
Ubiquitin-Specific Protease Inhibitors for Cancer Therapy: Recent Advances and Future Prospects

 ,
,  ,
,  and
and 
Abstract
1. Introduction
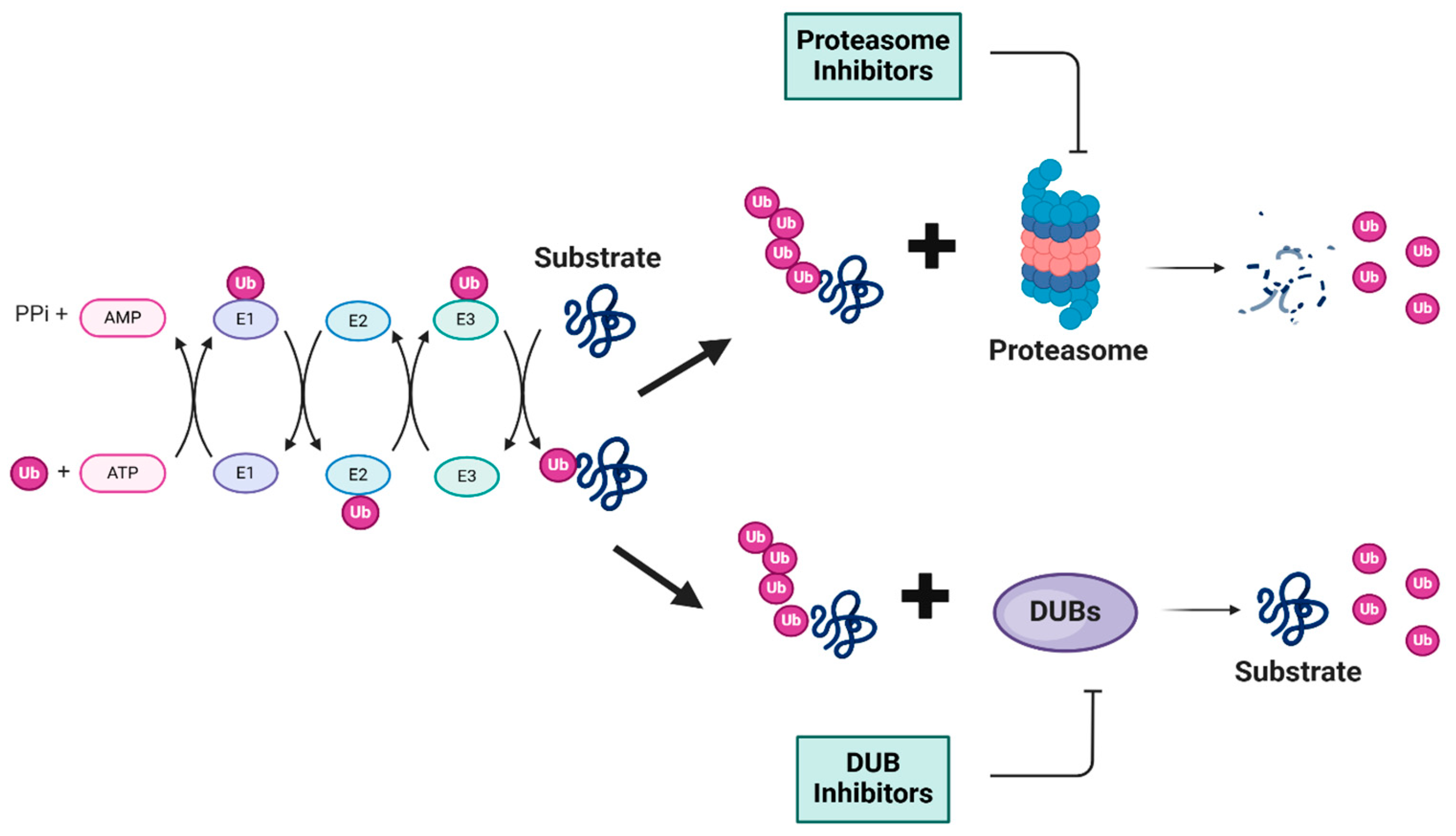
1.1. Overview of the Ubiquitin–Proteasome Pathway
1.2. Role of Deubiquitinating Enzymes
1.2.1. Metalloprotease DUBs
1.2.2. Cysteine Protease DUBs
Ubiquitin C-Terminal Hydrolases (UCHs)
Ovarian Tumor Proteases (OTUs)
Machado–Josephin Domain Proteases (MJDs)
Motif Interacting with Ubiquitin (MIU)-Containing Novel DUB Family (MINDY) and Zinc Finger with UFM1-Specific Peptidase Domain Protein (ZUFSP)
Ubiquitin-Specific Proteases (USPs)
2. USPs: Structure, Biological Function, and Role in Tumorigenesis and Cancer Progression
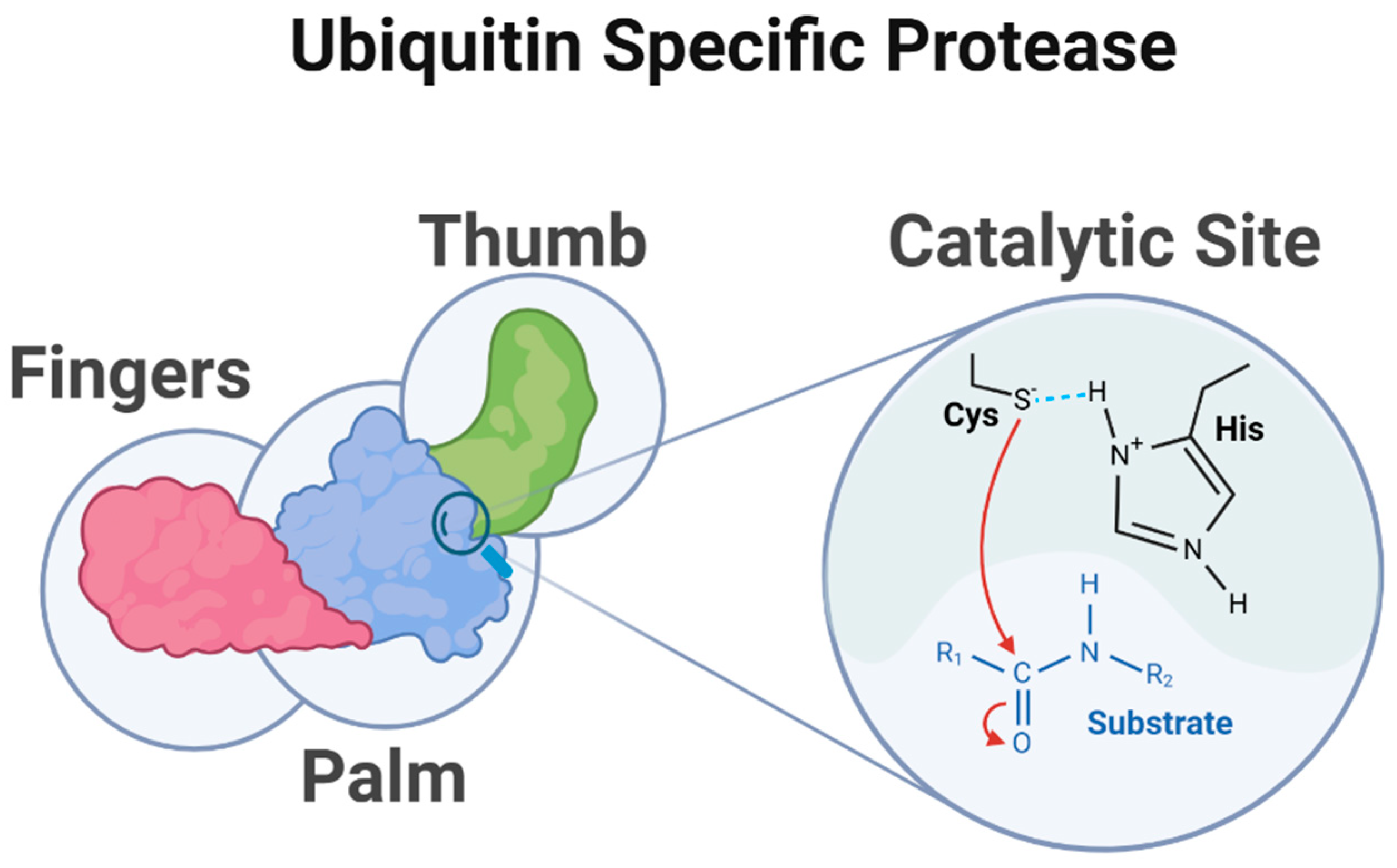
2.1. The Structure of USPs
2.1.1. Core Catalytic Domain
2.1.2. Ubiquitin-like Domains (UBL)
2.1.3. Ancillary Domains
2.2. The Roles of USPs in Tumorigenesis and Cancer Progression
2.3. Unique Structural and Functional Characteristics of Various USPs

{kind=link}
{kind=link}
{kind=link}
| USP Name | Domains | Catalytic Site | Cancer Association | References |
|---|---|---|---|---|
| USP1 | Conserved USP catalytic domain (thumb, palm, and finger sub-domains). | Three catalytic residues: Cys90, His593, and Asp751. | Osteosarcoma, renal clear cell carcinoma, colorectal carcinoma, non-small cell lung cancer, and gastric cancers | [42] |
| USP3 | Zinc finger (ZnF) domain and catalytic domain. | Conserved catalytic domain containing key Cys, Hys, and Asp/Asn residues critical for USP activity. | Gastric cancer and breast cancer | [55,56] |
| USP5 | Catalytic domain flanked by two ubiquitin-associated (UBA) domains and two Zinc finger (ZnF) ubiquitin-binding protein (UBP) domains. | Papain-like proteases have a Cys and a His residue, which form an ion pair with the negatively charged cysteine thiolate functioning as a nucleophile. | Breast, lung, colorectal, hepatocellular, pancreatic, and non-small cell lung cancer | [57,58,59] |
| USP7 | N-terminal MATH domain, central catalytic domain, and five C-terminal tandem ubiquitin-like domains (UbL). | Cys223, His464, and Asp481 residues are critical for deubiquitination. | Colorectal cancer, osteosarcoma, acute myeloid leukemia, breast cancer, prostate cancer, multiple myeloma, ovarian cancer, bladder cancer, esophageal squamous cell carcinoma, chronic lymphocytic lymphoma, and medulloblastoma | [54,60,61] |
| USP10 | N-terminus region, USP catalytic structure domain, and a smaller C-terminus region. | Conserved catalytic domain containing key Cys, His, and Asp/Asn residues critical for USP activity. | Colorectal cancer, prostate cancer, hepatocellular carcinoma, glioblastoma multiforme, non-small-cell lung cancer, chronic myeloid leukemia, and acute myeloid leukemia | [62] |
| USP19 | Two CHORD-SGT1/P23 domains (CS1 and CS2 at the N-terminus), USP catalytic domain. | Contains Cys and His residues critical for enzymatic activity and auto-inhibition mechanism. | Breast cancer and osteosarcoma | [63,64] |
| USP20 | ZnF-UBP domain, catalytic USP domain, and two domains present in ubiquitin-specific protease (DUSP) domains. | Conserved Cys and His residues that catalyze proteolysis of the isopeptide bond between a target protein lysine residue and a glycine residue of ubiquitin. | Breast cancer, lung cancer, colon cancer, gastric cancer, and adult T-cell leukemia | [44,65] |
| USP32 | USP catalytic domain, Domain in USP (DUSP), two UbL domains, and calcium-binding EF-hand with a signal transduction mechanism. | Conserved catalytic domain containing key Cys and His residues critical for USP activity. | Small cell lung cancer, gastric cancer, breast cancer, epithelial ovarian cancer, glioblastoma, gastrointestinal stromal tumor, AML, and pancreatic adenocarcinoma | [66] |
| USP36 | Conserved USP catalytic domain (thumb, palm, and finger sub-domains). | Key residues: Cys223, His464, Asp481 in the catalytic domain. | Colon cancer and breast cancer | [36,67] |
2.4. History of the Development of USP Inhibitors
2.5. Therapeutic Implications
3. Small Molecule Inhibitors Targeting USPs
3.1. Chemical Structure, Inhibition Potency, and Mechanism of Action of Small Molecule Inhibitors Targeting USPs
3.2. USP Inhibitors and Immunotherapy
3.2.1. Role of USP1 in Immune Response
3.2.2. Role of USP7 in Immune Response
3.2.3. Enhancing Immunotherapy by USP Inhibitors
3.3. Development of USP Inhibitors for Cancer Therapy: Clinical Trials Status
3.3.1. USP1 Inhibitors
3.3.2. USP14 Inhibitor
| Chemical Structure | Type of USP Inhibitor | Clinical Trials.gov ID | Trial Phase | Study Design | Results | References |
|---|---|---|---|---|---|---|
TNG348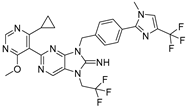 | USP1 IC50: 82 nM | NCT06065059 | Phase 1 dose escalation and Phase 2 dose expansion trial (terminated) |
|
| [194,200,201,202] |
KSQ-4279, also known as RO7623066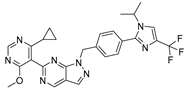 | USP1 IC50: 10 nM | NCT05240898 | Phase 1 (recruiting) |
|
| [71,196,203,,204] |
| XL309, also known as ISM3091 Chemical structure is not publicly disclosed | USP1 | NCT05932862 | Phase 1 (recruiting) |
|
| [197,205,206] |
| SIM0501 Chemical structure is not publicly disclosed. | USP1 | NCT06331559 | Phase 1 (recruiting) |
|
| [197,,207,208] |
| HSK39775 Chemical structure is not publicly disclosed. | USP1 | NCT06314373 | Phase 1 dose escalation and Phase 2 dose expansion trial (recruiting) |
|
| [197,209,210] |
VLX1570 | USP14/UCHL5 IC50: ~10 µM | NCT02372240 | Phase 1 dose escalation and Phase 2 dose expansion trial (terminated) |
|
| [198,199,211,212] |
4. Summary and Future Directions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bedford, L.; Lowe, J.; Dick, L.R.; Mayer, R.J.; Brownell, J.E. Ubiquitin-like protein conjugation and the ubiquitin-proteasome system as drug targets. Nat. Rev. Drug Discov. 2011, 10, 29–46. [Google Scholar] [CrossRef]
- Du, W.; Mei, Q.B. Ubiquitin-proteasome system, a new anti-tumor target. Acta Pharmacol. Sin. 2013, 34, 187–188. [Google Scholar] [CrossRef] [PubMed]
- Arkwright, R.; Pham, T.M.; Zonder, J.A.; Dou, Q.P. The preclinical discovery and development of bortezomib for the treatment of mantle cell lymphoma. Expert. Opin. Drug Discov. 2017, 12, 225–235. [Google Scholar] [CrossRef]
- Teicher, B.A.; Tomaszewski, J.E. Proteasome inhibitors. Biochem. Pharmacol. 2015, 96, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Snyder, N.A.; Silva, G.M. Deubiquitinating enzymes (DUBs): Regulation, homeostasis, and oxidative stress response. J. Biol. Chem. 2021, 297, 101077. [Google Scholar] [CrossRef]
- Johnston, S.C.; Larsen, C.N.; Cook, W.J.; Wilkinson, K.D.; Hill, C.P. Crystal structure of a deubiquitinating enzyme (human UCH-L3) at 1.8 A resolution. EMBO J. 1997, 16, 3787–3796. [Google Scholar] [CrossRef]
- Fang, Y.; Fu, D.; Shen, X.Z. The potential role of ubiquitin c-terminal hydrolases in oncogenesis. Biochim. Biophys. Acta 2010, 1806, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Lu, K.; Jiao, Q.; Chen, X.; Jia, F. The Regulation and Double-Edged Roles of the Deubiquitinase OTUD5. Cells 2023, 12, 1161. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, Y.; Okamoto, T.; Taniwaki, M.; Aizawa, M.; Inoue, M.; Katayama, S.; Kawakami, H.; Nakamura, S.; Nishimura, M.; Akiguchi, I.; et al. CAG expansions in a novel gene for Machado-Joseph disease at chromosome 14q32.1. Nat. Genet. 1994, 8, 221–228. [Google Scholar] [CrossRef]
- Shi, Z.; Chen, J.; Zhang, X.; Chu, J.; Han, Z.; Xu, D.; Gan, S.; Pan, X.; Ye, J.; Cui, X. Ataxin-3 promotes testicular cancer cell proliferation by inhibiting anti-oncogene PTEN. Biochem. Biophys. Res. Commun. 2018, 503, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Abdul Rehman, S.A.; Armstrong, L.A.; Lange, S.M.; Kristariyanto, Y.A.; Grawert, T.W.; Knebel, A.; Svergun, D.I.; Kulathu, Y. Mechanism of activation and regulation of deubiquitinase activity in MINDY1 and MINDY2. Mol. Cell 2021, 81, 4176–4190.e6. [Google Scholar] [CrossRef]
- Abdul Rehman, S.A.; Kristariyanto, Y.A.; Choi, S.Y.; Nkosi, P.J.; Weidlich, S.; Labib, K.; Hofmann, K.; Kulathu, Y. MINDY-1 Is a Member of an Evolutionarily Conserved and Structurally Distinct New Family of Deubiquitinating Enzymes. Mol. Cell 2016, 63, 146–155. [Google Scholar] [CrossRef]
- Trulsson, F.; Akimov, V.; Robu, M.; van Overbeek, N.; Berrocal, D.A.P.; Shah, R.G.; Cox, J.; Shah, G.M.; Blagoev, B.; Vertegaal, A.C.O. Deubiquitinating enzymes and the proteasome regulate preferential sets of ubiquitin substrates. Nat. Commun. 2022, 13, 2736. [Google Scholar] [CrossRef] [PubMed]
- Hermanns, T.; Pichlo, C.; Woiwode, I.; Klopffleisch, K.; Witting, K.F.; Ovaa, H.; Baumann, U.; Hofmann, K. A family of unconventional deubiquitinases with modular chain specificity determinants. Nat. Commun. 2018, 9, 799. [Google Scholar] [CrossRef]
- Harper, S.; Gratton, H.E.; Cornaciu, I.; Oberer, M.; Scott, D.J.; Emsley, J.; Dreveny, I. Structure and catalytic regulatory function of ubiquitin specific protease 11 N-terminal and ubiquitin-like domains. Biochemistry 2014, 53, 2966–2978. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.; Soares, P.; Correia, M. Ubiquitin-Specific Proteases: Players in Cancer Cellular Processes. Pharmaceuticals 2021, 14, 848. [Google Scholar] [CrossRef]
- Ye, Y.; Scheel, H.; Hofmann, K.; Komander, D. Dissection of USP catalytic domains reveals five common insertion points. Mol. Biosyst. 2009, 5, 1797–1808. [Google Scholar] [CrossRef]
- Harrigan, J.A.; Jacq, X.; Martin, N.M.; Jackson, S.P. Deubiquitylating enzymes and drug discovery: Emerging opportunities. Nat. Rev. Drug Discov. 2018, 17, 57–78. [Google Scholar] [CrossRef]
- Reyes-Turcu, F.E.; Wilkinson, K.D. Polyubiquitin binding and disassembly by deubiquitinating enzymes. Chem. Rev. 2009, 109, 1495–1508. [Google Scholar] [CrossRef]
- Reyes-Turcu, F.E.; Ventii, K.H.; Wilkinson, K.D. Regulation and cellular roles of ubiquitin-specific deubiquitinating enzymes. Annu. Rev. Biochem. 2009, 78, 363–397. [Google Scholar] [CrossRef]
- Komander, D. Mechanism, specificity and structure of the deubiquitinases. Subcell. Biochem. 2010, 54, 69–87. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Zhou, Z.; Shah, A.A.; Zou, H.; Tao, J.; Chen, Q.; Wan, Y. The emerging role of deubiquitinating enzymes in genomic integrity, diseases, and therapeutics. Cell Biosci. 2016, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Dikic, I.; Wakatsuki, S.; Walters, K.J. Ubiquitin-binding domains—From structures to functions. Nat. Rev. Mol. Cell Biol. 2009, 10, 659–671. [Google Scholar] [CrossRef]
- Drag, M.; Mikolajczyk, J.; Bekes, M.; Reyes-Turcu, F.E.; Ellman, J.A.; Wilkinson, K.D.; Salvesen, G.S. Positional-scanning fluorigenic substrate libraries reveal unexpected specificity determinants of DUBs (deubiquitinating enzymes). Biochem. J. 2008, 415, 367–375. [Google Scholar] [CrossRef]
- Fan, Y.H.; Yu, Y.; Mao, R.F.; Tan, X.J.; Xu, G.F.; Zhang, H.; Lu, X.B.; Fu, S.B.; Yang, J. USP4 targets TAK1 to downregulate TNFalpha-induced NF-kappaB activation. Cell Death Differ. 2011, 18, 1547–1560. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Tan, X.; Wang, H.; Sun, W.; Shi, Y.; Burlingame, S.; Gu, X.; Cao, G.; Zhang, T.; Qin, J.; et al. Ubiquitin-specific peptidase 21 inhibits tumor necrosis factor alpha-induced nuclear factor kappaB activation via binding to and deubiquitinating receptor-interacting protein 1. J. Biol. Chem. 2010, 285, 969–978. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Gaynor, R.B. Therapeutic potential of inhibition of the NF-kappaB pathway in the treatment of inflammation and cancer. J. Clin. Investig. 2001, 107, 135–142. [Google Scholar] [CrossRef]
- Yun, S.I.; Kim, H.H.; Yoon, J.H.; Park, W.S.; Hahn, M.J.; Kim, H.C.; Chung, C.H.; Kim, K.K. Ubiquitin specific protease 4 positively regulates the WNT/beta-catenin signaling in colorectal cancer. Mol. Oncol. 2015, 9, 1834–1851. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, N.; Kurzrock, R. Targeting the Wnt/beta-catenin pathway in cancer: Update on effectors and inhibitors. Cancer Treat. Rev. 2018, 62, 50–60. [Google Scholar] [CrossRef]
- Hu, X.; Li, J.; Fu, M.; Zhao, X.; Wang, W. The JAK/STAT signaling pathway: From bench to clinic. Signal Transduct. Target. Ther. 2021, 6, 402. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.; Liu, C.; Guan, X.; Wang, B.; Yao, Y.; Su, D.; Ma, Y.; Fang, L.; Zhang, Y. Ubiquitin specific peptidase 5 enhances STAT3 signaling and promotes migration and invasion in Pancreatic Cancer. J. Cancer 2020, 11, 6802–6811. [Google Scholar] [CrossRef] [PubMed]
- Qureshy, Z.; Johnson, D.E.; Grandis, J.R. Targeting the JAK/STAT pathway in solid tumors. J. Cancer Metastasis Treat 2020, 6, 21. [Google Scholar] [CrossRef]
- Lee, J.T.; Gu, W. The multiple levels of regulation by p53 ubiquitination. Cell Death Differ. 2010, 17, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, T.; Su, W.; Dou, Z.; Zhao, D.; Jin, X.; Lei, H.; Wang, J.; Xie, X.; Cheng, B.; et al. Mutant p53 in cancer: From molecular mechanism to therapeutic modulation. Cell Death Dis. 2022, 13, 974. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Yu, W.; Li, J.; Ma, H.; Wang, P.; Zhou, Y.; Wang, Y.; Zhang, J.; Shi, G. USP16 regulates castration-resistant prostate cancer cell proliferation by deubiquitinating and stablizing c-Myc. J. Exp. Clin. Cancer Res. 2021, 40, 59. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Luo, M. Cinobufotalin regulates the USP36/c-Myc axis to suppress malignant phenotypes of colon cancer cells in vitro and in vivo. Aging 2024, 16, 5526–5544. [Google Scholar] [CrossRef]
- Ruiz, E.J.; Pinto-Fernandez, A.; Turnbull, A.P.; Lan, L.; Charlton, T.M.; Scott, H.C.; Damianou, A.; Vere, G.; Riising, E.M.; Da Costa, C.; et al. USP28 deletion and small-molecule inhibition destabilizes c-MYC and elicits regression of squamous cell lung carcinoma. Elife 2021, 10, e71596. [Google Scholar] [CrossRef]
- Guo, J.; Zhao, J.; Sun, L.; Yang, C. Role of ubiquitin specific proteases in the immune microenvironment of prostate cancer: A new direction. Front. Oncol. 2022, 12, 955718. [Google Scholar] [CrossRef]
- Ji, M.; Shi, H.; Xie, Y.; Zhao, Z.; Li, S.; Chang, C.; Cheng, X.; Li, Y. Ubiquitin specific protease 22 promotes cell proliferation and tumor growth of epithelial ovarian cancer through synergy with transforming growth factor beta1. Oncol. Rep. 2015, 33, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xiang, H.; Lu, Y.; Wu, T. Role and clinical significance of TGF-beta1 and TGF-betaR1 in malignant tumors (Review). Int. J. Mol. Med. 2021, 47, 55. [Google Scholar] [CrossRef]
- Liu, J.; Kruswick, A.; Dang, H.; Tran, A.D.; Kwon, S.M.; Wang, X.W.; Oberdoerffer, P. Ubiquitin-specific protease 21 stabilizes BRCA2 to control DNA repair and tumor growth. Nat. Commun. 2017, 8, 137. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.M.; Kim, S.; Seo, S.U.; Kim, S.; Park, J.W.; Kim, G.; Choi, Y.R.; Hur, K.; Kwon, T.K. Inhibition of USP1 enhances anticancer drugs-induced cancer cell death through downregulation of survivin and miR-216a-5p-mediated upregulation of DR5. Cell Death Dis. 2022, 13, 821. [Google Scholar] [CrossRef]
- Foster, B.M.; Wang, Z.; Schmidt, C.K. DoUBLing up: Ubiquitin and ubiquitin-like proteases in genome stability. Biochem. J. 2024, 481, 515–545. [Google Scholar] [CrossRef]
- Li, Q.; Ye, C.; Tian, T.; Jiang, Q.; Zhao, P.; Wang, X.; Liu, F.; Shan, J.; Ruan, J. The emerging role of ubiquitin-specific protease 20 in tumorigenesis and cancer therapeutics. Cell Death Dis. 2022, 13, 434. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, S.; Narita, M.; Tsujimoto, Y. Bcl-2 family proteins regulate the release of apoptogenic cytochrome c by the mitochondrial channel VDAC. Nature 1999, 399, 483–487. [Google Scholar] [CrossRef]
- Trivigno, D.; Essmann, F.; Huber, S.M.; Rudner, J. Deubiquitinase USP9x confers radioresistance through stabilization of Mcl-1. Neoplasia 2012, 14, 893–904. [Google Scholar] [CrossRef]
- Ma, C.; Wang, D.; Tian, Z.; Gao, W.; Zang, Y.; Qian, L.; Xu, X.; Jia, J.; Liu, Z. USP13 deubiquitinates and stabilizes cyclin D1 to promote gastric cancer cell cycle progression and cell proliferation. Oncogene 2023, 42, 2249–2262. [Google Scholar] [CrossRef]
- Li, G.; Yang, T.; Chen, Y.; Bao, J.; Wu, D.; Hu, X.; Feng, C.; Xu, L.; Li, M.; Li, G.; et al. USP5 Sustains the Proliferation of Glioblastoma Through Stabilization of CyclinD1. Front. Pharmacol. 2021, 12, 720307. [Google Scholar] [CrossRef]
- Zhang, Z.; Cui, Z.; Xie, Z.; Li, C.; Xu, C.; Guo, X.; Yu, J.; Chen, T.; Facchinetti, F.; Bohnenberger, H.; et al. Deubiquitinase USP5 promotes non-small cell lung cancer cell proliferation by stabilizing cyclin D1. Transl. Lung Cancer Res. 2021, 10, 3995–4011. [Google Scholar] [CrossRef]
- Sun, H.; Ou, B.; Zhao, S.; Liu, X.; Song, L.; Liu, X.; Wang, R.; Peng, Z. USP11 promotes growth and metastasis of colorectal cancer via PPP1CA-mediated activation of ERK/MAPK signaling pathway. EBioMedicine 2019, 48, 236–247. [Google Scholar] [CrossRef]
- Brahimi-Horn, C.; Pouyssegur, J. When hypoxia signalling meets the ubiquitin-proteasomal pathway, new targets for cancer therapy. Crit. Rev. Oncol. Hematol. 2005, 53, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.C.; Zhao, C.J.; Jin, Z.F.; Zheng, J.; Ma, L.T. Targeted therapy based on ubiquitin-specific proteases, signalling pathways and E3 ligases in non-small-cell lung cancer. Front. Oncol. 2023, 13, 1120828. [Google Scholar] [CrossRef]
- Keijzer, N.; Priyanka, A.; Stijf-Bultsma, Y.; Fish, A.; Gersch, M.; Sixma, T.K. Variety in the USP deubiquitinase catalytic mechanism. Life Sci. Alliance 2024, 7, e202302533. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, Y.; Zhou, H. Advances in the Development Ubiquitin-Specific Peptidase (USP) Inhibitors. Int. J. Mol. Sci. 2021, 22, 4546. [Google Scholar] [CrossRef]
- Wang, Y.; Shi, Y.; Niu, K.; Yang, R.; Lv, Q.; Zhang, W.; Feng, K.; Zhang, Y. Ubiquitin specific peptidase 3: An emerging deubiquitinase that regulates physiology and diseases. Cell Death Discov. 2024, 10, 243. [Google Scholar] [CrossRef]
- Wu, X.; Wang, H.; Zhu, D.; Chai, Y.; Wang, J.; Dai, W.; Xiao, Y.; Tang, W.; Li, J.; Hong, L.; et al. USP3 promotes gastric cancer progression and metastasis by deubiquitination-dependent COL9A3/COL6A5 stabilisation. Cell Death Dis. 2021, 13, 10. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.; Layfield, R.; Oldham, N.J. Structural insights into interactions between ubiquitin specific protease 5 and its polyubiquitin substrates by mass spectrometry and ion mobility spectrometry. Protein Sci. 2015, 24, 1257–1263. [Google Scholar] [CrossRef] [PubMed]
- Novinec, M.; Lenarcic, B. Papain-like peptidases: Structure, function, and evolution. Biomol. Concepts 2013, 4, 287–308. [Google Scholar] [CrossRef]
- Yan, B.; Guo, J.; Deng, S.; Chen, D.; Huang, M. A pan-cancer analysis of the role of USP5 in human cancers. Sci. Rep. 2023, 13, 8972. [Google Scholar] [CrossRef]
- Nie, L.; Wang, C.; Liu, X.; Teng, H.; Li, S.; Huang, M.; Feng, X.; Pei, G.; Hang, Q.; Zhao, Z.; et al. USP7 substrates identified by proteomics analysis reveal the specificity of USP7. Genes. Dev. 2022, 36, 1016–1030. [Google Scholar] [CrossRef] [PubMed]
- Korenev, G.; Yakukhnov, S.; Druk, A.; Golovina, A.; Chasov, V.; Mirgayazova, R.; Ivanov, R.; Bulatov, E. USP7 Inhibitors in Cancer Immunotherapy: Current Status and Perspective. Cancers 2022, 14, 5539. [Google Scholar] [CrossRef]
- Tao, L.; Liu, X.; Jiang, X.; Zhang, K.; Wang, Y.; Li, X.; Jiang, S.; Han, T. USP10 as a Potential Therapeutic Target in Human Cancers. Genes. 2022, 13, 831. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.A.; Rossi, M. Emerging Role of Ubiquitin-Specific Protease 19 in Oncogenesis and Cancer Development. Front. Cell Dev. Biol. 2022, 10, 889166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; van Dinther, M.; Thorikay, M.; Gourabi, B.M.; Kruithof, B.P.T.; Ten Dijke, P. Opposing USP19 splice variants in TGF-beta signaling and TGF-beta-induced epithelial-mesenchymal transition of breast cancer cells. Cell Mol. Life Sci. 2023, 80, 43. [Google Scholar] [CrossRef] [PubMed]
- Komander, D.; Clague, M.J.; Urbe, S. Breaking the chains: Structure and function of the deubiquitinases. Nat. Rev. Mol. Cell Biol. 2009, 10, 550–563. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Song, Y.; Wang, K.; Liu, G.; Dong, X.; Yang, F.; Chen, G.; Cao, C.; Zhang, H.; Wang, M.; et al. USP32 deubiquitinase: Cellular functions, regulatory mechanisms, and potential as a cancer therapy target. Cell Death Discov. 2023, 9, 338. [Google Scholar] [CrossRef]
- O’Dea, R.; Kazi, N.; Hoffmann-Benito, A.; Zhao, Z.; Recknagel, S.; Wendrich, K.; Janning, P.; Gersch, M. Molecular basis for ubiquitin/Fubi cross-reactivity in USP16 and USP36. Nat. Chem. Biol. 2023, 19, 1394–1405. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Young, M.A.; Donato, N.J. Emerging potential of therapeutic targeting of ubiquitin-specific proteases in the treatment of cancer. Cancer Res. 2014, 74, 4955–4966. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.L.; Shen, G.T.; Li, N.L. Emerging potential of ubiquitin-specific proteases and ubiquitin-specific proteases inhibitors in breast cancer treatment. World J. Clin. Cases 2022, 10, 11690–11701. [Google Scholar] [CrossRef]
- Clague, M.J.; Urbe, S.; Komander, D. Breaking the chains: Deubiquitylating enzyme specificity begets function. Nat. Rev. Mol. Cell Biol. 2019, 20, 338–352. [Google Scholar] [CrossRef]
- Zhou, F.; Li, Z.; Zhou, L.; Zhu, W.; Zhang, J.; Yang, W.; Xue, L.; Qin, X.; Chen, P.; Tang, R. Abstract 6201: Identification of SP-002, a highly selective USP1 inhibitor effectively inhibits HRD tumor growth and displays low hematotoxicity risk. Cancer Res. 2023, 83, 6201. [Google Scholar] [CrossRef]
- Gore, S.D.; Carducci, M.A. Modifying histones to tame cancer: Clinical development of sodium phenylbutyrate and other histone deacetylase inhibitors. Expert. Opin. Investig. Drugs 2000, 9, 2923–2934. [Google Scholar] [CrossRef] [PubMed]
- Mistry, H.; Hsieh, G.; Buhrlage, S.J.; Huang, M.; Park, E.; Cuny, G.D.; Galinsky, I.; Stone, R.M.; Gray, N.S.; D’Andrea, A.D.; et al. Small-molecule inhibitors of USP1 target ID1 degradation in leukemic cells. Mol. Cancer Ther. 2013, 12, 2651–2662. [Google Scholar] [CrossRef] [PubMed]
- Shang, K.; Zhang, L.; Yu, Y.; Xiao, H.; Gao, Y.; Yang, L.; Huang, J.; Song, H.; Han, H. Disulfide-containing polymer delivery of C527 and a Platinum(IV) prodrug selectively inhibited protein ubiquitination and tumor growth on cisplatin resistant and patient-derived liver cancer models. Mater. Today Bio 2023, 18, 100548. [Google Scholar] [CrossRef]
- Das, D.S.; Das, A.; Ray, A.; Song, Y.; Samur, M.K.; Munshi, N.C.; Chauhan, D.; Anderson, K.C. Blockade of Deubiquitylating Enzyme USP1 Inhibits DNA Repair and Triggers Apoptosis in Multiple Myeloma Cells. Clin. Cancer Res. 2017, 23, 4280–4289. [Google Scholar] [CrossRef]
- Dexheimer, T.S.; Rosenthal, A.S.; Luci, D.K.; Liang, Q.; Villamil, M.A.; Chen, J.; Sun, H.; Kerns, E.H.; Simeonov, A.; Jadhav, A.; et al. Synthesis and structure-activity relationship studies of N-benzyl-2-phenylpyrimidin-4-amine derivatives as potent USP1/UAF1 deubiquitinase inhibitors with anticancer activity against nonsmall cell lung cancer. J. Med. Chem. 2014, 57, 8099–8110. [Google Scholar] [CrossRef] [PubMed]
- Rennie, M.L.; Arkinson, C.; Chaugule, V.K.; Walden, H. Cryo-EM reveals a mechanism of USP1 inhibition through a cryptic binding site. Sci. Adv. 2022, 8, eabq6353. [Google Scholar] [CrossRef] [PubMed]
- Dexheimer, T.S.; Rosenthal, A.S.; Liang, Q.; Chen, J.; Villamil, M.A.; Kerns, E.H.; Simeonov, A.; Jadhav, A.; Zhuang, Z.; Maloney, D.J. Discovery of ML323 as a Novel Inhibitor of the USP1/UAF1 Deubiquitinase Complex. In Probe Reports from the NIH Molecular Libraries Program; National Center for Biotechnology Information: Bethesda, MD, USA, 2010. [Google Scholar]
- Chen, J.; Dexheimer, T.S.; Ai, Y.; Liang, Q.; Villamil, M.A.; Inglese, J.; Maloney, D.J.; Jadhav, A.; Simeonov, A.; Zhuang, Z. Selective and cell-active inhibitors of the USP1/ UAF1 deubiquitinase complex reverse cisplatin resistance in non-small cell lung cancer cells. Chem. Biol. 2011, 18, 1390–1400. [Google Scholar] [CrossRef]
- Li, X.Y.; Wu, J.C.; Liu, P.; Li, Z.J.; Wang, Y.; Chen, B.Y.; Hu, C.L.; Fei, M.Y.; Yu, P.C.; Jiang, Y.L.; et al. Inhibition of USP1 reverses the chemotherapy resistance through destabilization of MAX in the relapsed/refractory B-cell lymphoma. Leukemia 2023, 37, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Dakir, E.H.; Pickard, A.; Srivastava, K.; McCrudden, C.M.; Gross, S.R.; Lloyd, S.; Zhang, S.D.; Margariti, A.; Morgan, R.; Rudland, P.S.; et al. The anti-psychotic drug pimozide is a novel chemotherapeutic for breast cancer. Oncotarget 2018, 9, 34889–34910. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Xia, Y.; Gao, T.; Xu, F.; Lei, Q.; Peng, C.; Yang, Y.; Xue, Q.; Hu, X.; Wang, Q.; et al. The antipsychotic agent trifluoperazine hydrochloride suppresses triple-negative breast cancer tumor growth and brain metastasis by inducing G0/G1 arrest and apoptosis. Cell Death Dis. 2018, 9, 1006. [Google Scholar] [CrossRef] [PubMed]
- Valacchi, G.; Pecorelli, A.; Sticozzi, C.; Torricelli, C.; Muscettola, M.; Aldinucci, C.; Maioli, E. Rottlerin exhibits antiangiogenic effects in vitro. Chem. Biol. Drug Des. 2011, 77, 460–470. [Google Scholar] [CrossRef]
- Kumar, D.; Shankar, S.; Srivastava, R.K. Rottlerin-induced autophagy leads to the apoptosis in breast cancer stem cells: Molecular mechanisms. Mol. Cancer 2013, 12, 171. [Google Scholar] [CrossRef] [PubMed]
- McClurg, U.L.; Summerscales, E.E.; Harle, V.J.; Gaughan, L.; Robson, C.N. Deubiquitinating enzyme Usp12 regulates the interaction between the androgen receptor and the Akt pathway. Oncotarget 2014, 5, 7081–7092. [Google Scholar] [CrossRef] [PubMed]
- Altun, M.; Kramer, H.B.; Willems, L.I.; McDermott, J.L.; Leach, C.A.; Goldenberg, S.J.; Kumar, K.G.; Konietzny, R.; Fischer, R.; Kogan, E.; et al. Activity-based chemical proteomics accelerates inhibitor development for deubiquitylating enzymes. Chem. Biol. 2011, 18, 1401–1412. [Google Scholar] [CrossRef]
- Lin, W.C.; Chiu, Y.L.; Kuo, K.L.; Chow, P.M.; Hsu, C.H.; Liao, S.M.; Dong, J.R.; Chang, S.C.; Liu, S.H.; Liu, T.J.; et al. Anti-tumor effects of deubiquitinating enzyme inhibitor PR-619 in human chondrosarcoma through reduced cell proliferation and endoplasmic reticulum stress-related apoptosis. Am. J. Cancer Res. 2023, 13, 3055–3066. [Google Scholar] [PubMed]
- Ohayon, S.; Refua, M.; Hendler, A.; Aharoni, A.; Brik, A. Harnessing the oxidation susceptibility of deubiquitinases for inhibition with small molecules. Angew. Chem. Int. Ed. Engl. 2015, 54, 599–603. [Google Scholar] [CrossRef]
- Zhang, S.; Guo, Y.; Zhang, S.; Wang, Z.; Zhang, Y.; Zuo, S. Targeting the deubiquitinase USP2 for malignant tumor therapy (Review). Oncol. Rep. 2023, 50, 176. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.I.; Pragani, R.; Fox, J.T.; Shen, M.; Parmar, K.; Gaudiano, E.F.; Liu, L.; Tanega, C.; McGee, L.; Hall, M.D.; et al. Small Molecule Inhibition of the Ubiquitin-specific Protease USP2 Accelerates cyclin D1 Degradation and Leads to Cell Cycle Arrest in Colorectal Cancer and Mantle Cell Lymphoma Models. J. Biol. Chem. 2016, 291, 24628–24640. [Google Scholar] [CrossRef] [PubMed]
- Magiera, K.; Tomala, M.; Kubica, K.; De Cesare, V.; Trost, M.; Zieba, B.J.; Kachamakova-Trojanowska, N.; Les, M.; Dubin, G.; Holak, T.A.; et al. Lithocholic Acid Hydroxyamide Destabilizes Cyclin D1 and Induces G(0)/G(1) Arrest by Inhibiting Deubiquitinase USP2a. Cell Chem. Biol. 2017, 24, 458–470.e18. [Google Scholar] [CrossRef]
- Tomala, M.D.; Magiera-Mularz, K.; Kubica, K.; Krzanik, S.; Zieba, B.; Musielak, B.; Pustula, M.; Popowicz, G.M.; Sattler, M.; Dubin, G.; et al. Identification of small-molecule inhibitors of USP2a. Eur. J. Med. Chem. 2018, 150, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.J.; Cheng, S.C.; Tang, H.C.; Sun, C.Y.; Chou, C.Y. 6-Thioguanine is a noncompetitive and slow binding inhibitor of human deubiquitinating protease USP2. Sci. Rep. 2018, 8, 3102. [Google Scholar] [CrossRef]
- Lin, H.C.; Kuan, Y.; Chu, H.F.; Cheng, S.C.; Pan, H.C.; Chen, W.Y.; Sun, C.Y.; Lin, T.H. Disulfiram and 6-Thioguanine synergistically inhibit the enzymatic activities of USP2 and USP21. Int. J. Biol. Macromol. 2021, 176, 490–497. [Google Scholar] [CrossRef]
- Issaenko, O.A.; Amerik, A.Y. Chalcone-based small-molecule inhibitors attenuate malignant phenotype via targeting deubiquitinating enzymes. Cell Cycle 2012, 11, 1804–1817. [Google Scholar] [CrossRef]
- Coughlin, K.; Anchoori, R.; Iizuka, Y.; Meints, J.; MacNeill, L.; Vogel, R.I.; Orlowski, R.Z.; Lee, M.K.; Roden, R.B.; Bazzaro, M. Small-molecule RA-9 inhibits proteasome-associated DUBs and ovarian cancer in vitro and in vivo via exacerbating unfolded protein responses. Clin. Cancer Res. 2014, 20, 3174–3186. [Google Scholar] [CrossRef] [PubMed]
- Mirzapoiazova, T.; Pozhitkov, A.; Nam, A.; Mambetsariev, I.; Nelson, M.S.; Tan, Y.C.; Zhang, K.; Raz, D.; Singhal, S.; Nasser, M.W.; et al. Effects of selected deubiquitinating enzyme inhibitors on the proliferation and motility of lung cancer and mesothelioma cell lines. Int. J. Oncol. 2020, 57, 80–86. [Google Scholar] [CrossRef]
- Nicholson, B.; Leach, C.A.; Goldenberg, S.J.; Francis, D.M.; Kodrasov, M.P.; Tian, X.; Shanks, J.; Sterner, D.E.; Bernal, A.; Mattern, M.R.; et al. Characterization of ubiquitin and ubiquitin-like-protein isopeptidase activities. Protein Sci. 2008, 17, 1035–1043. [Google Scholar] [CrossRef]
- Vamisetti, G.B.; Meledin, R.; Gopinath, P.; Brik, A. Halogen Substituents in the Isoquinoline Scaffold Switches the Selectivity of Inhibition between USP2 and USP7. Chembiochem 2019, 20, 282–286. [Google Scholar] [CrossRef]
- Okada, K.; Ye, Y.Q.; Taniguchi, K.; Yoshida, A.; Akiyama, T.; Yoshioka, Y.; Onose, J.; Koshino, H.; Takahashi, S.; Yajima, A.; et al. Vialinin A is a ubiquitin-specific peptidase inhibitor. Bioorg. Med. Chem. Lett. 2013, 23, 4328–4331. [Google Scholar] [CrossRef]
- Xu, J.; Chen, D.; Jin, L.; Chen, Z.; Tu, Y.; Huang, X.; Xue, F.; Xu, J.; Chen, M.; Wang, X.; et al. Ubiquitously specific protease 4 inhibitor-Vialinin A attenuates inflammation and fibrosis in S100-induced hepatitis mice through Rheb/mTOR signalling. J. Cell Mol. Med. 2021, 25, 1140–1150. [Google Scholar] [CrossRef]
- Bailly, C.; Vergoten, G. Binding of Vialinin A and p-Terphenyl Derivatives to Ubiquitin-Specific Protease 4 (USP4): A Molecular Docking Study. Molecules 2022, 27, 5909. [Google Scholar] [CrossRef]
- Mann, M.K.; Zepeda-Velázquez, C.A.; González-Álvarez, H.; Dong, A.; Kiyota, T.; Aman, A.M.; Loppnau, P.; Li, Y.; Wilson, B.; Arrowsmith, C.H.; et al. Structure-Activity Relationship of USP5 Inhibitors. J. Med. Chem. 2021, 64, 15017–15036. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.N.; Zhou, Y.B.; Gao, A.H.; Cao, J.Y.; Gao, L.X.; Sheng, L.; Xu, L.; Su, M.B.; Cao, X.C.; Han, M.M.; et al. Curcusone D, a novel ubiquitin-proteasome pathway inhibitor via ROS-induced DUB inhibition, is synergistic with bortezomib against multiple myeloma cell growth. Biochim. Biophys. Acta 2014, 1840, 2004–2013. [Google Scholar] [CrossRef]
- Kapuria, V.; Peterson, L.F.; Fang, D.; Bornmann, W.G.; Talpaz, M.; Donato, N.J. Deubiquitinase inhibition by small-molecule WP1130 triggers aggresome formation and tumor cell apoptosis. Cancer Res. 2010, 70, 9265–9276. [Google Scholar] [CrossRef]
- Kim, S.; Woo, S.M.; Min, K.J.; Seo, S.U.; Lee, T.J.; Kubatka, P.; Kim, D.E.; Kwon, T.K. WP1130 Enhances TRAIL-Induced Apoptosis through USP9X-Dependent miR-708-Mediated Downregulation of c-FLIP. Cancers 2019, 11, 344. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, H.M. Recent advances in the development of ubiquitin-specific-processing protease 7 (USP7) inhibitors. Eur. J. Med. Chem. 2020, 191, 112107. [Google Scholar] [CrossRef]
- D’Arcy, P.; Wang, X.; Linder, S. Deubiquitinase inhibition as a cancer therapeutic strategy. Pharmacol. Ther. 2015, 147, 32–54. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.F.; Sun, H.; Liu, Y.; Potu, H.; Kandarpa, M.; Ermann, M.; Courtney, S.M.; Young, M.; Showalter, H.D.; Sun, D.; et al. Targeting deubiquitinase activity with a novel small-molecule inhibitor as therapy for B-cell malignancies. Blood 2015, 125, 3588–3597. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, L.; Niu, X.; Guo, Y.; Zhao, J.; Li, L.; Zhao, J. EOAI, a ubiquitin-specific peptidase 5 inhibitor, prevents non-small cell lung cancer progression by inducing DNA damage. BMC Cancer 2023, 23, 28. [Google Scholar] [CrossRef] [PubMed]
- Colland, F.; Formstecher, E.; Jacq, X.; Reverdy, C.; Planquette, C.; Conrath, S.; Trouplin, V.; Bianchi, J.; Aushev, V.N.; Camonis, J.; et al. Small-molecule inhibitor of USP7/HAUSP ubiquitin protease stabilizes and activates p53 in cells. Mol. Cancer Ther. 2009, 8, 2286–2295. [Google Scholar] [CrossRef] [PubMed]
- Pal, A.; Donato, N.J. Ubiquitin-specific proteases as therapeutic targets for the treatment of breast cancer. Breast Cancer Res. 2014, 16, 461. [Google Scholar] [CrossRef] [PubMed]
- Reverdy, C.; Conrath, S.; Lopez, R.; Planquette, C.; Atmanene, C.; Collura, V.; Harpon, J.; Battaglia, V.; Vivat, V.; Sippl, W.; et al. Discovery of specific inhibitors of human USP7/HAUSP deubiquitinating enzyme. Chem. Biol. 2012, 19, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.M.; Cheng, G.; Cheng, X.D.; Xu, Z.; Xu, B.; Zhang, W.D.; Qin, J.J. Targeting USP7-Mediated Deubiquitination of MDM2/MDMX-p53 Pathway for Cancer Therapy: Are We There Yet? Front. Cell Dev. Biol. 2020, 8, 233. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Miyazaki, M.; Kodrasov, M.P.; Rotinsulu, H.; Losung, F.; Mangindaan, R.E.; de Voogd, N.J.; Yokosawa, H.; Nicholson, B.; Tsukamoto, S. Spongiacidin C, a pyrrole alkaloid from the marine sponge Stylissa massa, functions as a USP7 inhibitor. Bioorg Med. Chem. Lett. 2013, 23, 3884–3886. [Google Scholar] [CrossRef] [PubMed]
- Kategaya, L.; Di Lello, P.; Rouge, L.; Pastor, R.; Clark, K.R.; Drummond, J.; Kleinheinz, T.; Lin, E.; Upton, J.P.; Prakash, S.; et al. USP7 small-molecule inhibitors interfere with ubiquitin binding. Nature 2017, 550, 534–538. [Google Scholar] [CrossRef]
- Kategaya, L.; Di Lello, P.; Rougé, L.; Pastor , R.; Clark, K.R.; Drummond, J. Identified Selective USP7 Inhibitors Compete with Ubiquitin Binding. Cancer Discov. 2017, 7, 1365. [Google Scholar] [CrossRef]
- Wang, S.A.; Young, M.J.; Wang, Y.C.; Chen, S.H.; Liu, C.Y.; Lo, Y.A.; Jen, H.H.; Hsu, K.C.; Hung, J.J. USP24 promotes drug resistance during cancer therapy. Cell Death Differ. 2021, 28, 2690–2707. [Google Scholar] [CrossRef]
- Lamberto, I.; Liu, X.; Seo, H.S.; Schauer, N.J.; Iacob, R.E.; Hu, W.; Das, D.; Mikhailova, T.; Weisberg, E.L.; Engen, J.R.; et al. Structure-Guided Development of a Potent and Selective Non-covalent Active-Site Inhibitor of USP7. Cell Chem. Biol. 2017, 24, 1490–1500 e1411. [Google Scholar] [CrossRef] [PubMed]
- Schauer, N.J.; Liu, X.; Magin, R.S.; Doherty, L.M.; Chan, W.C.; Ficarro, S.B.; Hu, W.; Roberts, R.M.; Iacob, R.E.; Stolte, B.; et al. Selective USP7 inhibition elicits cancer cell killing through a p53-dependent mechanism. Sci. Rep. 2020, 10, 5324. [Google Scholar] [CrossRef] [PubMed]
- Gavory, G.; O’Dowd, C.R.; Helm, M.D.; Flasz, J.; Arkoudis, E.; Dossang, A.; Hughes, C.; Cassidy, E.; McClelland, K.; Odrzywol, E.; et al. Discovery and characterization of highly potent and selective allosteric USP7 inhibitors. Nat. Chem. Biol. 2018, 14, 118–125. [Google Scholar] [CrossRef]
- O’Dowd, C.R.; Helm, M.D.; Rountree, J.S.S.; Flasz, J.T.; Arkoudis, E.; Miel, H.; Hewitt, P.R.; Jordan, L.; Barker, O.; Hughes, C.; et al. Identification and Structure-Guided Development of Pyrimidinone Based USP7 Inhibitors. ACS Med. Chem. Lett. 2018, 9, 238–243. [Google Scholar] [CrossRef]
- Turnbull, A.P.; Ioannidis, S.; Krajewski, W.W.; Pinto-Fernandez, A.; Heride, C.; Martin, A.C.L.; Tonkin, L.M.; Townsend, E.C.; Buker, S.M.; Lancia, D.R.; et al. Molecular basis of USP7 inhibition by selective small-molecule inhibitors. Nature 2017, 550, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Benavente, C. Abstract PR04: USP7 inhibitors prevent triple negative breast cancer metastasis by inducing UHRF1 protein degradation. Cancer Res. 2024, 84, PR04. [Google Scholar] [CrossRef]
- Li, X.; Kong, L.; Yang, Q.; Duan, A.; Ju, X.; Cai, B.; Chen, L.; An, T.; Li, Y. Parthenolide inhibits ubiquitin-specific peptidase 7 (USP7), Wnt signaling, and colorectal cancer cell growth. J. Biol. Chem. 2020, 295, 3576–3589. [Google Scholar] [CrossRef]
- Cheng, Y.J.; Zhuang, Z.; Miao, Y.L.; Song, S.S.; Bao, X.B.; Yang, C.H.; He, J.X. Identification of YCH2823 as a novel USP7 inhibitor for cancer therapy. Biochem. Pharmacol. 2024, 222, 116071. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, E.L.; Schauer, N.J.; Yang, J.; Lamberto, I.; Doherty, L.; Bhatt, S.; Nonami, A.; Meng, C.; Letai, A.; Wright, R.; et al. Inhibition of USP10 induces degradation of oncogenic FLT3. Nat. Chem. Biol. 2017, 13, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Javaid, S.; Zadi, S.; Awais, M.; Wahab, A.T.; Zafar, H.; Maslennikov, I.; Choudhary, M.I. Identification of new leads against ubiquitin specific protease-7 (USP7): A step towards the potential treatment of cancers. RSC Adv. 2024, 14, 33080–33093. [Google Scholar] [CrossRef]
- Lee, G.; Oh, T.I.; Um, K.B.; Yoon, H.; Son, J.; Kim, B.M.; Kim, H.I.; Kim, H.; Kim, Y.J.; Lee, C.S.; et al. Small-molecule inhibitors of USP7 induce apoptosis through oxidative and endoplasmic reticulum stress in cancer cells. Biochem. Biophys. Res. Commun. 2016, 470, 181–186. [Google Scholar] [CrossRef]
- Fan, Y.H.; Cheng, J.; Vasudevan, S.A.; Dou, J.; Zhang, H.; Patel, R.H.; Ma, I.T.; Rojas, Y.; Zhao, Y.; Yu, Y.; et al. USP7 inhibitor P22077 inhibits neuroblastoma growth via inducing p53-mediated apoptosis. Cell Death Dis. 2013, 4, e867. [Google Scholar] [CrossRef]
- Cartel, M.; Mouchel, P.L.; Gotanegre, M.; David, L.; Bertoli, S.; Mansat-De Mas, V.; Besson, A.; Sarry, J.E.; Manenti, S.; Didier, C. Inhibition of ubiquitin-specific protease 7 sensitizes acute myeloid leukemia to chemotherapy. Leukemia 2021, 35, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Pozhidaeva, A.; Valles, G.; Wang, F.; Wu, J.; Sterner, D.E.; Nguyen, P.; Weinstock, J.; Kumar, K.G.S.; Kanyo, J.; Wright, D.; et al. USP7-Specific Inhibitors Target and Modify the Enzyme’s Active Site via Distinct Chemical Mechanisms. Cell Chem. Biol. 2017, 24, 1501–1512. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, D.; Tian, Z.; Nicholson, B.; Kumar, K.G.; Zhou, B.; Carrasco, R.; McDermott, J.L.; Leach, C.A.; Fulcinniti, M.; Kodrasov, M.P.; et al. A small molecule inhibitor of ubiquitin-specific protease-7 induces apoptosis in multiple myeloma cells and overcomes bortezomib resistance. Cancer Cell 2012, 22, 345–358. [Google Scholar] [CrossRef]
- An, T.; Gong, Y.; Li, X.; Kong, L.; Ma, P.; Gong, L.; Zhu, H.; Yu, C.; Liu, J.; Zhou, H.; et al. USP7 inhibitor P5091 inhibits Wnt signaling and colorectal tumor growth. Biochem. Pharmacol. 2017, 131, 29–39. [Google Scholar] [CrossRef]
- Granieri, L.; Marocchi, F.; Melixetian, M.; Mohammadi, N.; Nicoli, P.; Cuomo, A.; Bonaldi, T.; Confalonieri, S.; Pisati, F.; Giardina, G.; et al. Targeting the USP7/RRM2 axis drives senescence and sensitizes melanoma cells to HDAC/LSD1 inhibitors. Cell Rep. 2022, 40, 111396. [Google Scholar] [CrossRef]
- Weinstock, J.; Wu, J.; Cao, P.; Kingsbury, W.D.; McDermott, J.L.; Kodrasov, M.P.; McKelvey, D.M.; Suresh Kumar, K.G.; Goldenberg, S.J.; Mattern, M.R.; et al. Selective Dual Inhibitors of the Cancer-Related Deubiquitylating Proteases USP7 and USP47. ACS Med. Chem. Lett. 2012, 3, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; Vallese, S.; Peretto, I.; Jacq, X.; Rain, J.C.; Colland, F.; Guedat, P. Synthesis and biological evaluation of 9-oxo-9H-indeno[1,2-b]pyrazine-2,3-dicarbonitrile analogues as potential inhibitors of deubiquitinating enzymes. ChemMedChem 2010, 5, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Clancy, A.; Heride, C.; Pinto-Fernandez, A.; Elcocks, H.; Kallinos, A.; Kayser-Bricker, K.J.; Wang, W.; Smith, V.; Davis, S.; Fessler, S.; et al. The deubiquitylase USP9X controls ribosomal stalling. J. Cell Biol. 2021, 220, e202004211. [Google Scholar] [CrossRef]
- Yu, M.; Fang, Z.X.; Wang, W.W.; Zhang, Y.; Bu, Z.L.; Liu, M.; Xiao, X.H.; Zhang, Z.L.; Zhang, X.M.; Cao, Y.; et al. Wu-5, a novel USP10 inhibitor, enhances crenolanib-induced FLT3-ITD-positive AML cell death via inhibiting FLT3 and AMPK pathways. Acta Pharmacol. Sin. 2021, 42, 604–612. [Google Scholar] [CrossRef]
- Yang, J.; Meng, C.; Weisberg, E.; Case, A.; Lamberto, I.; Magin, R.S.; Adamia, S.; Wang, J.; Gray, N.; Liu, S.; et al. Inhibition of the deubiquitinase USP10 induces degradation of SYK. Br. J. Cancer 2020, 122, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xia, H.; Kim, M.; Xu, L.; Li, Y.; Zhang, L.; Cai, Y.; Norberg, H.V.; Zhang, T.; Furuya, T.; et al. Beclin1 controls the levels of p53 by regulating the deubiquitination activity of USP10 and USP13. Cell 2011, 147, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Ndubaku, C.; Tsui, V. Inhibiting the deubiquitinating enzymes (DUBs). J. Med. Chem. 2015, 58, 1581–1595. [Google Scholar] [CrossRef] [PubMed]
- Kunimasa, K.; Ikeda-Ishikawa, C.; Tani, Y.; Tsukahara, S.; Sakurai, J.; Okamoto, Y.; Koido, M.; Dan, S.; Tomida, A. Spautin-1 inhibits mitochondrial complex I and leads to suppression of the unfolded protein response and cell survival during glucose starvation. Sci. Rep. 2022, 12, 11533. [Google Scholar] [CrossRef]
- Burkhart, R.A.; Peng, Y.; Norris, Z.A.; Tholey, R.M.; Talbott, V.A.; Liang, Q.; Ai, Y.; Miller, K.; Lal, S.; Cozzitorto, J.A.; et al. Mitoxantrone targets human ubiquitin-specific peptidase 11 (USP11) and is a potent inhibitor of pancreatic cancer cell survival. Mol. Cancer Res. 2013, 11, 901–911. [Google Scholar] [CrossRef]
- McClurg, U.L.; Azizyan, M.; Dransfield, D.T.; Namdev, N.; Chit, N.; Nakjang, S.; Robson, C.N. The novel anti-androgen candidate galeterone targets deubiquitinating enzymes, USP12 and USP46, to control prostate cancer growth and survival. Oncotarget 2018, 9, 24992–25007. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Lee, M.J.; Park, S.; Oh, D.C.; Elsasser, S.; Chen, P.C.; Gartner, C.; Dimova, N.; Hanna, J.; Gygi, S.P.; et al. Enhancement of proteasome activity by a small-molecule inhibitor of USP14. Nature 2010, 467, 179–184. [Google Scholar] [CrossRef]
- Xu, L.; Wang, J.; Yuan, X.; Yang, S.; Xu, X.; Li, K.; He, Y.; Wei, L.; Zhang, J.; Tian, Y. IU1 suppresses proliferation of cervical cancer cells through MDM2 degradation. Int. J. Biol. Sci. 2020, 16, 2951–2963. [Google Scholar] [CrossRef] [PubMed]
- Boselli, M.; Lee, B.H.; Robert, J.; Prado, M.A.; Min, S.W.; Cheng, C.; Silva, M.C.; Seong, C.; Elsasser, S.; Hatle, K.M.; et al. An inhibitor of the proteasomal deubiquitinating enzyme USP14 induces tau elimination in cultured neurons. J. Biol. Chem. 2017, 292, 19209–19225. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, Y.; Ding, S.; Li, J.; Song, N.; Ren, Y.; Hong, D.; Wu, C.; Li, B.; Wang, F.; et al. Small molecule inhibitors reveal allosteric regulation of USP14 via steric blockade. Cell Res. 2018, 28, 1186–1194. [Google Scholar] [CrossRef]
- Li, P.; Zhu, X.; Qu, H.; Han, Z.; Yao, X.; Wei, Y.; Li, B.; Chen, H. Synergistic Effect of Ubiquitin-Specific Protease 14 and Poly(ADP-Ribose) Glycohydrolase Co-Inhibition in BRCA1-Mutant, Poly(ADP-Ribose) Polymerase Inhibitor-Resistant Triple-Negative Breast Cancer Cells. Onco Targets Ther. 2024, 17, 741–753. [Google Scholar] [CrossRef]
- Chen, X.; Yang, Q.; Xiao, L.; Tang, D.; Dou, Q.P.; Liu, J. Metal-based proteasomal deubiquitinase inhibitors as potential anticancer agents. Cancer Metastasis Rev. 2017, 36, 655–668. [Google Scholar] [CrossRef]
- Liu, N.; Li, X.; Huang, H.; Zhao, C.; Liao, S.; Yang, C.; Liu, S.; Song, W.; Lu, X.; Lan, X.; et al. Clinically used antirheumatic agent auranofin is a proteasomal deubiquitinase inhibitor and inhibits tumor growth. Oncotarget 2014, 5, 5453–5471. [Google Scholar] [CrossRef]
- Liu, N.; Liu, C.; Li, X.; Liao, S.; Song, W.; Yang, C.; Zhao, C.; Huang, H.; Guan, L.; Zhang, P.; et al. A novel proteasome inhibitor suppresses tumor growth via targeting both 19S proteasome deubiquitinases and 20S proteolytic peptidases. Sci. Rep. 2014, 4, 5240. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Chen, X.; Yang, C.; Zang, D.; Lan, X.; Liao, S.; Zhang, P.; Wu, J.; Li, X.; Liu, N.; et al. Repurposing an antidandruff agent to treating cancer: Zinc pyrithione inhibits tumor growth via targeting proteasome-associated deubiquitinases. Oncotarget 2017, 8, 13942–13956. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Chen, X.; Zang, D.; Lan, X.; Liao, S.; Yang, C.; Zhang, P.; Wu, J.; Li, X.; Liu, N.; et al. Platinum-containing compound platinum pyrithione is stronger and safer than cisplatin in cancer therapy. Biochem. Pharmacol. 2016, 116, 22–38. [Google Scholar] [CrossRef]
- Zhao, C.; Chen, X.; Zang, D.; Lan, X.; Liao, S.; Yang, C.; Zhang, P.; Wu, J.; Li, X.; Liu, N.; et al. A novel nickel complex works as a proteasomal deubiquitinase inhibitor for cancer therapy. Oncogene 2016, 35, 5916–5927. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Ning, S.; Yu, B.; Wang, Y. USP14: Structure, Function, and Target Inhibition. Front. Pharmacol. 2021, 12, 801328. [Google Scholar] [CrossRef]
- Wang, X.; Stafford, W.; Mazurkiewicz, M.; Fryknas, M.; Brjnic, S.; Zhang, X.; Gullbo, J.; Larsson, R.; Arner, E.S.; D’Arcy, P.; et al. The 19S Deubiquitinase inhibitor b-AP15 is enriched in cells and elicits rapid commitment to cell death. Mol. Pharmacol. 2014, 85, 932–945. [Google Scholar] [CrossRef]
- Ward, J.A.; Pinto-Fernandez, A.; Cornelissen, L.; Bonham, S.; Diaz-Saez, L.; Riant, O.; Huber, K.V.M.; Kessler, B.M.; Feron, O.; Tate, E.W. Re-Evaluating the Mechanism of Action of alpha, beta-Unsaturated Carbonyl DUB Inhibitors b-AP15 and VLX1570: A Paradigmatic Example of Unspecific Protein Cross-linking with Michael Acceptor Motif-Containing Drugs. J. Med. Chem. 2020, 63, 3756–3762. [Google Scholar] [CrossRef]
- Cai, J.; Xia, X.; Liao, Y.; Liu, N.; Guo, Z.; Chen, J.; Yang, L.; Long, H.; Yang, Q.; Zhang, X.; et al. A novel deubiquitinase inhibitor b-AP15 triggers apoptosis in both androgen receptor-dependent and -independent prostate cancers. Oncotarget 2017, 8, 63232–63246. [Google Scholar] [CrossRef]
- Gubat, J.; Selvaraju, K.; Sjostrand, L.; Kumar Singh, D.; Turkina, M.V.; Schmierer, B.; Sabatier, P.; Zubarev, R.A.; Linder, S.; D’Arcy, P. Comprehensive Target Screening and Cellular Profiling of the Cancer-Active Compound b-AP15 Indicate Abrogation of Protein Homeostasis and Organelle Dysfunction as the Primary Mechanism of Action. Front. Oncol. 2022, 12, 852980. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Gao, Q.; Jia, Y.; Wei, J.; Chaudhuri, S.M.; Wang, S.; Tang, A.; Mani, N.L.; Iyer, R.; Cheng, Y.; et al. Ubiquitin-specific peptidase 22 controls integrin-dependent cancer cell stemness and metastasis. iScience 2024, 27, 110592. [Google Scholar] [CrossRef]
- Prokakis, E.; Dyas, A.; Grun, R.; Fritzsche, S.; Bedi, U.; Kazerouni, Z.B.; Kosinsky, R.L.; Johnsen, S.A.; Wegwitz, F. USP22 promotes HER2-driven mammary carcinoma aggressiveness by suppressing the unfolded protein response. Oncogene 2021, 40, 4004–4018. [Google Scholar] [CrossRef] [PubMed]
- Montauti, E.; Weinberg, S.E.; Chu, P.; Chaudhuri, S.; Mani, N.L.; Iyer, R.; Zhou, Y.; Zhang, Y.; Liu, C.; Xin, C.; et al. A deubiquitination module essential for T(reg) fitness in the tumor microenvironment. Sci. Adv. 2022, 8, eabo4116. [Google Scholar] [CrossRef] [PubMed]
- Prokakis, E.; Bamahmoud, H.; Jansari, S.; Fritsche, L.; Dietz, A.; Boshnakovska, A.; Rehling, P.; Johnsen, S.A.; Gallwas, J.; Wegwitz, F. USP22 supports the aggressive behavior of basal-like breast cancer by stimulating cellular respiration. Cell Commun. Signal 2024, 22, 120. [Google Scholar] [CrossRef]
- Wang, S.A.; Wu, Y.C.; Yang, F.M.; Hsu, F.L.; Zhang, K.; Hung, J.J. NCI677397 targeting USP24-mediated induction of lipid peroxidation induces ferroptosis in drug-resistant cancer cells. Mol. Oncol. 2024, 18, 2255–2276. [Google Scholar] [CrossRef] [PubMed]
- Young, M.J.; Wang, S.A.; Chen, Y.C.; Liu, C.Y.; Hsu, K.C.; Tang, S.W.; Tseng, Y.L.; Wang, Y.C.; Lin, S.M.; Hung, J.J. USP24-i-101 targeting of USP24 activates autophagy to inhibit drug resistance acquired during cancer therapy. Cell Death Differ. 2024, 31, 574–591. [Google Scholar] [CrossRef] [PubMed]
- Wrigley, J.D.; Gavory, G.; Simpson, I.; Preston, M.; Plant, H.; Bradley, J.; Goeppert, A.U.; Rozycka, E.; Davies, G.; Walsh, J.; et al. Identification and Characterization of Dual Inhibitors of the USP25/28 Deubiquitinating Enzyme Subfamily. ACS Chem. Biol. 2017, 12, 3113–3125. [Google Scholar] [CrossRef] [PubMed]
- Patzke, J.V.; Sauer, F.; Nair, R.K.; Endres, E.; Proschak, E.; Hernandez-Olmos, V.; Sotriffer, C.; Kisker, C. Structural basis for the bi-specificity of USP25 and USP28 inhibitors. EMBO Rep. 2024, 25, 2950–2973. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Xu, Z.; Huang, Y.; Wang, H.; Zhu, X.; Zhang, W.; Song, W.; Gao, T.; Liu, T.; Wang, M.; et al. Structure-based discovery of potent USP28 inhibitors derived from Vismodegib. Eur. J. Med. Chem. 2023, 254, 115369. [Google Scholar] [CrossRef]
- Aditya, S.; Rattan, A. Vismodegib: A smoothened inhibitor for the treatment of advanced basal cell carcinoma. Indian. Dermatol. Online J. 2013, 4, 365–368. [Google Scholar] [CrossRef]
- Wang, H.; Meng, Q.; Ding, Y.; Xiong, M.; Zhu, M.; Yang, Y.; Su, H.; Gu, L.; Xu, Y.; Shi, L.; et al. USP28 and USP25 are downregulated by Vismodegib in vitro and in colorectal cancer cell lines. FEBS J. 2021, 288, 1325–1342. [Google Scholar] [CrossRef]
- Kluge, A.F.; Lagu, B.R.; Maiti, P.; Jaleel, M.; Webb, M.; Malhotra, J.; Mallat, A.; Srinivas, P.A.; Thompson, J.E. Novel highly selective inhibitors of ubiquitin specific protease 30 (USP30) accelerate mitophagy. Bioorg. Med. Chem. Lett. 2018, 28, 2655–2659. [Google Scholar] [CrossRef]
- Zhang, X.; Han, Y.; Liu, S.; Guo, B.; Xu, S.; He, Y.; Liu, L. MF-094 nanodelivery inhibits oral squamous cell carcinoma by targeting USP30. Cell Mol. Biol. Lett. 2022, 27, 107. [Google Scholar] [CrossRef]
- Li, X.; Wang, T.; Tao, Y.; Wang, X.; Li, L.; Liu, J. MF-094, a potent and selective USP30 inhibitor, accelerates diabetic wound healing by inhibiting the NLRP3 inflammasome. Exp. Cell Res. 2022, 410, 112967. [Google Scholar] [CrossRef]
- Wang, W.; Liu, H.; Wang, S.; Hao, X.; Li, L. A diterpenoid derivative 15-oxospiramilactone inhibits Wnt/beta-catenin signaling and colon cancer cell tumorigenesis. Cell Res. 2011, 21, 730–740. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Chen, Z.; Liu, H.; Yan, C.; Chen, M.; Feng, D.; Yan, C.; Wu, H.; Du, L.; Wang, Y.; et al. A small natural molecule promotes mitochondrial fusion through inhibition of the deubiquitinase USP30. Cell Res. 2014, 24, 482–496. [Google Scholar] [CrossRef]
- Waters, C.S.; Angenent, S.B.; Altschuler, S.J.; Wu, L.F. A PINK1 input threshold arises from positive feedback in the PINK1/Parkin mitophagy decision circuit. Cell Rep. 2023, 42, 113260. [Google Scholar] [CrossRef]
- Rusilowicz-Jones, E.V.; Barone, F.G.; Lopes, F.M.; Stephen, E.; Mortiboys, H.; Urbe, S.; Clague, M.J. Benchmarking a highly selective USP30 inhibitor for enhancement of mitophagy and pexophagy. Life Sci. Alliance 2022, 5, e202101287. [Google Scholar] [CrossRef]
- Li, M.; Wang, X.; Qiu, Y.; Zhang, Y.; Pan, X.; Tang, N.; Chen, T.; Ruan, B.; Shao, S.; He, L.; et al. Safety reassessment of cinobufotalin injection: New findings into cardiotoxicity. Toxicol. Res. 2020, 9, 390–398. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, Y.; Shin, H.; Hwang, S.; Park, J.; Song, E.J. Ubiquitin-proteasome system as a target for anticancer treatment-an update. Arch. Pharm. Res. 2023, 46, 573–597. [Google Scholar] [CrossRef]
- Gao, H.; Xi, Z.; Dai, J.; Xue, J.; Guan, X.; Zhao, L.; Chen, Z.; Xing, F. Drug resistance mechanisms and treatment strategies mediated by Ubiquitin-Specific Proteases (USPs) in cancers: New directions and therapeutic options. Mol. Cancer 2024, 23, 88. [Google Scholar] [CrossRef]
- Gao, H.; Yin, J.; Ji, C.; Yu, X.; Xue, J.; Guan, X.; Zhang, S.; Liu, X.; Xing, F. Targeting ubiquitin specific proteases (USPs) in cancer immunotherapy: From basic research to preclinical application. J. Exp. Clin. Cancer Res. 2023, 42, 225. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, P.; Zhan, X.; Zhang, Y.; Sheng, J.; He, S.; Chen, Y.; Nie, D.; You, X.; Mai, H.; et al. USP1-regulated reciprocal differentiation of Th17 cells and Treg cells by deubiquitinating and stabilizing TAZ. Cell Mol. Immunol. 2023, 20, 252–263. [Google Scholar] [CrossRef] [PubMed]
- van Loosdregt, J.; Fleskens, V.; Fu, J.; Brenkman, A.B.; Bekker, C.P.; Pals, C.E.; Meerding, J.; Berkers, C.R.; Barbi, J.; Grone, A.; et al. Stabilization of the transcription factor Foxp3 by the deubiquitinase USP7 increases Treg-cell-suppressive capacity. Immunity 2013, 39, 259–271. [Google Scholar] [CrossRef]
- Wang, L.; Kumar, S.; Dahiya, S.; Wang, F.; Wu, J.; Newick, K.; Han, R.; Samanta, A.; Beier, U.H.; Akimova, T.; et al. Ubiquitin-specific Protease-7 Inhibition Impairs Tip60-dependent Foxp3+ T-regulatory Cell Function and Promotes Antitumor Immunity. EBioMedicine 2016, 13, 99–112. [Google Scholar] [CrossRef]
- Wang, Z.; Kang, W.; Li, O.; Qi, F.; Wang, J.; You, Y.; He, P.; Suo, Z.; Zheng, Y.; Liu, H.M. Abrogation of USP7 is an alternative strategy to downregulate PD-L1 and sensitize gastric cancer cells to T cells killing. Acta Pharm. Sin. B 2021, 11, 694–707. [Google Scholar] [CrossRef]
- Selvaraju, K.; Mazurkiewicz, M.; Wang, X.; Gullbo, J.; Linder, S.; D’Arcy, P. Inhibition of proteasome deubiquitinase activity: A strategy to overcome resistance to conventional proteasome inhibitors? Drug Resist. Updat. 2015, 21–22, 20–29. [Google Scholar] [CrossRef]
- Li, J.; Yuan, S.; Norgard, R.J.; Yan, F.; Yamazoe, T.; Blanco, A.; Stanger, B.Z. Tumor Cell-Intrinsic USP22 Suppresses Antitumor Immunity in Pancreatic Cancer. Cancer Immunol. Res. 2020, 8, 282–291. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, Q.; Lou, Y.; Wang, J.; Zhao, X.; Wang, L.; Zhang, X.; Li, S.; Zhao, Y.; Chen, Q.; et al. USP22 Deubiquitinates CD274 to Suppress Anticancer Immunity. Cancer Immunol. Res. 2019, 7, 1580–1590. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Yu, H.; Yao, J.; Li, J.; Li, Z.; Jiang, Z. ncRNA-mediated overexpression of ubiquitin-specific proteinase 13 contributes to the progression of prostate cancer via modulating AR signaling, DNA damage repair and immune infiltration. BMC Cancer 2022, 22, 1350. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Wu, X.; Jian, Y.; Wang, J.; Huang, C.; Mo, S.; Li, Y.; Li, F.; Zhang, C.; Zhang, D.; et al. USP14 promotes tryptophan metabolism and immune suppression by stabilizing IDO1 in colorectal cancer. Nat. Commun. 2022, 13, 5644. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, A.; Murakami, K.; Elia, A.; Shibahara, Y.; Done, S.J.; Wood, S.A.; Donato, N.J.; Ohashi, P.S.; Reedijk, M. Therapeutic inhibition of USP9x-mediated Notch signaling in triple-negative breast cancer. Proc. Natl. Acad. Sci. USA 2021, 118, e2101592118. [Google Scholar] [CrossRef]
- Lim, K.S.; Li, H.; Roberts, E.A.; Gaudiano, E.F.; Clairmont, C.; Sambel, L.A.; Ponnienselvan, K.; Liu, J.C.; Yang, C.; Kozono, D.; et al. USP1 Is Required for Replication Fork Protection in BRCA1-Deficient Tumors. Mol. Cell 2018, 72, 925–941 e924. [Google Scholar] [CrossRef] [PubMed]
- Rennie, M.L.; Gundogdu, M.; Arkinson, C.; Liness, S.; Frame, S.; Walden, H. Structural and Biochemical Insights into the Mechanism of Action of the Clinical USP1 Inhibitor, KSQ-4279. J. Med. Chem. 2024, 67, 15557–15568. [Google Scholar] [CrossRef] [PubMed]
- Cadzow, L.; Brenneman, J.; Tobin, E.; Sullivan, P.; Nayak, S.; Ali, J.A.; Shenker, S.; Griffith, J.; McGuire, M.; Grasberger, P.; et al. The USP1 Inhibitor KSQ-4279 Overcomes PARP Inhibitor Resistance in Homologous Recombination-Deficient Tumors. Cancer Res. 2024, 84, 3419–3434. [Google Scholar] [CrossRef]
- Li, H.; Liu, B.J.; Xu, J.; Song, S.S.; Ba, R.; Zhang, J.; Huan, X.J.; Wang, D.; Miao, Z.H.; Liu, T.; et al. Design, synthesis, and biological evaluation of pyrido[2,3-d]pyrimidin-7(8H)-one derivatives as potent USP1 inhibitors. Eur. J. Med. Chem. 2024, 275, 116568. [Google Scholar] [CrossRef]
- Wang, X.; Mazurkiewicz, M.; Hillert, E.K.; Olofsson, M.H.; Pierrou, S.; Hillertz, P.; Gullbo, J.; Selvaraju, K.; Paulus, A.; Akhtar, S.; et al. The proteasome deubiquitinase inhibitor VLX1570 shows selectivity for ubiquitin-specific protease-14 and induces apoptosis of multiple myeloma cells. Sci. Rep. 2016, 6, 26979. [Google Scholar] [CrossRef]
- Rowinsky, E.K.; Paner, A.; Berdeja, J.G.; Paba-Prada, C.; Venugopal, P.; Porkka, K.; Gullbo, J.; Linder, S.; Loskog, A.; Richardson, P.G.; et al. Phase 1 study of the protein deubiquitinase inhibitor VLX1570 in patients with relapsed and/or refractory multiple myeloma. Investig. New Drugs 2020, 38, 1448–1453. [Google Scholar] [CrossRef]
- Simoneau, A.; Pratt, C.; Comer, G.; Wu, H.-J.; Choi, A.; Khendu, T.; Meier, S.; Yu, Y.; Liu, S.; Zhang, W.; et al. Abstract B054: TNG348, a selective USP1 inhibitor, shows strong preclinical combination activity with PARP inhibitors and other agents targeting DNA repair. Mol. Cancer Ther. 2023, 22, B054. [Google Scholar] [CrossRef]
- Simoneau, A.; Wu, H.-J.; Bandi, M.; Lazarides, K.; Sun, S.; Liu, S.; Meier, S.; Choi, A.; Zhang, H.; Shen, B.; et al. Abstract 4968: Characterization of the clinical development candidate TNG348 as a potent and selective inhibitor of USP1 for the treatment of BRCA1/2mut cancers. Cancer Res. 2023, 83, 4968. [Google Scholar] [CrossRef]
- Phase 1/2, Multi-Center, Open-Label Study to Evaluate the Safety, Tolerability, and Preliminary Antitumor Activity of TNG348 Single Agent and in Combination With a PARP Inhibitor in Patients With BRCA 1/2 Mutant or Other HRD+ Solid Tumors. 2024. Available online: https://www.dana-farber.org/clinical-trials/23-643 (accessed on 13 August 2024).
- A Phase 1 Study of RO7623066 Alone and in Combination in Patients With Advanced Solid Tumors. 2022. Available online: https://www.cancer.gov/research/participate/clinical-trials-search/v?id=NCI-2022-01620 (accessed on 4 February 2025).
- Yap, T.A.; Lakhani, N.J.; Patnaik, A.; Lee, E.K.; Gutierrez, M.; Moore, K.N.; Carneiro, B.A.; Hays, J.L.; Huang, M.; LoRusso, P.; et al. First-in-human phase I trial of the oral first-in-class ubiquitin specific peptidase 1 (USP1) inhibitor KSQ-4279 (KSQi), given as single agent (SA) and in combination with olaparib (OLA) or carboplatin (CARBO) in patients (pts) with advanced solid tumors, enriched for deleterious homologous recombination repair (HRR) mutations. J. Clin. Oncol. 2024, 42, 3005. [Google Scholar] [CrossRef]
- Li, Y.; Wu, J.; Liu, J.; Qin, L.; Cai, X.; Qiao, J.; Wang, L.; Rao, S.; Ren, F.; Zhavoronkov, A. Abstract 502: ISM3091, a novel selective USP1 inhibitor as a targeted anticancer therapy. Cancer Res. 2023, 83, 502. [Google Scholar] [CrossRef]
- An Open-Label, Multicenter Study to Investigate the Safety, Tolerability, Pharmacokinetics, and Preliminary Efficacy of XL309 (ISM3091) as Single-Agent and Combination Therapy in Patients With Advanced Solid Tumors. 2025. Available online: https://clinicaltrials.gov/study/NCT05932862 (accessed on 24 January 2025).
- A Phase I First-in-human, Open-label Trial to Investigate the Safety, Pharmacokinetics and Antitumor Activity of SIM0501 as Monotherapy and in Combination in Participants With Advanced Solid Tumors. 2024. Available online: https://www.medifind.com/articles/clinical-trial/507724029 (accessed on 4 February 2025).
- Yang, S.; Li, Z.; Zhou, L.; Zhou, F.; Qin, X.; Xue, L.; Xu, P.; Tang, J.; Wang, W.; Tang, R. Abstract LB271: Synergistic anti-tumor efficacy in olaparib-sensitive and -resistant models via simultaneously inhibition of USP1 and PARP. Cancer Res. 2024, 84, LB271. [Google Scholar] [CrossRef]
- Xu, J.; Li, Y.; Wang, J.; Tang, P.; Wang, J.; Zhang, C.; Yan, P. Abstract 7146: HSK39775: A USP1 inhibitor for the treatment of cancers with homologous recombination deficiencies. Cancer Res. 2024, 84, 7146. [Google Scholar] [CrossRef]
- A Phase I/II Study to Assess the Safety, Tolerability, PK/PD, and Preliminary Efficacy of HSK39775 Tablet Monotherapy in Participants With Advanced Solid Malignancies. 2024. Available online: https://adisinsight.springer.com/trials/700372061 (accessed on 22 March 2024).
- VLX1570 and Low-Dose Dexamethasone in Relapsed or Relapsed and Refractory Multiple Myeloma: A Clinical and Correlative Phase 1/2 Study. 2018. Available online: https://clinicaltrials.gov/study/NCT02372240 (accessed on 11 May 2018).
- Kharel, P.; Uprety, D.; Chandra, A.B.; Hu, Y.; Belur, A.A.; Dhakal, A. Bortezomib-Induced Pulmonary Toxicity: A Case Report and Review of Literature. Case Rep. Med. 2018, 2018, 2913124. [Google Scholar] [CrossRef]

| Pathway | Role of USPs | Cancers | References |
|---|---|---|---|
| NF-κB pathway | Nuclear factor-kappa B (NF-κB) is involved in the regulation of several key biological functions, including cell survival. USPs regulate the NF-κB pathway by modulating the deubiquitination of key signaling proteins. USP21 plays a key role in the downregulation of TNF- α induced NF-κB signaling through deubiquitination of receptor-interacting protein 1 (RIP1), while USP4 promotes TNF-α mediated NF-κB activation by deubiquitinating TGF- β-activated kinase 1, highlighting the diverse roles USPs can play in regulating NF-κB signaling. Downregulation of NF-κB promotes cell death and reduces cell survival. | Breast cancer, prostate cancer, lung cancer, colorectal cancer, and ovarian cancer | [25,26,27] |
| Wnt/β-catenin pathway | The Wnt/β-catenin pathway is a crucial signaling mechanism that regulates various biological processes, including cell proliferation, differentiation, and survival. β-catenin is a key signal transducer in this pathway and is positively regulated by USP4. The C-terminal catalytic domain of USP4 is responsible for binding and nuclear transport of β-catenin leading to increased transcription of oncogenes. USP4 knockdown in a cell line of colon cancer has also been found to reduce invasion and migration, while overexpression has been demonstrated to enhance β-catenin-regulated transcription. | Colorectal cancer, gastric cancer, breast cancer, lung cancer, pancreatic cancer, and melanoma | [28,29] |
| JAK/STAT pathway | USPs regulate the JAK/STAT pathway by modulating the stability and degradation of key signaling components, such as STAT3 which plays a crucial role in transcription. Aberrant activation of STAT3 can enhance cancer cell proliferation, increase cell survival by up-regulating anti-apoptotic genes, and may contribute to drug resistance as well as immune evasion. USP5 plays a significant role in the progression and metastasis of pancreatic cancer by stabilizing STAT3. | Breast cancer, lung cancer, colorectal cancer, gastric cancer, leukemia, and bladder cancer | [30,31,32] |
| p53 signaling pathway | Several USPs regulate the p53 signaling pathway. Importantly, USP7 has been demonstrated to regulate p53 by stabilizing MDM2, an E3 ubiquitin ligase that typically promotes p53 degradation. USP7 has been proposed to be a potential therapeutic target given the contribution it has to cancer pathogenesis through the downregulation of p53. This in turn allows damaged cells to evade apoptosis and proliferate abnormally. | Breast cancer, colorectal cancer, ovarian cancer, lung cancer, and head and neck cancer | [33,34] |
| c-MYC | c-MYC is a well-established oncogene that plays a critical role in the progression of various cancers. This pathway is significantly influenced by various USPs, such as USP16 and USP36, which interact directly with c-MYC by deubiquitinating and stabilizing it, promoting chemoresistance. USP1 has been demonstrated to promote c-MYC pathway activation, leading to increased transcription of MYC target genes. USP7 indirectly affects MYC levels by stabilizing MDM2 and MDMX which regulate p53 that functions as a MYC antagonist. | Colorectal cancer, breast cancer, non-small cell lung cancer, pancreatic ductal adenocarcinoma, glioblastoma, and ovarian cancer | [35,36,37] |
| TGF-beta pathway | TGF-beta signaling has been shown to drive epithelial-mesenchymal transition enhancing tumor migration and invasion through pathways regulated by USPs. USP22 is upregulated in many types of cancers. In epithelial ovarian cancer, higher USP22 levels are associated with higher stage and worse clinical outcomes. USP22 facilitates cell proliferation by inducing G1 cell cycle arrest through interaction with oncogenic TGF-β1. | Breast cancer, colorectal cancer, liver cancer, non-small cell lung cancer, pancreatic cancer, and epithelial ovarian cancer | [38,39,40] |
| DNA repair pathways | USP21 has shown positive regulatory effects on BRCA2 which plays a critical role in homologous recombination. USP1 also plays a critical role in DNA damage repair by modulating the ubiquitination of key regulators of DNA repair such as PCNA and FANCD2. | Breast cancer, non-small cell lung cancer, colorectal cancer, gastric cancer, ovarian cancer, and hepatocellular carcinoma | [3,41,42,43] |
| Apoptotic pathways | Apoptotic signaling pathways are regulated by USPs. BCL-2 family proteins regulate apoptosis by controlling the release of apoptogenic factors, with both pro-apoptotic and anti-apoptotic proteins regulated by this group of proteins. Overexpression of USP9x in certain cancers leads to the overexpression of Mcl-1, an anti-apoptotic protein, by way of deubiquitination. This has been shown to lead to increased resistance to radiation therapy. | Breast cancer, lung cancer, colorectal cancer, ovarian cancer, and hepatocellular carcinoma | [44,45,46] |
| Cell cycle regulation pathways | The cell cycle is characterized by stages G1, S, G2, and M. Cyclins, cyclin-dependent kinases (CDKs), cyclin-dependent kinase inhibitors (CDKIs), and aurora kinases function to regulate the cellular transition among these stages. USPs regulate cell cycle progression by stabilizing regulatory proteins such as CDKs. USP2, USP5, USP13, USP20, and USP22 have been found to promote the stability of cyclin D1 which facilitates cell cycle advancement and tumorigenesis. | Breast cancer, lung cancer, colorectal cancer, gastric cancer, and prostate cancer | [16,44,47,48,49] |
| MAPK pathway | The MAPK (mitogen-activated protein kinase) pathway is a system of intracellular communication that transmits signals from the cell surface to the nucleus. USP11 promotes the progression and chemoresistance of various cancers, and overexpression of USP11 has been associated with abnormal activation of the ERK-dependent MAPK pathway during the progression of colorectal carcinoma. | Breast cancer, colorectal cancer, lung cancer, gastric cancer, ovarian cancer, and hepatocellular carcinoma | [44,50] |
| Hypoxia pathways | Hypoxic conditions in tissues, particularly in cancer, are known to induce a cellular transcriptional response. Transcriptional complex hypoxia-inducible factors (HIF) are the key factors in this signaling pathway and the α-subunit of HIF triggers a cascade that leads to degradation by the 26S proteasome. USP22 has been shown to regulate the hypoxia pathway by stabilizing HIFs such as HIF-1α and enhancing their transcriptional activity, allowing cancer cells to adapt to hypoxic conditions. | Breast cancer, lung cancer, colorectal cancer, pancreatic cancer, cervical cancer, and head and neck cancer | [51,52] |
| USP Target | Name of the Inhibitor | Chemical Structure | IC50 Values | Mechanism of Action and Anticancer Activity | Refs |
|---|---|---|---|---|---|
| USP1 | SP-002 | Structure not disclosed. | 15.7 nM |
| [71,72] |
| USP1 + UAF1 | C527 | 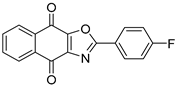 | 0.88 µM |
| [73,74] |
| SJB2-043 |  | 0.544 µM |
| [73,75] | |
| SJB3-019A | 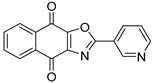 | 78.1 nM |
| ||
| ML323 |  | 76 nM |
| [76,77,78] | |
| USP1 + UAF1/USP7 | Pimozide | 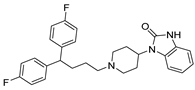 | 2 µM/47 µM |
| [79,80,81] |
| Trifluoperazine (TFP) |  | 8 µM/9 µM |
| [79,82] | |
| Flupenthixol |  | 7 µM/13 µM |
| [79] | |
| USP1 + UAF1/USP2/USP7/USP8 | Rottlerin |  | 8 µM/34 µM/13 µM/6 µM |
| [79,83,84] |
| USP1 + UAF1/USP12 | GW7647 |  | 5 µM/44 µM |
| [79,85] |
| USP 1/2/3/4/5/7/8/15/20/28/47/UCHL1/UCHL3/UCHL5 | PR619(Broad range or Pan DUBs inhibitor) |  | 3.9–8.9 µM |
| [54,86,87] |
| USP2 | Q29 | 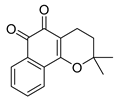 | <5 µM |
| [88,89] |
| ML364 |  | 1.1 µM |
| [89,90] | |
| LCAHA | 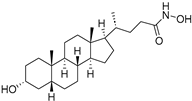 | 3.7–9.7 µM |
| [89,91] | |
| STD1T | 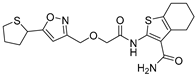 | 3.3 µM |
| [89,92] | |
| 6TG | 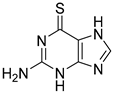 | 40 µM |
| [89,93,94] | |
| USP2/5/8/UCHL1/UCHL3 | RA-9 | 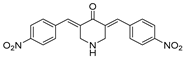 | Not determined by enzymatic assay |
| [54,89,95,96,97] |
| RA-14 | Chalcone derivative of RA-9 (structure not disclosed) | Not determined by enzymatic assay |
| [54,95] | |
| AM146 | Chalcone derivative of RA-9 (structure not disclosed) | Not determined by enzymatic assay |
| [54,95] | |
| USP2/7 | NSC632839 | 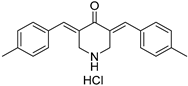 | 45 μM/37 μM |
| [89,98] |
| Compound 14 |  | 0.25 µM (USP2) |
| [54,89,99] | |
| USP4/5 | Vialinin A | 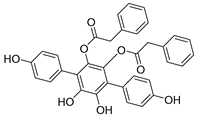 | 1.5 µM/5.9 µM |
| [100,101,102] |
| USP5 | USP5-IN-1 (compound 64) | 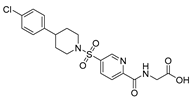 | 0.8–26 µM |
| [103] |
| USP5/7/8/13/14/15/22 | Curcusone D |  | Not determined by enzymatic assay |
| [54,104] |
| USP5/9X/14/24/37/UCHL5 | WP1130 (Degrasyn) | 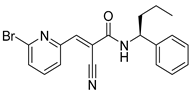 | 2.5–5 µM |
| [100,105,106,107,108] |
| EOAI3402143 (G9) |  | 1.6–5 µM |
| [49,107,109,110] | |
| USP7 | HBX-41108 | 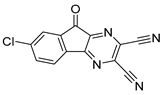 | 0.424 µM |
| [111,112] |
| HBX-28258 | 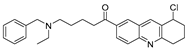 | 22.6 µM |
| [113,114] | |
| Spongiacidin C | 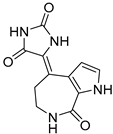 | 3.8 µM |
| [114,115] | |
| GNE-6640 |  | 0.75 µM |
| [116,117,118] | |
| GNE-6776 | 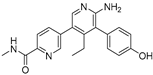 | 1.34 µM | |||
| XL188 |  | 90 nM |
| [119,120] | |
| ALM4 | 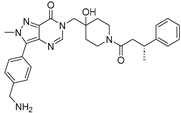 | 6 nM |
| [121,122] | |
| ALM46 |  | 0.09 µM | |||
| FT671 |  | 52 nM |
| [123,124] | |
| FT827 |  | Ki = 66 µM |
| [123] | |
| Parthenolide (PTL) | 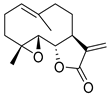 | 6.58 µM |
| [125] | |
| Costunolide | 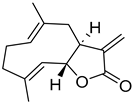 | Not determined by enzymatic assay |
| [125] | |
| YCH2823 | 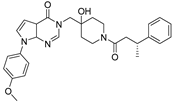 | 49.6 nM |
| [126] | |
| XL177A | 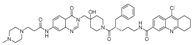 | 0.34 nM |
| [120] | |
| USP7/10 | HBX-19818 | 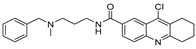 | 28.1 µM/14 µM/ |
| [113,114,127] |
| Compound 3 |  | 3.6 µM (USP7) |
| [54,62,127,128] | |
| USP7/47 | P22077 |  | 8 µM (USP7) |
| [54,86,129,130,131,132] |
| P5091 |  | 4.2 µM (USP7) |
| [54,133,134,135] | |
| P50429 |  | 0.42 µM/1 µM |
| [54,132,136] | |
| USP8 | HY50736 | 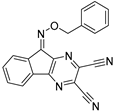 | 0.24 µM |
| [54,137] |
| HY50737A | 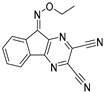 | 0.28 µM | |||
| USP9x | FT709 |  | 82 nM |
| [138] |
| USP10 | Compound 9 |  | Not determined by enzymatic assay |
| [54,62,127,128] |
| Wu-5 | 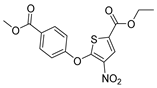 | 8.3 µM |
| [139,140] | |
| USP10/13 | Spautin-1 |  | 0.6–0.7 µM |
| [141,142,143] |
| USP11 | Mitoxantrone |  | 3.15 µM |
| [144] |
| USP12/USP46 | Galeterone | 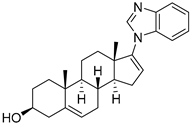 | 2.1–3.4/3.4–4.2 µM |
| [145] |
| USP14 | IU1 |  | 4–5 µM |
| [146,147] |
| IU1-47 | 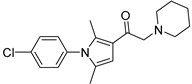 | 0.6 µM |
| [146,148,149,150] | |
| IU1-206 | 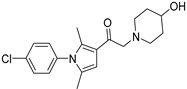 | Not determined by enzymatic assay | |||
| IU1-248 | 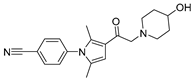 | 0.83 µM | |||
| USP14/UCHL5 | Auranofin | 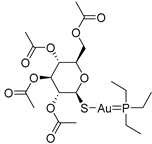 | Not determined by enzymatic assay |
| [151,152] |
| CuPT | 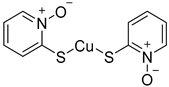 | Not determined by enzymatic assay |
| [153,154,155,156,157] | |
| PtPT |  | Not determined by enzymatic assay | |||
| NiPT |  | Not determined by enzymatic assay | |||
| b-AP15 |  | <10 µM |
| [158,159,160,161] | |
| USP22 | USP22i-S02 | 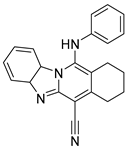 | Not determined by enzymatic assay |
| [162,163,164,165] |
| USP24 | NCI677397 | 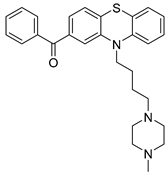 | Not determined by enzymatic assay |
| [118,166] |
| USP24-i-101 |  | Not determined by enzymatic assay |
| [167] | |
| USP25/28 | AZ1 | 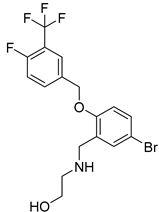 | 1.08 µM/1.76 µM |
| [168,169] |
| FT206 | 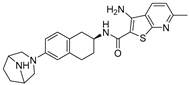 | 1.01/0.15 μM |
| [37,169] | |
| Vismodegib (VSM) | 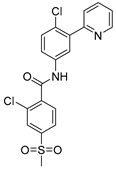 | 2.92/3.51–4.41 μM |
| [169,170,171,172] | |
| USP30 | MF-094 |  | 0.12 µM |
| [173,174,175] |
| 15-oxospiramilactone |  | Not determined by enzymatic assay |
| [176,177] | |
| Compound 39 (CMPD-39) | 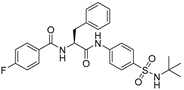 | 20 nM |
| [178,179,180] | |
| USP36 | Cinobufotalin |  | 2.75 μM |
| [36] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakkar, M.; Khalil, S.; Bhayekar, K.; Kushwaha, N.D.; Samarbakhsh, A.; Dorandish, S.; Edwards, H.; Dou, Q.P.; Ge, Y.; Gavande, N.S. Ubiquitin-Specific Protease Inhibitors for Cancer Therapy: Recent Advances and Future Prospects. Biomolecules 2025, 15, 240. https://doi.org/10.3390/biom15020240
Bakkar M, Khalil S, Bhayekar K, Kushwaha ND, Samarbakhsh A, Dorandish S, Edwards H, Dou QP, Ge Y, Gavande NS. Ubiquitin-Specific Protease Inhibitors for Cancer Therapy: Recent Advances and Future Prospects. Biomolecules. 2025; 15(2):240. https://doi.org/10.3390/biom15020240
Chicago/Turabian StyleBakkar, Mohamad, Sara Khalil, Komal Bhayekar, Narva Deshwar Kushwaha, Amirreza Samarbakhsh, Sadaf Dorandish, Holly Edwards, Q. Ping Dou, Yubin Ge, and Navnath S. Gavande. 2025. "Ubiquitin-Specific Protease Inhibitors for Cancer Therapy: Recent Advances and Future Prospects" Biomolecules 15, no. 2: 240. https://doi.org/10.3390/biom15020240
APA StyleBakkar, M., Khalil, S., Bhayekar, K., Kushwaha, N. D., Samarbakhsh, A., Dorandish, S., Edwards, H., Dou, Q. P., Ge, Y., & Gavande, N. S. (2025). Ubiquitin-Specific Protease Inhibitors for Cancer Therapy: Recent Advances and Future Prospects. Biomolecules, 15(2), 240. https://doi.org/10.3390/biom15020240