
A Collection of Melon (Cucumis melo) Fruit Cultivars with Varied Skin Appearances Provide Insight to the Contribution of Suberin in Periderm Formation and Reticulation



Abstract
:1. Introduction
2. Results
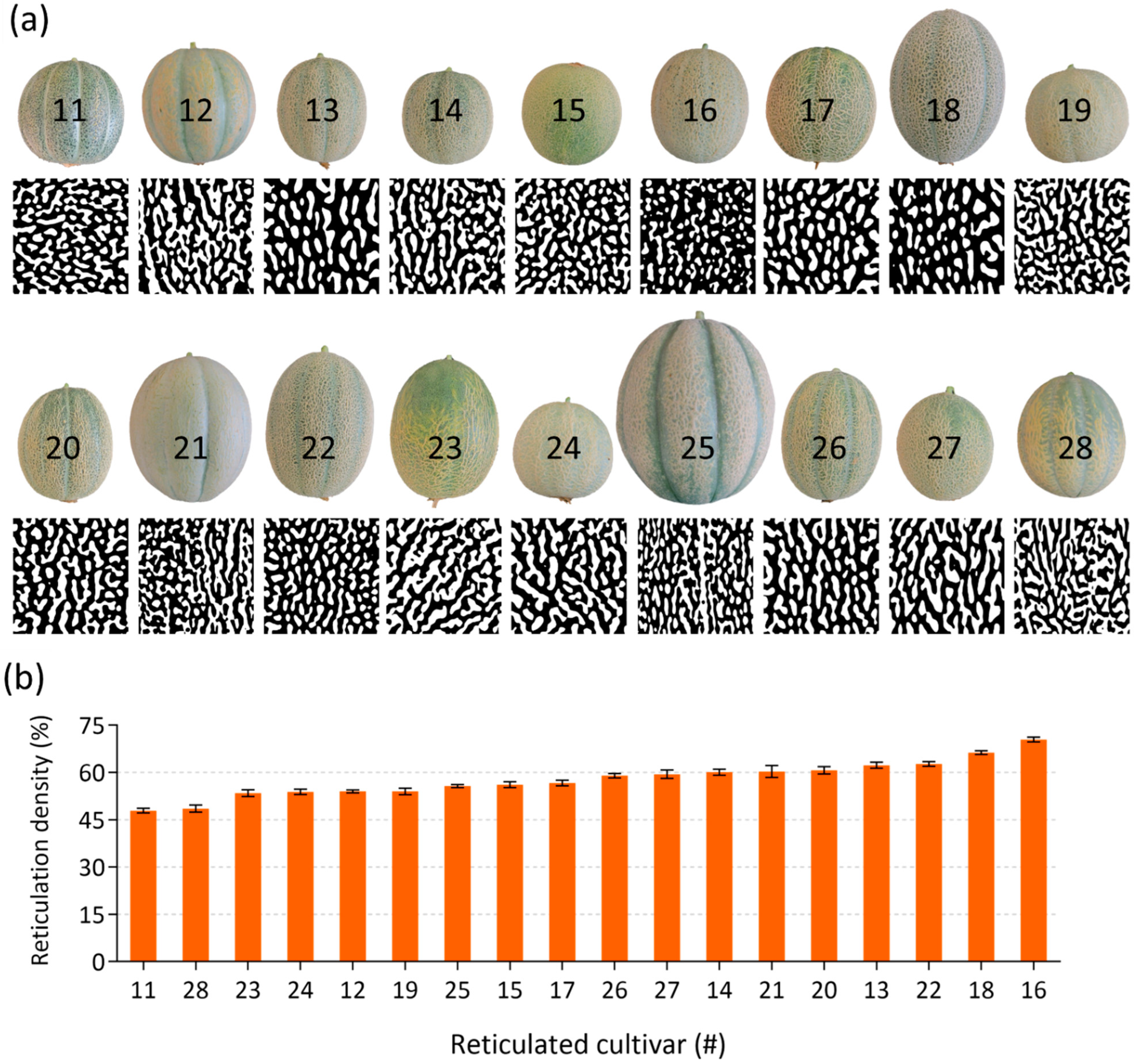
2.1. The Utilization of Melon (Cucumis melo) Fruit Cultivars with Varied Skin Appearances to Study the Contribution of Suberin in Periderm Formation and Reticulation
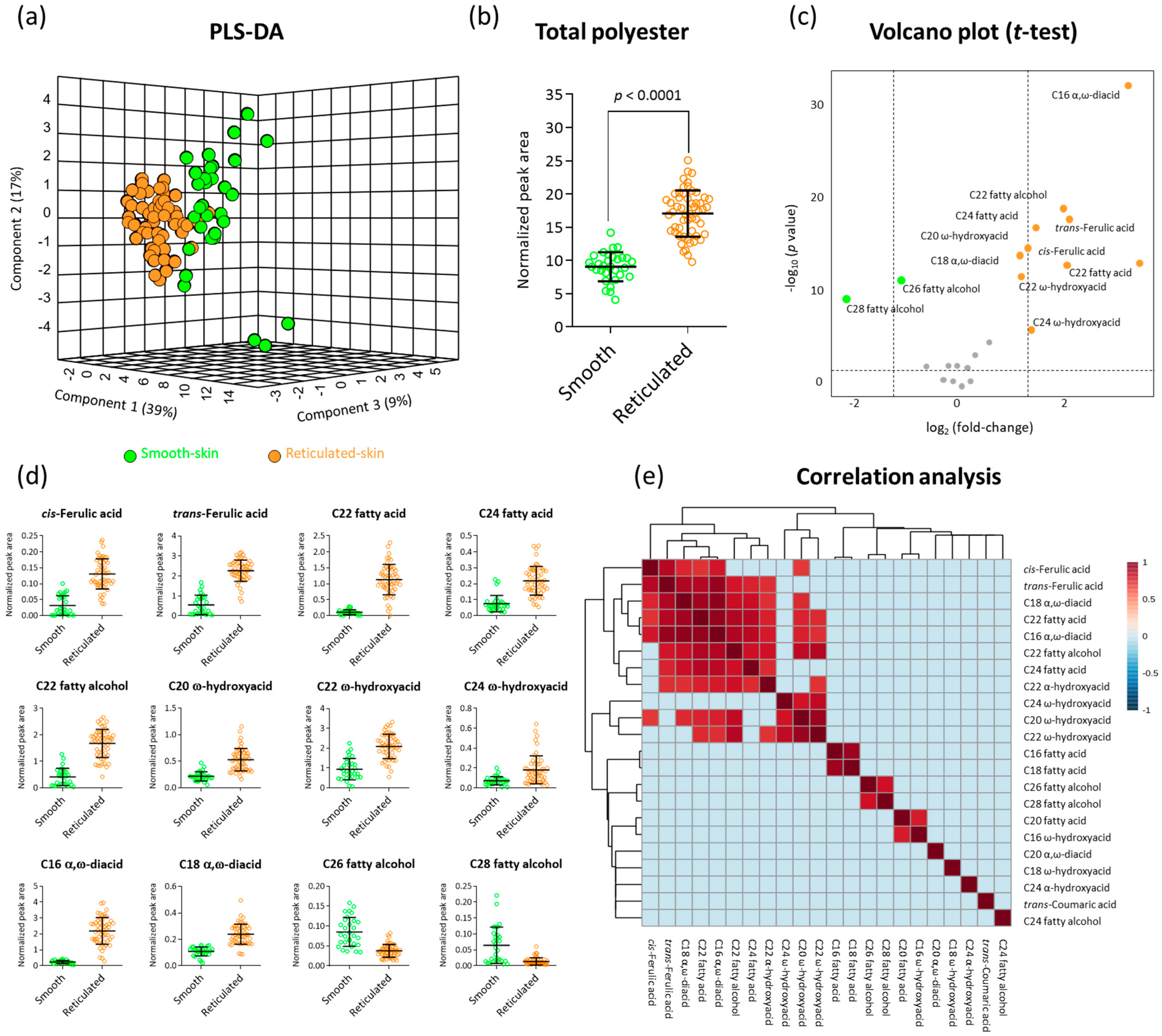
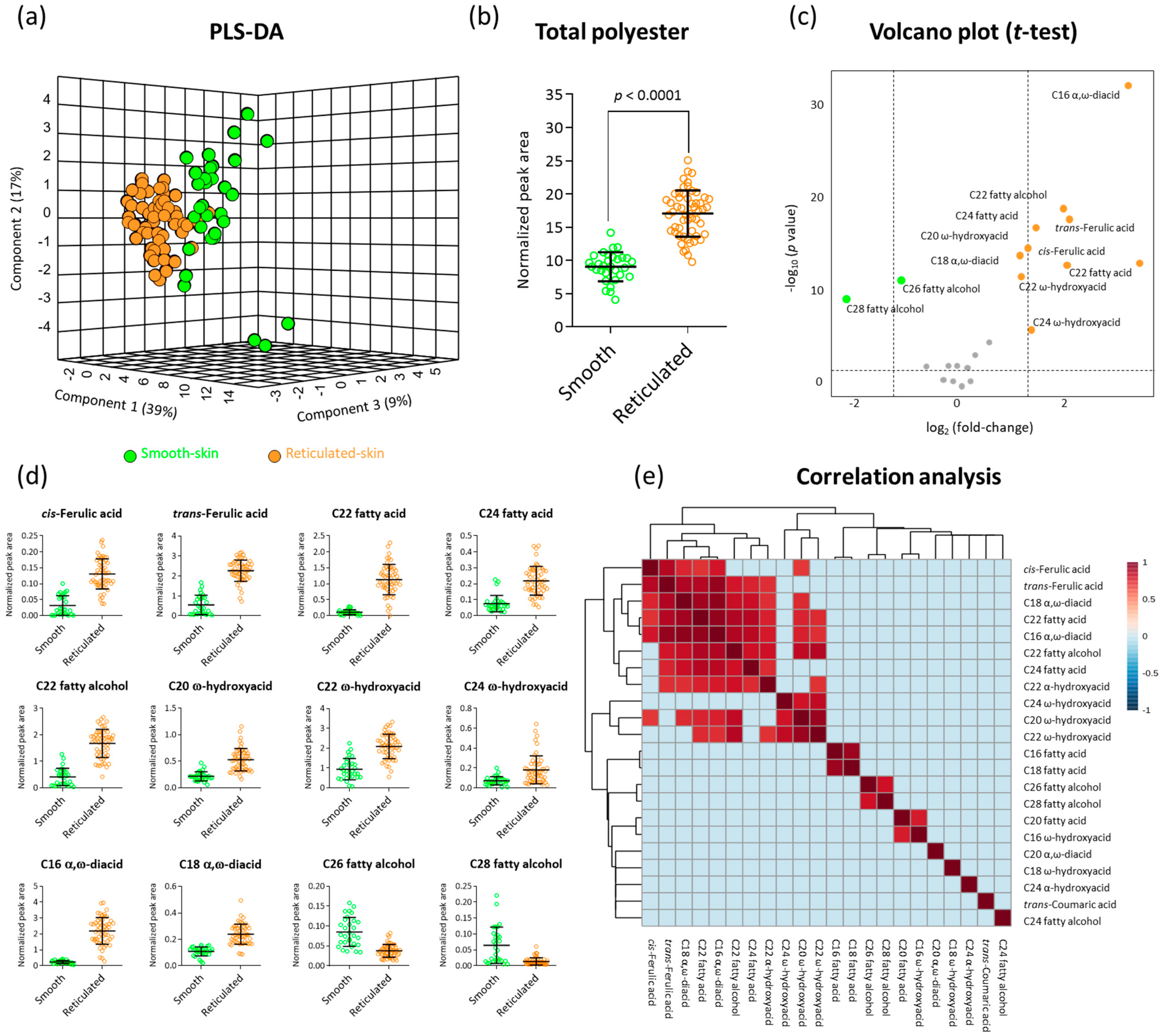
2.2. GC-MS Polyester Analysis Highlights Fundamental Chemical Differences between Skin Tissues of Smooth- and Reticulated-Skin Melon Fruit
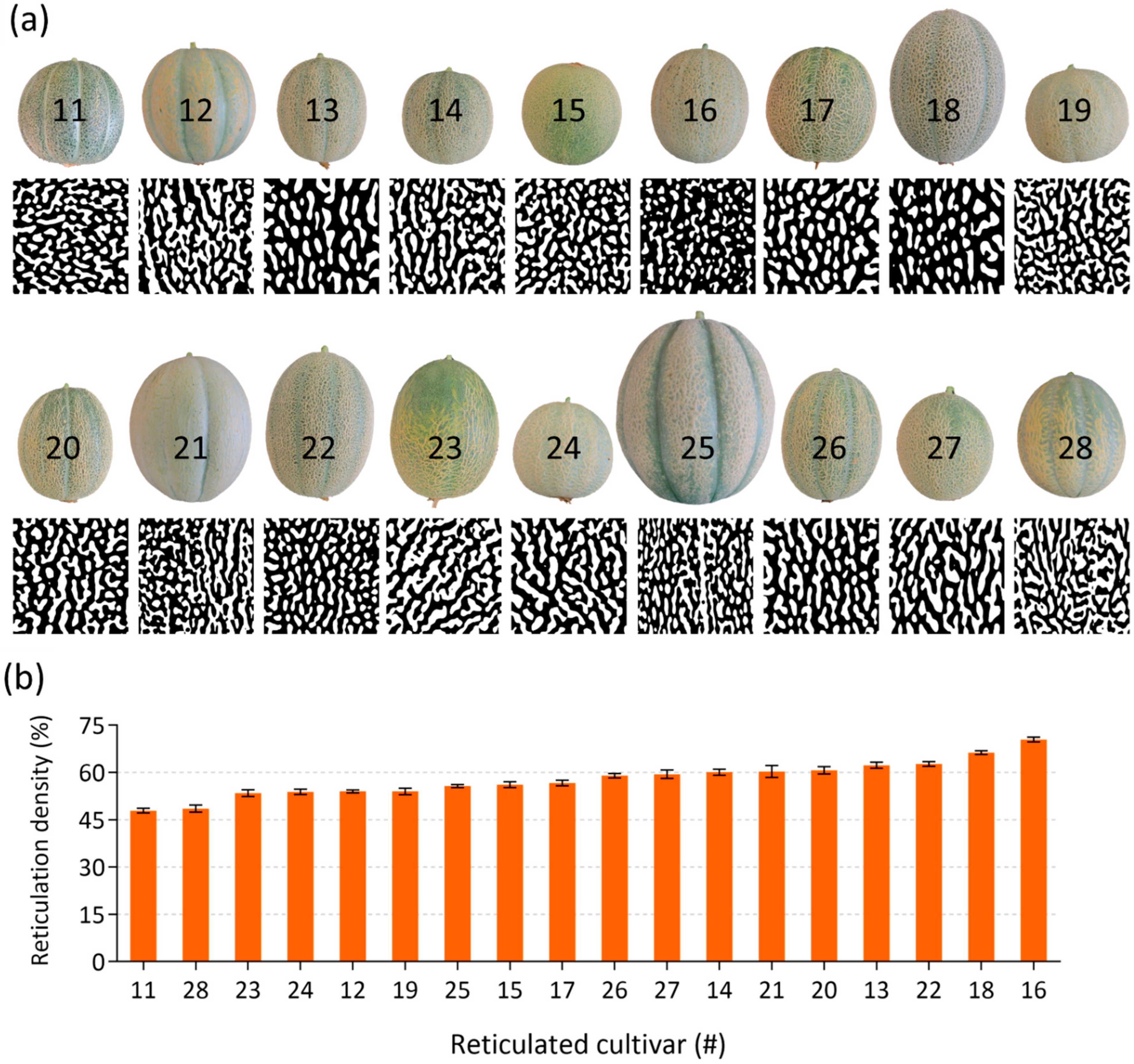
2.3. Image Analysis and Reticulation Density Calculations in Skin of Reticulated Fruit
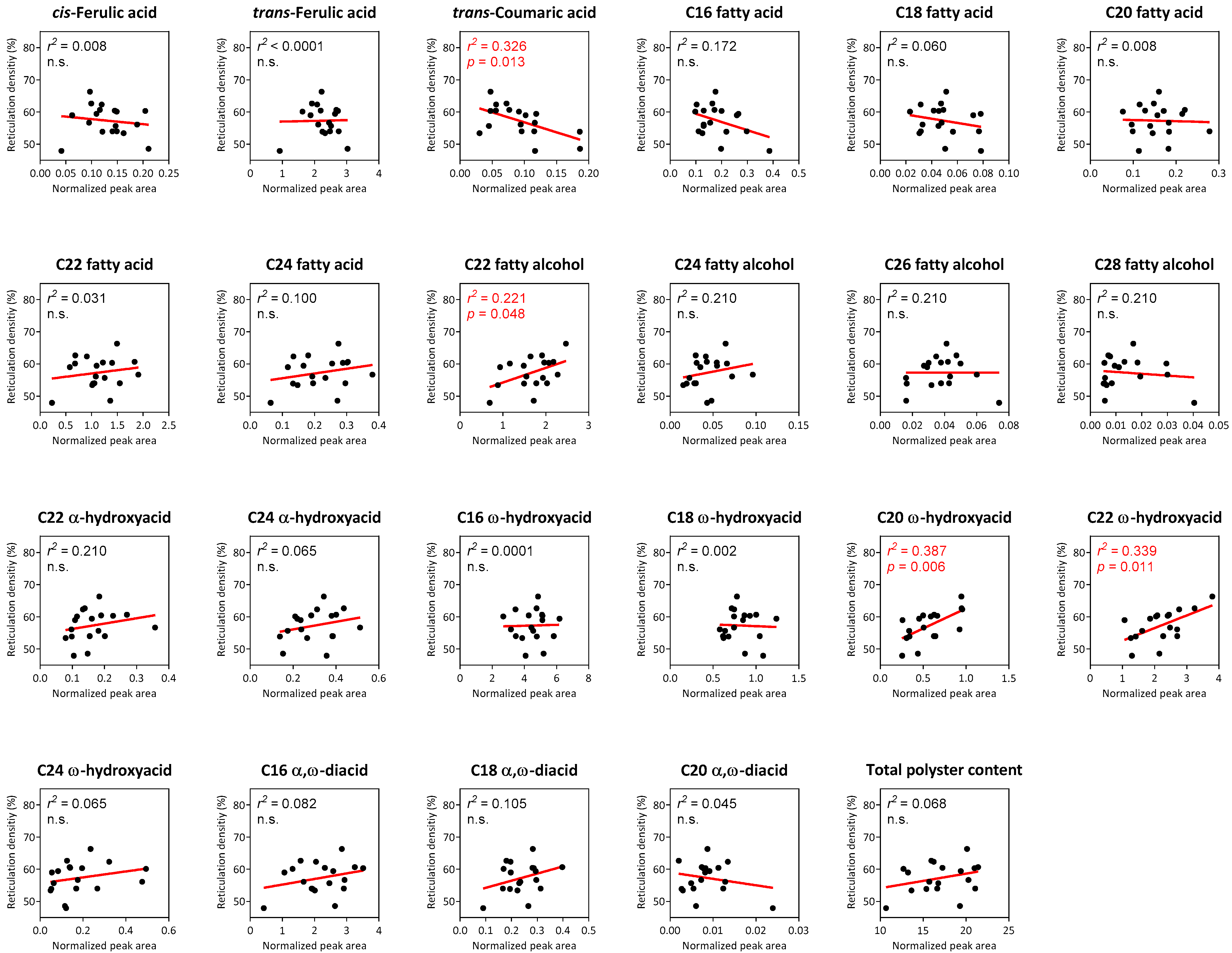
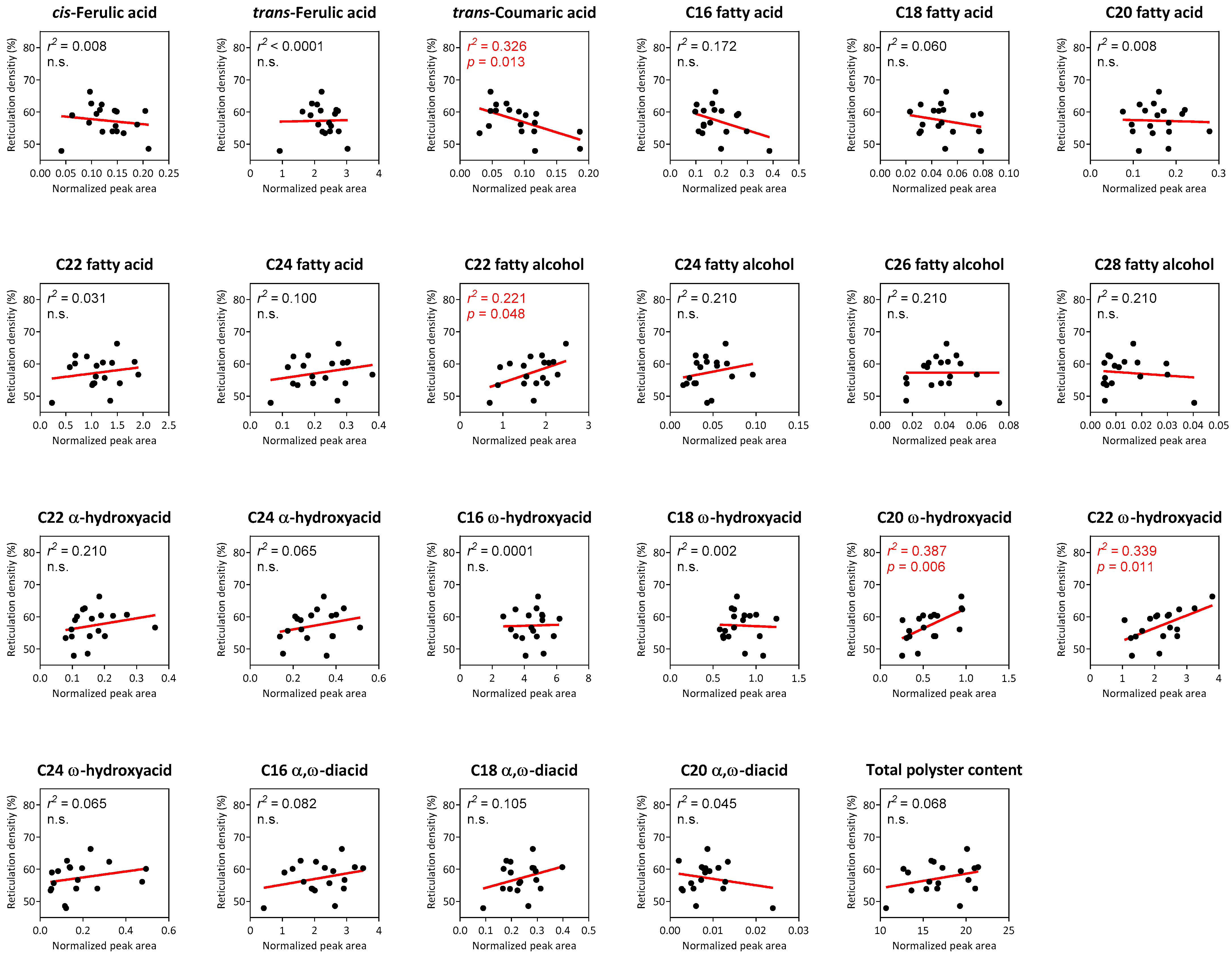
2.4. Correlation Analyses between Suberin Monomers and Reticulation Density in Reticulated-Skin Melon Cultivars
3. Discussion
4. Materials and Methods
4.1. Plant Material and Cultivation Conditions
4.2. Polyester Analysis via GC-MS
4.3. Image Analysis and Calculations of Reticulation Densities
4.4. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Monforte, A.J.; Diaz, A.; Cano-Delgado, A.; van der Knaap, E. The genetic basis of fruit morphology in horticultural crops: Lessons from tomato and melon. J. Exp. Bot. 2014, 65, 4625–4637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meissner, F. Die Korkbildung der Fruechte von Aesculusund Cucumis-Arten. Osstereichsche Bot. Z. 1952, 99, 606–624. [Google Scholar] [CrossRef]
- Cutler, E.G. Plant Anatomy: Experiment and Interpretation; Addison-Wesely: Menlo Park, CA, USA, 1969. [Google Scholar]
- Webster, B.D.; Craig, M.E. Net morphogenesis and characteristics of the surface of melon fruit. J. Am. Soc. Hortic. Sci. 1976, 101, 412–415. [Google Scholar]
- Keren-Keiserman, A.; Tanami, Z.; Shoseyov, O.; Ginzberg, I. Rind characteristics associated with melon (Cucumis melo) netting: Comperative study with smoothed-rind vareities. J. Hortic. Sci. Biotechnol. 2004, 79, 107–113. [Google Scholar] [CrossRef]
- Rose, J.K.C.; Hadfield, K.A.; Labavitch, J.M.; Bennett, A.B. Temporal sequence of cell wall disassembly in rapidly ripening melon fruit. Plant Physiol. 1998, 117, 345–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos-Santos, N.; Jimenez-Araujo, A.; Rodriguez-Arcos, R.; Fernandez-Trujillo, J.P. Cell wall polysaccharides of near-isogenic lines of melon (Cucumis melo L.) and their inbred parentals which show differential flesh firmness or physiological behavior. J. Agric. Food Chem. 2011, 59, 7773–7784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puthmee, T.; Takahashi, K.; Sugawara, M.; Kawamata, R.; Motomura, Y.; Nishizawa, T.; Aikawa, T.; Kumpoun, W. The role of net development as a barrier to moisture loss in netted melon fruit (Cucumis melo L.). HortScience 2013, 48, 1463–1469. [Google Scholar] [CrossRef]
- Saladié, M.; Matas, A.J.; Isaacson, T.; Jenks, M.A.; Goodwin, S.M.; Niklas, K.J.; Xiaolin, R.; Labavitch, J.M.; Shackel, K.A.; Fernie, A.R.; et al. A re-evaluation of the key factors that influence tomato fruit softening and integrity. Plant Physiol. 2007, 144, 1012–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, H.; Dong, Y.; Szymanski, J.; Lashbrooke, J.; Meir, S.; Almekias-Siegl, E.; Zeisler-Diehl, V.V.; Schreiber, L.; Aharoni, A. A multilevel study of melon fruit reticulation provides insight into skin ligno-suberizattion hallmarks. Plant Physiol. 2019, 179, 1486–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arya, G.C.; Dong, Y.; Heinig, U.; Shahf, N.; Kazachkova, Y.; Aviv-Sharon, E.; Nomberg, G.; Marinov, O.; Manasherova, E.; Aharoni, A.; et al. The metabolic and proteomic repertoires of periderm tissue in skin of the reticulated Sikkim cucumber fruit. Horticul. Res. 2022. [Google Scholar] [CrossRef]
- Khanal, B.P.; Grimm, E.; Knoche, M. Russeting in apple and pear: A plastic periderm replaces a stiff cuticle. AoB Plants 2013, 5, pls048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lashbrooke, J.; Aharoni, A.; Costa, F. Genome investigation suggests MdSHN3, an APETALA2-domain transcription factor gene, to be a positive regulator of apple fruit cuticle formation and an inhibitor of russet development. J. Exp. Bot. 2015, 66, 6579–6589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graca, J.; Pereira, H. Suberin structure in potato periderm: Glycerol, long-chain monomers, and glyceryl and feruloyl dimers. J. Agric. Food Chem. 2000, 48, 5476–5483. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Mao, L.; Li, J.; Chen, J.; Lu, W.; Huang, S. Abscisic acid mediates wound-healing in harvested tomato fruit. Postharvest Biol. Technol. 2016, 118, 128–133. [Google Scholar] [CrossRef]
- Han, X.; Mao, L.; Wei, X.; Lu, W. Stimulatory involvement of abscisic acid in wound suberization of postharvest kiwifruit. Sci. Hortic. 2017, 224, 244–250. [Google Scholar] [CrossRef]
- Pollard, M.; Beisson, F.; Li, Y.; Ohlrogge, J.B. Building Lipid Barriers: Biosynthesis of Cutin and Suberin. Trends Plant Sci. 2008, 13, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Arya, G.C.; Sarkar, S.; Manasherova, E.; Aharoni, A.; Cohen, H. The Plant Cuticle: An Ancient Guardian Barrier Set Against Long-Standing Rivals. Front. Plant Sci. 2021, 12, 1264. [Google Scholar] [CrossRef] [PubMed]
- Legay, S.; Guerriero, G.; Deleruelle, A.; Lateur, M.; Evers, D.; André, C.M.; Hausman, J.F. Apple russeting as seen through the RNA-seq lens: Strong alterations in the exocarp cell wall. Plant Mol. Biol. 2015, 88, 21–40. [Google Scholar] [CrossRef] [PubMed]
- Nomberg, G.; Marinov, O.; Arya, G.C.; Manasherova, E.; Cohen, H. The key enzymes in the suberin biosynthetic pathway in plants: An update. Plants 2022, 11, 392. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar Number | Cultivar Name | Skin Characteristics | Days to Mature Fruit |
|---|---|---|---|
| 1 | Banana | Smooth | 90 |
| 2 | Early Silver Line | Smooth | 76 |
| 3 | Petit Gris de Rennes | Smooth | 85 |
| 4 | Piel de Sapo | Smooth | 103 |
| 5 | Collective Farm Woman | Smooth | 83 |
| 6 | Crenshaw | Smooth | 110 |
| 7 | Green Flesh Honeydew | Smooth | 105 |
| 8 | Juane Canary | Smooth | 82 |
| 9 | Casaba Golden Beauty | Smooth | 100 |
| 10 | Tam Dew | Smooth | 100 |
| 11 | Emerald Green Gem | Reticulated | 78 |
| 12 | Golden Delicious | Reticulated | 83 |
| 13 | Green Nutmeg | Reticulated | 85 |
| 14 | Jenny Lind | Reticulated | 79 |
| 15 | Minnesota Midget | Reticulated | 70 |
| 16 | Planter’s Jumbo | Reticulated | 89 |
| 17 | Hale’s Best 36 | Reticulated | 85 |
| 18 | Hale’s Best Jumbo | Reticulated | 85 |
| 19 | Honey Rock | Reticulated | 100 |
| 20 | Rocky Ford Green Flesh | Reticulated | 89 |
| 21 | Schoon’s Hardshell | Reticulated | 88 |
| 22 | Sierra Gold | Reticulated | 90 |
| 23 | Edisto 47 Cantaloupe | Reticulated | 86 |
| 24 | Granite State | Reticulated | 83 |
| 25 | Iroquois | Reticulated | 75 |
| 26 | PMR 45 | Reticulated | 90 |
| 27 | Top Mark | Reticulated | 90 |
| 28 | Tuscany | Reticulated | 90 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manasherova, E.; Cohen, H. A Collection of Melon (Cucumis melo) Fruit Cultivars with Varied Skin Appearances Provide Insight to the Contribution of Suberin in Periderm Formation and Reticulation. Plants 2022, 11, 1336. https://doi.org/10.3390/plants11101336
Manasherova E, Cohen H. A Collection of Melon (Cucumis melo) Fruit Cultivars with Varied Skin Appearances Provide Insight to the Contribution of Suberin in Periderm Formation and Reticulation. Plants. 2022; 11(10):1336. https://doi.org/10.3390/plants11101336
Chicago/Turabian StyleManasherova, Ekaterina, and Hagai Cohen. 2022. "A Collection of Melon (Cucumis melo) Fruit Cultivars with Varied Skin Appearances Provide Insight to the Contribution of Suberin in Periderm Formation and Reticulation" Plants 11, no. 10: 1336. https://doi.org/10.3390/plants11101336
APA StyleManasherova, E., & Cohen, H. (2022). A Collection of Melon (Cucumis melo) Fruit Cultivars with Varied Skin Appearances Provide Insight to the Contribution of Suberin in Periderm Formation and Reticulation. Plants, 11(10), 1336. https://doi.org/10.3390/plants11101336