of acetyl-CoA carboxylase | accD | |
| Translational initiation | infA |
| Section (Subsection) | Species | Country | Collection | Voucher |
|---|---|---|---|---|
| Macrophylla | M. alejandrae García-Mor. and Iamonico | Mexico | XAL | M. Mata 1188b |
| M. dealbata Zucc. | Mexico | XAL | M. Mata 0866a | |
| Magnolia | M. iltisiana A. Vázquez | Mexico | XAL | S. Cházaro B. and M. Rodríguez 8590 |
| M. oaxacensis A. Vázquez | Mexico | IEB, MEXU | M.S. Samain and E. Martínez 2019-019 | |
| M. schiedeana Schltdl. | Mexico | IEB, MEXU | F. Aldaba 187 | |
| Talauma (Chocotalauma) | M. chiguila F. Arroyo, Á.J. Pérez and A. Vázquez | Ecuador | ECUAMZ | F. Arroyo and Á.J. Pérez 286 |
| Talauma (Cubenses) | M. cubensis subsp. cubensis Urb. | Cuba | HAJB | A. Palmarola et al. HFC-89195 |
| M. portoricensis Bello | Puerto Rico | GENT | E. Veltjen et al. 2016-033 | |
| Talauma (Dugandiodendron) | M. argyrothricha (G. Lozano C.) Govaerts | Colombia | “El Refugio” Natural Reserve | NA |
| M. chimantensis Steyerm. and Maguire | Venezuela | K | J. Steyermark 1191 | |
| M. neomagnifolia I.M. Turner | Colombia | “El Refugio” Natural Reserve | NA | |
| Talauma (Talauma) | M. costaricensis A. Vázquez | Costa Rica | USJ | J.E. Jiménez 4622 |
| M. hernandezii (G. Lozano C.) Govaerts | Colombia | K | J. Hernández et al. 1001 | |
| M. jaliscana A. Vázquez and R. Guzmán | Mexico | IBUG | J.A. Vázquez García et al. 9335 | |
| M. minor (Urb.) Govaerts | Cuba | HAJB | B. Falcón HFC88953 |
| Genus | Section (Subsection) | Species | NCBI reference |
|---|---|---|---|
| Liriodendron | NA | L. chinense (Hemsl.) Sarg. | MN990597 |
| L. tulipifera L. | MN990625 | ||
| Magnolia | Auriculata | M. fraseri Walter | MN990599 |
| Gwillimia (Blumiana) | M. coco (Lour.) DC. | MN990612 | |
| Gwillimia (Gwillimia) | M. liliifera (L.) Baill. | MN990610 | |
| Gynopodium | M. kachirachirai (Kaneh. and Yamam.) Dandy | MN990641 | |
| M. yunnanensis (H.H. Hu) Noot. | KF753638 | ||
| Kmeria | M. kwangsiensis Figlar and Noot. | MN990593 | |
| Manglietia | M. aromatica (Dandy) V.S. Kumar | MN990576 | |
| M. glaucifolia Noot. 1 | MF990565 | ||
| Manglietiastrum | M. sinica (Y.W. Law) Noot. | MN990584 | |
| Michelia (Aromadendron) | M. elegans (Blume) H. Keng | MN990630 | |
| Michelia (Elmerrillia) | M. ovalis (Miq.) Figlar 2 | MN990602 | |
| Michelia (Maingola) | M. cathcartii (Hook. f. and Thomson) Noot. | MN990570 | |
| Michelia (Michelia) | M. champaca var. champaca (L.) Baill. ex Pierre | MT269873 | |
| Rhytidospermum (Oyama) | M. sieboldii K. Koch | MN990583 | |
| M. wilsonii (Finet and Gagnep.) Rehder | MN990621 | ||
| Rhytidospermum (Rhytidospermum) | M. obovata Thunb. | MN990571 | |
| M. officinalis Rehder and E.H. Wilson | MN990572 | ||
| Yulania (Tulipastrum) | M. acuminata (L.) L. | MN990595 | |
| Yulania (Yulania) | M. kobus DC. | MN990635 | |
| M. liliiflora Desr. | MN990588 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guzmán-Díaz, S.; Núñez, F.A.A.; Veltjen, E.; Asselman, P.; Larridon, I.; Samain, M.-S. Comparison of Magnoliaceae Plastomes: Adding Neotropical Magnolia to the Discussion. Plants 2022, 11, 448. https://doi.org/10.3390/plants11030448
Guzmán-Díaz S, Núñez FAA, Veltjen E, Asselman P, Larridon I, Samain M-S. Comparison of Magnoliaceae Plastomes: Adding Neotropical Magnolia to the Discussion. Plants. 2022; 11(3):448. https://doi.org/10.3390/plants11030448
Chicago/Turabian StyleGuzmán-Díaz, Salvador, Fabián Augusto Aldaba Núñez, Emily Veltjen, Pieter Asselman, Isabel Larridon, and Marie-Stéphanie Samain. 2022. "Comparison of Magnoliaceae Plastomes: Adding Neotropical Magnolia to the Discussion" Plants 11, no. 3: 448. https://doi.org/10.3390/plants11030448
APA StyleGuzmán-Díaz, S., Núñez, F. A. A., Veltjen, E., Asselman, P., Larridon, I., & Samain, M.-S. (2022). Comparison of Magnoliaceae Plastomes: Adding Neotropical Magnolia to the Discussion. Plants, 11(3), 448. https://doi.org/10.3390/plants11030448