
Optimizing Nitrogen and Seed Rate Combination for Improving Grain Yield and Nitrogen Uptake Efficiency in Winter Wheat

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Physiological Traits
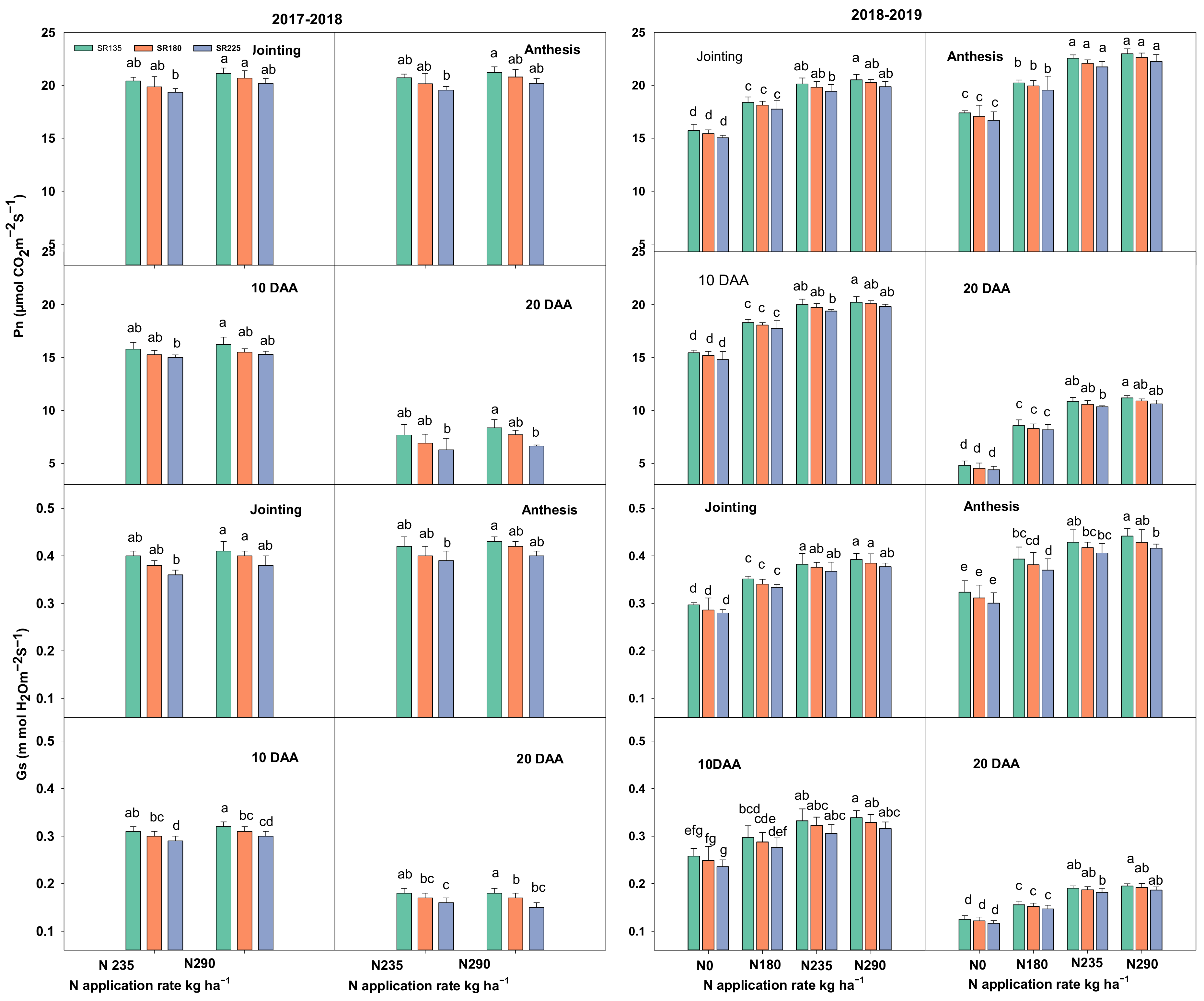
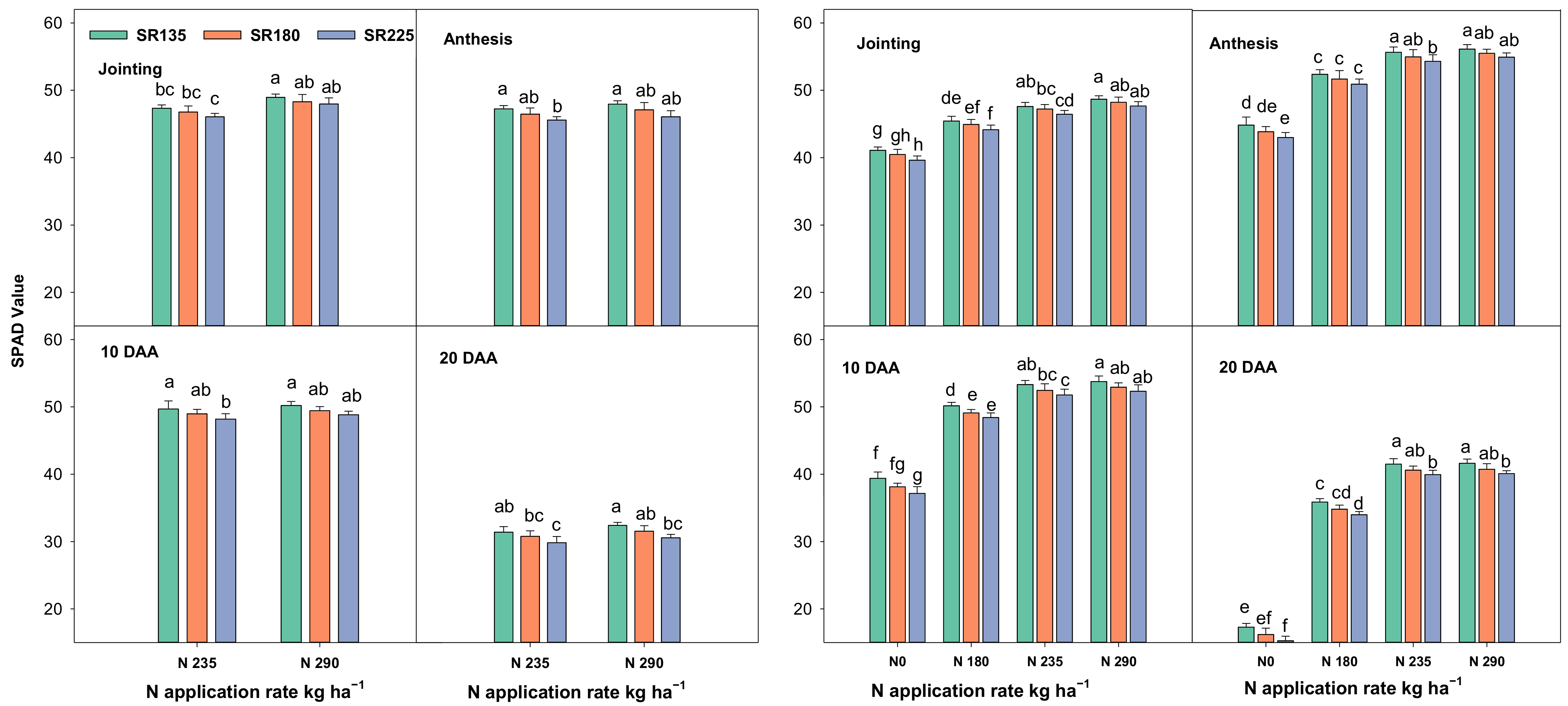
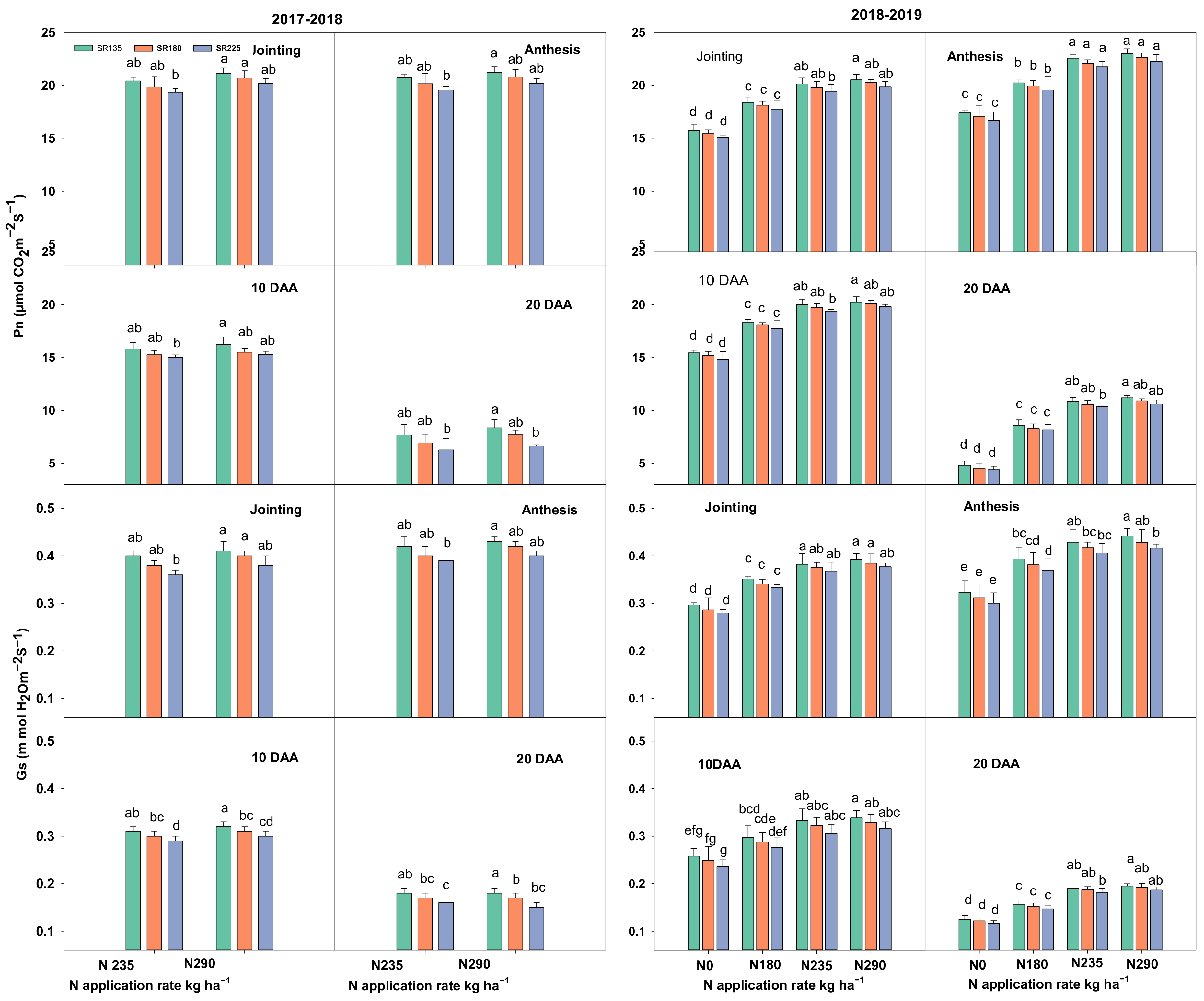
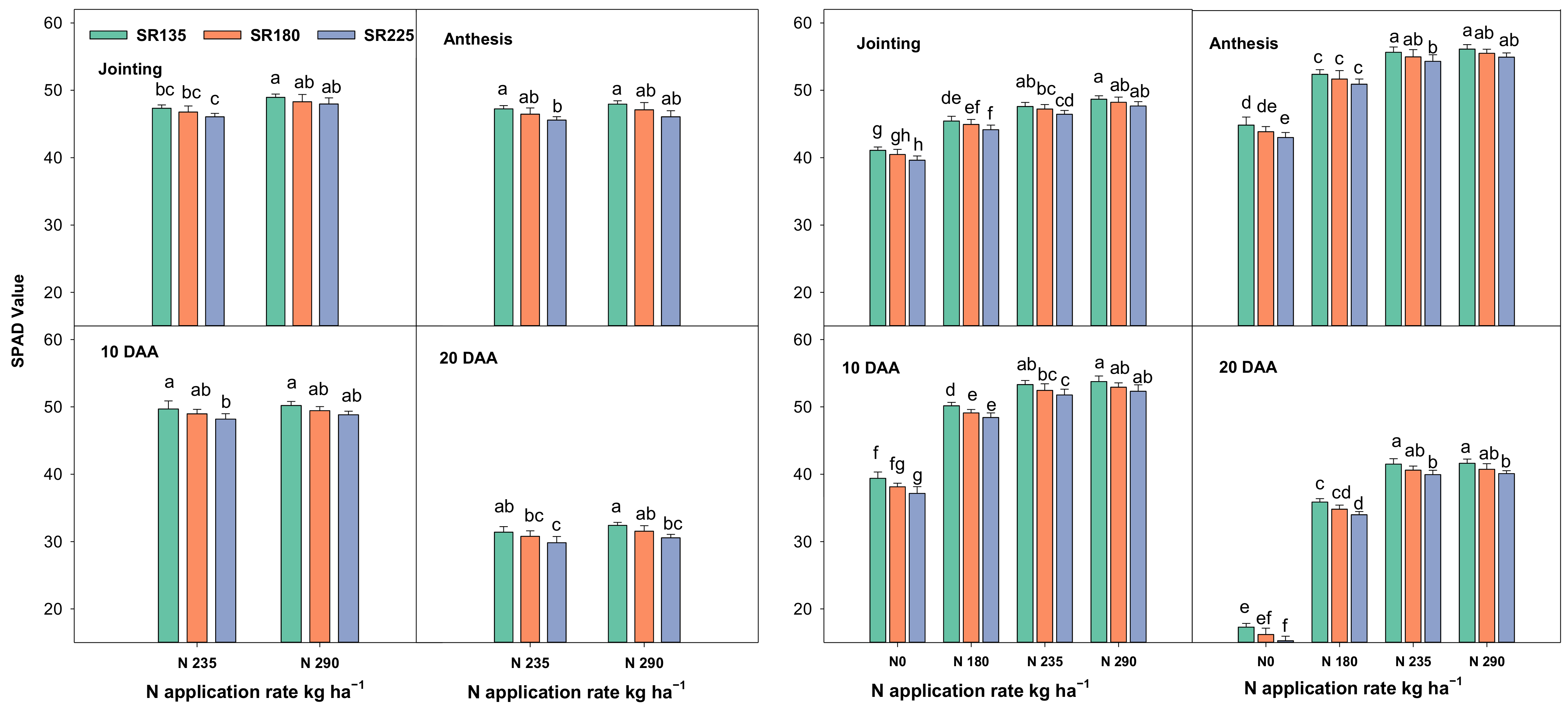
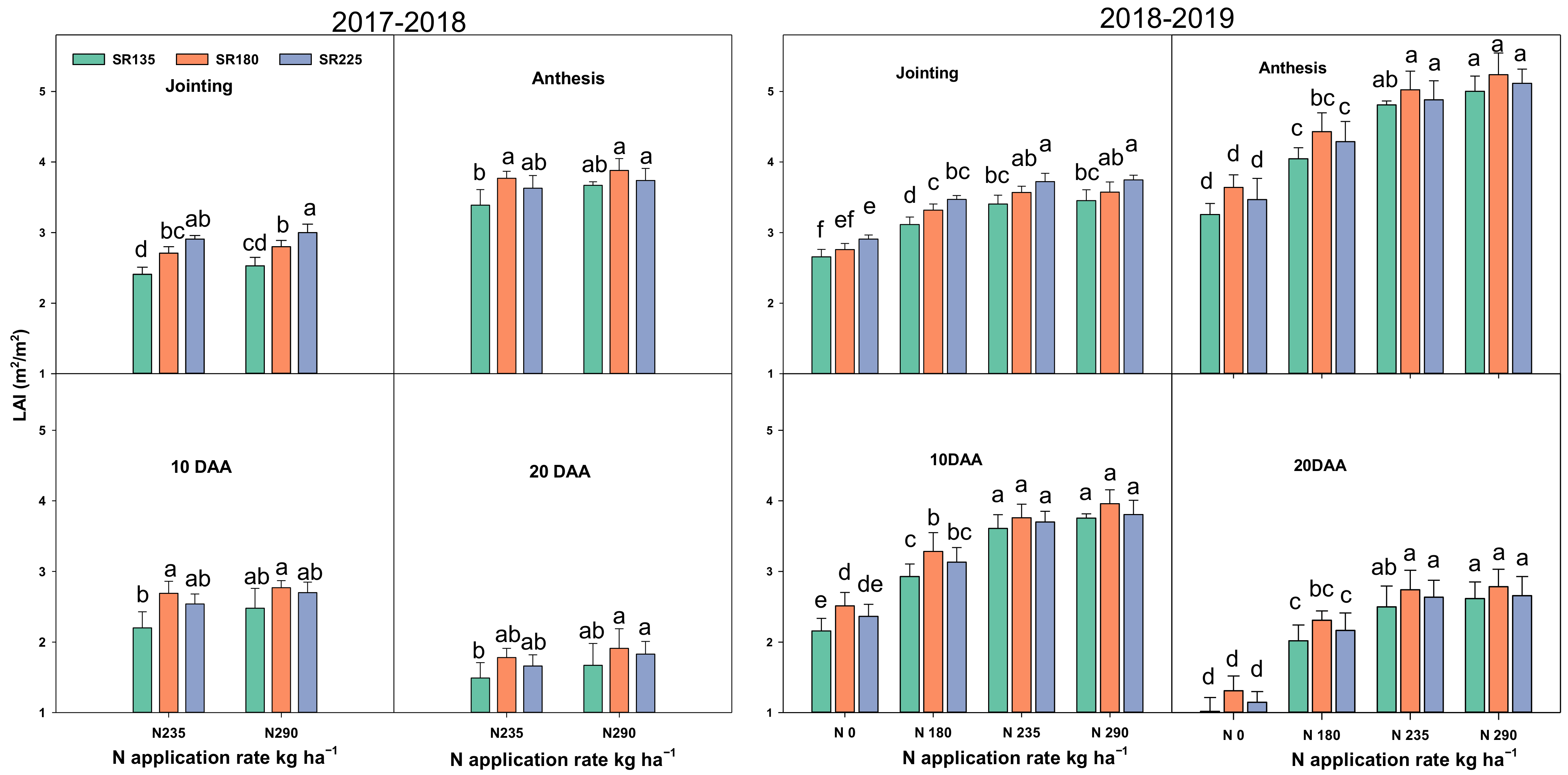
2.1.1. Photosynthetic Capacity, Chlorophyll Content, and Leaf Area Index
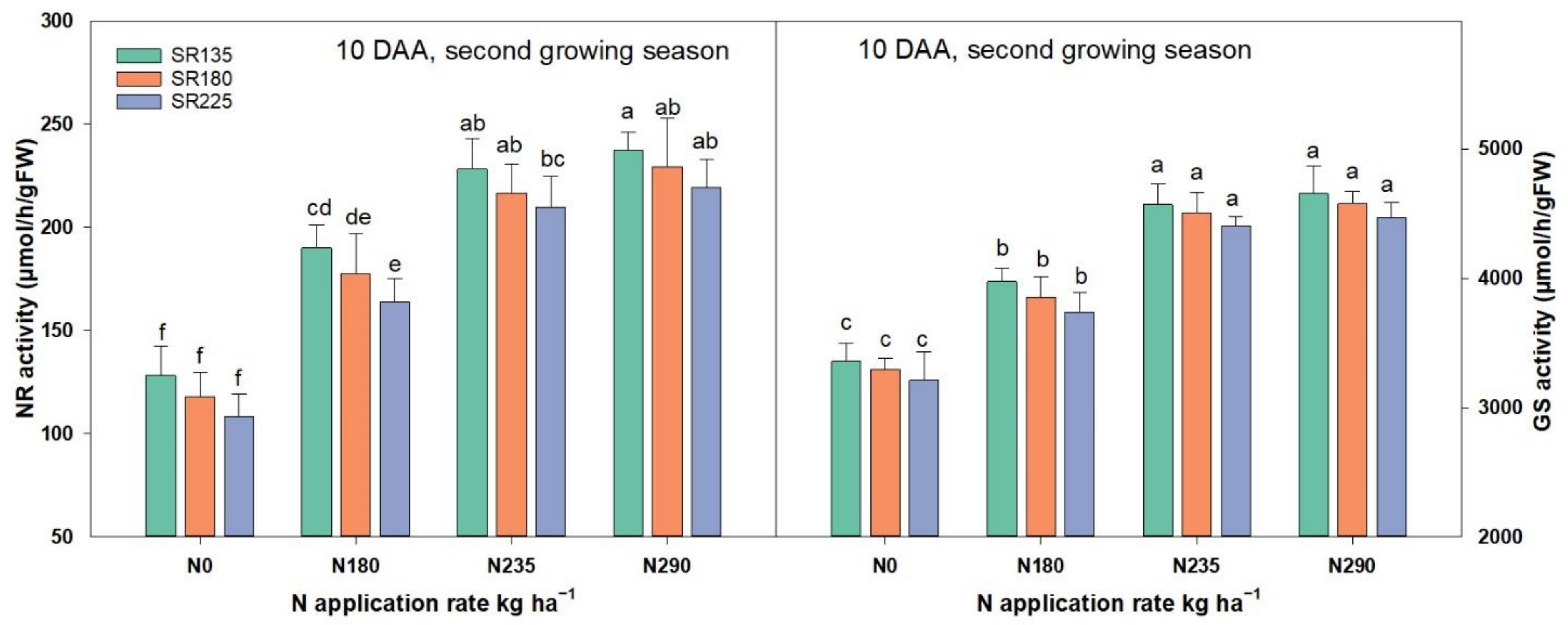
2.1.2. Enzymatic Activities of NR and GS
2.2. Grain Yield and Related Agronomic Characteristics
2.3. Accumulation, Translation, and Partitioning of DM
2.4. Accumulation, Translocation, and Partitioning of N
2.5. N Use Efficiency (NUE) Parameters
2.6. Correlation Analysis
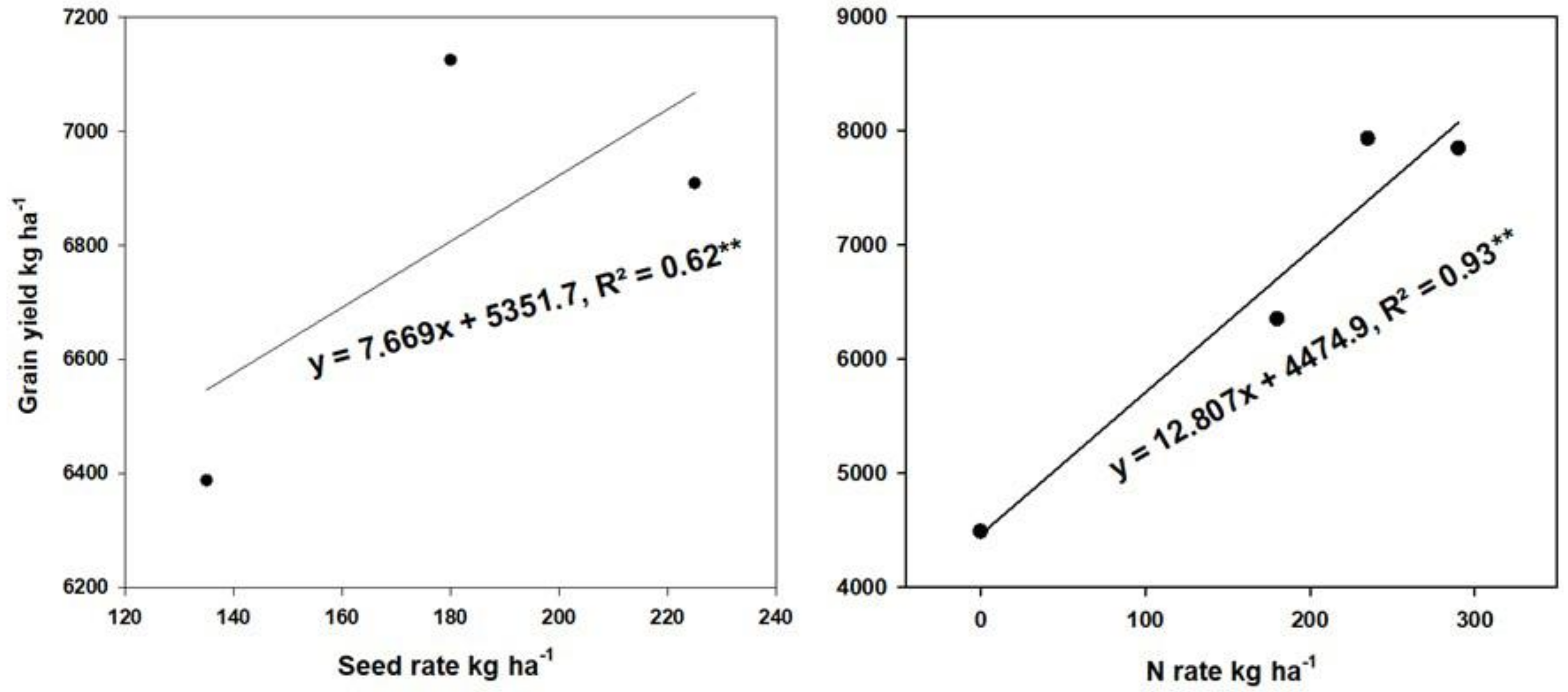
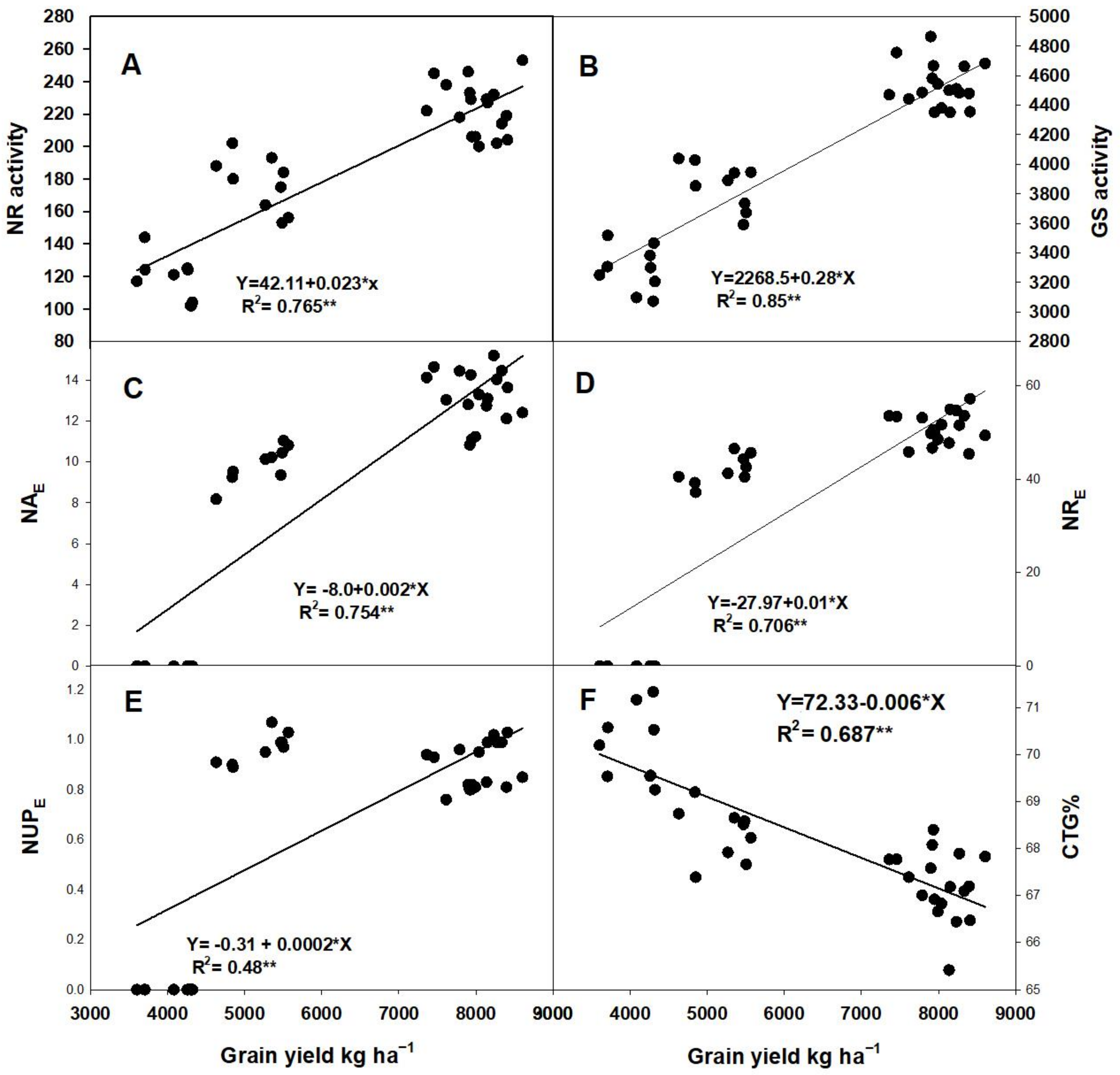
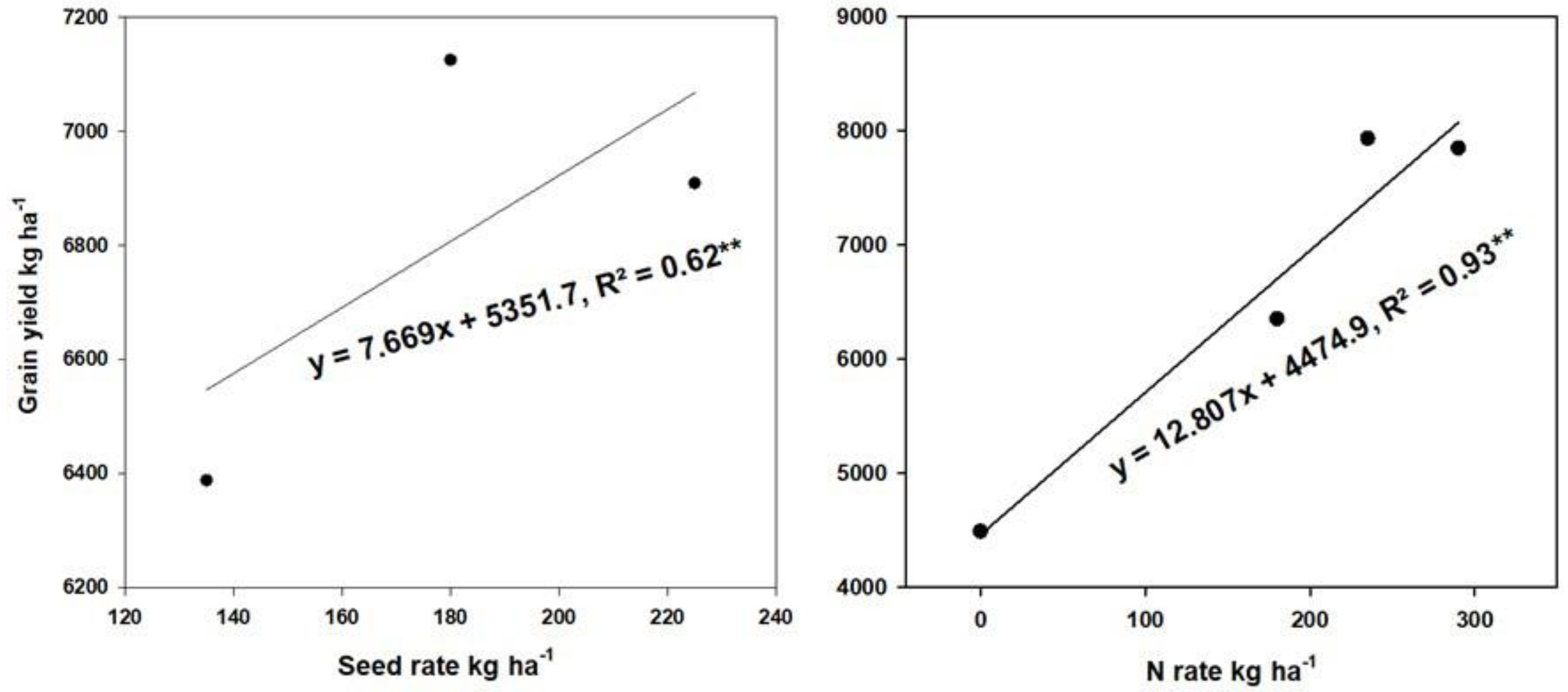
2.6.1. Correlation of GY with Agronomic and Photosynthesis Traits
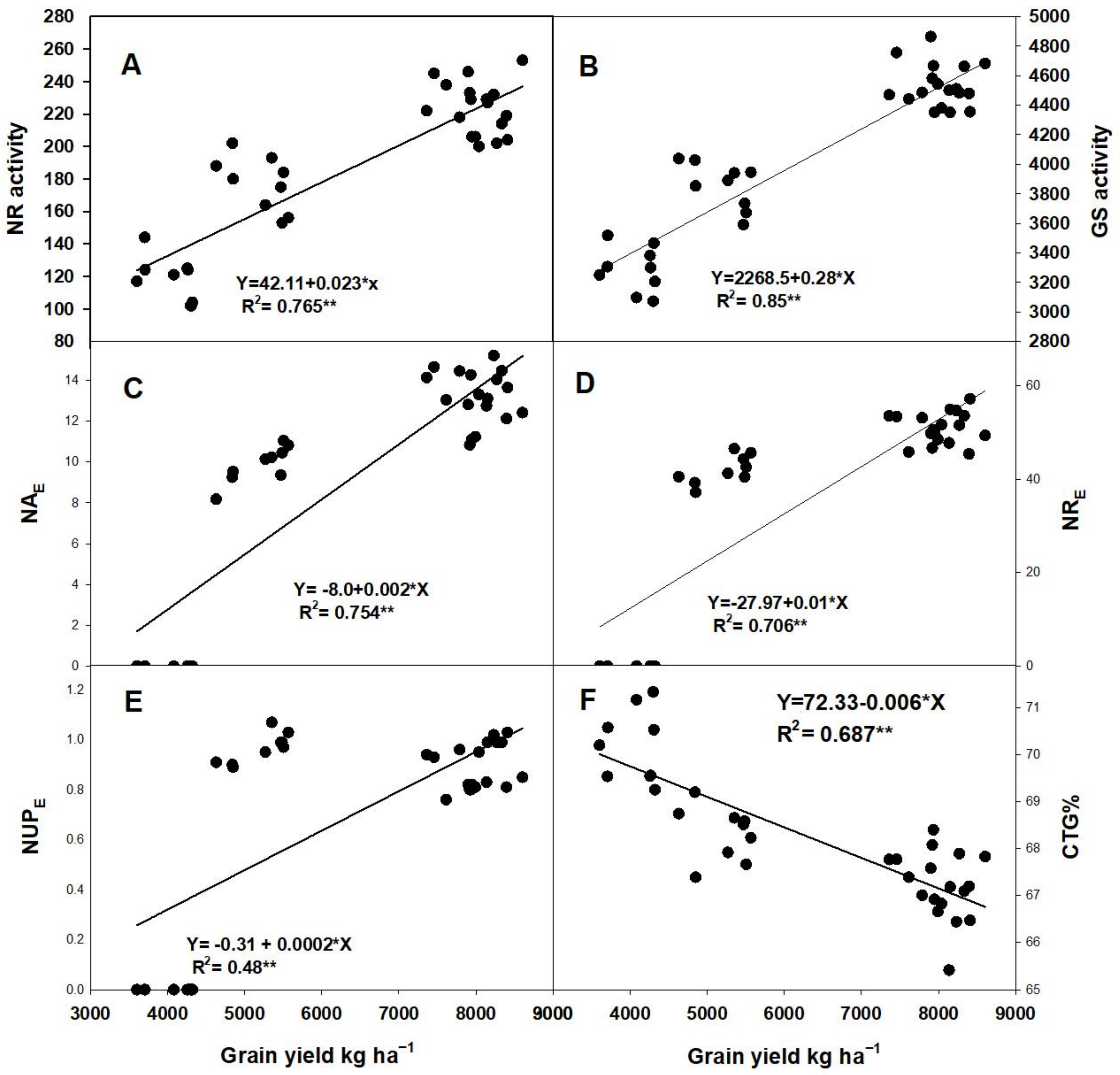
2.6.2. Relationship between GY and Enzyme Activities and NUE Parameters
3. Discussion
3.1. Physiological Characteristics
3.2. Grain Yield (GY) and N Use Efficiency (NUE)
3.3. Accumulation and Translocation of DM and N
3.4. Relationship of GY with Related Parameters
4. Materials and Methods
4.1. Plant Material and Experimental Site
4.2. Experiment Design
4.3. Grain Yield and Yield Components
4.4. Photosynthesis, SPAD, LAI, N, and Enzyme Activities in Leaves
4.5. Dry Matter (DM) and N Translocation
4.6. Use Efficiency (NUE) Parameters
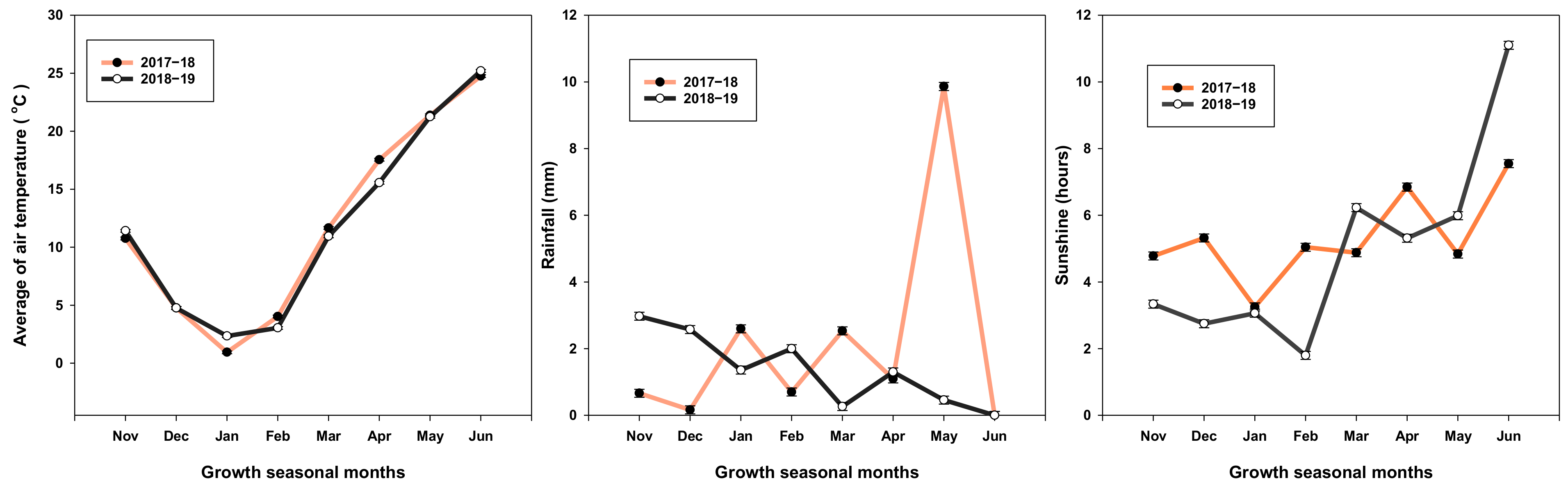
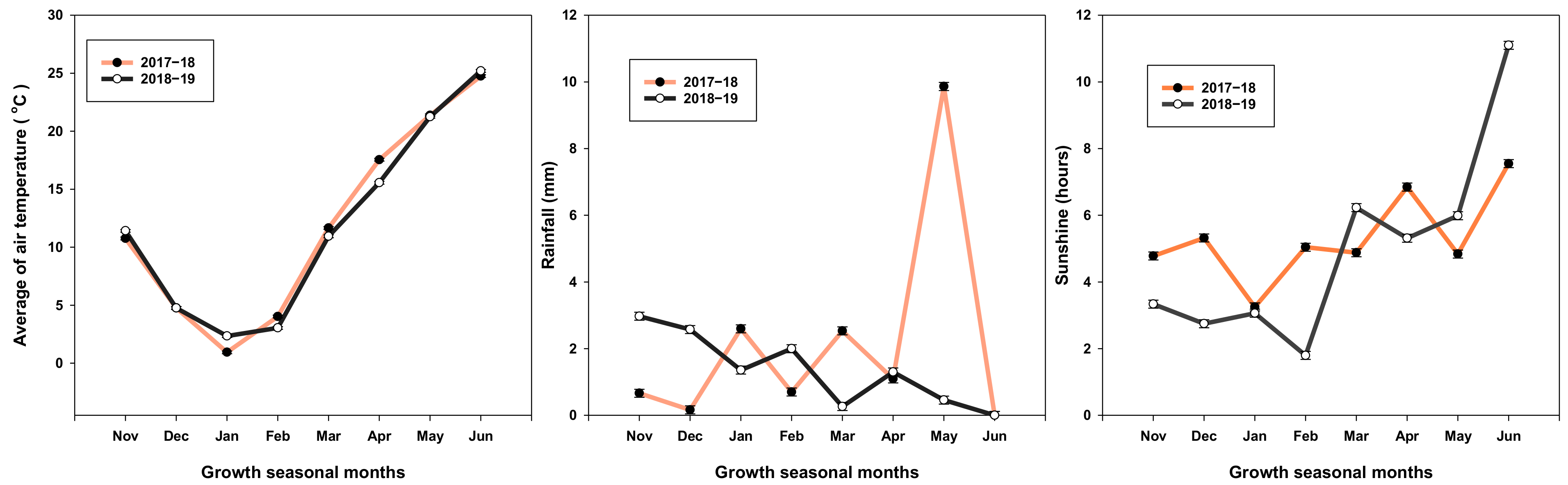
4.7. Weather Condition
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Slafer, G.A.; Savin, R.; Sadras, V.O. Coarse and fine regulation of wheat yield components in response to genotype and environment. Field Crops Res. 2014, 157, 71–83. [Google Scholar] [CrossRef]
- Lu, D.; Lu, F.; Yan, P.; Cui, Z.; Chen, X. Elucidating population establishment associated with N management and cultivars for wheat production in China. Field Crops Res. 2014, 163, 81–89. [Google Scholar] [CrossRef]
- Efretuei, A.; Gooding, M.; White, E.; Spink, J.; Hackett, R. Effect of nitrogen fertilizer application timing on nitrogen use efficiency and grain yield of winter wheat in Ireland. Ir. J. Agric. Food Res. 2016, 55, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Bly, A.G.; Woodard, H.J. Foliar nitrogen application timing influence on grain yield and protein concentration of hard red winter and spring wheat. Agron. J. 2003, 95, 335–338. [Google Scholar] [CrossRef]
- Fageria, N.; Baligar, V. Enhancing nitrogen use efficiency in crop plants. Adv. Agron. 2005, 88, 97–185. [Google Scholar]
- Lawlor, D.W.; Lemaire, G.; Gastal, F. Nitrogen, plant growth and crop yield. In Plant Nitrogen; Springer: Cham, Switzerland, 2001; pp. 343–367. [Google Scholar]
- Wang, Y.; Ren, T.; Lu, J.; Ming, R.; Li, P.; Hussain, S.; Cong, R.; Li, X. Heterogeneity in rice tillers yield associated with tillers formation and nitrogen fertilizer. Agron. J. 2016, 108, 1717–1725. [Google Scholar] [CrossRef]
- Huang, M.; Yang, C.; Ji, Q.; Jiang, L.; Tan, J.; Li, Y. Tillering responses of rice to plant density and nitrogen rate in a subtropical environment of southern China. Field Crops Res. 2013, 149, 187–192. [Google Scholar] [CrossRef]
- Balkcom, K.; Burmester, C. Nitrogen applications for wheat production across tillage systems in Alabama. Agron. J. 2015, 107, 425–434. [Google Scholar] [CrossRef]
- Terrile, I.I.; Miralles, D.J.; González, F.G. Fruiting efficiency in wheat (Triticum aestivum L): Trait response to different growing conditions and its relation to spike dry weight at anthesis and grain weight at harvest. Field Crops Res. 2017, 201, 86–96. [Google Scholar] [CrossRef]
- Yang, D.; Cai, T.; Luo, Y.; Wang, Z. Optimizing plant density and nitrogen application to manipulate tiller growth and increase grain yield and nitrogen-use efficiency in winter wheat. PeerJ 2019, 7, e6484. [Google Scholar] [CrossRef]
- Ju, X.-T.; Xing, G.-X.; Chen, X.-P.; Zhang, S.-L.; Zhang, L.-J.; Liu, X.-J.; Cui, Z.-L.; Yin, B.; Christie, P.; Zhu, Z.-L. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef] [Green Version]
- Mandic, V.; Krnjaja, V.; Tomic, Z.; Bijelic, Z.; Simic, A.; Ruzic Muslic, D.; Gogic, M. Nitrogen fertilizer influence on wheat yield and use efficiency under different environmental conditions. Chil. J. Agric. Res. 2015, 75, 92–97. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Christie, P.; Zhang, W.; Zhang, F. Crop Productivity, Fertilizer Use, and Soil Quality in China; CRC Press: Boca Raton, FL, USA, 2010; pp. 87–107. [Google Scholar]
- Ma, G.; Liu, W.; Li, S.; Zhang, P.; Wang, C.; Lu, H.; Xie, Y.; Ma, D.; Kang, G. Determining the optimal N input to improve grain yield and quality in winter wheat with reduced apparent N loss in the North China Plain. Front. Plant Sci. 2019, 10, 181. [Google Scholar] [CrossRef] [Green Version]
- Qingfeng, M.; Shanchao, Y.; Peng, H.; Zhenling, C.; Xinping, C. Improving yield and nitrogen use efficiency simultaneously for maize and wheat in China: A review. Pedosphere 2016, 26, 137–147. [Google Scholar]
- Wu, Y.; Xi, X.; Tang, X.; Luo, D.; Gu, B.; Lam, S.K.; Vitousek, P.M.; Chen, D. Policy distortions, farm size, and the overuse of agricultural chemicals in China. Proc. Natl. Acad. Sci. USA 2018, 115, 7010–7015. [Google Scholar] [CrossRef] [Green Version]
- Hirel, B.; Le Gouis, J.; Ney, B.; Gallais, A. The challenge of improving nitrogen use efficiency in crop plants: Towards a more central role for genetic variability and quantitative genetics within integrated approaches. J. Exp. Bot. 2007, 58, 2369–2387. [Google Scholar] [CrossRef]
- López-Bellido, L.; López-Bellido, R.J.; Redondo, R. Nitrogen efficiency in wheat under rainfed Mediterranean conditions as affected by split nitrogen application. Field Crops Res. 2005, 94, 86–97. [Google Scholar] [CrossRef]
- Zhai, B.; Li, S. Response to nitrogen deficiency and compensation on growth and yield of winter wheat. Plant Nutr. Fertitizer Sci. 2005, 11, 308–313. [Google Scholar]
- Tian, G.; Gao, L.; Kong, Y.; Hu, X.; Xie, K.; Zhang, R.; Ling, N.; Shen, Q.; Guo, S. Improving rice population productivity by reducing nitrogen rate and increasing plant density. PLoS ONE 2017, 12, e0182310. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Xiao, L.; Jia, D.; Kong, H.; Wang, Y.; Li, C.; Zhang, Y.; He, M. Increased plant density of winter wheat can enhance nitrogen–uptake from deep soil. Plant Soil 2014, 384, 141–152. [Google Scholar] [CrossRef]
- Meng, Q.; Yue, S.; Chen, X.; Cui, Z.; Ye, Y.; Ma, W.; Tong, Y.; Zhang, F. Understanding dry matter and nitrogen accumulation with time-course for high-yielding wheat production in China. PLoS ONE 2013, 8, e68783. [Google Scholar] [CrossRef] [Green Version]
- Dordas, C. Dry matter, nitrogen and phosphorus accumulation, partitioning and remobilization as affected by N and P fertilization and source–sink relations. Eur. J. Agron. 2009, 30, 129–139. [Google Scholar] [CrossRef]
- Jiang, D.; Dai, T.; Jing, Q.; Cao, W.; Zhou, Q.; Zhao, H.; Fan, X. Effects of long-term fertilization on leaf photosynthetic characteristics and grain yield in winter wheat. Photosynthetica 2004, 42, 439–446. [Google Scholar] [CrossRef]
- Bhatta, M.; Eskridge, K.M.; Rose, D.J.; Santra, D.K.; Baenziger, P.S.; Regassa, T. Seeding rate, genotype, and topdressed nitrogen effects on yield and agronomic characteristics of winter wheat. Crop Sci. 2017, 57, 951–963. [Google Scholar] [CrossRef] [Green Version]
- Geleta, B.; Atak, M.; Baenziger, P.; Nelson, L.; Baltenesperger, D.; Eskridge, K.M.; Shipman, M.; Shelton, D. Seeding rate and genotype effect on agronomic performance and end-use quality of winter wheat. Crop Sci. 2002, 42, 827–832. [Google Scholar]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [Green Version]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Dong, S.; Dai, X.; Wu, T.; Wang, X.; Bai, H.; Wang, L.; He, M. Combined effect of plant density and nitrogen input on grain yield, nitrogen uptake and utilization of winter wheat. Vegetos-Int. J. Plant Res. 2016, 29, 63–73. [Google Scholar] [CrossRef]
- Zhang, J.H.; Jian-Li, L.; Zhang, J.-B.; Fu-Tao, Z.; Cheng, Y.-N.; Wei-Peng, W. Effects of Nitrogen Application rates on translocation of dry matter and Nitrogen utilization in Rice and Wheat. Acta Agron. Sin. 2010, 36, 1736–1742. [Google Scholar] [CrossRef]
- Ma, D.-H.; Wang, Y.F.; Zhou, H.; Sun, H.J. Effect of Postanthesis Soil Water Status and Nitrogen on Grain Yield and Canopy Biomass Accumulation and Transportation of Winter Wheat. J. Triticeae Crops 2007, 5, 847–851. [Google Scholar]
- Arduini, I.; Masoni, A.; Ercoli, L.; Mariotti, M. Grain yield, and dry matter and nitrogen accumulation and remobilization in durum wheat as affected by variety and seeding rate. Eur. J. Agron. 2006, 25, 309–318. [Google Scholar] [CrossRef]
- Liu, W.; Wang, J.; Wang, C.; Ma, G.; Wei, Q.; Lu, H.; Xie, Y.; Ma, D.; Kang, G. Root growth, water and nitrogen use efficiencies in winter wheat under different irrigation and nitrogen regimes in North China Plain. Front. Plant Sci. 2018, 9, 1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.Z.; Zhou, G.S. Combined effects of water stress and high temperature on photosynthesis, nitrogen metabolism and lipid peroxidation of a perennial grass Leymus chinensis. Planta 2006, 224, 1080–1090. [Google Scholar] [CrossRef] [PubMed]
- Ferrario-Méry, S.; Valadier, M.-H.; Foyer, C.H. Overexpression of nitrate reductase in tobacco delays drought-induced decreases in nitrate reductase activity and mRNA. Plant Physiol. 1998, 117, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.-L.; Biswas, D.K. Precision nitrogen management for sustainable corn production. In Sustainable Agriculture Reviews; Springer: Cham, Switzerland, 2015; pp. 33–62. [Google Scholar]
- Biswas, D.K.; Ma, B.-L. Effect of nitrogen rate and fertilizer nitrogen source on physiology, yield, grain quality, and nitrogen use efficiency in corn. Can. J. Plant Sci. 2016, 96, 392–403. [Google Scholar] [CrossRef] [Green Version]
- Bremner, J.M.; Mulvaney, C. Nitrogen—Total. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1982; pp. 595–624. [Google Scholar]
- Zhang, C.; Peng, S.; Bennett, J. Glutamine synthetase and its isoforms in rice spikelets and rachis during grain development. J. Plant Physiol. 2000, 156, 230–233. [Google Scholar] [CrossRef]
- Wu, H.; Xiang, J.; Zhang, Y.; Zhang, Y.; Peng, S.; Chen, H.; Zhu, D. Effects of post-anthesis nitrogen uptake and translocation on photosynthetic production and rice yield. Sci. Rep. 2018, 8, 12891. [Google Scholar] [CrossRef]
- Pask, A.J.D.; Sylvester-Bradley, R.; Jamieson, P.D.; Foulkes, M.J. Quantifying how winter wheat crops accumulate and use nitrogen reserves during growth. Field Crops Res. 2012, 126, 104–118. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N kg ha−1 | SR kg ha−1 | TY t ha−1 | NS × 104 ha−1 | TGW(g) | NGS | HI | PH (cm) | |
|---|---|---|---|---|---|---|---|---|
| 2017–2018 | N235 | SR135 | 5.05 b | 357 c | 39.7 a | 35.63 a | 0.41 c | 73.4 c |
| SR180 | 5.78 a | 414 b | 39.5 ab | 35.38 a | 0.44 ab | 74.33 bc | ||
| SR225 | 5.7 a | 425 ab | 39.2 bc | 34.27 b | 0.45 a | 75.74 ab | ||
| N290 | SR135 | 5.03 b | 361 c | 39.1 bc | 35.63 a | 0.41 c | 76.43 ab | |
| SR180 | 5.77 a | 420 ab | 39 bc | 35.3 a | 0.43 bc | 77.23 a | ||
| SR225 | 5.77 a | 434 a | 38.9 c | 34.18 b | 0.42 bc | 77.57 a | ||
| F-Value | N | 0.113 | 2.475 | 13.65 ** | 0.098 | 7.478 * | 19.88 ** | |
| SR | 107.1 ** | 105.9 ** | 3.38 * | 21.65 ** | 11.46 ** | 2.999 | ||
| N*S | 0.331 | 0.123 | 0.63 | 0.025 | 1.637 | 0.426 | ||
| N0 | SR135 | 4 g | 391 g | 35.7 bc | 28.8 d | 0.35 e | 62.93 d | |
| SR180 | 4.8 f | 479 ef | 35.6 bc | 28.1 d | 0.4 cde | 63.62 d | ||
| SR225 | 4.8 f | 503 de | 35.5 c | 27.2 d | 0.4 cd | 64.07 d | ||
| 2018–2019 | N180 | SR135 | 5.6 e | 448 f | 36.4 ab | 34.5 c | 0.38 de | 72.87 c |
| SR180 | 6.7 d | 543 bc | 36.3 abc | 34.1 c | 0.41 cd | 74.02 bc | ||
| SR225 | 6.6 d | 555 b | 36.3 abc | 33.1 c | 0.42 bc | 74.47 b | ||
| N235 | SR135 | 7.5 c | 518 cd | 36.7 a | 39.6 ab | 0.45 ab | 73.97 bc | |
| SR180 | 8.3 a | 593 a | 36.7 a | 38.3 ab | 0.46 a | 75.15 ab | ||
| SR225 | 8.2 a | 597 a | 36.5 ab | 37.5 b | 0.46 a | 75.75 a | ||
| N290 | SR135 | 7.8 bc | 529 bc | 36.6 a | 40.4 a | 0.45 ab | 73.95 bc | |
| SR180 | 8.2 a | 587 a | 36.4 ab | 38.6 ab | 0.44 ab | 75.22 ab | ||
| SR225 | 8.1 ab | 591 a | 36.4 ab | 37.7 ab | 0.45 ab | 75.97 a | ||
| F-Value | N | 690 ** | 74.4 ** | 9.66 ** | 242.5 ** | 43.55 ** | 598.3 ** | |
| SR | 60.1 ** | 83.94 ** | 0.66 | 12.19 ** | 4.48 * | 17.8 ** | ||
| N*SR | 2.64 * | 1.36 | 0.027 | 0.394 | 1.77 | 0.25 |
| Total DM Accumulation kg ha−1 | DM Translocation | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| N kg ha−1 | SR kg ha−1 | So-JT | JT-An | An-M | So-M | PTA kg ha−1 | CPT% | PAA kg ha−1 | CPA% |
| N0 | SR135 | 1959 h | 6139 b | 2680 c | 10778 g | 1214.1 f | 30.3 a | 4256 d | 69.7 e |
| SR180 | 2314 g | 6514 b | 3250 bc | 12079 f | 1322.8 f | 27.6 b | 5072 d | 72.4 d | |
| SR225 | 2401 f | 6682 b | 3025 bc | 12108 f | 1439 e | 29.7 a | 5018 d | 70.3 e | |
| N180 | SR135 | 2395 f | 8585 a | 3708 b | 14689 e | 1529.4 de | 27.2 bc | 6053 c | 72.8 cd |
| SR180 | 2856 b | 8904 a | 4780 a | 16539 cd | 1633.8 cd | 24.3 d | 7276 b | 75.7 b | |
| SR225 | 2488 e | 8587 a | 4646 a | 15721 d | 1701.4 c | 25.6 cd | 7164 b | 74.4 bc | |
| N235 | SR135 | 2668 d | 9335 a | 4731 a | 16735 c | 1918.7 ab | 25.5 cd | 7804 ab | 74.5 bc |
| SR180 | 3019 a | 10027 a | 5130 a | 18177 ab | 1963.7 a | 23.6 de | 8727 a | 76.4 ab | |
| SR225 | 3033 a | 9804 a | 5064 a | 17902 ab | 2028.5 a | 24.9 d | 8490 a | 75.1 b | |
| N290 | SR135 | 2719 c | 9477 a | 5048 a | 17245 bc | 1905.8 ab | 24.4 d | 8141 ab | 75.6 b |
| SR180 | 2998 a | 10066 a | 5418 a | 18483 a | 1823.2 b | 22.1 e | 8776 a | 77.9 a | |
| SR225 | 3010 a | 9649 a | 5459 a | 18119 ab | 1906.7 ab | 24 de | 8518 a | 76.4 ab | |
| F-Value | N | 1817 ** | 28.1 ** | 32.1 ** | 258 ** | 167.9 ** | 50.4 ** | 92.454 ** | 50.4 ** |
| SR | 84 ** | 0.9 | 4.3 * | 23.8 ** | 11.1 ** | 16 ** | 9.546 ** | 16 ** | |
| N*SR | 67.1 ** | 0.1 | 0.3 | 0.4 | 1.9 | 0.2 | 0.28 | 0.2 | |
| N kg ha−1 | SR kg ha−1 | Grain | Rachis + Glumes | Culms + Sheaths | Leaves | ||||
|---|---|---|---|---|---|---|---|---|---|
| Anthesis | Maturity | Anthesis | Maturity | Anthesis | Maturity | Anthesis | Maturity | ||
| N0 | SR135 | 4005 g | 1576 c | 1465 f | 5442 g | 4462 c | 1080 f | 846 f | |
| SR180 | 4788 f | 1821 c | 1606 e | 5631 f | 4741 bc | 1376 e | 944 e | ||
| SR225 | 4851 f | 1994 c | 1606 e | 5701 f | 4682 bc | 1389 e | 968 e | ||
| N180 | SR135 | 5623 e | 2344 bc | 1959 d | 7217 d | 5932 a | 1420 e | 1175 d | |
| SR180 | 6712 d | 2496 bc | 2198 bc | 7426 c | 6299 a | 1837 c | 1330 c | ||
| SR225 | 6647 d | 2517 bc | 2218 bc | 6716 e | 5525 ab | 1841 c | 1330 c | ||
| N235 | SR135 | 7535 c | 3072 ab | 2187 c | 7156 d | 5642 a | 1775 d | 1370 b | |
| SR180 | 8324 a | 3597 a | 2366 a | 7375 c | 6017 a | 2075 a | 1468 a | ||
| SR225 | 8152 ab | 3426 ab | 2366 a | 7341 c | 5919 a | 2070 a | 1464 a | ||
| N290 | SR135 | 7814 bc | 3092 ab | 2232 b | 7199 d | 5807 a | 1906 b | 1391 b | |
| SR180 | 8244 a | 3358 ab | 2355 a | 7641 a | 6425 a | 2066 a | 1458 a | ||
| SR225 | 8087 ab | 3058 ab | 2337 a | 7545 b | 6241 a | 2057 a | 1453 a | ||
| F-Value | N | 690 ** | 14.8 ** | 2002 ** | 2858.3 ** | 18.4 ** | 955.9 ** | 1001 ** | |
| SR | 60.1 ** | 0.92 | 207 ** | 96.3 ** | 2.3 | 324.9 ** | 85.5 ** | ||
| N*SR | 2.64 * | 0.17 | 6.29 ** | 61.7 ** | 0.58 | 11.1 ** | 2.92 * | ||
| Total N Accumulation kg ha−1 | N Translocation | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| N kg ha−1 | SR kg ha−1 | So-JT | JT-An | An -M | So-M | NTA kg ha−1 | CTG% | NAA kg ha−1 | CAG% |
| N0 | SR135 | 25.6 h | 53.4 i | 12.4 g | 91.4 k | 48.3 j | 70.1 ab | 27.4 j | 29.9 de |
| SR180 | 28.5 g | 60.4 h | 14.5 f | 103.4 i | 55.5 i | 69.4 bc | 31.5 h | 30.6 cd | |
| SR225 | 28.8 g | 60.4 h | 11 g | 100.2 j | 55.3 i | 71 a | 29.2 i | 29 e | |
| N180 | SR135 | 41.3 f | 97.2 g | 23.2 e | 161.6 h | 84.3 h | 68.4 cd | 48.9 g | 31.6 bc |
| SR180 | 45.6 e | 108.5 e | 30.2 cd | 184.3 f | 97.8 f | 68.2 d | 56.5 e | 31.8 b | |
| SR225 | 46.1 e | 100.6 f | 29.1 d | 175.8 g | 94.7 g | 68.3 cd | 54.2 f | 31.7 bc | |
| N235 | SR135 | 57.4 d | 135.1 d | 29.7 cd | 222.2 e | 118.1 e | 67.5 de | 70 d | 32.5 ab |
| SR180 | 61.7 ab | 145.3 a | 31.5 c | 238.6 ab | 125.9 ab | 66.7 e | 76.9 a | 33.3 a | |
| SR225 | 61.6 ab | 136.3 d | 31.5 c | 229.4 d | 122.1 d | 67.3 de | 72.7 bc | 32.7 ab | |
| N290 | SR135 | 59 c | 140.1 b | 31.3 c | 230.4 d | 123.2 cd | 67.8 de | 72.1 c | 32.2 ab |
| SR180 | 61.9 a | 145.8 a | 34.2 b | 241.8 a | 126.9 a | 66.6 e | 77.7 a | 33.4 a | |
| SR225 | 61 b | 138 c | 36.1 a | 235 c | 124.6 bc | 67.4 de | 74.2 b | 32.6 ab | |
| F-Value | N | 1159 ** | 2229 ** | 628.1 ** | 6418 ** | 9584 ** | 44.2 ** | 436.6 ** | 44.2 ** |
| SR | 270.4 ** | 377.9 ** | 31.8 ** | 1283 ** | 195.7 ** | 5.7 * | 115.6 ** | 5.7 * | |
| N*SR | 7.1 ** | 31.7 ** | 8.72 ** | 44.3 ** | 15.81 ** | 1 | 3.03 * | 1 | |
| N kg ha−1 | SR kg ha−1 | N at Grain | N at Rachis + Glumes | NA at Culms + Sheaths | N at Leaves | ||||
|---|---|---|---|---|---|---|---|---|---|
| Anthesis | Maturity | Anthesis | Maturity | Anthesis | Maturity | Anthesis | Maturity | ||
| N0 | SR135 | 68.9 f | 15 g | 6.8 c | 34.9 g | 10.9 e | 29.2 i | 4.9 d | |
| SR180 | 80 e | 17.1 f | 7.1 c | 35.1 g | 11.2 e | 36.8 g | 5.2 d | ||
| SR225 | 77.8 ef | 18.2 f | 6.6 c | 34.9 g | 10.6 e | 36.1 h | 5.1 d | ||
| N180 | SR135 | 123.2 d | 25.7 e | 10 b | 65.2 e | 18.1 cd | 47.6 f | 10.3 c | |
| SR180 | 143.4 c | 26.2 e | 10.8 b | 67.1 d | 18.8 bc | 60.8 e | 11.3 b | ||
| SR225 | 138.5 c | 25.1 e | 10.3 b | 60.6 f | 15.9 d | 60.9 e | 11 bc | ||
| N235 | SR135 | 174.9 b | 40.3 c | 13.2 a | 87.7 b | 20.1 abc | 64.5 d | 14 a | |
| SR180 | 188.9 a | 45.4 a | 13.9 a | 88.4 b | 21 abc | 73.3 a | 14.7 a | ||
| SR225 | 181.4 ab | 41.2 c | 13.4 a | 85.7 c | 20.2 abc | 70.9 b | 14.4 a | ||
| N290 | SR135 | 181.7 ab | 40.8 c | 13.5 a | 89.1 b | 20.7 abc | 69.2 c | 14.4 a | |
| SR180 | 190.4 a | 43.5 b | 14.1 a | 91.2 a | 22.5 a | 73 a | 14.8 a | ||
| SR225 | 184.9 a | 38.1 d | 13.9 a | 89.5 ab | 21.7 ab | 71.4 b | 14.5 a | ||
| F-Value | N | 845.1 ** | 2010.4 ** | 322.7 ** | 5132.3 ** | 83.3 ** | 24379 ** | 595 ** | |
| SR | 19.80 ** | 36.8 ** | 4.35 * | 19.9 ** | 2.02 | 2256 ** | 3.63 * | ||
| N*SR | 0.98 | 13.9 ** | 0.2 | 5.6 * | 0.66 | 172.9 ** | 0.3 | ||
| N Rate kg ha−1 | SR kg ha−1 | NAE | NRE | NUPE | NPFP | NHI |
|---|---|---|---|---|---|---|
| kg kg−1 | % | kg kg−1 | kg kg−1 | % | ||
| N0 | SR135 | 0.75 f | ||||
| SR180 | 0.77 de | |||||
| SR225 | 0.77 de | |||||
| N180 | SR135 | 9.0 f | 39 e | 0.9 c | 31.2 c | 0.76 ef |
| SR180 | 10.7 de | 44.9 cd | 1.02 a | 37.3 a | 0.78 bc | |
| SR225 | 10 e | 42 de | 0.98 a | 36.9 a | 0.79 ab | |
| N235 | SR135 | 14.4 a | 53.4 a | 0.95 b | 30.8 c | 0.79 ab |
| SR180 | 14.4 a | 55.2 a | 1.02 a | 34 b | 0.79 ab | |
| SR225 | 13.5 b | 52.7 a | 0.98 b | 33.3 b | 0.8 a | |
| N290 | SR135 | 13.4 b | 48.8 b | 0.8 d | 27.4 e | 0.79 ab |
| SR180 | 12.1 c | 48.6 b | 0.83 d | 28.9 d | 0.79 ab | |
| SR225 | 11.4 cd | 47.3 bc | 0.81 d | 28.4 de | 0.79 ab | |
| F-value | N | 156.4 ** | 82.6 ** | 3774.8 ** | 180.8 ** | 17.6 ** |
| SR | 6.4 ** | 4.4 * | 20.04 ** | 55.4 ** | 7.1 ** | |
| N*SR | 8.76 ** | 2.16 | 4.27 ** | 9.12 ** | 1.88 |
| GY | NS | NGS | TGW | PH | HI | Pn | Gs | SPAD | LAI | |
|---|---|---|---|---|---|---|---|---|---|---|
| GY | 1 | 0.854 ** | 0.893 ** | 0.626 ** | 0.116 | 0.879 ** | 0.917 ** | 0.788 ** | 0.889 ** | 0.961 ** |
| NS | 1 | 0.535 ** | 0.418 * | 0.346* | 0.733 ** | 0.641 ** | 0.492 ** | 0.638 ** | 0.777 ** | |
| NGS | 1 | 0.613 ** | −0.08 | 0.804 ** | 0.942 ** | 0.860 ** | 0.909 ** | 0.900 ** | ||
| TGW | 1 | 0.212 | 0.432 ** | 0.676 ** | 0.566 ** | 0.699 ** | 0.633 ** | |||
| PH | 1 | −0.102 | 0.031 | −0.138 | 0.22 | 0.117 | ||||
| HI | 1 | 0.747 ** | 0.660 ** | 0.673 ** | 0.821 ** | |||||
| Pn | 1 | 0.852 ** | 0.951 ** | 0.923 ** | ||||||
| Gs | 1 | 0.813 ** | 0.814 ** | |||||||
| SPAD | 1 | 0.894 ** | ||||||||
| LAI | 1 |
| Parameters | Equation | Unit | |
|---|---|---|---|
| Abbreviation | Denotation | ||
| PTA | Pre-anthesis DM translocation | DM of vegetative parts at anthesis—at maturity | kg ha−1 |
| CPT | Contribution of pre-anthesis DM translocation to grain | PTA ÷ GY at maturity × 100 | % |
| PAA | Post-anthesis DM accumulation | Biomass at maturity- biomass at anthesis | kg ha−1 |
| CPA | Contribution of post-anthesis DM accumulation to grain | PAA ÷ GY at maturity × 100 | % |
| NTA | Pre-anthesis N translocation | N of vegetative parts at anthesis—at maturity | kg ha−1 |
| CTG | Contribution of pre-anthesis N translocation to grain | NTA ÷ grain N×100 | % |
| NAA | Post-anthesis N accumulation | Plant N accumulation at maturity—N accumulation at anthesis | kg ha−1 |
| CAG | Contribution rate of post-anthesis N accumulation to grain | NAA ÷ grain N ×100 | % |
| Parameters | Equation | Unit | |
|---|---|---|---|
| Abbreviation | Denotation | ||
| NAE | N agronomy efficiency | (GY with N—GY without N) ÷ N application rate | kg kg−1 |
| NRE | N recovery efficiency | (total N uptake with N- total N uptake without N) ÷ N application rate | % |
| NUPE | N uptake efficiency | Above-ground N at harvesting ÷ N application rate | % |
| NPFP | N partial factor productivity | GY ÷ N application rate | kg kg−1 |
| NHI | N harvest index | Grain N accumulation at maturity/plant N accumulation at maturity | mg mg−1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmood, H.; Cai, J.; Zhou, Q.; Wang, X.; Samo, A.; Huang, M.; Dai, T.; Jahan, M.S.; Jiang, D. Optimizing Nitrogen and Seed Rate Combination for Improving Grain Yield and Nitrogen Uptake Efficiency in Winter Wheat. Plants 2022, 11, 1745. https://doi.org/10.3390/plants11131745
Mahmood H, Cai J, Zhou Q, Wang X, Samo A, Huang M, Dai T, Jahan MS, Jiang D. Optimizing Nitrogen and Seed Rate Combination for Improving Grain Yield and Nitrogen Uptake Efficiency in Winter Wheat. Plants. 2022; 11(13):1745. https://doi.org/10.3390/plants11131745
Chicago/Turabian StyleMahmood, Hemat, Jian Cai, Qin Zhou, Xiao Wang, Allan Samo, Mei Huang, Tingbo Dai, Mohammad Shah Jahan, and Dong Jiang. 2022. "Optimizing Nitrogen and Seed Rate Combination for Improving Grain Yield and Nitrogen Uptake Efficiency in Winter Wheat" Plants 11, no. 13: 1745. https://doi.org/10.3390/plants11131745
APA StyleMahmood, H., Cai, J., Zhou, Q., Wang, X., Samo, A., Huang, M., Dai, T., Jahan, M. S., & Jiang, D. (2022). Optimizing Nitrogen and Seed Rate Combination for Improving Grain Yield and Nitrogen Uptake Efficiency in Winter Wheat. Plants, 11(13), 1745. https://doi.org/10.3390/plants11131745