
Beneficial Effects of the Five Isolates of Funneliformis mosseae on the Tomato Plants Were Not Related to Their Evolutionary Distances of SSU rDNA or PT1 Sequences in the Nutrition Solution Production


Abstract
:1. Introduction
2. Material and Methods
2.1. Experimental Site and Design
2.1.1. Isolation and Identification of the AM Fungi
2.1.2. DNA Extraction, Amplification and Sequencing
2.1.3. Sequence and Phylogenetic Analysis
2.2. Experimental Site and Design
2.2.1. Experimental Protocol and Growing Seasons
2.2.2. AM Fungi Material and Inoculation
2.2.3. Growth Conditions of Tomato Plants
2.3. An Analysis of Biomass, Yields, Nutrients, and Colonization Rates
2.4. Statistical Analysis
3. Results
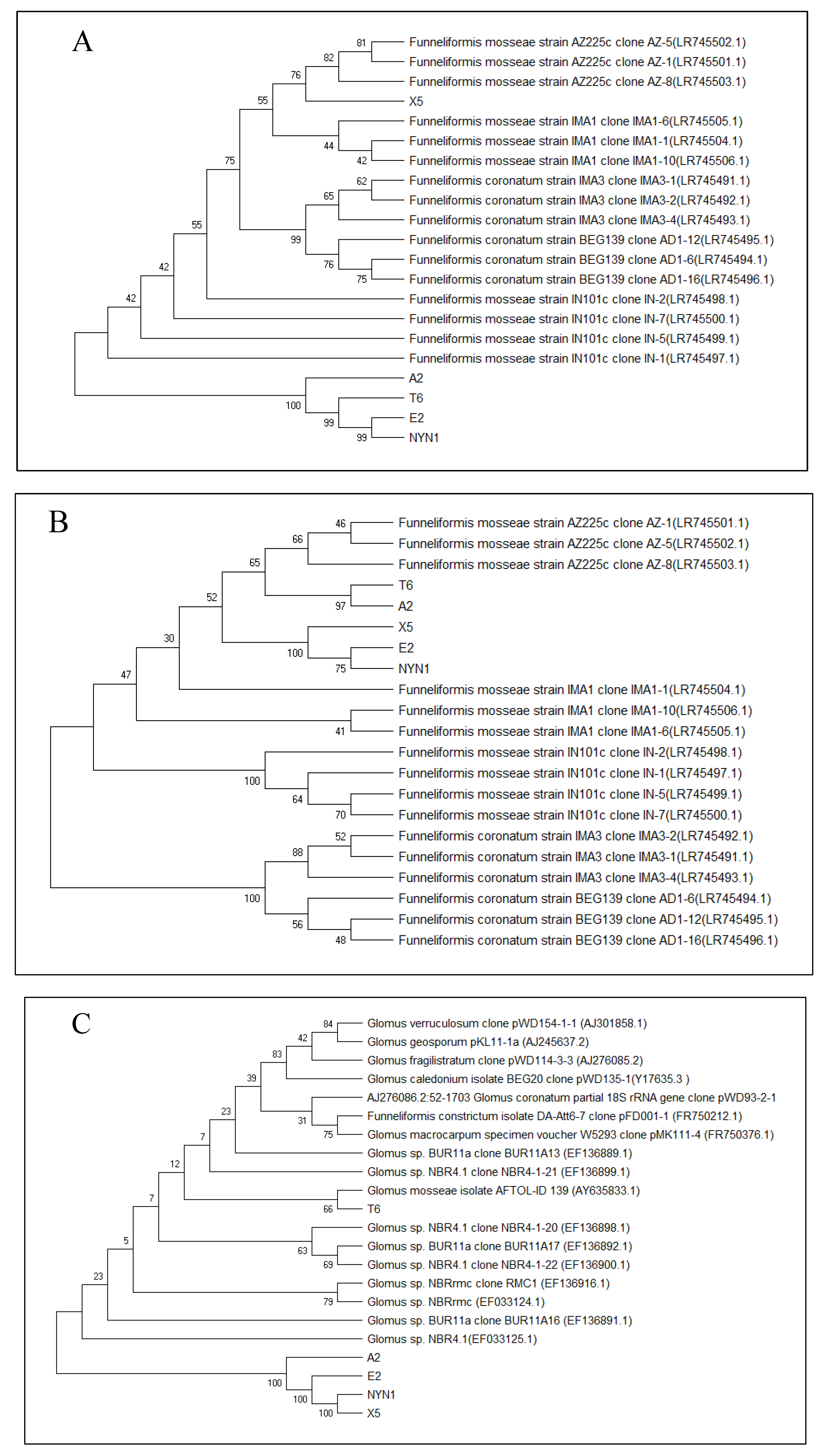
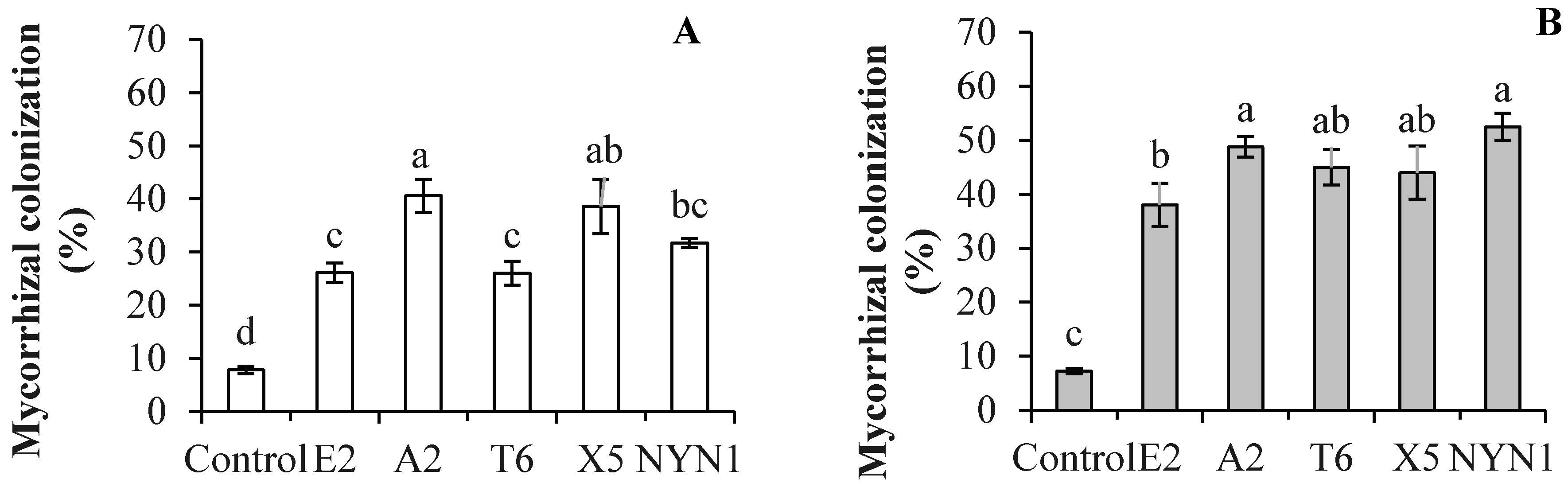
3.1. Isolate Identification and Their Root Colonization
3.2. Effects of Mycorrhizae on Plant Growth and Yields
3.3. Effects of Mycorrhizae on Plant Nutrient Uptake
4. Discussion
4.1. Mycorrhizal Colonization of Roots
4.2. Biomass and Yield Production
4.3. Nitrogen and Phosphorus Uptake
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in Plant nutrition and growth: New paradigms from cellularto eco-system scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Chen, D.; Zhang, H.; Song, Y.; Chen, H.; Tang, M. Funneliformis mosseae Enhances Root Development and Pb Phytostabilization in Robinia pseudoacacia in Pb-Contaminated Soil. Front. Microbiol. 2019, 10, 2591. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Chen, D.; Lu, K.; Sun, Z.; Zeng, R. Enhanced tomato disease resistance primed by arbuscular mycorrhizal fungus. Front. Plant Sci. 2015, 6, 786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAOSTAT. 2014. Available online: http://faostat3.fao.org/browse/Q/QC/E (accessed on 26 April 2019).
- Hariprasad, P.; Venkateswaran, G.; Niranjana, S.R. Diversity of cultivable rhizobacteria across tomato growing regions of Karnataka. Biol. Control. 2014, 72, 9–16. [Google Scholar] [CrossRef]
- Giovannetti, M.; Avio, L.; Barale, R.; Ceccarelli, N.; Cristofani, R.; Iezzi, A.; Mignolli, F.; Picciarelli, P.; Pinto, B.; Reali, D.; et al. Nutraceutical value and safety of tomato fruits produced by mycorrhizal plants. Br. J. Nutr. 2012, 107, 242–251. [Google Scholar] [CrossRef] [Green Version]
- Maboko, M.M.; Plooy, C.P.D.; Bertling, I. Comparative performance of tomato cultivars cultivated in two hydroponic pro-duction systems. S. Afr. J. Plant Soil 2011, 28, 97–102. [Google Scholar] [CrossRef]
- Kowalska, I.; Konieczny, A.; Gastol, M.; Sady, W.; Hanus-Fajerska, E. Effect of mycorrhiza and phosphorus content in nutrient solution on the yield and nutritional status of tomato plants grown on rockwool or coconut coir. Agric. Food Sci. 2015, 24, 39–51. [Google Scholar] [CrossRef]
- Mueller, A.; Franken, P.; Schwarz, D. Nutrient uptake and fruit quality of tomato colonized with mycorrhizal fungus Glomus mosseae (BEG) under deficient supply of nitrogen and phosphorus. Acta Hortic. 2009, 807, 383–388. [Google Scholar] [CrossRef]
- Maboko, M.M.; Bertling, I.; Du Plooy, C.P. Arbuscular mycorrhiza has limited effects on yield and quality of tomatoes grown under soilless cultivation. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2013, 63, 261–270. [Google Scholar] [CrossRef]
- Nedorost, L.; Pokluda, R. Effect of arbuscular mycorrhizal fungi on tomato yield and nutrient uptake under different ferti-lization levels. Acta Univ. Agric. Silvic. Mendel. Brun. 2012, 60, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Ar-buscular mycorrhizal fungi act as biostimulants in horticultural crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Azcoin, R.; Rodríguez, R.; Amora-Lazcano, E.; Ambrosano, E. Uptake and metabolism of nitrate in mycorrhizal plants as af-fected by water availability and N concentration in soil. Eur. J. Soil Sci. 2008, 59, 131–138. [Google Scholar] [CrossRef]
- Miransari, M. Arbuscular mycorrhizal fungi and nitrogen uptake. Review article. Arch. Microbiol. 2011, 193, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Rouphael, Y.; Di Mattia EEl-Nakhel, C.; Cardarelli, M. Co-inoculation of Glomus intraradices and Trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Sci. Food Agric. 2015, 95, 1706–1715. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Liu, W.X.; Guo, S.X.; Liu, R.J.; Li, M. Diversity of arbuscular mycorrhizal fungi in continuous cropping soils used for pepper production. Afr. J. Microbiol. Res. 2012, 6, 2469–2474. [Google Scholar] [CrossRef]
- Ortas, I.; Sari, N.; Akpinar, C. Effect of mycorrhizal inoculation and soil fumigation on the yield and nutrient uptake of some Solanaceae crops (tomato, eggplant and pepper) under field conditions. Agric. Mediterr. 2003, 133, 249–258. [Google Scholar]
- Inculet, C.; Mihalache, G.; Sellitto, V.M.; Hlihor, R.M.; Stoleru, V. The Effects of a Microorganisms-Based Commercial Product on the Morphological, Biochemical and Yield of Tomato Plants under Two Different Water Regimes. Microorganisms 2019, 7, 706. [Google Scholar] [CrossRef] [Green Version]
- Dasgan, H.Y.; Kusvuran, S.; Ortas, I. Response of soilless grown tomato plants to arbuscular mycorrhizal fungal (Glomus fasciculatum) colonization in re-cycling and open system. Afr. J. Biotechnol. 2008, 7, 3606–3613. [Google Scholar]
- Walder, F.; Niemann, H.; Natarajan, M.; Moritz FLehmann Boller, T.; Wiemken, A. Mycorrhizal networks: Common goods of plants shared under unequal terms of trade. Plant Physiol. 2012, 159, 789–797. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.A.; Smith, S.E. How harmonious are arbuscular mycorrhizal symbioses? Inconsistent concepts reflect different mindsets as well as results. New Phytol. 2015, 205, 1381–1384. [Google Scholar] [CrossRef]
- Feddermann, N.; Finlay, R.; Boller, T.; Elfstrand, M. Functional diversity in arbuscular mycorrhiza—The role of gene expression, phosphorous nutrition and symbiotic efficiency. Fungal Ecol. 2010, 3, 1–8. [Google Scholar] [CrossRef]
- Javier, R.; Jordi, G.; Ricardo, A.; Pozo, M.J.; Flors, V. Metabolic transition in mycorrhizal tomato roots. Front. Microbiol. 2015, 6, 598. [Google Scholar] [CrossRef]
- Pozo, M.J.; Azcoin-Aguilar, C. Unraveling mycorrhiza induced resistance. Curr. Opin. Plant Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef]
- Ortas, I. Effect of mycorrhiza application on plant growth and nutrient uptake in cucumber production under field conditions. Span. J. Agric. Res. 2010, 8, S116–S122.17. [Google Scholar] [CrossRef]
- Volpe, V.; Chitarra, W.; Cascone, P.; Volpe, M.G.; Bartolini, P.; Moneti, G.; Pieraccini, G.; Di Serio, C.; Maserti, B.; Guerrieri, E.; et al. The Association with two different arbuscular mycorrhizal fungi differently affects water stress tolerance in tomato. Front. Plant Sci. 2018, 9, 1480. [Google Scholar] [CrossRef] [PubMed]
- Ngwene, B.; Gabriel, E.; George, E. Influence of different mineral nitrogen sources (NO3-N vs. NH4-N) on arbuscular my-corrhiza development and N transfer in a Glomus intraradices cowpea symbiosis. Mycorrhiza 2013, 23, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Krüger, K.; Stockinger, H.; Krüger, C.; Schüßler, A. DNA-based species level detection of Glomeromycota: One PCR primer set for all arbuscular mycorrhizal fungi. New Phytol. 2009, 183, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Sokolski, S.; Dalpé, Y.; Piché, Y. Phosphate transporter genes as reliable gene markers for the identification and discrimi-nation of arbuscular mycorrhizal fungi in the Genus Glomus. Appl. Environ. Microbiol. 2011, 77, 1888–1891. [Google Scholar] [CrossRef] [Green Version]
- Daniels, B.A.; Skipper, H.D. Methods for the recovery and quantitative estimation of propagules from soil. In Methods and Principles of Mycorrhizal Research; Schenck, N.C., Ed.; American Phytopathological Society: St. Paul, MN, USA, 1982; pp. 29–35. [Google Scholar]
- Schwarzott, D.; Schüßle, A. A simple and reliable method for SSU rRNA gene DNA extraction, amplification, and cloning from single AM fungal spores. Mycorrhiza 2001, 10, 203–207. [Google Scholar] [CrossRef]
- Giovannini, L.; Sbrana, C.; Avio, L.; Turrini, A. Diversity of a phosphate transporter gene among species and isolates of ar-buscular mycorrhizal fungi. FEMS Microbiol. Lett. 2020, 367, fnaa024. [Google Scholar] [CrossRef]
- Schüßle, A.; Walker, C. The Glomeromycota: A Species List with New Families and Genera; The Royal Botanic Garden: Edinburgh, UK; Kew, UK; Botanische Staatssammlung Munich: Munich, Germany; Oregon State University: Corvallis, OR, USA, 2010. [Google Scholar]
- Oehl, F.; de Silva, G.A.; Goto, B.T.; Sieverding, E. Glomeromycota: Three new genera and glomoid species reorganized. Mycotaxon. 2011, 116, 75–120. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K.; Mega, X. Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Circular. Calif. Agric. Exp. Stn. 1950, 347, 357–359. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N. A modified procedure for staining roots to detect VA mycorrhizas. Mycol. Res. 1989, 92, 486–488. [Google Scholar] [CrossRef]
- Brundrett, M. Mycorrhizas in natural ecosystems. In Advances in Ecological Research; Academic Press: Cambridge, MA, USA, 1991; Volume 21, pp. 171–313. [Google Scholar] [CrossRef]
- Al-Karaki, G.N. Nursery inoculation of tomato with arbuscular mycorrhizal fungi and subsequent performance under irrigation with saline water. Sci. Hortic. 2006, 109, 1–7. [Google Scholar] [CrossRef]
- Cwala, Y.; Laubscher, C.P.; Ndakidemi, P.A.; Meyer, A.H. Mycorrhizal root colonization and the subsequent host plant re-sponse of soil less grown tomato plants in the presence and absence of the mycorrhizal stimulant, Mycotech. Afr. J. Microbiol. Res. 2010, 4, 414–419. [Google Scholar] [CrossRef]
- Martín-Robles, N.; Lehmann, A.; Seco, E.; Aroca, R.; Rillig, M.C.; Milla, R. Impacts of domestication on the arbuscular mycorrhizal symbiosis of 27 crop species. New Phytol. 2018, 218, 322–334. [Google Scholar] [CrossRef] [Green Version]
- Rafique, M.; Ortas, I. Nutrient uptake-modification of different plant species in Mediterranean climate by arbuscular mycorrhizal fungi. Eur. J. Hortic. Sci. 2018, 83, 65–71. [Google Scholar] [CrossRef]
- Affokpon, A.; Coyne, D.L.; Lawouin, L.; Tossou, C.; Agbèdè, R.D.; Jozef, C. Effectiveness of native West African arbuscular mycorrhizal fungi in protecting vegetable crops against root-knot nematodes. Biol. Fertil. Soils 2011, 47, 207–217. [Google Scholar] [CrossRef]
- Subramanian, K.S.; Santhanakrishnan, P.; Balasubramanian, P. Responses of field grown tomato plants to arbuscular mycor-rhizal fungal colonization under varying intensities of drought stress. Sci. Hortic. 2006, 107, 245–253. [Google Scholar] [CrossRef]
- Sellitto, V.; Golubkina, N.; Pietrantonio, L.; Cozzolino, E.; Cuciniello, A.; Cenvinzo, V.; Florin, I.; Caruso, G. Tomato Yield, Quality, Mineral Composition and Antioxidants as Affected by Beneficial Microorganisms under Soil Salinity Induced by Balanced Nutrient Solutions. Agriculture 2019, 9, 110. [Google Scholar] [CrossRef] [Green Version]
- Salvioli, A.; Zouari, I.; Chalot, M.; Bonfante, P. The arbuscular mycorrhizal status has an impact on the transcriptome profile and amino acid composition of tomato fruit. BMC Plant Biol. 2012, 12, 44. [Google Scholar] [CrossRef] [Green Version]
- Labour, K.; Jolicoeur, M.; St-Arnaud, M. Arbuscular mycorrhizal responsiveness of in vitro tomato root lines is not related to growth and nutrient uptake rates. Can. J. Bot. 2003, 81, 645–656. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Treatment | Shoot Biomass (g plant−1) | Root Biomass (g plant−1) | Fruit Biomass (g plant−1) | Total Biomass (g plant−1) | Yield (kg m−2) |
|---|---|---|---|---|---|
| Autumn–Winter (cv. ‘Fortesa’) | |||||
| Control | 47.2 ± 1.69bc | 5.15 ± 0.15d | 45.8 ± 0.71b | 98.2 ± 2.02c | 3.09 ± 0.08b |
| Isolate E2 | 46.7 ± 2.81c | 5.55 ± 0.21cd | 49.9 ± 3.73ab | 102.2 ± 6.21bc | 3.31 ± 0.15ab |
| Isolate A2 | 55.3 ± 3.33ab | 6.24 ± 0.34ab | 55.2 ± 1.78ab | 116.8 ± 3.28a | 3.46 ± 0.05a |
| Isolate T6 | 56.7 ± 2.31a | 6.61 ± 0.17a | 46.8 ± 2.70b | 110.2 ± 1.46abc | 3.21 ± 0.05ab |
| Isolate X5 | 57.0 ± 4.03a | 5.70 ± 0.23bcd | 52.0 ± 2.32ab | 114.7 ± 3.53ab | 3.43 ± 0.11a |
| Isolate NYN1 | 57.6 ± 1.14a | 5.81 ± 0.07bc | 59.7 ± 8.47a | 123.1 ± 8.24a | 3.37 ± 0.06a |
| Spring–Summer (cv.‘Qidali’) | |||||
| Control | 84.9 ± 5.12b | 4.68 ± 0.34b | 159.9 ± 4.08b | 249.5 ± 3.43b | 13.3 ± 0.08c |
| Isolate E2 | 100.7 ± 9.30ab | 7.26 ± 1.45a | 162.2 ± 15.0ab | 270.1 ± 16.4ab | 13.6 ± 0.15bc |
| Isolate A2 | 90.4 ± 5.93ab | 5.23 ± 0.30ab | 182.0 ± 3.26a | 277.7 ± 3.99a | 14.2 ± 0.05ab |
| Isolate T6 | 106.3 ± 3.92a | 6.91 ± 0.23a | 182.2 ± 3.05a | 295.4 ± 4.65a | 14.2 ± 0.05ab |
| Isolate X5 | 90.9 ± 7.04ab | 6.00 ± 0.86ab | 177.0 ± 3.73ab | 273.9 ± 9.82ab | 14.0 ± 0.11abc |
| Isolate NYN1 | 102.2 ± 6.08ab | 7.04 ± 0.26a | 181.4 ± 1.40a | 290.6 ± 5.58a | 14.5 ± 0.06a |
| Treatment | N Concentration (mg g−1) | P Concentration (mg g−1) | ||
|---|---|---|---|---|
| Shoot | Root | Shoot | Root | |
| Autumn–Winter(cv.‘Fortesa’) | ||||
| Control | 25.9 ± 0.14b | 16.7 ± 0.34bc | 5.57 ± 0.06d | 5.10 ± 0.24d |
| Isolate E2 | 26.1 ± 0.96ab | 16.9 ± 0.77bc | 5.74 ± 0.18cd | 5.29 ± 0.08cd |
| Isolate A2 | 27.2 ± 0.62ab | 19.1 ± 0.63a | 5.78 ± 0.03bcd | 6.36 ± 0.38ab |
| Isolate T6 | 26.4 ± 0.12ab | 21.5 ± 1.61a | 6.01 ± 0.06abc | 6.65 ± 0.27a |
| Isolate X5 | 28.0 ± 0.40a | 17.2 ± 0.57bc | 6.08 ± 0.06ab | 5.85 ± 0.16bc |
| Isolate NYN1 | 26.7 ± 1.09ab | 16.4 ± 0.19c | 6.23 ± 0.17a | 5.33 ± 0.11cd |
| Spring–Summer(cv.‘Qidali’) | ||||
| Control | 25.9 ± 0.48b | 24.3 ± 1.18ab | 7.34 ± 0.04b | 5.55 ± 0.28a |
| Isolate E2 | 27.7 ± 0.87ab | 24.7 ± 0.80ab | 7.64 ± 0.12ab | 6.17 ± 0.39a |
| Isolate A2 | 28.2 ± 0.97a | 26.0 ± 0.57a | 7.75 ± 0.21ab | 5.99 ± 1.28a |
| Isolate T6 | 28.5 ± 0.62a | 25.8 ± 0.54a | 7.93 ± 0.23a | 6.07 ± 0.48a |
| Isolate X5 | 29.2 ± 0.36a | 22.4 ± 1.01b | 7.78 ± 0.11ab | 5.86 ± 0.47a |
| Isolate NYN1 | 28.3 ± 0.65a | 24.8 ± 1.02ab | 7.63 ± 0.08ab | 6.68 ± 0.11a |
| N Content | P Content | |||||
|---|---|---|---|---|---|---|
| Shoot (g plant−1) | Root (mg plant−1) | Total (g plant−1) | Shoot (mg plant−1) | Root (mg plant−1) | Total (mg plant−1) | |
| Autumn–Winter (cv. ‘Fortesa’) | ||||||
| Control | 1.22 ± 0.04b | 86.2 ± 3.51c | 1.31 ± 0.04b | 263.0 ± 10.2b | 26.2 ± 1.37c | 289.2 ± 9.37b |
| Isolate E2 | 1.23 ± 0.12b | 93.7 ± 4.43c | 1.32 ± 0.11b | 268.8 ± 21.1b | 29.4 ± 1.51bc | 298.2 ± 22.4b |
| Isolate A2 | 1.50 ± 0.08a | 118.4 ± 3.23b | 1.62 ± 0.08a | 319.8 ± 20.2a | 39.5 ± 2.42a | 359.3 ± 20.3a |
| Isolate T6 | 1.50 ± 0.06a | 141.5 ± 7.93a | 1.64 ± 0.07a | 341.0 ± 16.3a | 43.9 ± 1.20a | 384.9 ± 17.0a |
| Isolate X5 | 1.60 ± 0.11a | 97.7 ± 3.40c | 1.69 ± 0.11a | 345.8 ± 21.1a | 33.3 ± 1.23b | 379.1 ± 21.8a |
| Isolate NYN1 | 1.54 ± 0.05a | 95.4 ± 1.67c | 1.63 ± 0.05a | 358.4 ± 7.57a | 30.9 ± 0.86bc | 389.3 ± 8.05a |
| Spring–Summer (cv. ‘Qidali’) | ||||||
| Control | 2.20 ± 0.14b | 113.0 ± 6.33b | 2.31 ± 0.14b | 623.3 ± 35.6c | 26.0 ± 2.23b | 649.2 ± 36.4c |
| Isolate E2 | 2.80 ± 0.31ab | 176.3 ± 30.5a | 2.98 ± 0.33ab | 766.1 ± 59.7abc | 45.2 ± 10.8a | 811.3 ± 65.7ab |
| Isolate A2 | 2.57 ± 0.25ab | 135.9 ± 5.27ab | 2.71 ± 0.25ab | 697.3 ± 31.2bc | 31.5 ± 7.65ab | 728.9 ± 28.6bc |
| Isolate T6 | 3.04 ± 0.17a | 178.4 ± 8.11a | 3.21 ± 0.18a | 844.4 ± 49.2a | 42.0 ± 3.89ab | 886.4 ± 52.0a |
| Isolate X5 | 2.67 ± 0.24ab | 136.5 ± 24.4ab | 2.80 ± 0.26ab | 707.1 ± 56.2abc | 35.2 ± 5.49ab | 742.3 ± 59abc |
| Isolate NYN1 | 2.89 ± 0.17a | 174.2 ± 5.65a | 3.07 ± 0.17a | 781.0 ± 51.4ab | 47.0 ± 1.05a | 828.0 ± 50.8ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.; Huang, Z.; Zhang, Y.; Rui, W.; Lei, X.; Li, Z. Beneficial Effects of the Five Isolates of Funneliformis mosseae on the Tomato Plants Were Not Related to Their Evolutionary Distances of SSU rDNA or PT1 Sequences in the Nutrition Solution Production. Plants 2021, 10, 1948. https://doi.org/10.3390/plants10091948
Feng J, Huang Z, Zhang Y, Rui W, Lei X, Li Z. Beneficial Effects of the Five Isolates of Funneliformis mosseae on the Tomato Plants Were Not Related to Their Evolutionary Distances of SSU rDNA or PT1 Sequences in the Nutrition Solution Production. Plants. 2021; 10(9):1948. https://doi.org/10.3390/plants10091948
Chicago/Turabian StyleFeng, Jingyu, Zhe Huang, Yongbin Zhang, Wenjing Rui, Xihong Lei, and Zhifang Li. 2021. "Beneficial Effects of the Five Isolates of Funneliformis mosseae on the Tomato Plants Were Not Related to Their Evolutionary Distances of SSU rDNA or PT1 Sequences in the Nutrition Solution Production" Plants 10, no. 9: 1948. https://doi.org/10.3390/plants10091948
APA StyleFeng, J., Huang, Z., Zhang, Y., Rui, W., Lei, X., & Li, Z. (2021). Beneficial Effects of the Five Isolates of Funneliformis mosseae on the Tomato Plants Were Not Related to Their Evolutionary Distances of SSU rDNA or PT1 Sequences in the Nutrition Solution Production. Plants, 10(9), 1948. https://doi.org/10.3390/plants10091948