Abstract
The Epipactis helleborine (L.) Crantz group is one of the most taxonomically challenging species complexes within the genus Epipactis. Because of the exceptionally high levels of morphological variability and the ability to readily cross with other species, ninety different taxa at various taxonomic ranks have already been described within its nominative subspecies, but the taxonomic status of most of them is uncertain, widely disputed, and sometimes even irrelevant. The present review is based on results of the most recent research devoted to the E. helleborine group taxonomy. In addition, we analysed data about taxa belonging to this group presented in some research articles and monographs devoted directly to the genus Epipactis or to orchids in certain area(s). Based on the reviewed literature and data collected in four taxonomic databases available online, we propose an updated list of the 10 currently accepted taxa in the E. helleborine group (two species, six subspecies, and two varieties), which includes E. helleborine (L.) Crantz subsp. helleborine; E. helleborine subsp. bithynica (Robatsch) Kreutz; E. helleborine subsp. distans (Arv.-Touv.) R.Engel and P.Quentin; E. helleborine subsp. neerlandica (Verm.) Buttler; E. helleborine var. tangutica (Schltr.) S.C.Chen and G.H.Zhu; E. helleborine subsp. tremolsii (Pau) E.Klein; E. helleborine subsp. voethii (Robatsch) Jakubska-Busse, Żołubak, and Łobas, stat. nov.; E. condensata Boiss. ex D.P.Young; E. condensata var. kuenkeleana (Akhalk., H.Baumann, R.Lorenz, and Mosul.) Popovich; and E. cupaniana C.Brullo, D’Emerico, and Pulv.
1. Introduction
The long and turbulent history of changes in taxonomy of the genus Epipactis Zinn, 1757 (Orchidaceae) is well documented [1,2,3,4,5,6,7,8,9,10,11,12]. The most widely contested aspect of its taxonomy is whether to treat many of its morphologically different but poorly defined taxa as new species or if is more appropriate to transfer some of them to lower taxonomic ranks, such as subspecies, variety, or form [13,14]. The currently adopted classification system of Epipactis does not take sufficient account of the variation range of morphological characters within its taxa, nor does it explain the underlying sources of this variability. Moreover, the species delimitation within Epipactis is often complicated by the ease with which the interspecific hybrids are formed in the locations inside the native ranges of the parental species, the existence of which is a common cause of taxa misidentification [15,16,17,18,19,20,21]. The fluidity of the morphological boundaries between various Epipactis species causes serious difficulties in determinig the diagnostic characters useful in species identification [22,23,24]. As a result, there is still no official account of the species included within the genus Epipactis. The estimates range from a few to several dozen depending on the source (e.g., [6,17,25,26,27,28,29,30,31,32,33,34,35,36,37,38]).
The primary aim of taxonomic research is to provide a comprehensive classification system, which reflects the observed relationships between the taxa at the morphological, geographical, and genetic levels [39]. The major impediment in achieving a taxonomic consensus within the genus Epipactis is its exceptionally high morphological variability, which is still insufficiently documented and requires further clarification [11,14]. The main source of this variability, referred to the phenotypic plasticity, is manifested in natural populations of many Epipactis species through the slight differences observed in the vegetative (e.g., shoots and leaves) and generative (i.e., flowers and their separate elements) parts of the individual plants [4,15,40,41,42]. This broad spectrum of morphological variation also provided a wide range of characters that delimitate and group the species within the genus Epipactis over the past few decades (e.g., [14,17,21,23,28,29,30,31,32,43,44,45,46,47,48,49,50,51,52,53,54]). As a consequence, a large number of morphologically similar species and infraspecific taxa (i.e., subspecies, varieties, or forms), usually of local or narrowly restricted occurrence, have been described within Epipactis [17,23,31,55]. However, the taxonomy and systematic position of the majority of these taxa are still not well understood and highly debated [4,11,13,24,42,56,57,58,59].
One of these taxonomically problematic species is Epipactis helleborine (L.) Crantz, native to Eurasia and North Africa and widely naturalised in North America [27,31,32,58,60]. It is a common cross-pollinating (allogamous) orchid species with a very wide ecological amplitude. It grows in areas with nutrient rich soils and a broad pH spectrum (usually in alkaline conditions) and, unlike the other species of Epipactis, has a highly variable habitat preference. Its natural populations are usually found in forests, amongst shrubs, or in partly disturbed vegetation sites, ranging from lowland floodplains to mountain spruce forests up to the altitude of 2200 m a.s.l. [31,37,51,52,53,61,62,63]. It is also increasingly observed in the areas strongly impacted by human activity, such as roadsides, cemeteries, railway embankments, gravel pits, gardens, and urban parks [64,65].
The recently increased interest in the evolutionary history of the genus Epipactis has resulted in some significant changes in its taxonomy [11]. The most important and widely challenged one of them is considered the present division of this genus into nine morphologically distinct species complexes [66,67,68,69]. Among them, there is also a group devoted to E. helleborine, for which the circumscription has already been reorganized by numerous scientists [3,5,8,10,17,23,30,31,47,70,71]. Interestingly, the taxonomic status of individual taxa included in this group is still chaotic and in need of clarification.
Because of the general confusion concerning the taxonomy of the genus Epipactis, caused mainly by the frequent changes in its infrageneric classification, we aimed to present here an updated list of the 10 currently accepted taxa included in the E. helleborine group. As a decisive criterion for the selection of individual taxa to our circumscription, we used the results of recent genetic and morphometric analysis in relation to the total 41 taxa that have been included in this group.
2. Recent Taxonomic Publications Devoted to the Epipactis helleborine Group
So far, a number of the research articles and monographs have been published by representing the description and taxonomic treatments of taxa of the Epipactis helleborine group [3,5,8,10,17,23,30,31,47,70,71] (see Table 1). At the beginning, Tyteca and Dufrêne [47] conducted the medium-scale biostatistical study of the genus Epipactis focused on only seven allogamous species (autogamous taxa were explicitly excluded) from the south-western limit of its distribution range in Europe. But the authors concluded that at least five species (i.e., E. helleborine s.str., E. distans Arv.-Touv., E. neerlandica (Verm.) Devillers-Tersch. and Devillers, E. tremolsii Pau, and E. lusitanica D.Tyteca) should be included within the E. helleborine group. Tyteca and Dufrêne [47] also used the results of multivariate analysis of 28 carefully chosen characters of floral and vegetative morphology (particularly the differences in the flower structure) to prove that the four taxa included in this group are sufficiently different from E. helleborine s.str. and should be treated as independent species rather than at the subspecific rank.

Table 1.
Nomenclatural and taxonomic changes within the Epipactis helleborine group.
Later, the circumscription of the Epipactis helleborine group was delimited by a yet another set of clearly defined morphological characters, including the appearance of the shoot, labellum, ovary, and pedicel. However, the morphometric analysis of these characters was not as detailed as that of the other published taxonomic treatments (e.g., [23,47]). Delforge [30] divided the 23 species belonging to the E. helleborine group into three subgroups, i.e., the E. leptochila subgroup (five taxa), the E. helleborine subgroup (13 taxa), and the E. tremolsii subgroup (five taxa). Within the E. helleborine subgroup, this author included E. helleborine s.str. and 12 other morphologically similar species (Table 1). This subgroup was also further sub-divided into two additional sections: one with the cross-pollinating species and another with autogamous taxa only.
The increase in number of new taxa described within the genus Epipactis has led to some significant changes in its infrageneric classification. As a consequence, two new characters were added by Delforge [17] to the circumscription of the E. helleborine group, i.e., the leaf and the inflorescence morphology. Four of the previously used characters, i.e., the appearance of the shoot, labellum, ovary, and pedicel have also been redefined. Thus, Delforge’s newly circumscribed E. helleborine group included 13 taxa (11 at the rank of species and two varieties). Six of which were included in the author’s previous study [30], where one (i.e., E. helleborine var. youngiana A.J.Richards and A.F.Porter) Kreutz) has changed its taxonomic rank (Table 1).
Subsequently, Brullo et al. [23] have expanded the Epipactis helleborine group by including E. cupaniana C.Brullo, D’Emerico, and Pulv., a newly described endemic from the mesophilous Holm oak woods in north-central Sicily. Their circumscription of the E. helleborine group included 11 additional species and was broadly based on the system proposed by Delforge in 2006 (Table 1). The authors also conducted a morphometric analysis of a broad range of 37 characters of floral and vegetative morphology. The obtained results suggest that E. cupaniana does indeed belong to the E. helleborine group. This taxon is morphologically and karyologically different from E. helleborine s.str. and can be accepted as a separate species. Despite this conclusion, Delforge [31] did not include E. cupaniana in his latest concept of the E. helleborine group.
One year later, in 2014, Epipactis condensata subsp. kuenkeleana (Akhalk., H.Baumann, R.Lorenz, and Mosul.) Kreutz, Fateryga, and Efimov was published as a new combination for the species formerly known as E. viridiflora subsp. kuenkeleana Akhalk., H.Baumann, R.Lorenz, and Mosul. [8], where then Delforge raised this latter taxon to full species status (i.e., E. kuenkeleana (Akhalk., H.Baumann, R.Lorenz and Mosul.) P.Delforge) [72]. Thereafter, E. condensata subsp. kuenkeleana was put into synonymy with the nominative subspecies [10,70]. However, finally, in 2020, plants within the same taxon corresponding to the former subsp. kuenkeleana were considered as a phenotypic form, confined to shady forest communities, and described as E. condensata var. kuenkeleana (Akhalk., H.Baumann, R.Lorenz, and Mosul.) Popovich [70].
The most recent taxonomic treatment of the Epipactis helleborine group in Europe, North Africa, and the Middle East [31], is broadly based on an earlier account by the same author [17] and expands to comprise 17 taxa, five of which are included here for the first time, and one taxon (i.e., E. pontica Taubenheim) which is transferred to E. leptochila group (Table 1).
3. List of Names of Infraspecific Taxa in Epipactis helleborine and its Current Taxonomic Status
The seemingly endless morphological variation observed across the entire distribution range of Epipactis helleborine s.str. is clearly reflected by the list of its infraspecific taxa presented below in Table 2.
Table 2.
An overview of names of infraspecific taxa published in Epipactis helleborine.
As it turns out, in the light of the data collected in four taxonomic databases available online [73,74,75,76], as many as ninety morphologically similar taxa have been distinguished within Epipactis helleborine s.str. at various taxonomic ranks since its original description as Serapias helleborine L. by Carl Linnaeus in 1753 [33,77,78,79,80,81,82]. Among these, at the ranks of variety and subspecies have been classified respectively 37 and 36 names of taxa, and at the rank of form, 17 have been classified (Figure 1).
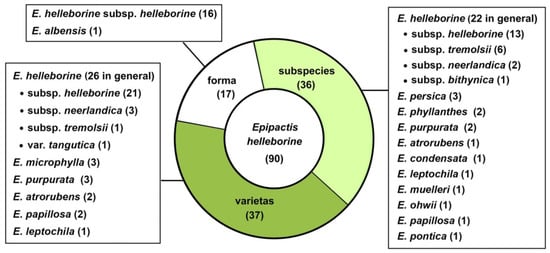
Figure 1.
The current status of names of infraspecific taxa published in Epipactis helleborine categorised according to their taxonomic ranks, based on [73,74,75,76]. The number of names synonymised with E. helleborine s.l. and with other species is shown in parentheses.
It should be noted that most of them (64) were synonymised with Epipactis helleborine s.l. almost as soon as they were published and currently only five infraspecific taxa are accepted, i.e., E. helleborine (L.) Crantz subsp. helleborine, E. helleborine subsp. bithynica (Robatsch) Kreutz, E. helleborine subsp. neerlandica (Verm.) Buttler, E. helleborine var. tangutica (Schltr.) S.C.Chen and G.H.Zhu, and E. helleborine subsp. tremolsii (Pau) E.Klein [3,63,71,83,84]. The remaining 26 published names of taxa do not currently have a taxonomic relationship with E. helleborine s.l. (Figure 2).
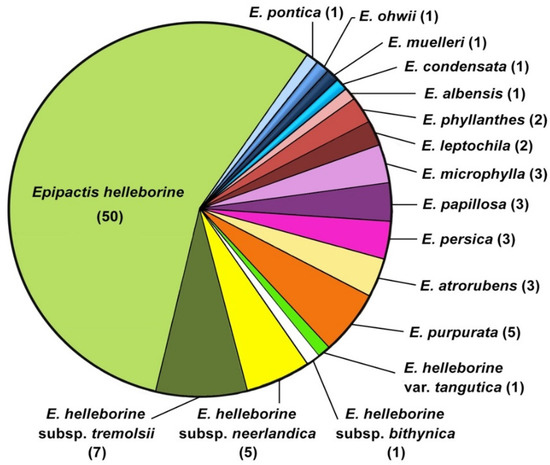
Figure 2.
The current status of names of infraspecific taxa published in Epipactis helleborine, based on [73,74,75,76]. The number of names synonymised with E. helleborine s.l. and to other species is shown in parentheses.
4. Conclusions
Since the genus Epipactis has been divided into several species complexes based on morphological characters, more than forty taxa have been classified into the E. helleborine group (see Table 1). These contain such taxa as E. danubialis Robatsch and Rydlo, E. greuteri H.Baumann & Künkele, E. halacsyi Robatsch, E. leptochila (Godfery) Godfery, E. muelleri Godfery, E. naousaensis Robatsch, E. olympica Robatsch, E. pontica or E. purpurata Sm., which, because of their distinct morphological phenotype, were excluded from it over time and (in some cases) provided a basis for effective distinguishing of other groups. Despite the fact that 15 of these taxa were originally included as separate species, they are being considered currently as three out of the five infraspecific taxa published in E. helleborine (i.e., E. helleborine subsp. helleborine, E. helleborine subsp. neerlandica, and E. helleborine subsp. tremolsii). Furthermore, two other infraspecific taxa have been genetically confirmed as well-founded, i.e., E. helleborine subsp. distans (Arv.-Touv.) R.Engel and P.Quentin and E. helleborine subsp. voethii, although the latter one still has not been officially distinguished at this rank. As it appears, E. bugacensis Robatsch and E. rhodanensis Gévaudan and Robatsch have in fact a similarly close genetic relationship with E. dunensis (T.Stephenson and T.A.Stephenson) Godfery (originally included in the E. helleborine group), which, in our opinion, due to its floral morphologies (typical of autogamous taxa) should not be classified in this group. Although E. nordeniorum Robatsch was for a long time assigned to the E. helleborine group, as a result of recent genetic analysis it turned out to be molecularly similar to E. albensis Nováková and Rydlo, classified in a separate group. Some taxa, such as E. condensata Boiss. ex D.P.Young and E. cupaniana, based on results of a detailed morphological analysis of their floral and vegetative characters, should be retained in the E. helleborine group, although these taxa are still not included there in the most recently published accounts of the genus Epipactis.
The boundaries between individual species within the Epipactis helleborine group are unclear, making it difficult to determine reliable taxonomic characters useful in the construction of an identification key which would be unambiguously interpreted by different users. In the light of the scientific literature published worldwide, especially because of the impact of the genetic research on our current understanding of the boundaries between various species of Epipactis, we think it is appropriate to maintain the E. helleborine group, but we propose to update its circumscription to better reflect the taxonomic changes summarised in Table 1 that have occurred for its individual members over the past few decades.
Our proposed taxonomic circumscription of the Epipactis helleborine group therefore consists of the following only allogamous taxa: E. helleborine subsp. helleborine, E. helleborine subsp. bithynica, E. helleborine subsp. distans, E. helleborine subsp. neerlandica, E. helleborine var. tangutica, E. helleborine subsp. tremolsii, E. helleborine subsp. voethii (Robatsch) Jakubska-Busse, Żołubak, and Łobas, stat. nov., E. condensata, E. condensata var. kuenkeleana and E. cupaniana.
Although the proposed list of taxa in the Epipactis helleborine group seems to be appropriate at the moment, we treat it as legitimate until new methods of genetic and morphometric analysis are developed, which would allow more precise definition of the Epipactis separate species concept in the future.
Proposal of a New Status for Epipactis helleborine subsp. voethii
Epipactis helleborine subsp. voethii (Robatsch) Jakubska-Busse, Żołubak, and Łobas, stat. nov.
Basionym: Epipactis voethii Robatsch, Mitteilungen der Abteilung für Botanik am Landesmuseum Joanneum in Graz 21/22: 22 (1993).
Comments: This subspecies differs from typical Epipactis helleborine s.str. through few developed clinandrium, as well as the slight differences observed in the morphological characters, i.e., the green colour of stems, leaves, and flowers of the individual plants, which are almost lacking in any violet coloration. In the fruiting stage, taxa can be distinguished by the shape of the seeds: in E. helleborine s.str. the seeds are worm-like, and club-shaped in E. helleborine subsp. voethii.
Author Contributions
Conceptualization, Z.Ł. and A.J.-B.; methodology, Z.Ł., E.Ż. and A.J.-B.; investigation, Z.Ł., E.Ż. and A.J.-B.; writing—original draft preparation, Z.Ł., A.K., E.Ż. and A.J.-B.; writing—review and editing, Z.Ł., A.K., E.Ż. and A.J.-B.; visualization, E.Ż. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Publicly available datasets were analysed in this study.
Acknowledgments
The authors wish to thank to Jacek Wajer (Natural History Museum) for improving the English version of the text.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Claessens, J.; Kleynen, J.; Wiellnga, R. Some Notes on Epipactis helleborine (L.) Crantz ssp. neerlandica (Vermeulen) Buttler and Epipactis renzii K. Robatsch. Eurorchis 1998, 10, 55–64. [Google Scholar]
- Mereďa, P.; Potůček, O. Epipactis futakii, spec. nova (Orchidaceae)—Eine Neue Kleistogam Blühende Sitter—Art aus der Slovakei. Preslia 1998, 70, 247–258. [Google Scholar]
- Chen, S.C.; Zhu, G. Nomenclatural changes in Epipactis (Orchidaceae) from China. Novon 2003, 13, 423–424. [Google Scholar] [CrossRef]
- Hollingsworth, P.M.; Squirrell, J.; Hollingsworth, M.L.; Richards, A.J.; Bateman, R.M. Taxonomic complexity, conservation and recurrent origins of self-pollination in Epipactis (Orchidaceae). In Current Taxonomic Research on the British and European Flora; Bailey, J.P., Ellis, R.G., Eds.; Botanical Society of the British Isles: London, UK, 2006; pp. 27–44. [Google Scholar]
- Efimov, P.G. Notes on Epipactis condensata, E. rechingeri and E. purpurata (Orchidaceae) in the Caucasus and Crimea. Willdenowia 2008, 38, 71–80. [Google Scholar] [CrossRef]
- Szlachetko, D.L. Flora Polski. Storczyki; Multico: Warsaw, Poland, 2009. [Google Scholar]
- Fateryga, A.V.; Kreutz, C.A.J. A new Epipactis species from the Crimea, South Ukraine (Orchidaceae). J. Eur. Orchid. 2012, 44, 199–206. [Google Scholar]
- Fateryga, A.V.; Kreutz, C.A.J.; Fateryga, V.V.; Efimov, P.G. Epipactis krymmontana (Orchidaceae), a new species endemic to the Crimean Mountains and notes on the related taxa in the Crimea and bordering Russian Caucasus. Phytotaxa 2014, 172, 22–30. [Google Scholar] [CrossRef]
- Chase, M.W.; Cameron, K.M.; Freudenstein, J.V.; Pridgeon, A.M.; Salazar, G.; Van den Berg, C.; Schuiteman, A. An updated classification of Orchidaceae. Bot. J. Linn. Soc. 2015, 177, 151–174. [Google Scholar] [CrossRef]
- Fateryga, A.V.; Fateryga, V.V. The genus Epipactis Zinn (Orchidaceae) in the flora of Russia. Turczaninowia 2018, 21, 19–34. [Google Scholar]
- Bateman, R.M. Implications of next-generation sequencing for the systematics and evolution of the terrestrial orchid genus Epipactis, with particular reference to the British Isles. Kew Bull. 2020, 75, 1–22. [Google Scholar] [CrossRef]
- Clark, M. Epipactis leptochila var. cordata. J. Hardy Orchid Soc. 2021, 18, 26. [Google Scholar]
- Squirrell, J.; Hollingsworth, P.M.; Bateman, R.M.; Tebbitt, M.C.; Hollingsworth, M.L. Taxonomic complexity and breeding system transitions: Conservation genetics of the Epipactis leptochila complex (Orchidaceae). Mol. Ecol. 2002, 11, 1957–1964. [Google Scholar] [CrossRef]
- Krajnc, A.U.; Ivanuš, A.; Luthar, Z.; Lipovšek, M. Morphological variability and taxonomic concepts of Broad-leaved Helleborine ingroup Epipactis helleborine (L.) Crantz. Folia Biol. Geol. 2020, 61, 97–125. [Google Scholar] [CrossRef]
- Ehlers, B.K.; Olesen, J.M.; Ågren, J. Floral morphology and reproductive success in the orchid Epipactis helleborine: Regional and local across-habitat variation. Plant Syst. Evol. 2002, 236, 19–32. [Google Scholar] [CrossRef]
- Foley, M.; Clarke, S. Orchids of the British Isles; Griffin: Cheltenham, UK, 2005. [Google Scholar]
- Delforge, P. Orchids of Europe, North Africa and the Middle East; A&C Black Publishers Ltd.: London, UK, 2006. [Google Scholar]
- Jakubska-Busse, A.; Gola, E.M. Morphological variability of Helleborines. I. Diagnostic significance of morphological features in Epipactis helleborine (L.) Crantz, Epipactis atrorubens (Hoffm.) Besser and their hybrid, Epipactis x schmalhausenii Richt. (Orchidaceae, Neottieae). Acta Soc. Bot. Pol. 2010, 79, 207–213. [Google Scholar] [CrossRef][Green Version]
- Jacquemyn, H.; van der Meer, S.; Brys, R. The impact of hybridization on long-term persistence of polyploid Dactylorhiza species. Am. J. Bot. 2016, 103, 1829–1837. [Google Scholar] [CrossRef]
- Jakubska-Busse, A.; Żołubak, E.; Łobas, Z. Epipactis ×subtilis (Orchidaceae), a new hybrid between E. albensis and E. purpurata. Ann. Bot. Fenn. 2017, 54, 139–144. [Google Scholar] [CrossRef]
- Ivanuš, A. Morphological and Molecular Analyses of Individual Genotypes from the Group of Epipactis helleborine s.l. in the Region of Goričko. Diploma Thesis, University of Maribor, Maribor, Slovenia, 2018. [Google Scholar]
- Jakubska-Busse, A.; Proćków, J.; Górniak, M.; Gola, E.M. Is Epipactis pseudopurpurata distinct from E. purpurata (Orchidaceae)? Evidence from morphology, anatomy, DNA and pollination biology. Bot. J. Linn. Soc. 2012, 170, 243–256. [Google Scholar] [CrossRef][Green Version]
- Brullo, C.; D’Emerico, S.; Pulvirenti, S. Karyological and taxonomical considerations on Epipactis cupaniana sp. nov. (Orchidaceae) from Sicily. Nord. J. Bot. 2013, 31, 577–589. [Google Scholar] [CrossRef]
- Sramkó, G.; Paun, O.; Brandrud, M.K.; Laczkó, L.; Molnár, A.; Bateman, R.M. Iterative allogamy–autogamy transitions drive actual and incipient speciation during the ongoing evolutionary radiation within the orchid genus Epipactis (Orchidaceae). Ann. Bot. 2019, 124, 481–497. [Google Scholar] [CrossRef]
- Sundermann, H. Europäische und Mediterrane Orchideen; Brücke Verlag Kurt Schmersow: Hildesheim, Germany, 1975. [Google Scholar]
- Richards, A.J. The influence of minor structural changes in the flower on breeding systems and speciation in Epipactis Zinn. (Orchidaceae). In Pollination and Evolution; Armstrong, J.A., Powell, J.M., Richards, A.J., Eds.; Royal Botanic Gardens: Sydney, Australia, 1982; pp. 47–53. [Google Scholar]
- Davies, P.; Davies, J.; Huxley, A. Wild orchids of Britain and Europe; The Hogarth Press: London, UK, 1983. [Google Scholar]
- Buttler, K.P. Field Guide to Orchids of Britain and Europe; The Crowd Press: Marlborough, UK, 1991. [Google Scholar]
- Delforge, P. Guide des Orchidées d’Europe d’Afrique du Nord et du Proche-Orient; Delachaux et Niestlé: Lausanne, Switzerland, 1994. [Google Scholar]
- Delforge, P. Orchids of Britain and Europe. Collins Photo Guide; Harper Collins Publishers: London, UK, 1995. [Google Scholar]
- Delforge, P. Orchidées d’Europe, d‘Afrique du Nord et du Proche-Orient, 4th ed.; Delachaux et Niestlé: Paris, France, 2016. [Google Scholar]
- Baumann, H.; Künkele, S.; Lorenz, R. Ulmer Naturführer Orchideen Europas mit Angrenzenden Gebieten; Ulmer Eugen Verlag: Stuttgart, Germany, 2006. [Google Scholar]
- Xinqi, C.; Zhongjian, L.; Guanghua, Z.; Kaiyong, L.; Zhanhe, J.; Yibo, L.; Xiaohua, J.; Cribb, P.; Wood, J.; Gale, S.; et al. Flora of China, 25: Orchidaceae; Missouri Botanical Garden Press: Saint Louis, MO, USA, 2009. [Google Scholar]
- Štěpánková, J.; Chrtek, J., Jr.; Kaplan, Z. Květena České republiky, 8th ed.; Academia: Praha, Czech Republic, 2011. [Google Scholar]
- Mossberg, B.; Pedersen, H.A. Orchids; HarperCollins Publishers: Glasgow, Scotland, 2017. [Google Scholar]
- Youssef, S.; Galalaey, A.; Mahmood, A.; Mahdi, H.; Véla, E. Wild Orchids of the Kurdistan Region Areas: A Scientific Window on the Unexpected Nature of the North-Western Zagros; Société Méditerrannéenne d’Orchidologie: La Motte-d’Aigues, France, 2019. [Google Scholar]
- Cole, S.; Waller, M. Britain’s Orchids: A Field Guide to the Orchids of Great Britain and Ireland; Princeton University Press: Woodstock, UK, 2020. [Google Scholar]
- Efimov, P.G. Orchids of Russia: Annotated checklist and geographic distribution. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 1–18. [Google Scholar] [CrossRef]
- Rouhan, G.; Gaudeul, M. Plant taxonomy: A historical perspective, current challenges, and perspective. In Molecular Plant Taxonomy. Methods in Molecular Biology; Besse, P., Ed.; Humana: New York, NY, USA, 2021; Volume 2222. [Google Scholar]
- Squirrell, J.; Hollingsworth, P.M.; Bateman, R.M.; Dickson, J.H.; Light, M.H.S.; MacConaill, M.; Tebbitt, M.C. Partitioning and diversity of nuclear and organelle markers in native and introduced populations of Epipactis helleborine (Orchidaceae). Am. J. Bot. 2001, 88, 1409–1418. [Google Scholar] [CrossRef]
- Bateman, R.M. How many orchid species are currently native to the British Isles. In Current Taxonomic Research on the British and European Flora; Bailey, J.P., Ellis, R.G., Eds.; Botanical Society of the British Isles: London, UK, 2006; pp. 89–110. [Google Scholar]
- Tranchida-Lombardo, V.; Cafasso, D.; Cristaudo, A.; Cozzolino, S. Phylogeographic patterns, genetic affinities and morphological differentiation between Epipactis helleborine and related lineages in a Mediterranean glacial refugium. Ann. Bot. 2011, 107, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Holub, J. Epipactis leptochila (Godf.) Godf., a Epipactis muelleri Godf.—Nové druhy československé flóry. Preslia 1970, 42, 330–349. [Google Scholar]
- Senghas, K.; Sundermann, H. Probleme der Orchideengattung Epipactis. Jahresberichte des Naturwissenschaftlischen Vereins in Wuppertal 1970, 23, 1–132. [Google Scholar]
- Procházka, F.; Velísek, V. Orchideje Naší Přírody; Academia Ved: Prague, Czech Republic, 1983. [Google Scholar]
- Reinhard, H.R.; Gölz, P.; Peter, R.; Wildermuth, H. Die Orchideen de Schweiz und Angrenzender Gebiete; Fotorotar AG: Egg, Germany, 1991. [Google Scholar]
- Tyteca, D.; Dufrêne, M. Biostatistical studies of western European allogamous populations of the Epipactis helleborine (L.) Crantz species group (Orchidaceae). Syst. Bot. 1994, 19, 424–442. [Google Scholar] [CrossRef]
- Potůček, O.; Čačko, Ľ. Všechno o Orchidejích; Slovart: Prague, Czech Republic, 1996. [Google Scholar]
- Szlachetko, D.L.; Skakuj, M. Storczyki Polski; Sorus: Poznań, Poland, 1996. [Google Scholar]
- Mereďa, P. Kl’úč na určovanie druhov rodu Epipactis Zinn publikovaných z územia Slovenska. Bulletin Slovenskej Botanickej Spoločnosti. 1999, 21, 131–142. [Google Scholar]
- Průša, D. Orchideje České Republiky; Computer Press: Brno, Czech Republic, 2005. [Google Scholar]
- Průša, D. Orchideje České Republiky; CPress: Brno, Czech Republic, 2019. [Google Scholar]
- Harrap, A.; Harrap, S. Orchids of Britain and Ireland. A Field and Site Guide; A&C Black Publishers Ltd.: London, UK, 2009. [Google Scholar]
- Batoušek, P.; Kežlínek, Z. Kruštíky České Republiky; Český svaz ochránců přírody ZO Hořepník: Prostějov, Czech Republic, 2012. [Google Scholar]
- Bateman, R.M. Circumscribing species in the European orchid flora: Multiple datasets interpreted in the context of speciation mechanisms. Berichte aus den Arbeitskreisen Heimische Orchideen Beiheft 2012, 29, 160–212. [Google Scholar]
- Ehlers, B.K.; Pedersen, H.Æ. Genetic variation in three species of Epipactis (Orchidaceae): Geographic scale and evolutionary inferences. Biol. J. Linn. Soc. 2000, 69, 411–430. [Google Scholar] [CrossRef]
- Brzosko, E.; Wróblewska, A.; Talalaj, I. Genetic variation and genotypic diversity in Epipactis helleborine populations from NE Poland. Plant Syst. Evol. 2004, 248, 57–69. [Google Scholar] [CrossRef]
- Bateman, R.M.; Hollingsworth, P.M.; Squirrell, J.; Hollingsworth, M.L. Tribe Neottieae. Phylogenetics. In Genera Orchidacearum 4. Epidendroideae 1; Pridgeon, A.M., Cribb, P.J., Chase, M.W., Rasmussen, F.N., Eds.; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Zhou, T.; Jin, X.H. Molecular systematics and the evolution of mycoheterotrophy of tribe Neottieae (Orchidaceae, Epidendroideae). PhytoKeys 2018, 94, 39–49. [Google Scholar] [CrossRef]
- Xing, X.; Gao, Y.; Zhao, Z.; Waud, M.; Duffy, K.J.; Selosse, M.A.; Jakalski, M.; Liu, N.; Jacquemyn, H.; Guo, S. Similarity in mycorrhizal communities associating with two widespread terrestrial orchids decays with distance. J. Biogeogr. 2020, 47, 421–433. [Google Scholar] [CrossRef]
- Lang, D. Britain’s Orchids, a Guide to the Identification and Ecology of the Wild Orchids of Britain and Ireland; WILDGuides Ltd.: Old Basing, UK, 2004. [Google Scholar]
- Wittig, R.; Wittig, M. Epipactis helleborine (L.) Crantz—The first (semi) ruderal orchid species of Central Europe. Feddes Repert. 2007, 118, 46–50. [Google Scholar] [CrossRef]
- Kühn, R.; Pedersen, H.Æ.; Cribb, P. Field Guide to the Orchids of Europe and the Mediterranean; Kew Publishing: Kew, UK, 2019. [Google Scholar]
- Kolanowska, M. Niche conservatism and the future potential range of Epipactis helleborine (Orchidaceae). PLoS ONE 2013, 8, e77352. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rewicz, A.; Rewers, M.; Jędrzejczyk, I.; Rewicz, T.; Kołodziejek, J.; Jakubska-Busse, A. Morphology and genome size of Epipactis helleborine (L.) Crantz (Orchidaceae) growing in anthropogenic and natural habitats. PeerJ 2018, 6, e5992. [Google Scholar] [CrossRef] [PubMed]
- Young, D.P. Bestimmung und Verbreitung der autogamen Epipactis-Arten. Jahresberichte Naturwissenschaftlichen Vereins Wuppertal 1970, 23, 43–52. [Google Scholar]
- Bayer, M. Die Gattung Epipactis Zinn in Baden-Wuirttemberg. Mitteilungsblättern Arbeitskreis Heimische Orchideen Baden-Württemberg 1980, 12, 219–268. [Google Scholar]
- Robatsch, K. Beitrage zur Bliitenbiologie und Autogamie der Gattung Epipactis. Jahresberichte Naturwissenschaftlichen Vereins Wuppertal 1983, 36, 25–32. [Google Scholar]
- Engel, R. Epipactis allogames et autogames. Monde Plantes 1986, 425–426, 12–18. [Google Scholar]
- Popovich, A.V.; Averyanova, E.A.; Shagarov, L.M. Orchids of the Black Sea coast of Krasnodarsky Krai (Russia): Current state, new records, conservation. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 46–68. [Google Scholar] [CrossRef]
- Kreutz, C.A.J. Kompendium der Europäischen Orchideen. Catalogue of European Orchids; Kreutz C.A.J.: Landgraaf, The Netherlands, 2004. [Google Scholar]
- Delforge, P. Nouvelles contributions taxonomiques et nomenclaturales aux Orchidées d’Europe. Natural. Belges 2015, 96, 14–21. [Google Scholar]
- POWO—Plants of the World Online. Facilitated by the Royal Botanic Gardens: Kew. 2019. Available online: http://www.plantsoftheworldonline.org/ (accessed on 9 August 2021).
- WCSP—World Checklist of Selected Plant Families. Facilitated by the Royal Botanic Gardens: Kew. 2021. Available online: http://wcsp.science.kew.org/ (accessed on 9 August 2021).
- WCVP—World Checklist of Vascular Plants, version 2.0. Facilitated by the Royal Botanic Gardens: Kew. 2021. Available online: http://wcvp.science.kew.org/ (accessed on 9 August 2021).
- WFO—World Flora Online. 2021. Available online: http://www.worldfloraonline.org/ (accessed on 9 August 2021).
- Linnaeus, C. Species Plantarum, 1st ed.; L. Salvius: Stockholm, Sweden, 1753. [Google Scholar]
- Govaerts, R.H.A. World Checklist of Monocotyledons Database in ACCESS: 1-71827; The Board of Trustees of the Royal Botanic Gardens: Kew, UK, 2003. [Google Scholar]
- Govaerts, R.H.A. World Checklist of Selected Plant Families published update; Facilitated by the Trustees of the Royal Botanic Gardens: Kew, UK, 2011. [Google Scholar]
- Govaerts, R.H.A. World Checklist of Vascular Plants (WCVP Database); The Board of Trustees of the Royal Botanic Gardens: Kew, UK, 2019. [Google Scholar]
- Gruppo Italiano per la Ricerca sulle Orchidee Spontanee (G.I.R.O.S.). Orchidee d’Italia—Guida alle Orchidee Spontanee; Il Castello: Milano, Italy, 2009; Available online: http://www.giros.it/ (accessed on 9 August 2021).
- Efimov, P.G.; Verkhozina, A.V. Epipactis helleborine var. tangutica (Orchidaceae), a new taxon for the flora of Russia and Middle Asia. Botanicheskii Zhurnal 2014, 99, 91–95. [Google Scholar]
- Klein, E. Revision der spanischen Epipactis-Taxa E. atrorubens (Hoffm.) Schult. subsp. parviflora A. et C. Nieschalk, „E. atrorubenti-microphylla“ und E. tremolsii C. Pau. Die Orchidee 1979, 30, 49–51. [Google Scholar]
- Greuter, W.; Raus, T. Med-Checklist Notulae, 13. Willdenowia 1986, 16, 103–116. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).