
On the Reproductive Potential in Primula veris L. (Primulaceae): Embryological Features, Pollen and Seed Viability, Genetic Diversity



Abstract
1. Introduction
2. Results and Discussion
2.1. Embryological Features
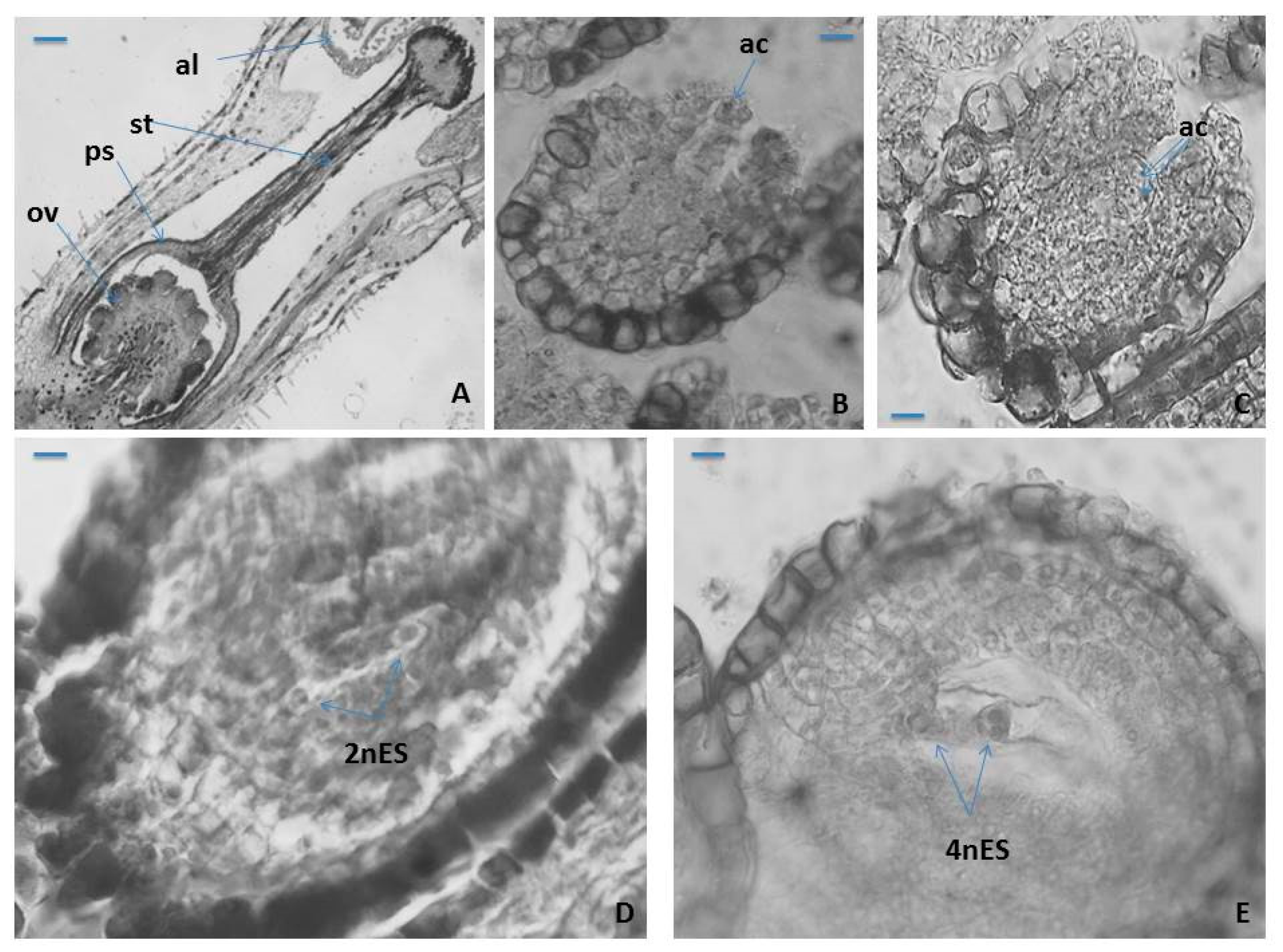
2.1.1. Anther, Microsporogenesis and Male Gametophyte Development
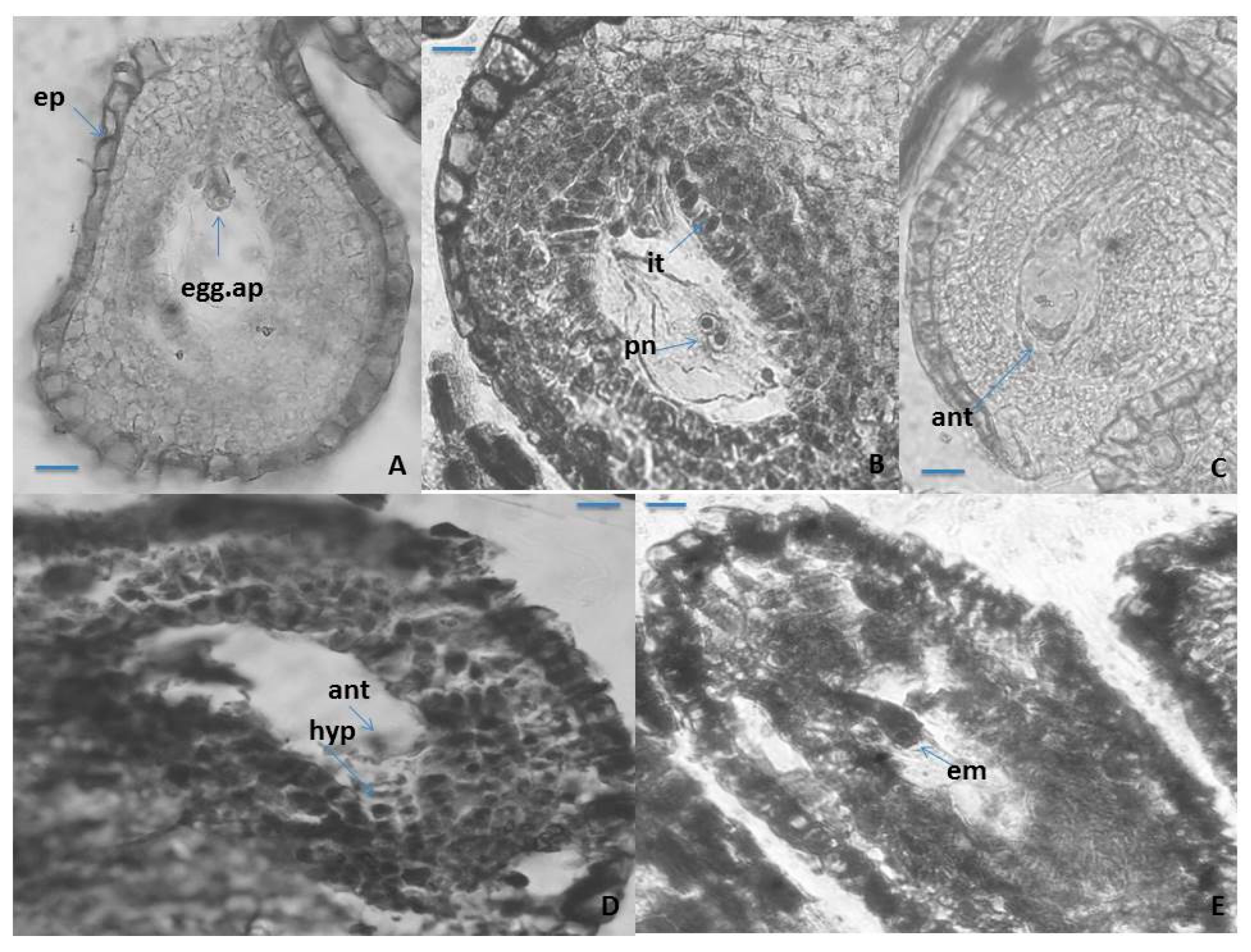
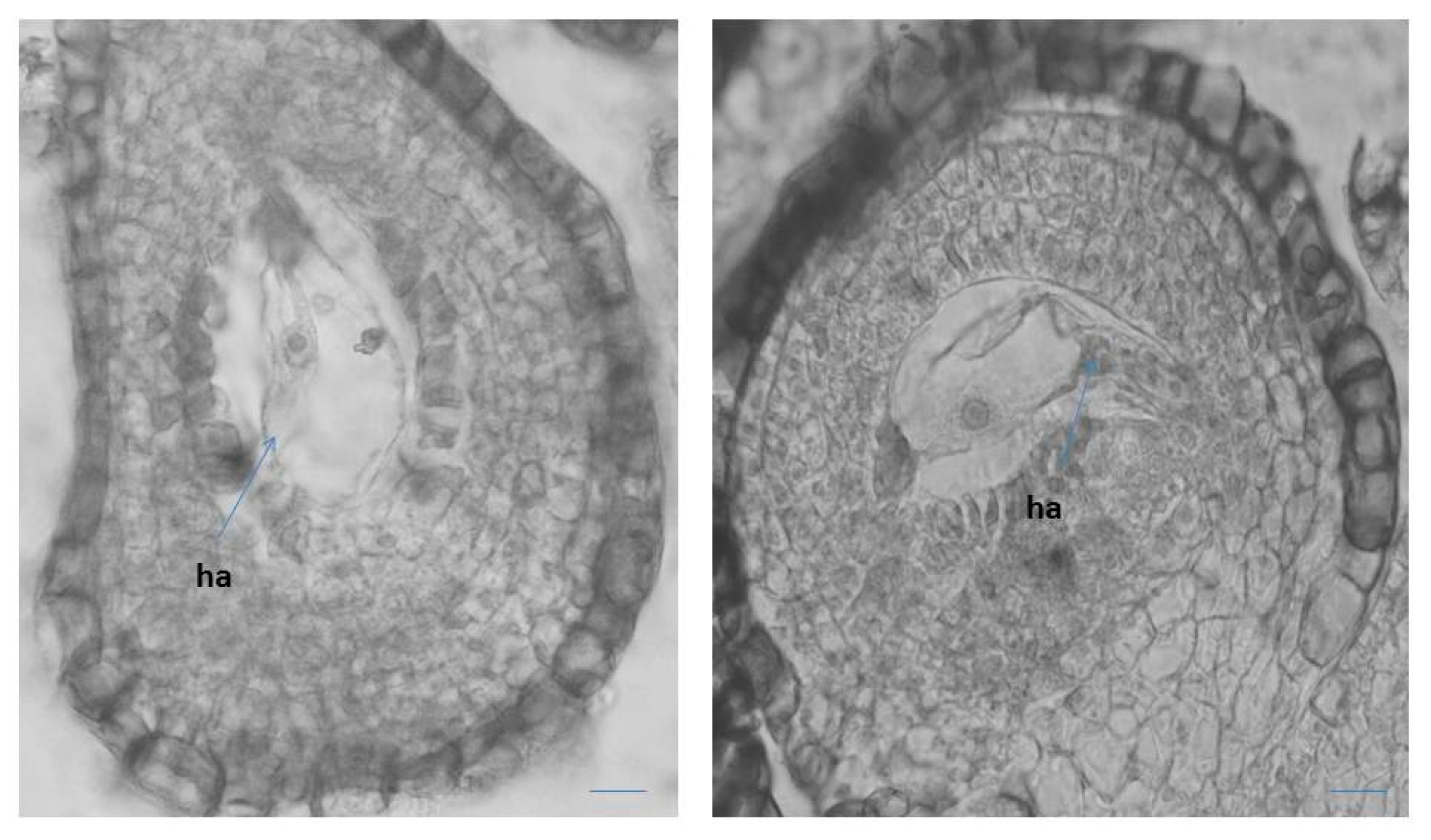
2.1.2. Ovule, Megasporogenesis and Female Gametophyte Development
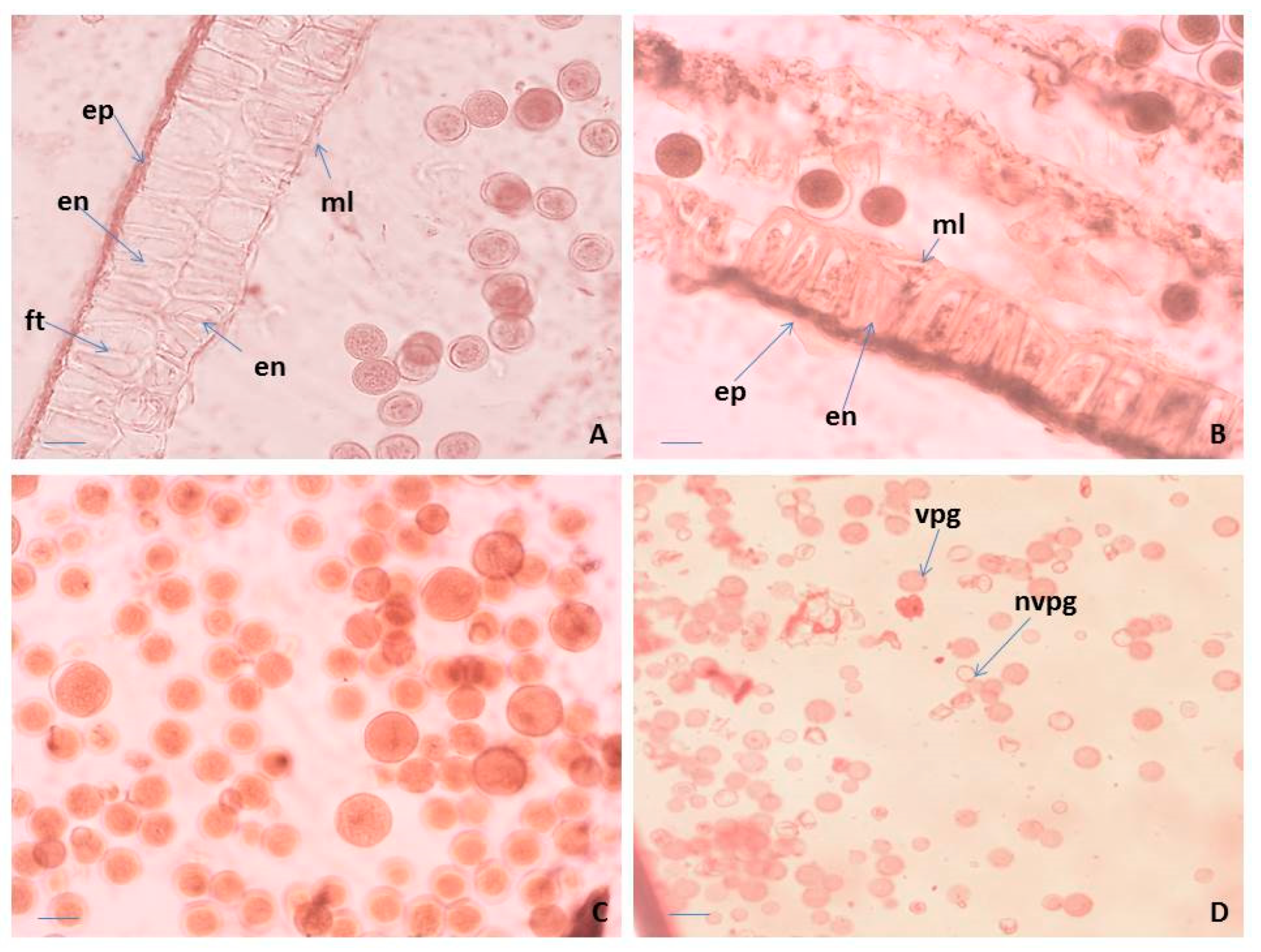
2.2. Pollen Viability
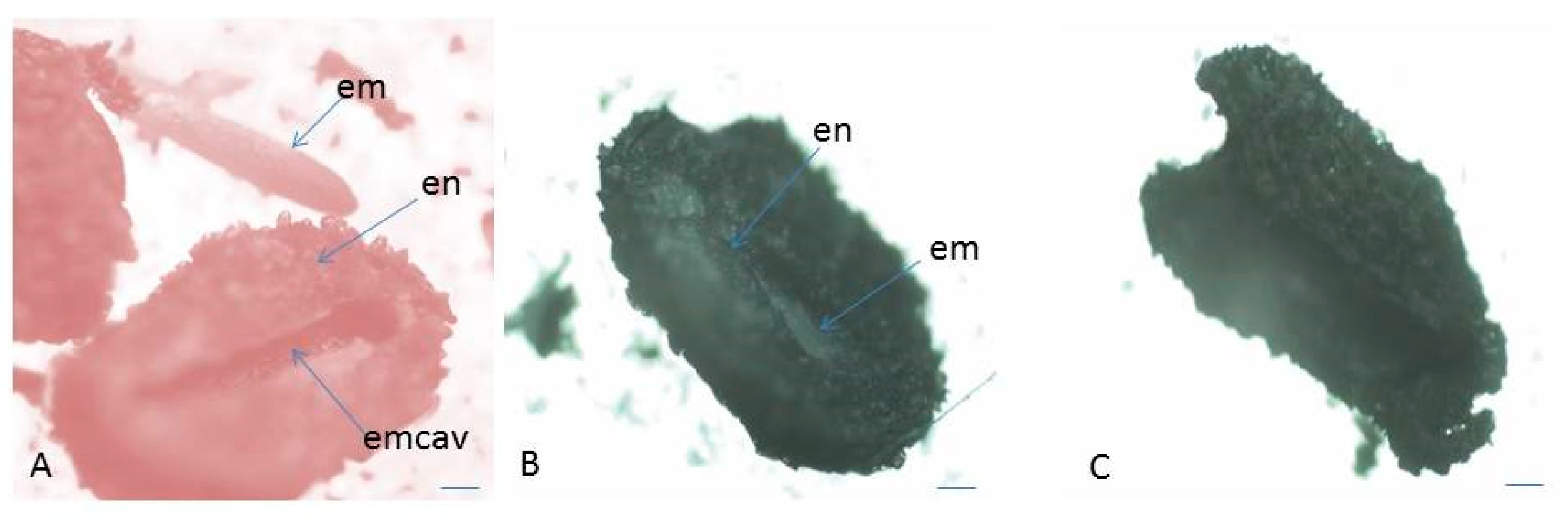
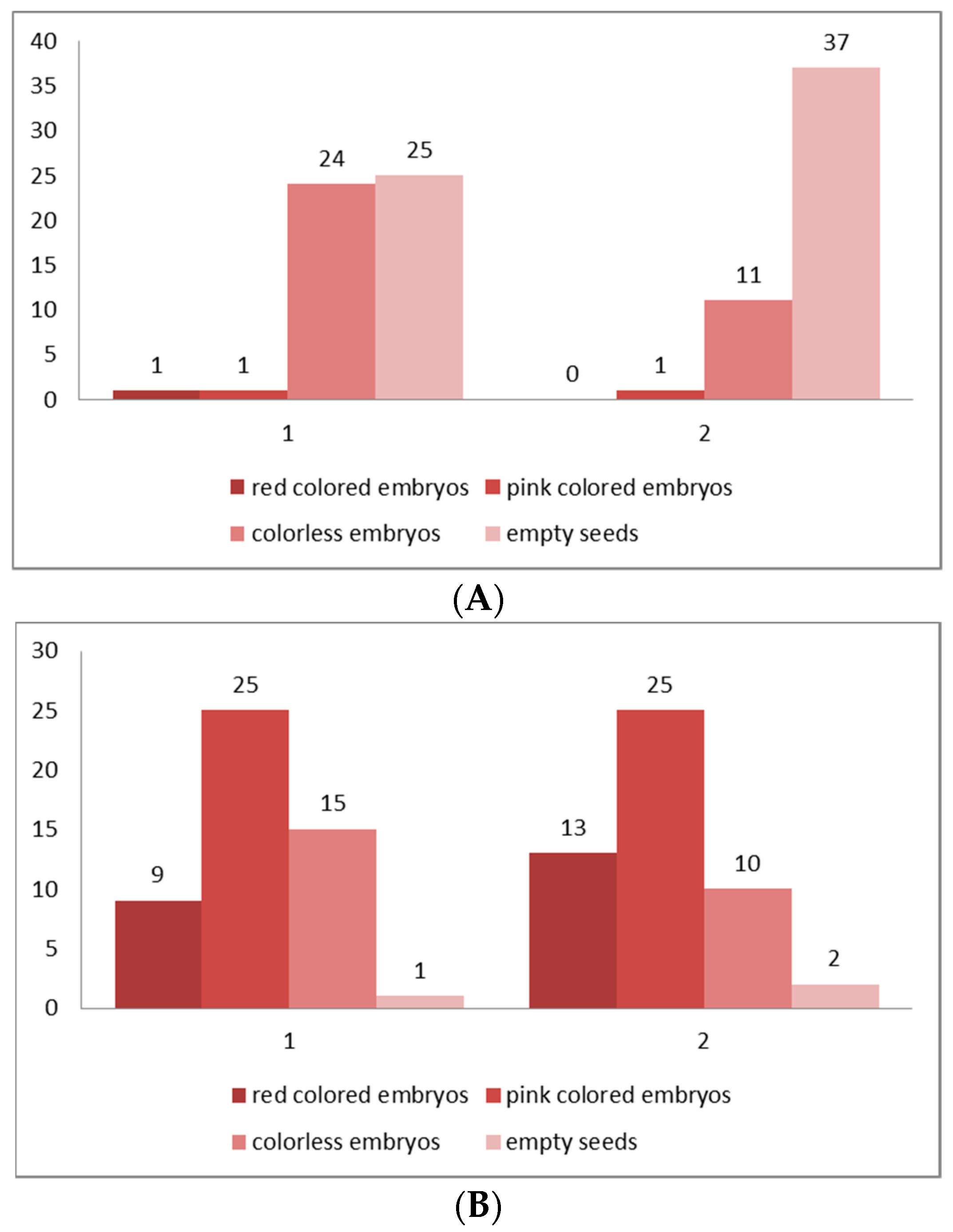
2.3. Seed Viability
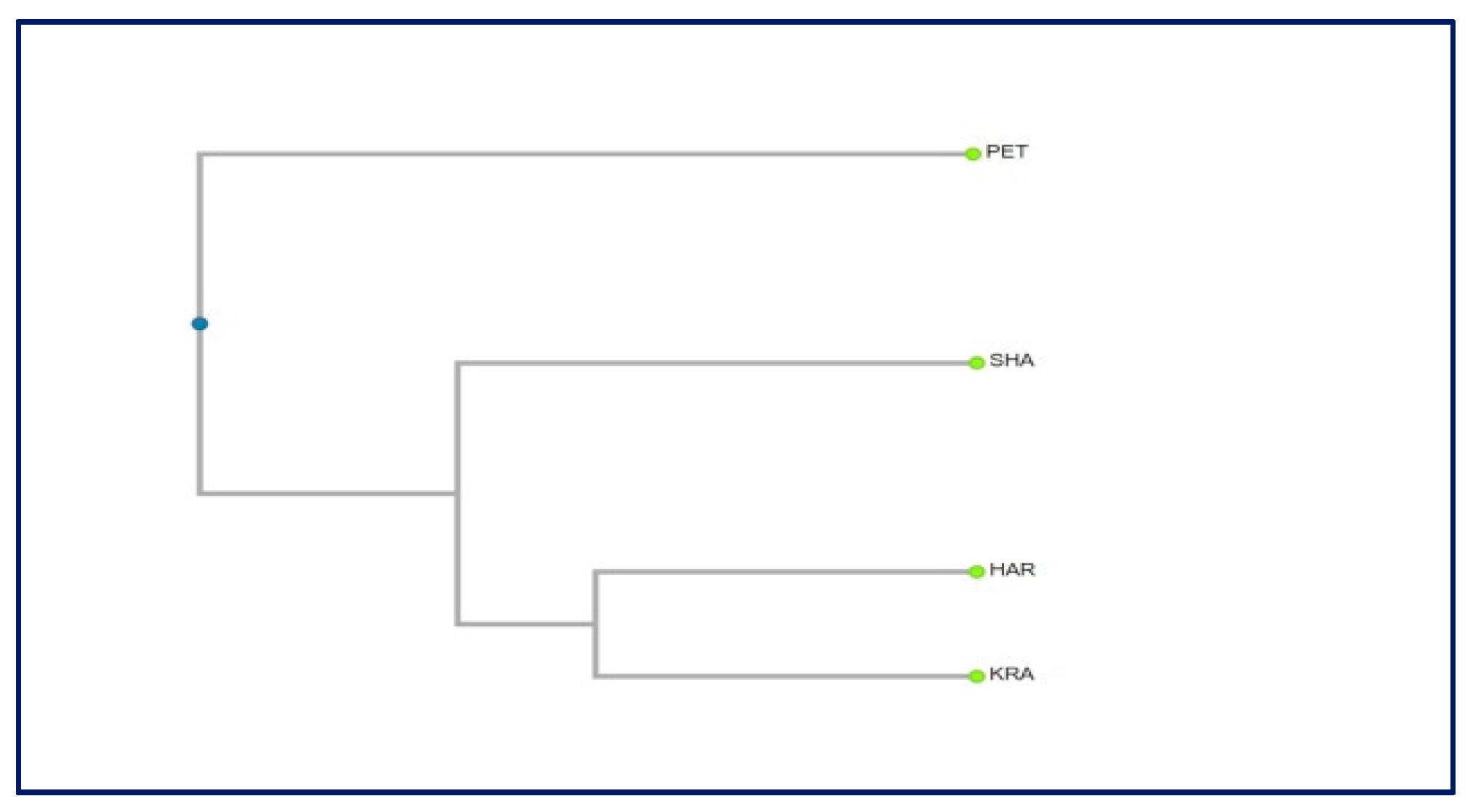
2.4. Genetic Diversity
3. Materials and Methods
3.1. Study of Reproductive Capacity
3.1.1. Embryological Study
3.1.2. Pollen Viability
3.1.3. Seed Viability
3.2. Study of Genetic Diversity in the Target Species
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Richards, J. Primula L., 2nd ed.; Timber Press: Portland, OR, USA, 2003; 386p. [Google Scholar]
- Zhang, L.B.; Kadereit, J.W. Classification of Primula sect. Auricula (Primulaceae) based on two molecular data sets (ITS, AFLPs), morphology and geographical distribution. Bot. J. Linn. Soc. 2004, 146, 1–26. [Google Scholar]
- Darwin, C.R. The Different Forms of Flowers on Plants of the Same Species; John Murray: London, UK, 1877; p. 351. [Google Scholar]
- Barrett, S.C.H.; Jesson, L.K.; Baker, A.M. The evolution and function of stylar polymorphisms in flowering plants. Ann. Bot. 2000, 85, 253–265. [Google Scholar] [CrossRef]
- Brys, R.; Jacquemyn, H. The impact of individual inaccuracy of reciprocal herkogamy on legitimate pollen deposition and seed set in a distylous self-incompatible herb. J. Ecol. 2020, 108, 81–93. [Google Scholar] [CrossRef]
- Naiki, A. Hetrostyly and the possibility of its breakdown by polyploidization. Plant Species Biol. 2012, 27, 3–29. [Google Scholar] [CrossRef]
- Schmidt-Lebuhn, A.N.; de Vos, J.M.; Keller, B.; Conti, E. Phylogenetic analysis of Primula section Primula reveals rampant non-monophyly among morphologically distinct species. Mol. Phylogenet. Evol. 2012, 65, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Clapham, A.R.; Tutin, T.G.; Moore, D.M. Flora of the British Isles, 3rd ed.; Cambridge University Press: Cambridge, UK, 1987; 688p. [Google Scholar]
- Nowak, M.D.; Russo, G.; Schlapbach, R.; Huu, C.N.; Lenhard, M.; Conti, E. The draft genome of Primula veris yields insights into the molecular basis of heterostyly. Genome Biol. 2015, 16, 12. [Google Scholar] [CrossRef] [PubMed]
- Peev, D. Genus Primula L. In Flora of the Republic of Bulgaria; Velchev, V., Ed.; Bulgarian Academy of Sciences: Sofia, Bulgaria, 1982; Volume 8, pp. 324–336. (In Bulgarian) [Google Scholar]
- Assyov, B.; Petrova, A.; Dimitrov, D.; Vassilev, P. Conspectus of the Bulgarian Vascular Flora. Distribution Maps and Floristic Elements, 4th ed.; Assyov, B., Petrova, A., Eds.; Bulgarian Biodiversity Foundation: Sofia, Bulgaria, 2012; pp. 331–332. [Google Scholar]
- Keller, B.; de Vos, J.M.; Conti, E. Decrease of sexual organ reciprocity between heterostylous primrose species, with possible functional and evolutionary implications. Ann. Bot. 2012, 110, 1233–1244. [Google Scholar] [CrossRef] [PubMed]
- Brys, R.; Jacquemyn, H.; Endels, P.; Van Rossum, F.; Hermy, M.; Triest, L.; De Bruyn, L.; Blust, G.D.E. Reduced reproductive success in small populations of the self-incompatible Primula vulgaris. J. Ecol. 2004, 92, 5–14. [Google Scholar] [CrossRef]
- Van Rossum, F.; Triest, L. Within-population genetic variation in the distylous Primula veris: Does floral morph anisoplethy matter in fragmented habitats? Perspectives in Plant Ecology. Evol. Syst. 2006, 7, 263–273. [Google Scholar]
- Berisha, N.; Millaku, F.; Gashi, B.; Krasniqi, E.; Novak, J. Initial determination of DNA polymorphism of some Primula veris L. populations from Kosovo and Austria. Physiol. Mol. Biol. Plants 2015, 21, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Müller, A.; Ganzera, M.; Stuppner, H. Analysis of phenolic glycosides and saponins in Primula elatior and Primula veris (primula root) by liquid chromatography, evaporative light scattering detection and mass spectrometry. J. Chromatogr. A 2006, 1112, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Bączek, K.; Przybył, J.L.; Mirgos, M.; Kosakowska, O.; Szymborska-Sandhu, I.; Węglarz, Z. Phenolics in Primula veris L. and P. elatior (L.) Hill Raw Materials. Int. J. Anal. Chem. 2017, 1, 2871579. [Google Scholar] [CrossRef] [PubMed]
- Martins, B.C.; Oberprieler, C.; Hellwig, F.H. A phylogenetic analysis of Primulaceae s.l. based on internal transcribed spacer (ITS) DNA sequence data. Plant Syst. Evol. 2003, 237, 75–85. [Google Scholar] [CrossRef]
- Brys, R.; Jacquemyn, H.; Endels, P.; Hermy, M.; De Blust, G. The relationship between reproductive success and demographic structure in remnant populations of Primula veris. Acta Oecol. 2003, 24, 247–253. [Google Scholar] [CrossRef]
- Deschepper, P.; Brys, R.; Jacquemyn, H. The impact of flower morphology and pollinator community composition on pollen transfer in the distylous Primula veris. Bot. J. Linn. Soc. 2017, 186, 414–424. [Google Scholar] [CrossRef]
- Preston, C.D.; Hill, M.O. The geographical relationships of British and Irish vascular plants. Bot. J. Linn. Soc. 1997, 124, 1–120. [Google Scholar] [CrossRef]
- Wichtl, M. Herbal Drugs and Phytopharmaceuticals, A Handbook of Practice on a Scientific Basis, 3rd ed.; Medpharm Scientific Publisher: Stuttgart, Germany, 2004; 704p. [Google Scholar]
- Ornduff, R. Pollen flow in a population of Primula vulgaris Huds. Bot. J. Linn. Soc. 1979, 78, 1–10. [Google Scholar] [CrossRef]
- Allen, D.; Bilz, M.; Leaman, D.J.; Miller, R.M.; Timoshyna, A.; Window, J. European Red List of Medicinal Plants; Publications Office of the European Union: Luxembourg, 2014; 62p. [Google Scholar]
- European Pharmacopoeia. Primula root (Primulae radix). In European Directorate for the Quality of Medicines and Health Care (EDQM), 5th ed.; Council of Europe: Strasbourg, France, 2006; p. 1588. [Google Scholar]
- Biological Diversity Act. 2002. Promulgated, State Gazette SG, No. 77/9.08.2002, Last amended, in SG No. 58/26.07.2016 (Bg). Available online: http://eea.government.bg/bg/legislation/biodiversity/ZBR_en_26_07_2016.pdf (accessed on 2 September 2021).
- Medicinal Plants Act. 2000. Promulgated State Gazette (SG), issue 29/7.04.2000. Last amended in SG, Number 98. 28 November 2014; (In Bulgarian). Available online: http://eea.government.bg/bg/legislation/biodiversity/ZLR_en.pdf (accessed on 2 September 2021).
- EMA (European Medicines Agency). Assessment Report on Primula veris L. and/or Primula elatior (L.) Hill, Flos. EMA/HMPC/136583/2012; European Medicines Agency: Amsterdam, The Netherlands, 2012. [Google Scholar]
- EMA (European Medicines Agency). Assessment Report on Primula veris L. and/or Primula elatior (L.) Hill, Radix. EMA/HMPC/113577/2012; European Medicines Agency: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Tarapatskyy, M.; Gumienna, A.; Sowa, P.; Capusta, I.; Puchalski, C. Bioactive Phenolic Compounds from Primula veris L.: Influence of the Extraction Conditions and Purification. Molecules 2021, 26, 997. [Google Scholar] [CrossRef]
- Pamukov, D.; Ahthardzhiev, H. Natural Pharmacy; Zemizdat: Sofia, Bulgaria, 1989; 327p. (In Bulgarian) [Google Scholar]
- Richards, A.J. Plant Breeding Systems, 2nd ed.; Chapman and Hall: London, UK, 1997. [Google Scholar]
- Brys, R.; Jacquemyn, H. Biological Flora of the British Isles: Primula veris L. J. Ecol. 2009, 97, 581–600. [Google Scholar] [CrossRef]
- <monospace>Cardoso, J.C.; Viana, M.L.; Matias, R.; Furtado, M.T.; Caetano, A.P.; Consolaro, H.; Brito, V.L. Towards a unified terminology for angiosperm reproductive system. Acta Bot. Bras. 2018, 32, 329–348. [Google Scholar]
- Li, J.; Webster, M.A.; Smith, M.C.; Gilmartin, P.M. Floral heteromorphy in Primula vulgaris: Progress towards isolation and characterization of the S locus. Ann. Bot. 2011, 108, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C.R. On the Various Contrivances by Which British and Foreign Orchids Are Fertilized by Insects, and on the Good Effects of Intercrossing; John Murray: London, UK, 1862; pp. 1–377. [Google Scholar]
- Minuto, L.; Guerrina, M.; Roccotiello, E.; Roccatagliata, N.; Mariotti, M.G.; Casazza, G. Pollination ecology in the narrow endemic winter-flowering Primula allionii (Primulaceae). J. Plant Prot. Res. 2014, 127, 141–150. [Google Scholar] [CrossRef]
- Larson, B.M.H.; Barrett, S.C.H. Reproductive biology of island and mainland populations of Primula mistassinica (Primulaceae) on Lake Huron shorelines. Can. J. Bot. 1998, 76, 1819–1827. [Google Scholar]
- Wolff, D.; Liede-Schumann, S. Evolution of flower morphology, pollen dimorphism, and nectar composition in Arcytophyllum, a distylous genus of Rubiaceae. Org. Divers. Evol. 2007, 7, 106–123. [Google Scholar] [CrossRef]
- Dahlgren, K.V.O. Zytologische und embryologische Studien über die Reihen Primulales und Plumpaginales. Kongl. Svenska Vetensk.-Akad. Handl. 1916, 56, 360–363. [Google Scholar]
- Davis, G.L. Systematic Embryology of the Angiosperms; John Wiley: New York, NY, USA, 1966; 528p. [Google Scholar]
- Poddubnaya-Arnoldi, V.A. Characteristics of Flowering Plants Families According to the Cytoembryological Features; Nauka: Moscow, Russia, 1982. (In Russian) [Google Scholar]
- Lersten, N.R.; Eilers, L.J. Binucleate Tapetum in Two Species of Lysimachia (Primulaceae). Proc. Iowa Acad. Sci. 1974, 81, 93–94. [Google Scholar]
- Gachechiladze, M.I. The embryology of Primula bayernii (Primulaceae). Bot. J. 1993, 78, 93–96. (In Russian) [Google Scholar]
- Mameteva, T.B. Family Primulaceae. In Comparative Embryology of Flowering Plants; Yakovlev, M.S., Ed.; Nauka: Leningrad, Russia, 1983; pp. 241–243. (In Russian) [Google Scholar]
- Anderberg, A.A. Primulaceae. In The Families and Genera of Vascular Plants; Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 2004; Volume 6, pp. 313–319. [Google Scholar]
- Morozowska, M.; Idzikowska, K. Morphological differentiation of Primula veris L. pollen from natural and cultivated populations. Acta Soc. Bot. Pol. 2004, 73, 229–232. [Google Scholar] [CrossRef][Green Version]
- Kálmán, K.; Medvegy, A.; Mihalik, E. Quantitative characteristics of the gynoecium in Primula vulgaris. Acta Biol. Szeged. 1999, 43, 5–14. [Google Scholar]
- Petrova, S.E.; Kozhin, M.N. Heterostyly in Primula nutans ssp. finmarchica (Jacq.) Á. Löve & D. Löve (Primulaceae) from three northern coenopopulations (Kandalaksha Bay, White Sea). Wulfenia 2018, 25, 231–241. [Google Scholar]
- Akhalkatsi, M.; Gvaladze, G.; Taralashvili, N. Embryology of Primula algida and Primula amoena (Primulaceae). Bull. Georg. Aca. Sci. 1998, 157, 98–101. [Google Scholar]
- Raju, M.V.S. Embryology of Anagallis pumila Swartz. Proc. Indian Acad. Sci. Sect. B 1952, 36, 34–42. [Google Scholar] [CrossRef]
- Kapil, N.R.; Tiwari, S.C. The integumentary tapetum. Bot. Rev. 1978, 44, 457–490. [Google Scholar] [CrossRef]
- Van Rossum, F.; De Sousa, S.C.; Triest, L. Morph-specific differences in reproductive success in the distylous Primula veris in a context of habitat fragmentation. Acta Oecol. 2006, 30, 426–433. [Google Scholar] [CrossRef]
- Veillet-Bartoszewska, M. La polyembryonie chez le Primula auricula L. Bull. Soc. bot. Fr. 1957, 104, 473–475. [Google Scholar] [CrossRef]
- Maheshwari, P. Polyembryony in Angiosperms. Palaeobotanist 1952, 1, 319–329. [Google Scholar]
- Dafni, A.; Firmage, D. Pollen viability and longevity: Practical, ecological and evolutionary implications. Plant Syst. Evol. 2000, 222, 113–132. [Google Scholar] [CrossRef]
- Lincoln, R.J.; Boxshall, G.A.; Clark, P.F. A Dictionary of Ecology, Evolution and Systematics; Cambridge University Press: New York, NY, USA, 1982; 313p. [Google Scholar]
- Aronne, G.; Iovane, M.; Strumia, S. Temperature and humidity affect pollen viability and may trigger distyly disruption in threatened species. Ann. Bot. 2021, 11, 77–82. [Google Scholar]
- Copeland, L.O.; McDonald, M.F. Seed Viability and Viability Testing. In Principles of Seed Science and Technology, 4th ed.; Copeland, L.O., McDonald, M.F., Eds.; Kluwer Academic Publisher: New York, USA, 2001; pp. 124–139. [Google Scholar]
- AOSA. Tetrazolium Testing Handbook; Contribution No. 29; Association of Official Seed Analysts: Lincoln, NE, USA, 2000; Available online: http://gsem.weebly.com/uploads/9/3/5/1/9351412/tetrazolium_testing_handbook__2001-2002__-_part_i.pdf (accessed on 2 September 2021).
- Moore, R.P. Handbook on Tetrazolium Testing; International Seed Testing Association: Zürich, Switzerland, 1985; 99p. [Google Scholar]
- Richards, A.J. Primulas of the British Isles; Shire Publications: Princes Risborough, UK, 1989; 24p. [Google Scholar]
- Jacquemyn, H.; Brys, R. Population growth rates of the forest herb Primula elatior increase with stand age in post-agricultural forests. Ecology 2008, 89, 3480–3489. [Google Scholar] [CrossRef] [PubMed]
- Grime, J.P.; Hodgson, J.G.; Hunt, R. Comparative Plant Ecology. A Functional Approach to Common British Species; Unwin Hyman: London, UK, 1988; 679p. [Google Scholar]
- Thompson, K.; Bakker, J.P.; Bekker, R.M. The Soil Seed Bank of North West Europe: Methodology, Density and Longevity; Cambridge University Press: Cambridge, UK, 1988. [Google Scholar]
- Antrobus, S.; Lack, A.J. Genetics of colonizing and established populations of Primula veris. Heredity 1993, 71, 252–258. [Google Scholar] [CrossRef][Green Version]
- Milberg, P. Germination ecology of the polycarpic grassland perennials Primula veris and Trollius europaeus. Ecography 1994, 17, 3–8. [Google Scholar] [CrossRef]
- Crema, S.; Christofolini, G.; Rossi, M.; Conte, L. High genetic diversity detected in the endemic Primula apennina Widmer (Primulaceae) using ISSR fingerprinting. Plant Syst. Evol. 2009, 280, 29–36. [Google Scholar] [CrossRef]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef]
- Noroozisharafa, A.; Hatamzadeha, A.; Samizadeh Lahiji, H.; Bakhshi, D. Genetic diversity of endangered primrose (Primula heterochroma Stapf.) accessions from Iran revealed by ISSR and IRAP markers. Sci. Hortic. 2015, 190, 173–178. [Google Scholar] [CrossRef]
- Rossetto, M.; Weaver, P.K.; Dixon, K.W. Use of RAPD analysis indevising conservation strategies for the rare and endangered Grevillea scapigera (Proteaceae). Mol. Ecol. 1995, 4, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Peev, D. Chromosome numbers and critical notes on the taxonomy of some Primulaceae species in Bulgaria. Fitologiya 1976, 4, 16–24. [Google Scholar]
- Nikolov, N.A. Chromosome numbers of Bulgarian Angiosperms from North Pirin Mountain: Reserve “Bajuvi Dupki -Džindžirica”. Fitologiya 1991, 41, 70–75. [Google Scholar]
- Romeis, B. Mikroskopische Technik; Oldenbourg Wissenschaftsverlag: München, Germany, 1948; 695p. [Google Scholar]
- Heidenhain, R. Eine neue Verwendung des haematoxylin. Archiv. Mikrosk. Anat. 1885, 24, 468–470. [Google Scholar] [CrossRef]
- Singh, R.J. Plant Cytogenetics, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2003; 488p. [Google Scholar]
- Peters, J. (Ed.) Tetrazolium Testing Handbook; Association of Official Seed Analysts: Ithaca, NY, USA, 2007; 88p. [Google Scholar]
- Petrova, G.; Dzhambazova, T.; Moyankova, D.; Georgieva, D.; Michova, A.; Djilianov, D.; Möller, M. Morphological variation, genetic diversity and genome size of critically endangered Haberlea (Gesneriaceae) populations in Bulgaria do not support the recognition of two different species. Plant Syst. Evol. 2013, 300, 29–41. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Husband, B.C.; Schemske, D.W. Evolution of the magnitude and timing of inbreeding depression in plants. Evolution 1996, 50, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Kéry, M.; Matthies, D.; Spillman, H.H. Reduced fertility and offspring performance in small populations of the declining grassland plants Primula veris and Gentiana lutea. J. Ecol. 2000, 88, 17–30. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Number of Tested Seeds | Number of Viable Embryos | Viable Embryos % | Unviable Embryos % |
|---|---|---|---|---|
| Golo Bardo Mt | 50 | 2 | 4 | 96 |
| Ilindentsi village | 50 | 1 | 2 | 98 |
| Population | Number of Tested Seeds | Number of Viable Embryos | Viable Embryos % | Unviable Embryos % |
|---|---|---|---|---|
| Golo Bardo Mt | 50 | 34 | 68 | 32 |
| Ilindentsi village | 50 | 38 | 76 | 24 |
| Population | Geographic Coordinates | Altitude (m) | Percent of Polymorphic Bands | Gene Diversity He (±SD) | Average Intra-Population Diversity HS (±SD) |
|---|---|---|---|---|---|
| Shabran (Slavyanka Mts) | 41°23′19″ N 23°36′30″ E | 1900 | 69 | 0.238 ± 0.155 | 0.236 ± 0.249 |
| Kraishte (Western Rhodopes) | 41°57′28″ N 23°39′12″ E | 1180 | 74 | 0.256 ± 0.108 | 0.302 ± 0.248 |
| Petrokhan (Western Stara planina) | 43°7′39″ N 23°7′28″ E | 1450 | 72 | 0.192 ± 0.134 | 0.256 ± 0.154 |
| Gorno Harsovo (Rila Mts) | 42°00′47″ N 23°12′30″ E | 820 | 67 | 0.201 ± 0.164 | 0.238 ± 0.203 |
| ISSR Primer (Name and Sequence) | Optimal Annealing Temperature | Total Number of Bands | Number of Polymorphic Bands |
|---|---|---|---|
| UBC-807 AGAGAGAGAGAGAGAGT | 59 | 16 | 11 |
| UBC-808 AGAGAGAGAGAGAGAGC | 57 | 10 | 8 |
| UBC-811 GAGAGAGAGAGAGAGAC | 53 | 12 | 10 |
| UBC-827 ACACACACACACACACG | 57 | 6 | 5 |
| UBC-835 AGAGAGAGAGAGAGAGYC | 54 | 19 | 17 |
| UBC-845 CTCTCTCT CTCTCTCTRG | 54 | 21 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yankova-Tsvetkova, E.; Yurukova-Grancharova, P.; Aneva, I.; Zhelev, P. On the Reproductive Potential in Primula veris L. (Primulaceae): Embryological Features, Pollen and Seed Viability, Genetic Diversity. Plants 2021, 10, 2296. https://doi.org/10.3390/plants10112296
Yankova-Tsvetkova E, Yurukova-Grancharova P, Aneva I, Zhelev P. On the Reproductive Potential in Primula veris L. (Primulaceae): Embryological Features, Pollen and Seed Viability, Genetic Diversity. Plants. 2021; 10(11):2296. https://doi.org/10.3390/plants10112296
Chicago/Turabian StyleYankova-Tsvetkova, Elina, Petka Yurukova-Grancharova, Ina Aneva, and Petar Zhelev. 2021. "On the Reproductive Potential in Primula veris L. (Primulaceae): Embryological Features, Pollen and Seed Viability, Genetic Diversity" Plants 10, no. 11: 2296. https://doi.org/10.3390/plants10112296
APA StyleYankova-Tsvetkova, E., Yurukova-Grancharova, P., Aneva, I., & Zhelev, P. (2021). On the Reproductive Potential in Primula veris L. (Primulaceae): Embryological Features, Pollen and Seed Viability, Genetic Diversity. Plants, 10(11), 2296. https://doi.org/10.3390/plants10112296