
Improving the Carotenoid Content in Maize by Using Isonuclear Lines

 ,
,  ,
,  , ,
, ,
Abstract
1. Introduction
2. Results and Discussion
2.1. Cytoplasm Influence on Isonuclear Lines
2.2. Cytoplasm × Nucleus Influence on Isonuclear Lines
2.3. Cytoplasm Influence on Hybrids
2.4. Cytoplasm × Nucleus Influence on Hybrids
2.5. Cytoplasm × Nucleus × Tester Influence on Hybrids
3. Materials and Methods
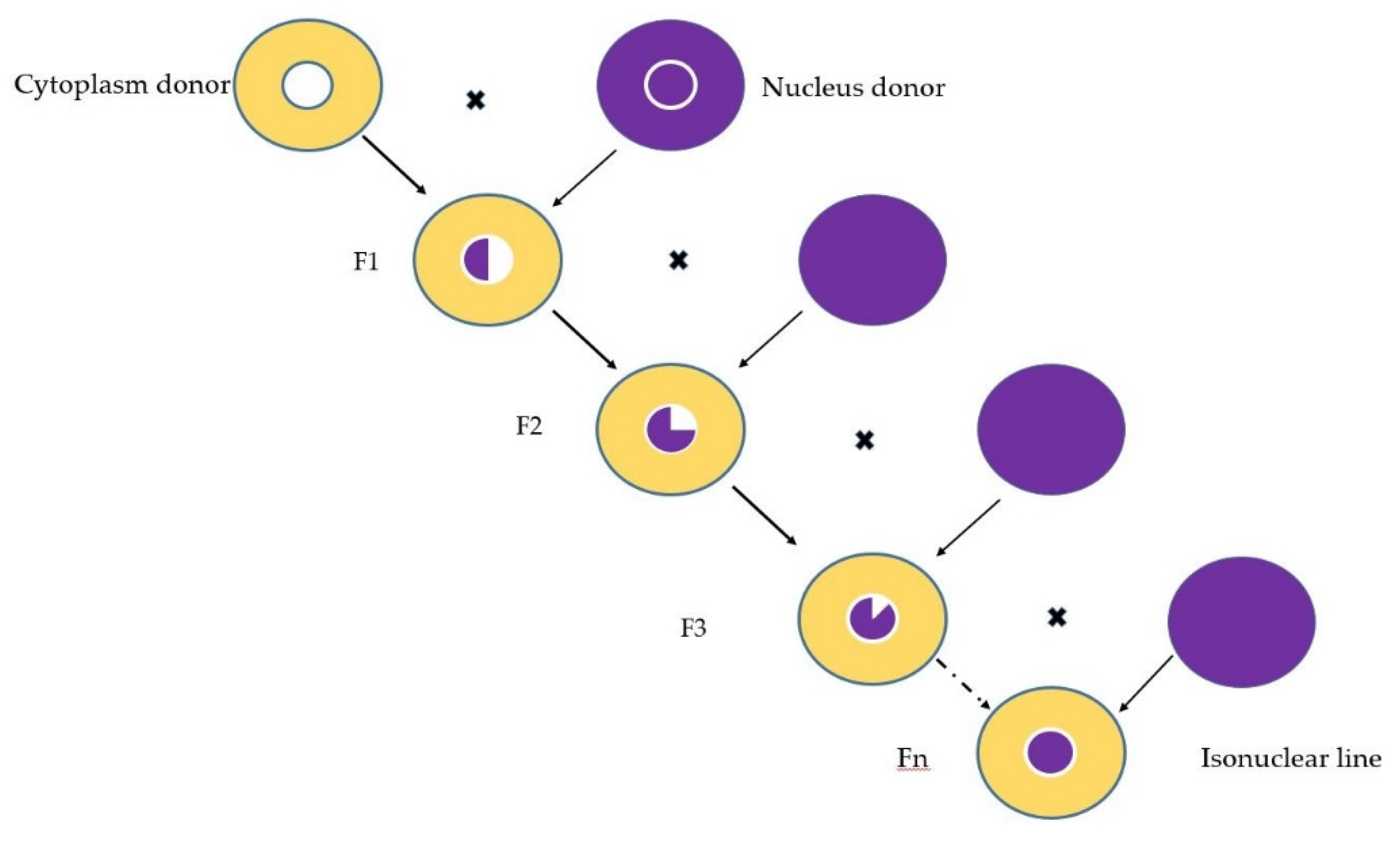
3.1. Biological Material and Experimental Design
3.2. Materials
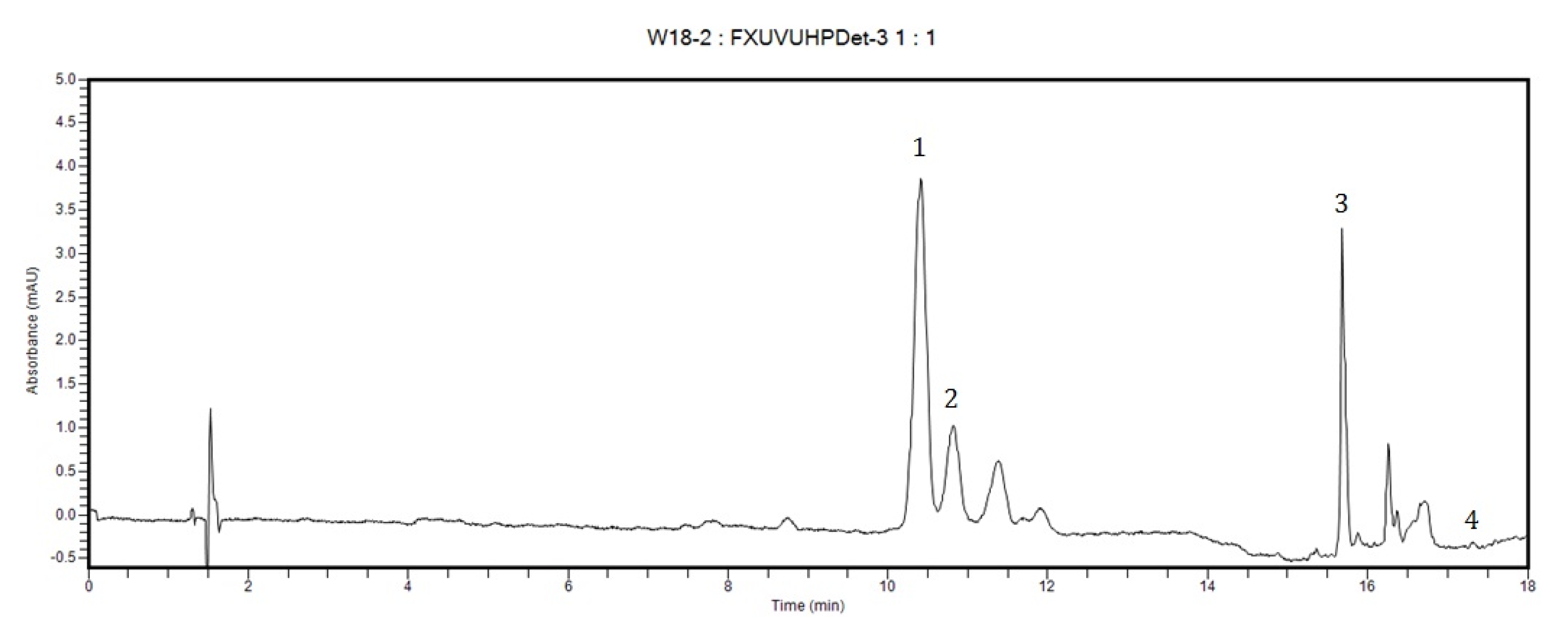
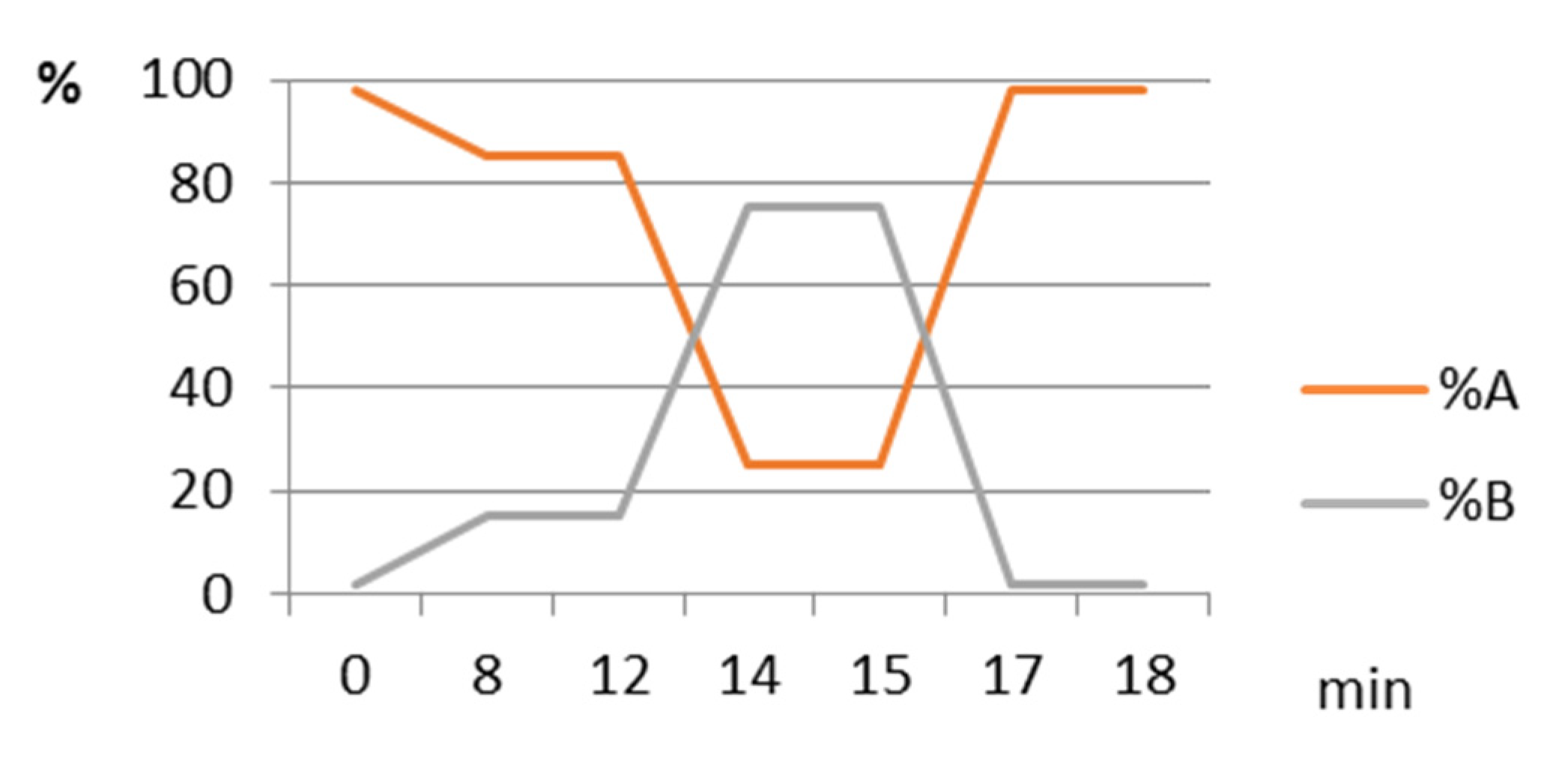
3.3. Quantification of Carotenoids
3.4. Data Processing
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data (accessed on 21 February 2022).
- Porumb-Ministerul Agriculturii Si Dezvoltarii Rurale. Available online: https://www.madr.ro/culturi-de-camp/cereale/porumb.html (accessed on 7 April 2022).
- Popescu, M.; Cristea, S.; Chiriac, A.-R.; Ţeican, D.-I. The Evolution of Maize Production in Romania. In 9th Edition of the International Symposium, Proceedings of the Agrarian Economy and Rural Development-Realities and Perspectives for Romania, November 2018, Bucharest, Romania; The Research Institute for Agricultural Economy and Rural Development (ICEADR): Bucharest, Romania, 2018; pp. 132–136. [Google Scholar]
- Ghețe, A.B.; Haș, V.; Vidican, R.; Copândean, A.; Ranta, O.; Moldovan, C.M.; Crișan, I.; Duda, M.M. Influence of Detasseling Methods on Seed Yield of Some Parent Inbred Lines of Turda Maize Hybrids. Agronomy 2020, 10, 729. [Google Scholar] [CrossRef]
- Zurak, D.; Grbeša, D.; Duvnjak, M.; Kiš, G.; Međimurec, T.; Kljak, K. Carotenoid Content and Bioaccessibility in Commercial Maize Hybrids. Agriculture 2021, 11, 586. [Google Scholar] [CrossRef]
- Burt, A.J.; Grainger, C.M.; Young, J.C.; Shelp, B.J.; Lee, E.A. Impact of Postharvest Handling on Carotenoid Concentration and Composition in High-Carotenoid Maize (Zea Mays L.) Kernels. J. Agric. Food Chem. 2010, 58, 8286–8292. [Google Scholar] [CrossRef]
- Asson-Batres, M.A.; Rochette-Egly, C. Biochemistry of Retinoid Signaling II; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Iseghohi, I.; Abe, A.; Meseka, S.; Mengesha, W.; Gedil, M.; Menkir, A. Assessing Effect of Marker-Based Improvement of Maize Synthetics on Agronomic Performance, Carotenoid Content, Combining Ability and Heterosis. Agronomy 2020, 10, 1625. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, R.B. Eye Nutrition in Context: Mechanisms, Implementation, and Future Directions. Nutrients 2013, 5, 2483–2501. [Google Scholar] [CrossRef]
- Ribaya-Mercado, J.D.; Blumberg, J.B. Lutein and Zeaxanthin and Their Potential Roles in Disease Prevention. J. Am. Coll. Nutr. 2004, 23, 567S–587S. [Google Scholar] [CrossRef]
- Gammone, M.A. Prevention of Cardiovascular Diseases with Carotenoids. Front. Biosci. 2017, 9, 165–171. [Google Scholar] [CrossRef]
- Cooper, D.A. Carotenoids in Health and Disease: Recent Scientific Evaluations, Research Recommendations and the Consumer. J. Nutr. 2004, 134, 221S–224S. [Google Scholar] [CrossRef]
- Muntean, E. Carotenoids in Several Transylvanian Maize Hybrids. In Proceedings of the 1st International Electronic Conference on Plant Science; MDPI: Basel, Switzerland, 2020; p. 8761. Available online: Sciforum.net (accessed on 21 February 2022).
- Jaswir, I.; Noviendri, D.; Hasrini, R.F.; Octavianti, F. Carotenoids: Sources, Medicinal Properties and Their Application in Food and Nutraceutical Industry. J. Med. Plants Res. 2011, 5, 7119–7131. [Google Scholar] [CrossRef]
- Kuhnen, S.; Menel Lemos, P.M.; Campestrini, L.H.; Ogliari, J.B.; Dias, P.F.; Maraschin, M. Carotenoid and Anthocyanin Contents of Grains of Brazilian Maize Landraces. J. Sci. Food Agric. 2011, 91, 1548–1553. [Google Scholar] [CrossRef]
- Koushan, K.; Rusovici, R.; Li, W.; Ferguson, L.R.; Chalam, K.V. The Role of Lutein in Eye-Related Disease. Nutrients 2013, 5, 1823–1839. [Google Scholar] [CrossRef]
- Hwang, T.; Ndolo, V.U.; Katundu, M.; Nyirenda, B.; Bezner-Kerr, R.; Arntfield, S.; Beta, T. Provitamin A Potential of Landrace Orange Maize Variety (Zea Mays L.) Grown in Different Geographical Locations of Central Malawi. Food Chem. 2016, 196, 1315–1324. [Google Scholar] [CrossRef]
- Wilson Reichert Júnior, F.; Rafael Silva de Oliveira, C.; Henrique da Silva Júnior, A.; Mulinari, J. Chemical Composition Os Maize Landraces and Their Importance to Human Health. In Proceedings of the Ciência, Tecnologia e Inovação: Do campo à mesa; Instituto Internacional Despertando Vocações, 2020. Available online: https://www.researchgate.net/publication/347665265_Chemical_Composition_Of_Maize_Landraces_And_Their_Importance_To_Human_Health (accessed on 20 April 2022).
- Tanumihardjo, S.A.; Yang, Z. Carotenoids | Epidemiology of Health Effects. In Encyclopedia of Human Nutrition, 2nd ed.; Caballero, B., Ed.; Elsevier: Oxford, UK, 2005; pp. 339–345. ISBN 978-0-12-226694-2. [Google Scholar]
- Cerhan, J.R.; Saag, K.G.; Merlino, L.A.; Mikuls, T.R.; Criswell, L.A. Antioxidant Micronutrients and Risk of Rheumatoid Arthritis in a Cohort of Older Women. Am. J. Epidemiol. 2003, 157, 345–354. [Google Scholar] [CrossRef]
- Pattison, D.J.; Symmons, D.P.; Lunt, M.; Welch, A.; Bingham, S.A.; Day, N.E.; Silman, A.J. Dietary β-Cryptoxanthin and Inflammatory Polyarthritis: Results from a Population-Based Prospective Study. Am. J. Clin. Nutr. 2005, 82, 451–455. [Google Scholar] [CrossRef]
- Zuma, M.K.; Kolanisi, U.; Modi, A.T. The Potential of Integrating Provitamin A-Biofortified Maize in Smallholder Farming Systems to Reduce Malnourishment in South Africa. Int. J. Environ. Res. Public Health 2018, 15, 805. [Google Scholar] [CrossRef]
- Zhu, C.; Naqvi, S.; Breitenbach, J.; Sandmann, G.; Christou, P.; Capell, T. Combinatorial Genetic Transformation Generates a Library of Metabolic Phenotypes for the Carotenoid Pathway in Maize. Proc. Natl. Acad. Sci. USA 2008, 105, 18232–18237. [Google Scholar] [CrossRef]
- Messias, R.D.S.; Galli, V.; Silva, S.D.D.A.e.; Rombaldi, C.V. Carotenoid Biosynthetic and Catabolic Pathways: Gene Expression and Carotenoid Content in Grains of Maize Landraces. Nutrients 2014, 6, 546–563. [Google Scholar] [CrossRef]
- Harjes, C.E.; Rocheford, T.R.; Bai, L.; Brutnell, T.P.; Kandianis, C.B.; Sowinski, S.G.; Stapleton, A.E.; Vallabhaneni, R.; Williams, M.; Wurtzel, E.T.; et al. Natural Genetic Variation in Lycopene Epsilon Cyclase Tapped for Maize Biofortification. Science 2008, 319, 330–333. [Google Scholar] [CrossRef]
- Trono, D. Carotenoids in Cereal Food Crops: Composition and Retention throughout Grain Storage and Food Processing. Plants 2019, 8, 551. [Google Scholar] [CrossRef]
- Kurilich, A.C.; Juvik, J.A. Quantification of Carotenoid and Tocopherol Antioxidants in Zea Mays. J. Agric. Food Chem. 1999, 47, 1948–1955. [Google Scholar] [CrossRef]
- Ortiz-Monasterio, J.I.; Palacios-Rojas, N.; Meng, E.; Pixley, K.; Trethowan, R.; Peña, R.J. Enhancing the Mineral and Vitamin Content of Wheat and Maize through Plant Breeding. J. Cereal Sci. 2007, 46, 293–307. [Google Scholar] [CrossRef]
- Menkir, A.; Liu, W.; White, W.S.; Maziya-Dixon, B.; Rocheford, T. Carotenoid Diversity in Tropical-Adapted Yellow Maize Inbred Lines. Food Chem. 2008, 109, 521–529. [Google Scholar] [CrossRef]
- Pillay, K.; Siwela, M.; Derera, J.; Veldman, F.J. Provitamin A Carotenoids in Biofortified Maize and Their Retention during Processing and Preparation of South African Maize Foods. J. Food Sci. Technol. 2014, 51, 634–644. [Google Scholar] [CrossRef]
- Suwarno, W.B.; Pixley, K.V.; Palacios-Rojas, N.; Kaeppler, S.M.; Babu, R. Formation of Heterotic Groups and Understanding Genetic Effects in a Provitamin A Biofortified Maize Breeding Program. Crop Sci. 2014, 54, 14–24. [Google Scholar] [CrossRef]
- Menkir, A.; Maziya-Dixon, B.; Mengesha, W.; Rocheford, T.; Alamu, E.O. Accruing Genetic Gain in Pro-Vitamin A Enrichment from Harnessing Diverse Maize Germplasm. Euphytica 2017, 213, 105. [Google Scholar] [CrossRef]
- Garg, M.; Sharma, N.; Sharma, S.; Kapoor, P.; Kumar, A.; Chunduri, V.; Arora, P. Biofortified Crops Generated by Breeding, Agronomy, and Transgenic Approaches Are Improving Lives of Millions of People around the World. Front. Nutr. 2018, 5, 12. [Google Scholar] [CrossRef]
- Muthusamy, V.; Hossain, F.; Thirunavukkarasu, N.; Choudhary, M.; Saha, S.; Bhat, J.S.; Prasanna, B.M.; Gupta, H.S. Development of β-Carotene Rich Maize Hybrids through Marker-Assisted Introgression of β-Carotene Hydroxylase Allele. PLoS ONE 2014, 9, e113583. [Google Scholar] [CrossRef]
- Liu, L.; Jeffers, D.; Zhang, Y.; Ding, M.; Chen, W.; Kang, M.S.; Fan, X. Introgression of the CrtRB1 Gene into Quality Protein Maize Inbred Lines Using Molecular Markers. Mol. Breed. 2015, 35, 154. [Google Scholar] [CrossRef]
- Duo, H.; Hossain, F.; Muthusamy, V.; Zunjare, R.U.; Goswami, R.; Chand, G.; Mishra, S.J.; Chhabra, R.; Gowda, M.M.; Pal, S.; et al. Development of Sub-Tropically Adapted Diverse Provitamin-A Rich Maize Inbreds through Marker-Assisted Pedigree Selection, Their Characterization and Utilization in Hybrid Breeding. PLoS ONE 2021, 16, e0245497. [Google Scholar] [CrossRef]
- Das, A.K.; Gowda, M.M.; Muthusamy, V.; Zunjare, R.U.; Chauhan, H.S.; Baveja, A.; Bhatt, V.; Chand, G.; Bhat, J.S.; Guleria, S.K.; et al. Development of Maize Hybrids With Enhanced Vitamin-E, Vitamin-A, Lysine, and Tryptophan Through Molecular Breeding. Front. Plant Sci. 2021, 12, 659381. [Google Scholar] [CrossRef]
- Singh, J.; Sharma, S.; Kaur, A.; Vikal, Y.; Cheema, A.K.; Bains, B.K.; Kaur, N.; Gill, G.K.; Malhotra, P.K.; Kumar, A.; et al. Marker-Assisted Pyramiding of Lycopene-ε-Cyclase, β-Carotene Hydroxylase1 and Opaque2 Genes for Development of Biofortified Maize Hybrids. Sci. Rep. 2021, 11, 12642. [Google Scholar] [CrossRef]
- Chandran, S.; Pukalenthy, B.; Adhimoolam, K.; Manickam, D.; Sampathrajan, V.; Chocklingam, V.; Eswaran, K.; Arunachalam, K.; Joikumar meetei, L.; Rajasekaran, R.; et al. Marker-Assisted Selection to Pyramid the Opaque-2 (O2) and β-Carotene (CrtRB1) Genes in Maize. Front. Genet. 2019, 10, 859. [Google Scholar] [CrossRef]
- Zunjare, R.U.; Hossain, F.; Muthusamy, V.; Baveja, A.; Chauhan, H.S.; Bhat, J.S.; Thirunavukkarasu, N.; Saha, S.; Gupta, H.S. Development of Biofortified Maize Hybrids through Marker-Assisted Stacking of β-Carotene Hydroxylase, Lycopene-ε-Cyclase and Opaque2 Genes. Front. Plant Sci. 2018, 9, 178. [Google Scholar] [CrossRef]
- Sagare, D.B.; Shetti, P.; Surender, M.; Reddy, S.S. Marker-Assisted Backcross Breeding for Enhancing β-Carotene of QPM Inbreds. Mol. Breed. 2019, 39, 31. [Google Scholar] [CrossRef]
- Yang, R.; Yan, Z.; Wang, Q.; Li, X.; Feng, F. Marker-Assisted Backcrossing of LcyE for Enhancement of ProA in Sweet Corn. Euphytica 2018, 214, 130. [Google Scholar] [CrossRef]
- Mehta, B.K.; Muthusamy, V.; Zunjare, R.U.; Baveja, A.; Chauhan, H.S.; Chhabra, R.; Singh, A.K.; Hossain, F. Biofortification of Sweet Corn Hybrids for Provitamin-A, Lysine and Tryptophan Using Molecular Breeding. J. Cereal Sci. 2020, 96, 103093. [Google Scholar] [CrossRef]
- Zhu, C.; Farré, G.; Zanga, D.; Lloveras, J.; Michelena, A.; Ferrio, J.P.; Voltas, J.; Slafer, G.; Savin, R.; Albajes, R.; et al. High-Carotenoid Maize: Development of Plant Biotechnology Prototypes for Human and Animal Health and Nutrition. Phytochem. Rev. 2018, 17, 195–209. [Google Scholar] [CrossRef]
- Naqvi, S.; Zhu, C.; Farre, G.; Ramessar, K.; Bassie, L.; Breitenbach, J.; Conesa, D.P.; Ros, G.; Sandmann, G.; Capell, T.; et al. Transgenic Multivitamin Corn through Biofortification of Endosperm with Three Vitamins Representing Three Distinct Metabolic Pathways. Proc. Natl. Acad. Sci. USA 2009, 106, 7762–7767. [Google Scholar] [CrossRef]
- Nogareda, C.; Moreno, J.A.; Angulo, E.; Sandmann, G.; Portero, M.; Capell, T.; Zhu, C.; Christou, P. Carotenoid-Enriched Transgenic Corn Delivers Bioavailable Carotenoids to Poultry and Protects Them against Coccidiosis. Plant Biotechnol. J. 2016, 14, 160–168. [Google Scholar] [CrossRef]
- Siwela, M.; Pillay, K.; Govender, L.; Lottering, S.; Mudau, F.N.; Modi, A.T.; Mabhaudhi, T. Biofortified Crops for Combating Hidden Hunger in South Africa: Availability, Acceptability, Micronutrient Retention and Bioavailability. Foods 2020, 9, 815. [Google Scholar] [CrossRef]
- Sangoi, L.; Salvador, R. Agronomic Performance of Male-Sterile and Fertile Maize Genotypes at Two Plant Populations. Ciência Rural. 1996, 26, 377–383. [Google Scholar] [CrossRef][Green Version]
- Allen, J.O. Effect of Teosinte Cytoplasmic Genomes on Maize Phenotype. Genetics 2005, 169, 863–880. [Google Scholar] [CrossRef][Green Version]
- Racz, C.; Has, I.; Has, V.; Schiop, T.; Coste, I.D. The Cytoplasm Origin Influence, the Tester Influence and the Nucleus-Cytoplasm Interactions Influence on Plant Traits for Isonuclear Lines. Available online: https://www.rjas.ro/paper_detail/648 (accessed on 21 February 2022).
- Rodrigues, L.A.; Pinho, R.G.V.; Junior, L.A.Y.B.; Souza, V.F.d.; Pereira, F.d.C.; Brito, A.H.d.; Filho, E.M.F. Influence of Cytoplasmic Genetic Male Sterility in the Grain Yield of Maize Hybrids. Afr. J. Agric. Res. 2018, 13, 2610–2617. [Google Scholar] [CrossRef][Green Version]
- Calugar, R.E.; Has, V.V.; Varga, A.; Vana, C.D.; Copandean, A.; Has, I. The Role of Cytoplasmatic Diversification on Some Productivity Traits of Maize. Euphytica 2018, 214, 90. [Google Scholar] [CrossRef]
- Kaeser, O.; Weingartner, U.; Camp, K.-H.; Chowchong, S.; Stamp, P. Impact of Different Cms Types on Grain Yield of Dent × Flint Hybrids of Maize (Zea mays L.). Maydica 2003, 48, 15–20. [Google Scholar]
- Calugar, R.; Rotar, I.; Has, V. Cytoplasmic Diversification Effect on Certain Plant Vegetative Traits Of Some Maize (Zea mays L.) Inbred Lines. Res. J. Agric. Sci. 2016, 48, 28–35. [Google Scholar]
- Calugar, R.; Rotar, I.; Haș, V.; Varga, A.; Vana, C.; Copândean, A.; Racz, I.; Haș, I. The Influence of Cytoplasmic Diversification on Some Plant Traits in Maize (Zea Mays). ProEnvironment 2017, 10, 26–32. [Google Scholar]
- Coste, I.D.; Has, I.; Schiop, T.; Chicinaş, C.; Ifrim, S. Genic, Cytoplasmatic And Nucleo-Cytoplasmatic Interactions Involved In Protein Content Determinism In A Series Of Isonuclear Inbred Corn Lines. Res. J. Agric. Sci. 2011, 43, 161–167. [Google Scholar]
- Schiop, T.; Haş, I.; Haş, V.; Coste, I.D.; Chicinaş, C.; Tritean, N. Phenotipic And Genetic Value Of Isonuclear Inbred Lines Of Maize. V. Phenotipic And Genetic Study Of Starch Content. An. INCDA Fundulea 2011, 79, 225–236. [Google Scholar]
- Ostry, V.; Ovesna, J.; Skarkova, J.; Pouchova, V.; Ruprich, J. A Review on Comparative Data Concerning Fusarium Mycotoxins in Bt Maize and Non-Bt Isogenic Maize. Mycotoxin Res. 2010, 26, 141–145. [Google Scholar] [CrossRef]
- Sopterean, L.; Valean, A.-M.; Calugar, R.; Muresanu, F.; Puia, C.; Suciu, A.L. Study On The Isonuclear Inbred Lines Reaction Under Natural Infection Conditions With Fusarium Spp. Rom. Agric. Res. 2018, 35, 121–128. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Varieties | Zeaxanthin | Lutein | β-Cryptoxanthin | β-Carotene | Reference |
|---|---|---|---|---|---|
| 44 sweet and dent inbred lines | 0.01–7.7 | 0.0–27.5 | 0.07–2.4 | 0.07–7.6 | [27] |
| Over 1000 improved genotypes and 400 landraces | 0.38–34.88 | 1.33–32.31 | 0.0–6.13 | 0.0–5.81 | [28] |
| 288 inbred lines (204 yellow lines) | 0.76–43.9 | 0.0–31.0 | 0.16–10.8 | 0.07–13.6 | [25] |
| 430 Tropical adapted inbred lines | 0.3–21.5 | 0.4–19 | 0.3–4.3 | 0.3–4.3 | [29] |
| 26 landraces | 0.07–10.7 | 0.03–3.69 | 0.01–0.10 | [15] | |
| F2—different colors | 0.09–11.8 | 1.1–19.1 | 0.01–5.4 | [30] | |
| 127 inbred lines | 2.77–14.88 | 0.01–7.22 | 1.65–9.71 | [31] | |
| 22 landraces varieties and one commercial hybrid | 1.85–26.95 | 3.50–35.30 | 0.30–13.85 | 0.30–6.10 | [24] |
| 2 landraces from 4 locations | 0.05–14.90 | 0.12–24.99 | 0.08–8.83 | [17] | |
| 477 lines (2014) and 496 lines (2016) | 0.0–42.8 | 0.0–52.9 | 0.1–9.6 | [32] |
| Cytoplasm | Total Carotenoids | Lutein | Zeaxanthin | β-Cryptoxanthin | β-Carotene |
|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |
| Original cyt | 9.70 ± 0.78 | 5.37 ± 0.37 | 1.84 ± 0.17 | 0.77 ± 0.09 | 0.39 ± 0.04 |
| Cyt T248 | 10.11 ± 0.80 | 5.37 ± 0.37 | 1.84 ± 0.14 | 0.74 ± 0.11 | 0.41 ± 0.06 |
| Cyt TB329 | 10.13 ± 0.80 | 5.58 ± 0.37 | 1.87 ± 0.13 | 0.78 ± 0.10 | 0.50 ± 0.08 *** |
| Cyt TC177 | 9.89 ± 0.74 | 5.56 ± 0.35 | 1.79 ± 0.13 | 0.71 ± 0.10 | 0.41 ± 0.08 |
| Cyt TC221 | 9.78 ± 0.64 | 5.35 ± 0.32 | 1.66 ± 0.08 0 | 0.83 ± 0.14 | 0.47 ± 0.09 *** |
| LSD 5% | 0.70 | 0.37 | 0.18 | 0.08 | 0.05 |
| LSD 1% | 0.93 | 0.50 | 0.24 | 0.11 | 0.07 |
| LSD 0.1% | 1.23 | 0.6 | 0.32 | 0.14 | 0.09 |
| Cytoplasm | Total Carotenoids | Lutein | Zeaxanthin | β-Cryptoxanthin | β-Carotene |
|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |
| TC209 | |||||
| Original cyt | 8.15 ± 0.31 | 4.67 ± 0.35 | 1.61 ± 0.14 | 0.74 ± 0.04 | 0.26 ± 0.03 |
| T248 | 10.06 ± 0.36 * | 6.31 ± 0.39 * | 1.78 ± 0.06 | 0.67 ± 0.06 | 0.27 ± 0.02 |
| TB329 | 9.23 ± 0.06 | 5.56 ± 0.25 * | 1.44 ± 0.03 | 0.75 ± 0.04 | 0.52 ± 0.04 *** |
| TC177 | 8.92 ± 0.42 | 5.32 ± 0.35 | 2.09 ± 0.04 * | 0.58 ± 0.05 | 0.25 ± 0.03 |
| TC221 | 9.42 ± 0.40 | 5.43 ± 0.35 | 1.80 ± 0.09 | 0.62 ± 0.06 | 0.23 ± 0.02 |
| TC316 | |||||
| Original cyt | 16.0 ± 0.42 | 8.1 ± 0.41 | 3.21 ± 0.30 | 1.53 ± 0.10 | 0.43 ± 0.05 |
| T248 | 15.42 ± 0.65 | 7.01 ± 0.51 0 | 2.99 ± 0.13 | 1.62 ± 0.13 | 0.76 ± 0.04 *** |
| TB329 | 15.78 ± 0.86 | 7.30 ± 0.49 | 2.85 ± 0.11 | 1.58 ± 0.14 | 0.80 ± 0.03 *** |
| TC177 | 15.32 ± 0.87 | 7.65 ± 0.32 | 2.70 ± 0.24 0 | 1.53 ± 0.08 | 0.56 ± 0.03 * |
| TC221 | 14.41 ± 0.65 0 | 7.26 ± 0.46 0 | 2.06 ± 0.13 000 | 2.04 ± 0.09 *** | 0.69 ± 0.06 *** |
| TC243 | |||||
| Original cyt | 9.39 ± 0.42 | 5.58 ± 0.19 | 1.43 ± 0.07 | 0.66 ± 0.04 | 0.28 ± 0.02 |
| T248 | 10.50 ± 0.75 | 6.13 ± 0.34 | 1.55 ± 0.10 | 0.69 ± 0.05 | 0.39 ± 0.04 * |
| TB329 | 9.08 ± 0.49 | 5.60 ± 0.52 | 1.34 ± 0.08 | 0.59 ± 0.03 | 0.26 ± 0.02 |
| TC177 | 10.09 ± 0.47 | 5.96 ± 0.57 | 1.32 ± 0.11 | 0.60 ± 0.03 | 0.31 ± 0.01 |
| TC221 | 9.19 ± 0.61 | 5.40 ± 0.47 | 1.26 ± 0.08 | 0.68 ± 0.04 | 0.38 ± 0.03 |
| TB367 | |||||
| Original cyt | 8.91 ± 0.33 | 4.99 ± 0.27 | 1.29 ± 0.11 | 0.58 ± 0.06 | 0.65 ± 0.05 |
| T248 | 9.82 ± 0.38 | 4.68 ± 0.21 | 1.42 ± 0.09 | 0.55 ± 0.05 | 0.63 ± 0.06 |
| TB329 | 11.25 ± 0.27 ** | 6.40 ± 0.46 ** | 2.10 ± 0.05 *** | 0.71 ± 0.07 | 0.93 ± 0.06 *** |
| TC177 | 9.32 ± 0.32 | 5.60 ± 0.32 | 1.25 ± 0.10 | 0.50 ± 0.04 | 0.95 ± 0.09 *** |
| TC221 | 9.66 ± 0.49 | 5.26 ± 0.23 | 1.32 ± 0.06 | 0.53 ± 0.03 | 1.08 ± 0.07 *** |
| D105 | |||||
| Original cyt | 6.06 ± 0.36 | 3.53 ± 0.34 | 1.68 ± 0.11 | 0.34 ± 0.02 | 0.31± 0.01 |
| T248 | 4.74 ± 0.12 | 2.75 ± 0.09 | 1.47 ± 0.14 | 0.19 ± 0.01 | 0± 0 000 |
| TB329 | 5.34 ± 0.29 | 3.03 ± 0.24 | 1.61 ± 0.06 | 0.29 ± 0.02 | 0± 0 000 |
| TC177 | 5.82 ± 0.42 | 3.26 ± 0.20 | 1.58 ± 0.04 | 0.37 ± 0.03 | 0± 0 000 |
| TC221 | 6.22 ± 0.39 | 3.38 ± 0.21 | 1.88 ± 0.14 | 0.28 ± 0.03 | 0± 0 000 |
| LSD 5% | 1.56 | 1.56 | 0.41 | 0.18 | 0.11 |
| LSD 1% | 2.09 | 2.09 | 0.55 | 0.24 | 0.15 |
| LSD 0.1% | 2.74 | 2.74 | 0.72 | 0.32 | 0.19 |
| Cytoplasm | Total Carotenoids | Lutein | Zeaxanthin | β-Cryptoxanthin | β-Carotene |
|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |
| Original cyt | 10.21 ± 0.44 | 5.14 ± 0.18 | 1.68 ± 0.08 | 1.27 ± 0.05 | 0.67 ± 0.04 |
| Cyt T248 | 10.54 ± 0.45 | 5.32 ± 0.20 | 1.78 ± 0.08 | 1.34 ± 0.06 | 0.68 ± 0.04 |
| Cyt TB329 | 10.40 ± 0.40 | 5.41 ± 0.16 * | 1.63 ± 0.07 | 1.30 ± 0.05 | 0.61 ± 0.04 |
| Cyt TC177 | 9.87 ± 0.38 | 5.11 ± 0.16 | 1.57 ± 0.08 0 | 1.25 ± 0.06 | 0.61 ± 0.05 |
| Cyt TC221 | 10.38 ± 0.30 | 5.42 ± 0.14 * | 1.68 ± 0.06 | 1.48 ± 0.05 *** | 0.63 ± 0.04 |
| LSD 5% | 0.47 | 0.26 | 0.10 | 0.10 | 0.07 |
| LSD 1% | 0.62 | 0.34 | 0.14 | 0.13 | 0.10 |
| LSD 0.1% | 0.80 | 0.44 | 0.17 | 0.16 | 0.12 |
| Cytoplasm | Total Carotenoids | Lutein | Zeaxanthin | β-Cryptoxanthin | β-Carotene |
|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |
| TC209 | |||||
| Original cyt | 12.43 ±1.03 | 6.73 ± 0.45 | 1.81 ± 0.13 | 1.59 ± 0.08 | 0.60 ± 0.05 |
| T248 | 13.40 ±1.17 | 7.12 ± 0.56 | 1.97 ± 0.18 | 1.71 ± 0.10 | 0.66 ± 0.07 |
| TB329 | 12.68 ±1.07 | 7.04 ± 0.34 | 1.70 ± 0.17 | 1.25 ± 0.11 00 | 0.49 ± 0.05 |
| TC177 | 12.19 ± 0.73 | 6.56 ± 0.28 | 1.84 ± 0.17 | 1.29 ± 0.12 00 | 0.46 ± 0.07 |
| TC221 | 11.41 ± 0.63 | 6.20 ± 0.27 | 1.64 ± 0.15 | 1.47 ± 0.10 | 0.60 ± 0.06 |
| TC316 | |||||
| Original cyt | 14.17 ± 0.70 | 6.32 ± 0.21 | 2.45 ± 0.17 | 1.89 ± 0.11 | 1.00 ± 0.07 |
| T248 | 13.57 ± 0.57 | 5.98 ± 0.22 | 2.36 ± 0.14 | 1.90 ± 0.13 | 0.84 ± 0.05 |
| TB329 | 13.31 ± 0.66 | 5.97 ± 0.24 | 2.26 ± 0.20 | 1.89 ± 0.12 | 0.76 ± 0.07 |
| TC177 | 11.89 ± 0.81 000 | 5.52 ± 0.31 0 | 1.95 ± 0.18 000 | 1.78 ± 0.14 | 0.85 ± 0.06 |
| TC221 | 12.24 ± 0.60 000 | 5.76 ± 0.23 | 2.01 ± 0.13 000 | 2.17 ± 0.14 ** | 0.84 ± 0.10 |
| TC243 | |||||
| Original cyt | 8.71 ± 0.49 | 4.30 ± 0.12 | 1.16 ± 0.09 | 1.07 ± 0.11 | 0.70 ± 0.08 |
| T248 | 8.29 ± 0.52 | 4.75 ± 0.23 | 1.15 ± 0.13 | 1.11 ± 0.16 | 0.67 ± 0.10 |
| TB329 | 9.82 ± 0.50 * | 5.34 ± 0.23 *** | 1.25 ± 0.10 | 1.32 ± 0.12 * | 0.62 ± 0.08 |
| TC177 | 7.86 ± 0.65 | 4.66 ± 0.32 | 0.94 ± 0.09 | 1.00 ± 0.14 | 0.54 ± 0.11 |
| TC221 | 9.64 ± 0.56 | 5.40 ± 0.24 *** | 1.26 ± 0.10 | 1.57 ± 0.12 *** | 0.58 ± 0.08 |
| TB367 | |||||
| Original cyt | 9.77 ± 0.67 | 4.98 ± 0.30 | 1.64 ± 0.13 | 1.12 ± 0.13 | 0.80 ± 0.06 |
| T248 | 10.31 ± 0.77 | 5.08 ± 0.35 | 1.77 ± 0.16 | 1.17 ± 0.15 | 0.93 ± 0.08 |
| TB329 | 9.12 ± 0.59 | 4.77 ± 0.23 | 1.53 ± 0.12 | 1.09 ± 0.13 | 0.83 ± 0.11 |
| TC177 | 9.58 ± 0.76 | 4.76 ± 0.31 | 1.56 ± 0.13 | 1.07 ± 0.15 | 0.87 ± 0.14 |
| TC221 | 10.60 ± 0.75 | 5.56 ± 0.41 * | 1.72 ± 0.15 | 1.32 ± 0.12 | 0.86 ± 0.11 |
| D105 | |||||
| Original cyt | 5.96 ± 0.32 | 3.39 ± 0.17 | 1.35 ± 0.11 | 0.68 ± 0.09 | 0.10 ± 0.05 |
| T248 | 7.13 ± 0.54 * | 3.69 ± 0.21 | 1.64 ± 0.18 * | 0.83 ± 0.12 | 0.29 ± 0.05 * |
| TB329 | 7.09 ± 0.38 * | 3.93 ± 0.19 | 1.41 ± 0.12 | 0.94 ± 0.10 * | 0.34 ± 0.06 ** |
| TC177 | 7.82 ± 0.57 *** | 4.05 ± 0.24 | 1.54 ± 0.15 | 1.10 ± 0.10 *** | 0.33 ± 0.09 ** |
| TC221 | 8.02 ± 0.30 *** | 4.19 ± 0.21 | 1.78 ± 0.11 *** | 0.86 ± 0.13 | 0.28 ± 0.02 * |
| LSD 5% | 1.06 | 0.58 | 0.23 | 0.22 | 0.16 |
| LSD 1% | 1.36 | 0.76 | 0.30 | 0.28 | 0.22 |
| LSD 0.1% | 1.79 | 0.98 | 0.39 | 0.37 | 0.28 |
| Hybrid | Original Cyt | Cyt T248 | Cyt TB329 | Cyt TC177 | Cyt TC221 |
|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |
| TC209 × TA367 | 14.38 ± 1.42 | 10.82 ± 0.80 00 | 11.33 ± 1.09 00 | 10.45 ± 0.75 000 | |
| TC209 × TC344 | 14.59 ± 1.20 | 17.36 ± 0.97 * | 11.33 ± 1.01 00 | ||
| TC209 × TC385A | 7.09 ± 0.57 | 9.22 ± 0.72 * | 9.55 ± 0.79 * | ||
| TC316 × TA367 | 16.07 ± 1.58 | 13.21 ± 1.09 00 | 13.54 ± 1.06 0 | 9.78 ± 0.52 000 | 12.66 ± 1.06 00 |
| TC316 × TA385A | 11.75 ± 1.09 | 9.00 ± 0.79 0 | |||
| TC316 × TC344 | 14.46 ± 1.09 | 11.78 ± 1.07 0 | |||
| TC243 × TA367 | 8.90 ± 0.75 | 11.30 ± 0.26 * | |||
| TC243 × TC385A | 6.88 ± 0.55 | −4.66 ± 0.38 0 | |||
| TB367 × TA367 | 10.61 ± 0.82 | 12.79 ± 0.50 * | |||
| TB367 × TC344 | 9.67 ± 0.85 | 13.17 ± 1.03 ** | |||
| TB367 × TE356 | 12.43 ± 1.17 | 10.23 ± 0.96 0 | |||
| D105 × TA367 | 5.91 ± 0.25 | 9.47 ± 0.68 ** | 8.66 ± 0.65 * | ||
| D105 × TC385A | 4.19 ± 0.21 | 8.16 ± 0.78 *** | |||
| D105 × TE356 | 7.15 ± 0.11 | 9.83 ± 0.81 * | |||
| LSD 5% | 2.11 | ||||
| LSD 1% | 2.78 | ||||
| LSD 0.1% | 3.58 |
| Hybrid | Original Cyt | Cyt T248 | Cyt TB329 | Cyt TC177 | Cyt TC221 |
|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |
| TC209 × TA367 | 8.14 ± 0.58 | 5.88 ± 0.50 000 | 6.69 ± 0.35 0 | 6.03 ± 0.54 000 | |
| TC209 × TC344 | 7.20 ± 0.65 | 5.47 ± 0.35 000 | |||
| TC209 × TC385A | 4.20 ± 0.38 | 6.33 ± 0.49 *** | 6.60 ± 0.57 *** | ||
| TC209 × TE356 | 7.37 ± 0.43 | 8.64 ± 0.74 * | |||
| TC316 × TA367 | 6.95 ± 0.50 | 4.88 ± 0.34 000 | |||
| TC316 × TC385A | 6.61 ± 0.47 | 4.74 ± 0.35 00 | 5.29 ± 0.45 0 | ||
| TC243 × TA367 | 4.30 ± 0.21 | 6.24 ± 0.27 ** | 5.80 ± 0.51 * | 5.74 ± 0.39 * | |
| TC243 × TC344 | 4.28 ± 0.13 | 5.54 ± 0.50 * | |||
| TC243 × TE356 | 4.50 ± 0.27 | 5.69 ± 0.31 * | 5.81 ± 0.52 * | ||
| TB367 × TC344 | 4.83 ± 0.36 | 7.09 ± 0.66 *** | |||
| D105 × TA367 | 3.34 ± 0.15 | 5.01 ± 0.42 ** | |||
| D105 × TC385A | 2.62 ± 0.21 | 4.59 ± 0.40 *** | |||
| LSD 5% | 1.15 | ||||
| LSD 1% | 1.52 | ||||
| LSD 0.1% | 1.96 |
| Hybrid | Original Cyt | Cyt T248 | Cyt TB329 | Cyt TC177 | Cyt TC221 |
|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |
| TC209 × TA367 | 2.18 ± 0.19 | 1.35 ± 0.11 00 | 1.69 ± 0.18 0 | 1.42 ± 0.13 00 | |
| TC209 × TC344 | 1.99 ± 0.11 | 2.74 ± 0.25 ** | |||
| TC316 × TA367 | 2.88 ± 0.23 | 2.01 ± 0.18 000 | 1.5 ± 0.13 000 | 1.96 ± 0.13 000 | |
| TC316 × TC344 | 2.42 ± 0.22 | 1.92 ± 0.08 0 | |||
| TC243 × TE356 | 1.67 ± 0.10 | 1.17 ± 0.08 0 | |||
| TB367 × TA367 | 1.63 ± 0.16 | 2.12 ± 0.21 * | |||
| TB367 × TC344 | 1.61 ± 0.13 | 2.17 ± 0.15 * | |||
| D105 × TA367 | 1.03 ± 0.09 | 1.59 ± 0.15 * | |||
| D105 × TC385A | 0.93 ± 0.09 | 1.64 ± 0.16 ** | |||
| D105 × TE356 | 1.97 ± 0.08 | 2.49 ± 0.20 * | |||
| LSD 5% | 0.46 | ||||
| LSD 1% | 0.61 | ||||
| LSD 0.1% | 0.78 |
| Hybrid | Original Cyt | Cyt T248 | Cyt TB329 | Cyt TC177 | Cyt TC221 |
|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |
| TC209 × TA367 | 1.31 ± 0.16 | 0.58 ± 0.17 00 | 0.43 ± 0.24 000 | ||
| TC209 × TC344 | 3.10 ± 0.15 | 2.48 ± 0.22 00 | 1.95 ± 0.15 000 | 2.04 ± 0.19 000 | |
| TC209 × TE356 | 1.13 ± 0.21 | 1.68 ± 0.17 * | 1.68 ± 0.15 * | ||
| TC316 × TC344 | 2.78 ± 0.18 | 3.28 ± 0.13 * | 3.23 ± 0.20 * | ||
| TC316 × TC385A | 1.40 ± 0.16 | 1.94 ± 0.20 * | |||
| TC316 × TE356 | 1.95 ± 0.20 | 1.41 ± 0.27 0 | |||
| TC243 × TA367 | 0.97 ± 0.17 | 1.45 ± 0.15 * | 1.49 ± 0.18 * | ||
| TC243 × TC344 | 1.91 ± 0.19 | 3.15 ± 0.29 *** | |||
| TB367 × TC344 | 1.84 ± 0.23 | 2.73 ± 0.16 *** | |||
| D105 × TE356 | 0.59 ± 0.18 | 1.12 ± 0.15 * | 1.12 ± 0.13 * | 1.38 ± 0.15 *** | |
| LSD 5% | 0.43 | ||||
| LSD 1% | 0.57 | ||||
| LSD 0.1% | 0.73 |
| Hybrid | Original Cyt | Cyt T248 | Cyt TB329 | Cyt TC177 | Cyt TC221 | ||||
|---|---|---|---|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |||||
| TC209 × TC344 | 1.0 ± 0.06 | 0.56 ± 0.09 00 | |||||||
| TC209 × TE356 | 0.40 ± 0.04 | 0.77 ± 0.05 * | |||||||
| TC316 × TC344 | 1.06 ± 0.06 | 0.68 ± 0.12 0 | |||||||
| TC316 × TC385A | 0.76 ± 0.11 | 0.37 ± 0.07 0 | |||||||
| TC316 × TE356 | 1.40 ± 0.14 | 0.82 ± 0.09 000 | |||||||
| TC243 × TA367 | 0.65 ± 0.09 | 1.08 ± 0.07 * | |||||||
| TC243 × TE356 | 1.28 ± 0.11 | 0.85 ± 0.10 0 | 0.69 ± 0.08 000 | 0.68 ± 0.04 000 | |||||
| D105 × TC344 | 0 ± 0 | 0.55 ± 0.07 ** | |||||||
| D105 × TE356 | 0.15 ± 0.01 | 0.56 ± 0.06 * | 0.49 ± 0.04 * | ||||||
| LSD 5% | 0.33 | ||||||||
| LSD 1% | 0.43 | ||||||||
| LSD 0.1% | 0.56 | ||||||||
| Hybrid | Total Carotenoids | Lutein | Zeaxanthin | β-Cryptoxanthin | β-Carotene |
|---|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |
| TC209 × TC344 (control) | 14.59 ± 1.20 | 7.2 ± 0.65 | 1.99 ± 0.11 | 3.1 ± 0.15 | 1.0 ± 0.06 |
| TC209(cyt T248) × TC344 | 17.36 ± 0.97 * | 8.24 ± 0.43 | 2.75 ± 0.25 ** | 3.4 ± 0.12 | 1.03 ± 0.07 |
| TC209 × TC385A (control) | 7.09 ± 0.57 | 4.2 ± 0.38 | 1.05 ± 0.11 | 0.84 ± 0.21 | 0.33 ± 0.03 |
| TC209(cytTC221) × TC385A | 9.55 ± 0.79 * | 6.6 ± 0.57 *** | 1.12 ± 0.08 | 1.04 ± 0.18 | 0.44 ± 0.03 |
| TC209 × TE356 (control) | 13.64 ± 1.35 | 7.37 ± 0.43 | 2.03 ± 0.20 | 1.13 ± 0.21 | 0.4 ± 0.04 |
| TC209(cyt T248) × TE356 | 15.71 ± 1.51 | 8.64 ± 0.74 * | 2.05 ± 0.15 | 1.5 ± 0.16 | 0.74 ± 0.05 * |
| TC209(cyt TB329) × TE356 | 15.64 ± 1.37 | 8.32 ± 0.49 | 2.05 ± 0.21 | 1.16 ± 0.17 | 0.51 ± 0.02 |
| TC209(cyt TC177) × TE356 | 13.86 ± 1.29 | 7.39 ± 0.53 | 2.31 ± 0.26 | 1.68 ± 0.17 ** | 0.62 ± 0.09 |
| TC243 × TA367 (control) | 8.90 ± 0.75 | 4.3 ± 0.21 | 0.95 ± 0.08 | 0.97 ± 0.17 | 0.65 ± 0.09 |
| TC243(cyt TB329) × TA367 | 11.3 ± 0.26* | 6.24 ± 0.27 ** | 1.39 ± 0.12 | 1.45 ± 0.15 * | 0.82 ± 0.08 |
| TC243 × TC344 (control) | 8.12 ± 0.52 | 4.28 ± 0.13 | 0.97 ± 0.10 | 1.91 ± 0.19 | 0.56 ± 0.07 |
| TC243(cyt TB329) × TC344 | 10.12 ± 0.70 | 4.9 ± 0.40 | 1.18 ± 0.04 | 1.98 ± 0.18 | 0.66 ± 0.09 |
| TC243(cyt TC221) × TC344 | 10.12 ± 0.91 | 5.54 ± 0.50 * | 1.13 ± 0.05 | 3.15 ± 0.29 *** | 0.81 ± 0.05 |
| TB367 × TC344 (control) | 9.67 ± 0.85 | 4.83 ± 0.36 | 1.61 ± 0.13 | 1.84 ± 0.23 | 0.68 ± 0.03 |
| TB367(cyt TB329) × TC344 | 10.39 ± 0.63 | 5.46 ± 0.45 | 1.77 ± 0.13 | 1.95 ± 0.21 | 0.69 ± 0.07 |
| TB367(cyt TC221) × TC344 | 13.16 ± 1.03 ** | 7.09 ± 0.66 *** | 2.17 ± 0.15 * | 2.73 ± 0.16 *** | 0.83 ± 0.03 |
| TB367 × TE356 (control) | 12.43 ± 1.17 | 5.81 ± 0.50 | 2.26 ± 0.18 | 1.02 ± 0.20 | 1.37 ± 0.08 |
| TB367(cyt T248) × TE356 | 12.72 ± 1.08 | 6.31 ± 0.39 | 2.35 ± 0.23 | 1.16 ± 0.32 | 1.44 ± 0.09 |
| D105 × TA367 (control) | 5.91 ± 0.25 | 3.34 ± 0.15 | 1.03 ± 0.09 | 0.62 ± 0.21 | 0.43 ± 0.03 |
| D105(cyt TC177) × TA367 | 9.47 ± 0.68 ** | 5.01 ± 0.42 ** | 1.48 ± 0.29 | 1.04 ± 0.17 | 0.67 ± 0.10 |
| D105 × TC344 (control) | 6.59 ± 0.49 | 3.75 ± 0.29 | 1.48 ± 0.09 | 1.2 ± 0.17 | 0 ± 0 |
| D105(cyt TB329) × TC344 | 7.52 ± 0.66 | 3.97 ± 0.37 | 1.59 ± 0.16 | 1.31 ± 0.18 | 0.55 ± 0.07 *** |
| D105 × TC385A (control) | 4.19 ± 0.21 | 2.62 ± 0.21 | 0.93 ± 0.09 | 0.27 ± 0.15 | 0 ± 0 |
| D105(cyt TB329) × TC385A | 5.06 ± 0.28 | 3.06 ± 0.15 | 0.96 ± 0.06 | 0.47 ± 0.14 | 0.19 ± 0.02 |
| D105(cyt TC177) × TC385A | 5.11 ± 0.42 | 2.95 ± 0.22 | 1.04 ± 0.06 | 0.6 ± 0.19 | 0.08 ± 0.01 |
| D105(cyt TC221) × TC385A | 8.16 ± 0.78 *** | 4.96 ± 0.40 *** | 1.64 ± 0.16 ** | 0.68 ± 0.22 | 0.17 ± 0.01 |
| D105 × TE356 (control) | 7.15 ± 0.11 | 3.86 ± 0.36 | 1.97 ± 0.08 | 0.59 ± 0.18 | 0.15 ± 0.01 |
| D105(cyt T248) × TE356 | 9.1 ± 0.21 | 4.31 ± 0.10 | 2.49 ± 0.20 * | 1.12 ± 0.15 * | 0.56 ± 0.06 * |
| D105(cyt TC177) × TE356 | 9.82 ± 0.81 ** | 4.36 ± 0.30 | 2.35 ± 0.11 | 1.39 ± 0.15 *** | 0.48 ± 0.04 * |
| LSD 5% | 2.11 | 1.15 | 0.46 | 0.43 | 0.33 |
| LSD 1% | 2.78 | 1.52 | 0.61 | 0.57 | 0.43 |
| LSD 0.1% | 3.58 | 1.96 | 0.78 | 0.73 | 0.56 |
| Inbred Line | Kernel Type | Kernel Color | Cob Color | |
|---|---|---|---|---|
| Nucleus donor | TC 209 | dent × flint | dark yellow | red |
| TC 316 | dent × flint | dark yellow | red | |
| TC 243 | dent | yellow | white | |
| TB 367 | flint | dark yellow | red | |
| D 105 | flint | yellow | white | |
| Cytoplasm donor | T 248 | dent | yellow | red |
| TB 329 | dent | dark yellow | red | |
| TC 177 | flint | dark yellow | white | |
| TC 221 | flint | orange | white | |
| Tester | TA 367 | flint × dent | yellow | red |
| TC 344 | dent × flint | dark yellow | red | |
| TC 385A | dent | light tallow | white | |
| TE 356 | dent × flint | dark yellow | red | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calugar, R.E.; Muntean, E.; Varga, A.; Vana, C.D.; Has, V.V.; Tritean, N.; Ceclan, L.A. Improving the Carotenoid Content in Maize by Using Isonuclear Lines. Plants 2022, 11, 1632. https://doi.org/10.3390/plants11131632
Calugar RE, Muntean E, Varga A, Vana CD, Has VV, Tritean N, Ceclan LA. Improving the Carotenoid Content in Maize by Using Isonuclear Lines. Plants. 2022; 11(13):1632. https://doi.org/10.3390/plants11131632
Chicago/Turabian StyleCalugar, Roxana Elena, Edward Muntean, Andrei Varga, Carmen Daniela Vana, Voichita Virginia Has, Nicolae Tritean, and Loredana Anca Ceclan. 2022. "Improving the Carotenoid Content in Maize by Using Isonuclear Lines" Plants 11, no. 13: 1632. https://doi.org/10.3390/plants11131632
APA StyleCalugar, R. E., Muntean, E., Varga, A., Vana, C. D., Has, V. V., Tritean, N., & Ceclan, L. A. (2022). Improving the Carotenoid Content in Maize by Using Isonuclear Lines. Plants, 11(13), 1632. https://doi.org/10.3390/plants11131632