
Spatiotemporal Variation in Pollination Deficits in an Insect-Pollinated Dioecious Crop

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Contribution of Wind to Kiwifruit Pollination
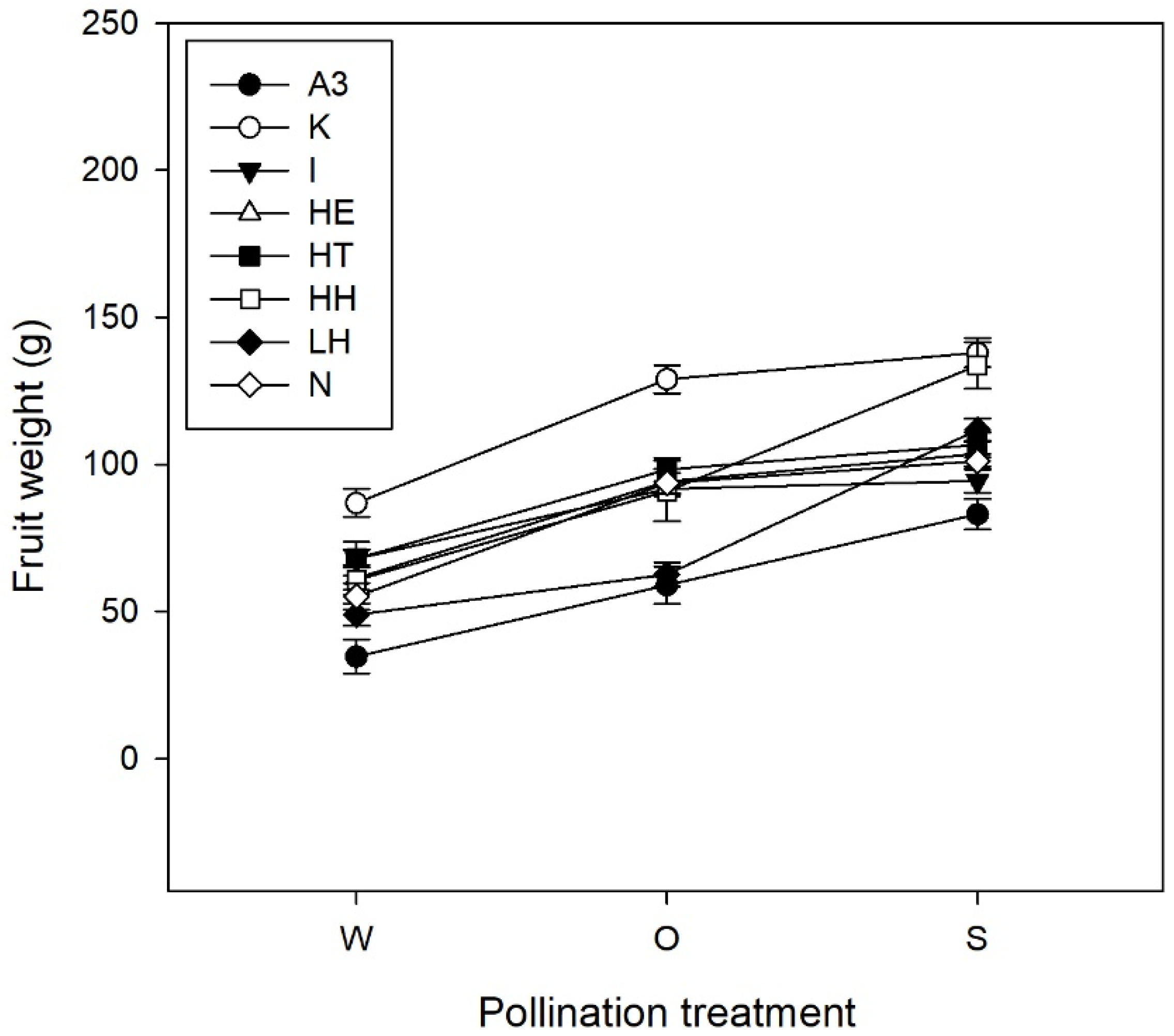
2.2. Pollination Deficits
2.3. Linking Pollination Deficits and Production with Environmental and Pollinator’s Related Variables
2.4. Linking Pollination Deficit with Production and Market Value
3. Discussion
3.1. Contribution of Wind to Kiwifruit Pollination
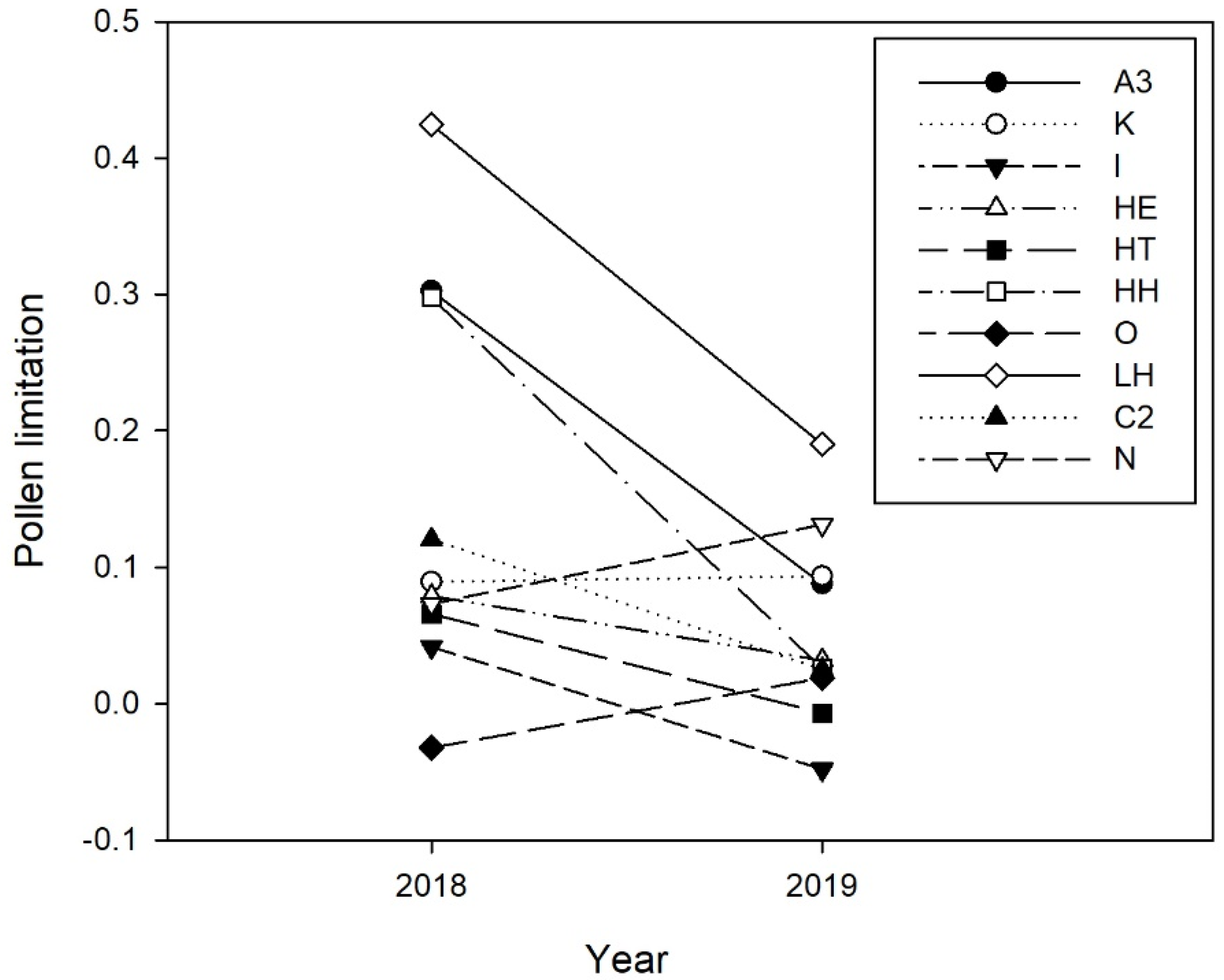
3.2. Spatio-Temporal Variation of Pollination Deficit
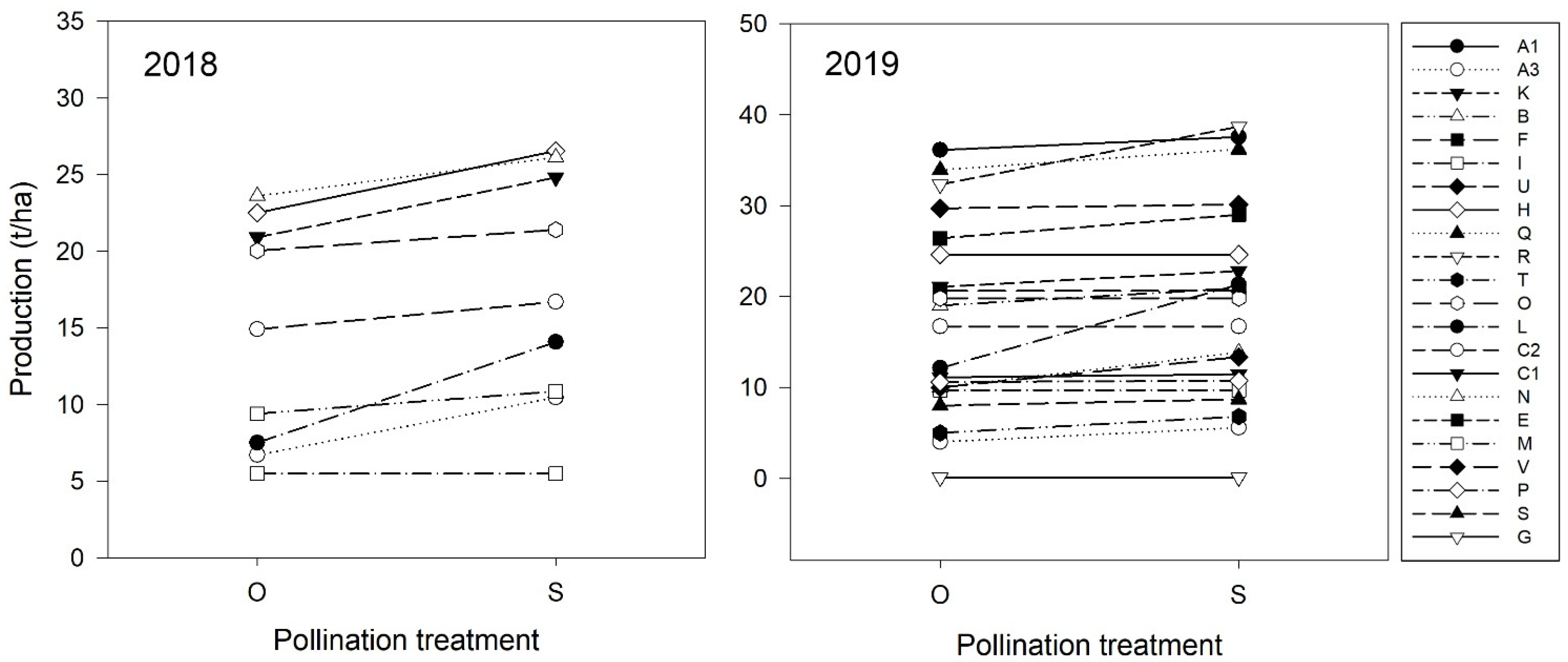
3.3. Pollination Services, Production and Market Value
4. Materials and Methods
4.1. Studied Orchards
4.2. Pollination Experiments
4.3. Fruit Processing
4.4. Pollination Deficit
4.5. Commercial Grading
4.6. Economic Analysis
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Aizen, M.A.; Garibaldi, L.A.; Cunningham, S.A.; Klein, A.M. How much does agriculture depend on pollinators? Lessons from long-term trends in crop production. Ann. Bot. 2009, 103, 1579–1588. [Google Scholar] [CrossRef] [PubMed]
- Knight, T.M.; Steets, J.A.; Vamosi, J.C.; Mazer, S.J.; Burd, M.; Campbell, D.R.; Dudash, M.R.; Johnston, M.O.; Mitchell, R.J.; Ashman, T.-L. Pollen Limitation of Plant Reproduction: Pattern and Process. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 467–497. [Google Scholar] [CrossRef] [Green Version]
- Vaissière, B.; Freitas, B.; Gemmill-Herren, B. Protocol to Detect and Assess Pollination Deficits in Crops: A Handbook for Its Use; Food and Agiculture Organization of the United Nations: Rome, Italy, 2011. [Google Scholar]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Freitas, B.M.; Vaissiére, B.E.; Saraiva, A.M.; Carvalheiro, L.G.; Garibaldi, L.A.; Ngo, H. Identifying and assessing pollina-tion deficits in crops. In Pollination Services to Agriculture: Sustaining and Enhancing a Key Ecosystem Service; Taylor & Francis: Oxfordshire, UK, 2016; pp. 17–32. ISBN 9781317445678. [Google Scholar]
- Lebuhn, G.; Droege, S.; Connor, E.; Gemmill-Herren, B.; Azzu, N. Protocol to Detect and Monitor Pollinator Communities–Guidance for Practitioners; Food and Agiculture Organization of the United Nations: Rome, Italy, 2016; ISBN 9789251089781. [Google Scholar]
- Garratt, M.P.D.; Evans, D.J.; Moss, R.L.; Dodson, E.D.; Jenner, C.; Potts, J.C. Pollination deficits in UK apple orchards. J. Pollinat. Ecol. 2014, 12, 9–14. [Google Scholar]
- Knapp, J.L.; Osborne, J.L. Courgette Production: Pollination Demand, Supply, and Value. J. Econ. Èntomol. 2017, 110, 1973–1979. [Google Scholar] [CrossRef] [Green Version]
- Webber, S.M.; Garratt, M.P.D.; Lukac, M.; Bailey, A.P.; Huxley, T.; Potts, S.G. Quantifying crop pollinator-dependence and pollination deficits: The effects of experimental scale on yield and quality assessments. Agric. Ecosyst. Environ. 2020, 304, 107106. [Google Scholar] [CrossRef]
- Pomeroy, N.; Fisher, R.M. Pollination of kiwifruit (Actinidia deliciosa) by bumble bees (Bombus terrestris): Effects of bee density and patterns of flower visitation. N. Z. Èntomol. 2002, 25, 41–49. [Google Scholar] [CrossRef]
- Testolin, R.; Vizzotio, G.; Costa, G. Kiwifruit pollination by wind and insects in Italy. N. Z. J. Crop. Hortic. Sci. 1991, 19, 381–384. [Google Scholar] [CrossRef]
- Costa, G.; Testolin, R.; Vizzotto, G. Kiwifruit pollination: An unbiased estimate of wind and bee contribution. N. Z. J. Crop. Hortic. Sci. 1993, 21, 189–195. [Google Scholar] [CrossRef]
- Ferguson, A.R. Kiwifruit: The Wild and the Cultivated Plants. Adv. Food Nutr. Res. 2013, 68, 15–32. [Google Scholar] [CrossRef]
- Ferguson, A.R. Kiwifruit: A Botanical Review. In Horticultural Reviews; Janick, J., Ed.; AVI Publishing Company, INC: Westport, CT, USA, 1984; pp. 1–64. [Google Scholar]
- Tacconi, G.; Michelotti, V.; Cacioppo, O.; Vittone, G. Kiwifruit pollination: The interaction between pollen quality, pollination systems and flowering stage. J. Berry Res. 2016, 6, 417–426. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, M.; Veloso, A.; Teixeira, M.; Veloso, F.; Antunes, D. Avaliação do efeito da polinização complementar em actinídea deliciosa CV. “Hayward” na Região de Entre—Douro e Minho. In Proceedings of the I Simpósium Nacional de Fruticultura, Alcobaça, Portugal, 12–13 October 2006; Ramos, A., Martins, J.M.S., Eds.; Associação Portuguesa de Horticultura: Lisbon, Portugal, 2009; pp. 97–101. [Google Scholar]
- Miñarro, M.; Twizell, K.W. Pollination services provided by wild insects to kiwifruit (Actinidia deliciosa). Apidologie 2015, 46, 276–285. [Google Scholar] [CrossRef] [Green Version]
- González, M.V.; Coque, M.; Herrero, M. Influence of pollination systems on fruit set and fruit quality in kiwifruit (Actinidia deliciosa). Ann. Appl. Biol. 1998, 132, 349–355. [Google Scholar] [CrossRef]
- Tacconi, G.; Michelotti, V. Artificial Pollination in Kiwifruit and Olive Trees. In Pollination in Plants; Segneanu, A.E., Orbeci, C., Lazau, C., Sfirloaga, P., Vlazan, P., Bandas, C., Grozescu, I., Eds.; InTech: London, UK, 2018; p. 29. ISBN 978-1-78923-237-0. [Google Scholar]
- Vicens, N.; Bosch, J. Weather-Dependent Pollinator Activity in an Apple Orchard, with Special Reference to Osmia cornuta and Apis mellifera (Hymenoptera: Megachilidae and Apidae). Environ. Èntomol. 2000, 29, 413–420. [Google Scholar] [CrossRef]
- Donati, I.; Cellini, A.; Buriani, G.; Mauri, S.; Kay, C.; Tacconi, G.; Spinelli, F. Pathways of flower infection and pollen-mediated dispersion of Pseudomonas syringae pv. actinidiae, the causal agent of kiwifruit bacterial canker. Hortic. Res. 2018, 5, 56. [Google Scholar] [CrossRef] [Green Version]
- Testolin, R.; Ferguson, A.R. Kiwifruit (Actinidia spp.) Production and Marketing in Italy. N. Z. J. Crop Hortic. Sci. 2009, 37, 1–32. [Google Scholar] [CrossRef]
- Goodwin, M. Pollination of Crops in Australia and New Zealand; Rural Industries Research and Development Corporation: Wagga Wagga, Australia, 2012; ISBN 9781742544021. [Google Scholar]
- NZKGI. Kiwifruit Book; NZKGI: Mount Maunganui, New Zealand, 2016. [Google Scholar]
- Antunes, M.D.; Terra, C. Da Produção à Comercialização; Universidade do Algarve: Algarve, Portugal, 2008. [Google Scholar]
- Gaspar, H. Impact of Insect Diversity and Abundance in Kiwi Orchard’s Production; University of Coimbra: Coimbra, Portugal, 2020. [Google Scholar]
- Martins, K.T.; Gonzalez, A.; Lechowicz, M.J. Pollination services are mediated by bee functional diversity and landscape context. Agric. Ecosyst. Environ. 2015, 200, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Fijen, T.P.M.; Scheper, J.A.; Boekelo, B.; Raemakers, I.; Kleijn, D. Effects of landscape complexity on pollinators are moderated by pollinators’ association with mass-flowering crops. Proc. R. Soc. B Boil. Sci. 2019, 286, 20190387. [Google Scholar] [CrossRef] [Green Version]
- Craig, J.L.; Stewart, A.M.; Pomeroy, N.; Heath, A.C.G.; Goodwin, R.M. A review of kiwifruit pollination: Where to next? N. Z. J. Exp. Agric. 1988, 16, 385–399. [Google Scholar] [CrossRef]
- Corbet, S.A.; Chapman, H.; Saville, N. Vibratory Pollen Collection and Flower Form: Bumble-Bees on Actinidia, Symphytum, Borago and Polygonatum. Funct. Ecol. 1988, 2, 147. [Google Scholar] [CrossRef]
- Goodwin, R.M.; Congdon, N.M. Recognition and attractiveness of staminate and pistillate kiwifruit flowers (Actinidia deliciosa var. deliciosa) by honey bees (Apis mellifera L.). N. Z. J. Crop. Hortic. Sci. 2017, 46, 72–80. [Google Scholar] [CrossRef]
- McPherson, H.G.; Hall, A.J.; Stanley, C.J. Seasonal and regional variation in budbreak and flowering of kiwifruit vines in New Zealand. N. Z. J. Crop. Hortic. Sci. 1994, 22, 263–276. [Google Scholar] [CrossRef]
- Tuell, J.K.; Isaacs, R. Weather during bloom affects pollination and yield of highbush blueberry. J. Econ. Èntomol. 2010, 103, 557–562. [Google Scholar] [CrossRef]
- Blitzer, E.J.; Gibbs, J.; Park, M.G.; Danforth, B. Pollination services for apple are dependent on diverse wild bee communities. Agric. Ecosyst. Environ. 2016, 221, 1–7. [Google Scholar] [CrossRef] [Green Version]
- MacInnis, G.; Forrest, J.R.K. Pollination by wild bees yields larger strawberries than pollination by honey bees. J. Appl. Ecol. 2019, 56, 824–832. [Google Scholar] [CrossRef]
- Connelly, H.; Poveda, K.; Loeb, G. Landscape simplification decreases wild bee pollination services to strawberry. Agric. Ecosyst. Environ. 2015, 211, 51–56. [Google Scholar] [CrossRef]
- Nicholson, C.C.; Koh, I.; Richardson, L.L.; Beauchemin, A.; Ricketts, T.H. Farm and landscape factors interact to affect the supply of pollination services. Agric. Ecosyst. Environ. 2017, 250, 113–122. [Google Scholar] [CrossRef]
- Bartual, A.M.; Sutter, L.; Bocci, G.; Moonen, A.-C.; Cresswell, J.; Entling, M.; Giffard, B.; Jacot, K.; Jeanneret, P.; Holland, J.; et al. The potential of different semi-natural habitats to sustain pollinators and natural enemies in European agricultural landscapes. Agric. Ecosyst. Environ. 2019, 279, 43–52. [Google Scholar] [CrossRef]
- Eeraerts, M.; Smagghe, G.; Meeus, I. Pollinator diversity, floral resources and semi-natural habitat, instead of honey bees and intensive agriculture, enhance pollination service to sweet cherry. Agric. Ecosyst. Environ. 2019, 284, 106586. [Google Scholar] [CrossRef]
- Lázaro, A.; Alomar, D. Landscape heterogeneity increases the spatial stability of pollination services to almond trees through the stability of pollinator visits. Agric. Ecosyst. Environ. 2019, 279, 149–155. [Google Scholar] [CrossRef]
- Bartholomée, O.; Aullo, A.; Becquet, J.; Vannier, C.; Lavorel, S. Pollinator presence in orchards depends on landscape-scale habitats more than in-field flower resources. Agric. Ecosyst. Environ. 2020, 293, 106806. [Google Scholar] [CrossRef]
- Evans, L.J.; Cutting, B.T.; Jochym, M.; Janke, M.A.; Felman, C.; Cross, S.; Jacob, M.; Goodwin, M. Netted crop covers reduce honeybee foraging activity and colony strength in a mass flowering crop. Ecol. Evol. 2019, 9, 5708–5719. [Google Scholar] [CrossRef]
- Antunes, M.; Oliveira, M.; Teixeira, M.; Veloso, A.; Veloso, F.; Panagopoulos, T. Evaluation of the effect of complementary pollination on Actinidia deliciosa ’hayward’ in northwest portugal. Acta Hortic. 2007, 753, 347–352. [Google Scholar] [CrossRef]
- Goodwin, R.; McBrydie, H.; Taylor, M. Wind and honey bee pollination of kiwifruit (Actinidia chinensis‘HORT16A’). N. Z. J. Bot. 2013, 51, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Castle, D.; Grass, I.; Westphal, C. Fruit quantity and quality of strawberries benefit from enhanced pollinator abundance at hedgerows in agricultural landscapes. Agric. Ecosyst. Environ. 2019, 275, 14–22. [Google Scholar] [CrossRef]
- Karamaouna, F.; Kati, V.; Volakakis, N.; Varikou, K.; Garantonakis, N.; Economou, L.; Birouraki, A.; Markellou, E.; Liberopoulou, S.; Edwards, M. Ground cover management with mixtures of flowering plants to enhance insect pollinators and natural enemies of pests in olive groves. Agric. Ecosyst. Environ. 2019, 274, 76–89. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sanchez-Diaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Osaki, M.; Shinano, T.; Tadano, T. Redistribution of carbon and nitrogen compounds from the shoot to the harvesting organs during maturation in field crops. Soil Sci. Plant Nutr. 1991, 37, 117–128. [Google Scholar] [CrossRef]
- Larson, B.M.H.; Barrett, S.C.H. A comparative analysis of pollen limitation in flowering plants. Biol. J. Linn. Soc. Linn. Soc. 2000, 69, 503–520. [Google Scholar] [CrossRef]
- Lawes, G.S.; Woolley, D.J.; Lai, R. Seeds and Other Factors Affecting Fruit Size in Kiwifruit. Acta Hortic. 1990, 257–264. [Google Scholar] [CrossRef]
- Vasilakakis, M.; Papadopoulos, K.; Papageorgiou, E. Factors affecting the fruit size of “hayward” kiwifruit. Acta Hortic. 1997, 419–424. [Google Scholar] [CrossRef]
- McPherson, H.G.; Richardson, A.C.; Snelgar, W.P.; Patterson, K.J.; Currie, M.B. Flower quality and fruit size in kiwifruit (Actinidia deliciosa). N. Z. J. Crop. Hortic. Sci. 2001, 29, 93–101. [Google Scholar] [CrossRef] [Green Version]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Statistics for Biology and Health; Springer: New York, NY, USA, 2009; ISBN 978-0-387-87457-9. [Google Scholar]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Bates, D.; Mächler, M. Package ‘lme4’: Linear mixed-effects models using eigen and S4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Means ± SE | X2 or F | p Values | ||
|---|---|---|---|---|---|
| Wind | Open | Supplement | |||
| Fruit set (%) | 66.52 ± 3.18 (b) | 73.20 ± 2.81 (b) | 85.60 ± 2.22 (a) | X22 = 24.72 | <0.001 |
| Weight (g) | 61.82 ± 2.02 (c) | 89.76 ± 2.01 (b) | 107.17 ± 1.87 (a) | F2, 527.6 = 164.51 | <0.001 |
| Dry matter (%) | 14.75 ± 0.28 (b) | 15.14 ± 0.21 (a, b) | 15.66 ± 0.21 (a) | F2, 214.0 = 164.51 | <0.001 |
| Flesh firmness (N) | 40.59 ± 2.08 (b) | 42.74 ± 1.85 (a) | 43.96 ± 1.75 (a) | F2, 215.03 = 6.12 | 0.003 |
| °Brix (%) | 6.03 ± 0.23 | 6.12 ± 0.20 | 6.21 ± 0.20 | F2, 215.0 = 0.246 | 0.783 |
| Acidity (%) | 1.46 ± 0.03 (b) | 1.55 ± 0.03 (a) | 1.50 ± 0.03 (a, b) | F2, 208.25 = 3.175 | 0.044 |
| S:L | 0.88 ± 0.00 (a) | 0.87 ± 0.00 (a, b) | 0.86 ± 0.00 (b) | F2, 515.9 = 3.08 | 0.047 |
| TAA (µmol eq. galic acid/g) | 3.08 ± 0.12 | 3.07 ± 0.10 | 2.98 ± 0.10 | F2, 211.0 = 0.354 | 0.702 |
| TSS (mg/g) | 4.81 ± 0.18 (b) | 5.33 ± 0.18 (a) | 5.31 ± 0.19 (a, b) | F2, 201.2 = 3.61 | 0.029 |
| Starch (mg/g) | 2.89 ± 0.06 | 2.93 ± 0.04 | 2.90 ± 0.05 | F2, 204.3 = 0.118 | 0.889 |
| Factors | Year | Year: Pollination Treatment | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Variables | 2018 | 2019 | X2 or F | p Values | 2018 | 2019 | X2 or F | p Values | ||
| Open | Supplement | Open | Supplement | |||||||
| Fruit set (%) | 78.68 ± 1.57 | 85.94 ± 0.88 | X21 = 1.51 | 0.219 | 73.24 ± 2.40 | 84.12 ± 1.99 | 82.99 ± 1.34 | 88.92 ± 1.13 | X22 = 26.27 | <0.001 |
| Weight (g) | 97.95 ± 1.22 | 103.65 ± 0.75 | F1, 20.26 = 0.256 | 0.619 | 91.89 ± 1.61 | 104.56 ± 1.53 | 100.57 ± 1.08 | 106.67 ± 1.02 | F2, 1865 =44.30 | <0.001 |
| Dry matter (%) | 15.56 ± 0.14 | 14.03 ± 0.07 | F1, 20.0 = 0.063 | <0.001 | 15.45 ± 0.19 | 15.88 ± 0.19 | 14.00 ± 0.10 | 14.07 ± 0.10 | F2, 1140 = 2.72 | 0.066 |
| Flesh firmness (N) | 47.24 ± 0.90 | 70.38 ± 0.35 | F1, 7.9 =65.65 | <0.001 | 46.89 ± 1.30 | 47.57 ± 1.24 | 70.14 ± 0.78 | 70.62 ± 1.24 | F2, 1519 = 1.26 | 0.284 |
| °Brix (%) | 6.19 ± 0.12 | 6.56± 0.05 | F1, 19.0 = 1.00 | 0.331 | 6.34 ± 0.16 | 6.26 ± 0.16 | 6.42 ± 0.07 | 6.57 ± 0.07 | F2, 1479 = 0.21 | 0.814 |
| Acidity (%) | 1.52 ± 0.02 | 1.54 ± 0.01 | F1, 12.4 = 0.13 | 0.728 | 1.54 ± 0.03 | 1.51 ± 0.02 | 1.54 ± 0.01 | 1.55 ±0.01 | F2, 1511= 0.69 | 0.501 |
| S:L | 0.88 ± 0.00 | 0.90 ± 0.00 | F1, 10.6 = 11.52 | 0.006 | 0.88 ± 0.00 | 0.90 ± 0.00 | 0.88 ± 0.00 | 0.90 ± 0.00 | F2, 1789= 1.29 | 0.274 |
| Pollination deficit | 0.11 ± 0.01 | 0.06 ± 0.01 | F1, 8.4 = 5.47 | 0.046 | - | - | - | - | - | - |
| Large fruits (%) | 50.98 ± 2.11 | 58.35 ± 1.34 | X21 = 0.35 | 0.554 | 42.86 ± 2.89 | 59.93 ± 3.00 | 54.45 ± 1.92 | 62.19 ± 1.85 | X22 = 29.33 | <0.001 |
| Medium fruits (%) | 24.06 ± 1.81 | 20.53 ± 1.10 | X21 = 1.17 | 0.282 | 24.49 ± 2.51 | 23.60 ± 2.60 | 20.18 ± 1.55 | 20.88 ± 1.55 | X22 = 0.21 | 0.901 |
| Small fruits (%) | 24.96 ± 1.83 | 21.12 ± 1.11 | X21 = 0.67 | 0.412 | 32.65 ± 2.74 | 16.48 ± 2.27 | 25.37 ± 1.68 | 16.93 ± 1.43 | X22 = 38.40 | <0.001 |
| Production (t/ha) | 15.97 ± 1.76 | 18.07 ± 1.63 | F1, 7.3 = 0.07 | 0.801 | 14.56 ± 2.46 | 17.38 ± 2.57 | 17.18 ± 2.27 | 18.97 ± 2.38 | F1, 28 = 11.71 | <0.001 |
| Monetary gain (€/ha) | 14,842.25± 1820.73 | 18,479.98± 2105.59 | F1, 7.3 = 0.01 | 0.948 | 12,385.04 ± 2912.32 | 17,299.47 ± 2664.70 | 16,935.04 ± 2340.69 | 20,024.43 ± 3075.26 | F1, 28 = 7.35 | 0.003 |
| Variables | Pollination Deficit | Production | ||||
|---|---|---|---|---|---|---|
| Estimate | t Value | p | Estimate | t Value | p | |
| Pollinator richness | 0.0007 | 0.267 | 0.792 | −0.603 | −2.600 | 0.021 |
| Honeybee abundance | −0.0004 | −2.286 | 0.034 | 0.039 | 2.399 | 0.031 |
| Bombus spp. abundance | −0.0003 | −0.822 | 0.421 | 0.077 | 2.206 | 0.045 |
| Precipitation | 0.0005 | 0.502 | 0.622 | 0.106 | 1.274 | 0.224 |
| Temperature | 0.0186 | 0.698 | 0.494 | −2.086 | −0.726 | 0.480 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castro, H.; Siopa, C.; Casais, V.; Castro, M.; Loureiro, J.; Gaspar, H.; Dias, M.C.; Castro, S. Spatiotemporal Variation in Pollination Deficits in an Insect-Pollinated Dioecious Crop. Plants 2021, 10, 1273. https://doi.org/10.3390/plants10071273
Castro H, Siopa C, Casais V, Castro M, Loureiro J, Gaspar H, Dias MC, Castro S. Spatiotemporal Variation in Pollination Deficits in an Insect-Pollinated Dioecious Crop. Plants. 2021; 10(7):1273. https://doi.org/10.3390/plants10071273
Chicago/Turabian StyleCastro, Helena, Catarina Siopa, Vinícius Casais, Mariana Castro, João Loureiro, Hugo Gaspar, Maria Celeste Dias, and Sílvia Castro. 2021. "Spatiotemporal Variation in Pollination Deficits in an Insect-Pollinated Dioecious Crop" Plants 10, no. 7: 1273. https://doi.org/10.3390/plants10071273
APA StyleCastro, H., Siopa, C., Casais, V., Castro, M., Loureiro, J., Gaspar, H., Dias, M. C., & Castro, S. (2021). Spatiotemporal Variation in Pollination Deficits in an Insect-Pollinated Dioecious Crop. Plants, 10(7), 1273. https://doi.org/10.3390/plants10071273