

Metabolomics Analysis and Biochemical Profiling of Arsenic-Induced Metabolic Impairment and Disease Susceptibility

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Arsenic Solution
2.2. Preparation of Resveratrol Solution
2.3. Preparation of Metformin Solution
2.4. Study Design—Animal Grouping, Dosing Schedule and Routine Monitoring
- Group-1 (CONT): Control group, given purified water.
- Group-2 (Na3AsO4): Arsenic-exposed group, administered a 50-ppm solution of sodium arsenate (50 mg/L) in purified water.
- Group-3 (MFT + Na3AsO4): Arsenic-exposed group treated with Metformin (250 mg/kg body weight/day).
- Group-4 (RSV + Na3AsO4): Arsenic-exposed group treated with resveratrol in peanut oil at a dosage of 8 mg/kg body weight/day.
2.5. Biochemical Analysis
2.6. Biomarker Analysis for Estimation of Metabolic Impairment
2.6.1. Estimation of Glycemic Index Biomarkers
2.6.2. Estimation of Carbohydrate Metabolism Biomarkers
2.6.3. Estimation of Oxidative Stress Biomarkers
2.6.4. Estimation of Liver Function Biomarkers
2.6.5. Estimation of Kidney Function Biomarker
2.6.6. Estimation of Inflammatory Biomarkers
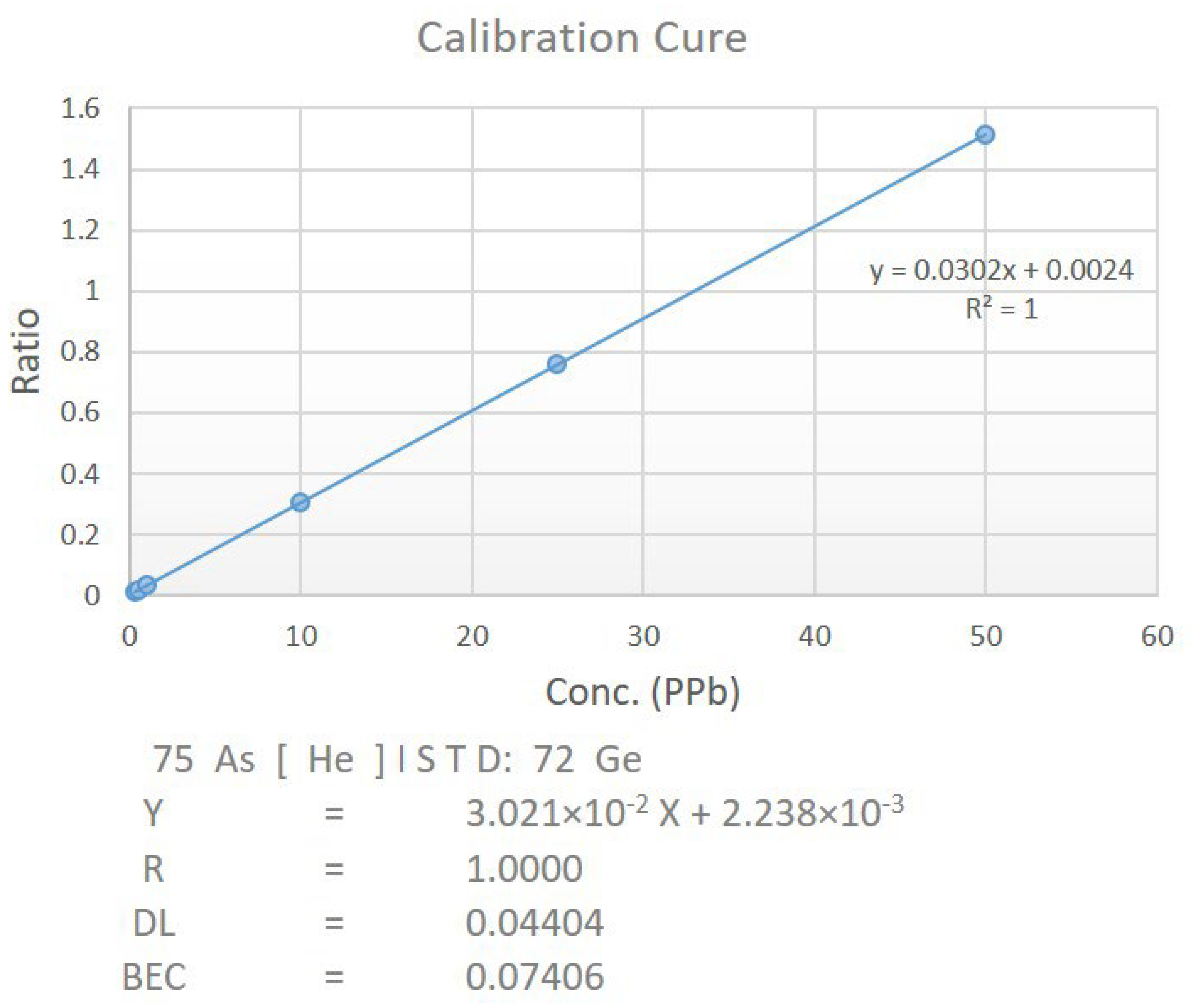
2.7. Estimation of Arsenic by ICP-MS
2.8. Estimation of Gene Expression Metabolizing Enzymes
2.8.1. RNA Isolation
2.8.2. cDNA Synthesis
2.8.3. qRT-PCR Procedure
2.9. Analysis of Amino Acid
2.10. Metabolomics Analysis
Qualitative Analysis of Metabolomes with MS/MS
2.11. Histopathological Assessment
2.12. Statistical Analysis
3. Results
3.1. Effect on Body Weight
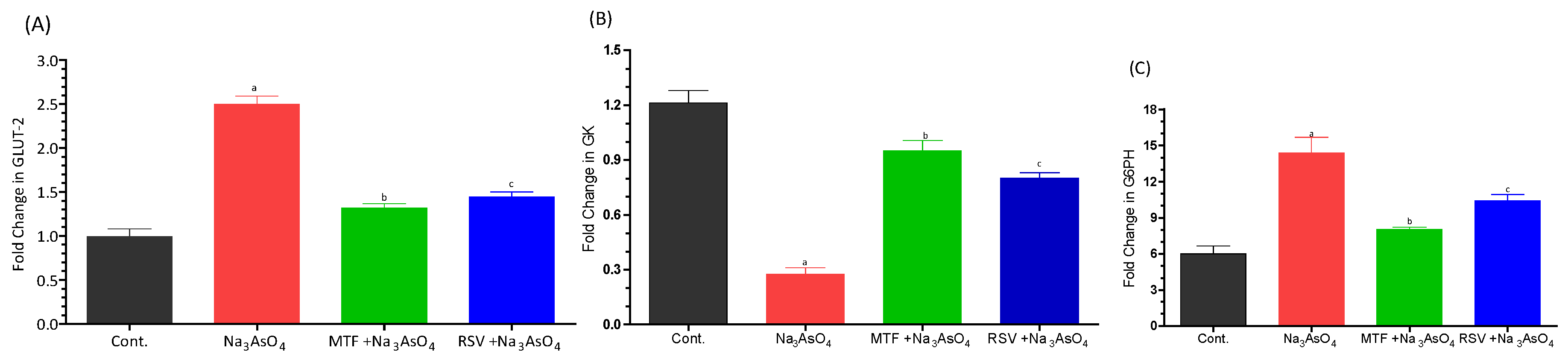
3.2. Effect on Glycemic Index Biomarkers
3.3. Effect on Carbohydrate Metabolism Biomarkers
3.4. Effect on Biomarkers of Oxidative Stress and Lipid Peroxidation
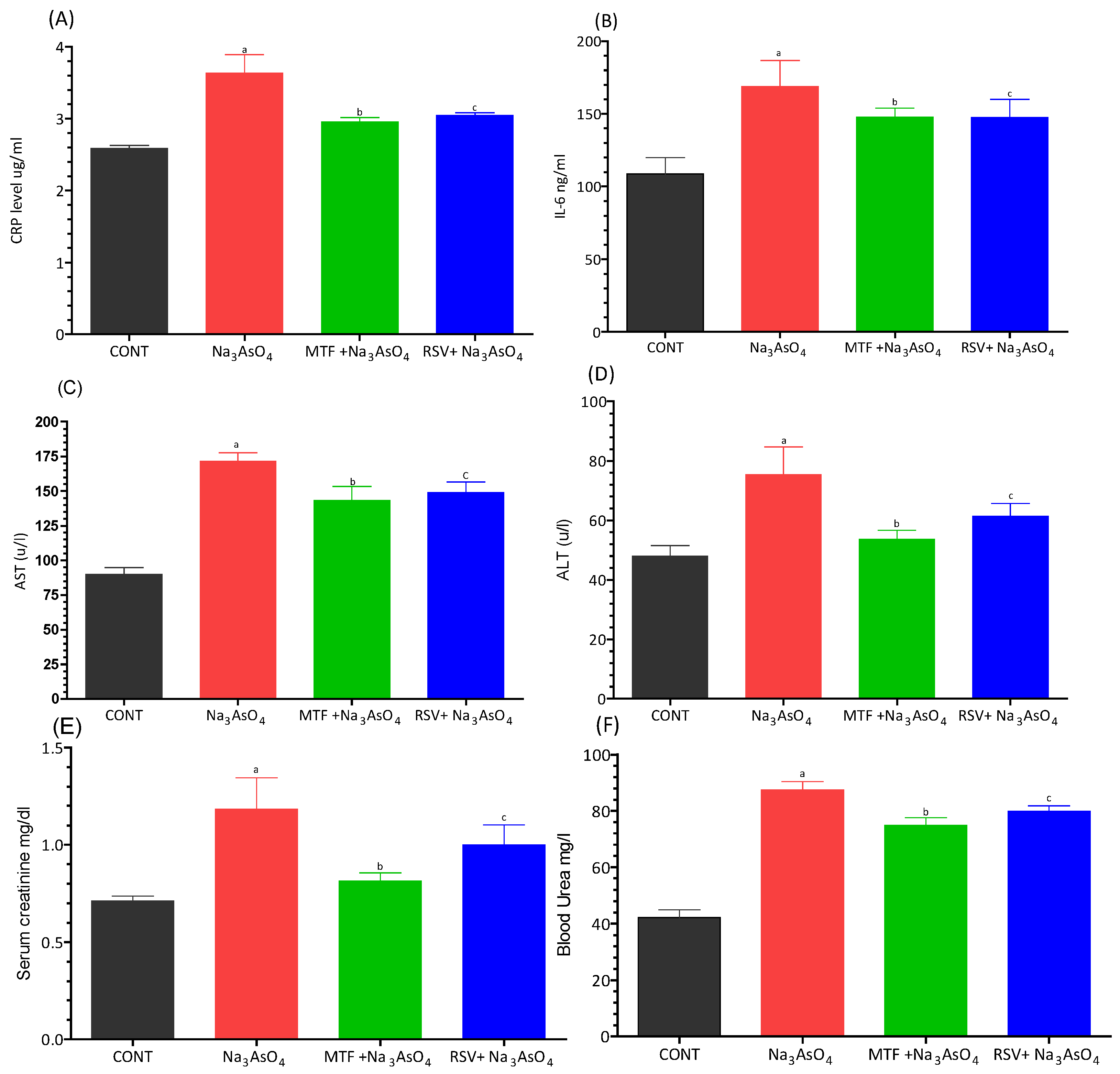
3.5. Effect on Inflammatory, Kidney and Liver Biomarkers
3.6. Estimation of Arsenic by ICP-MS
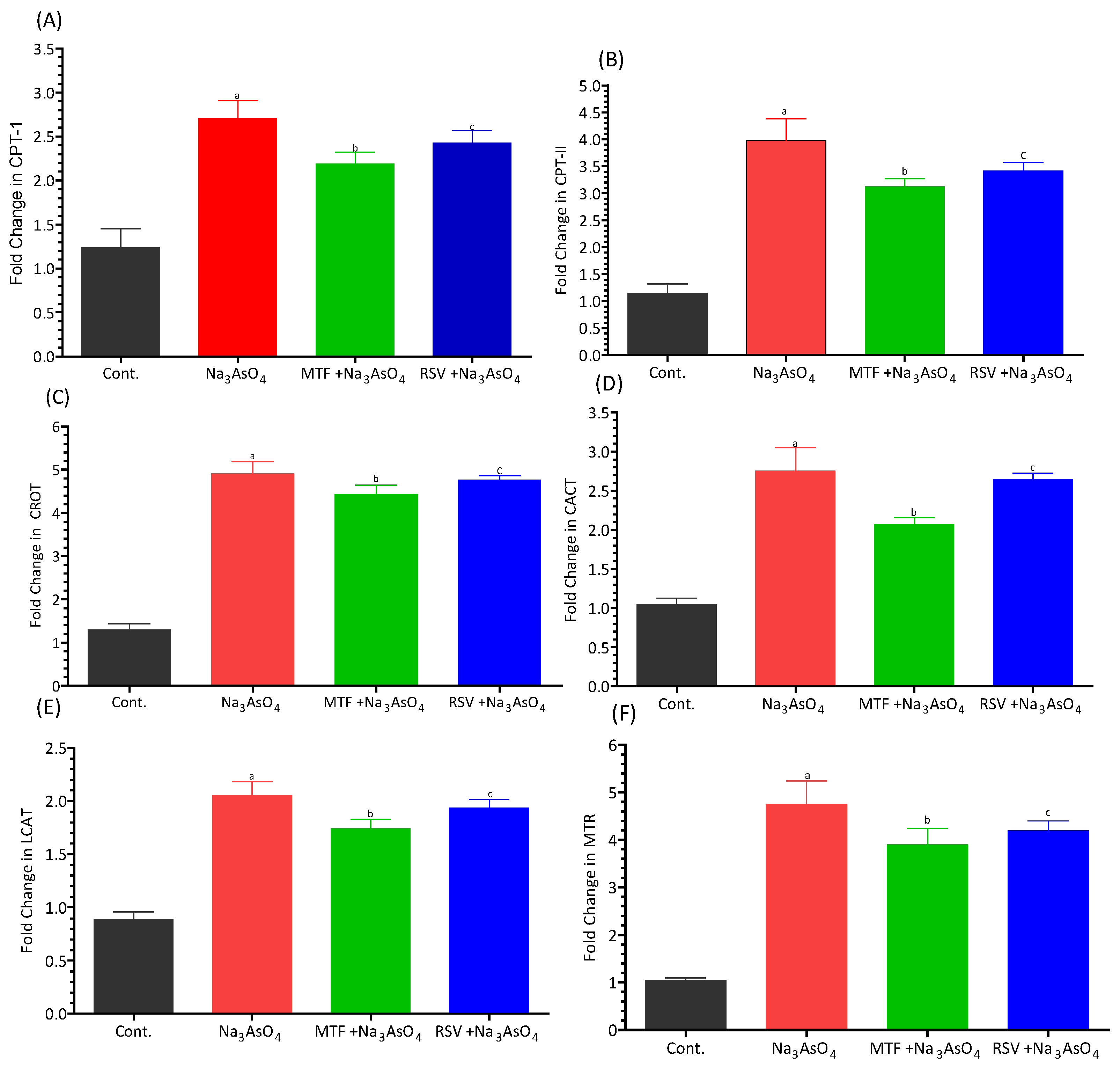
3.7. Gene Expression
3.8. Analysis of Amino Acid
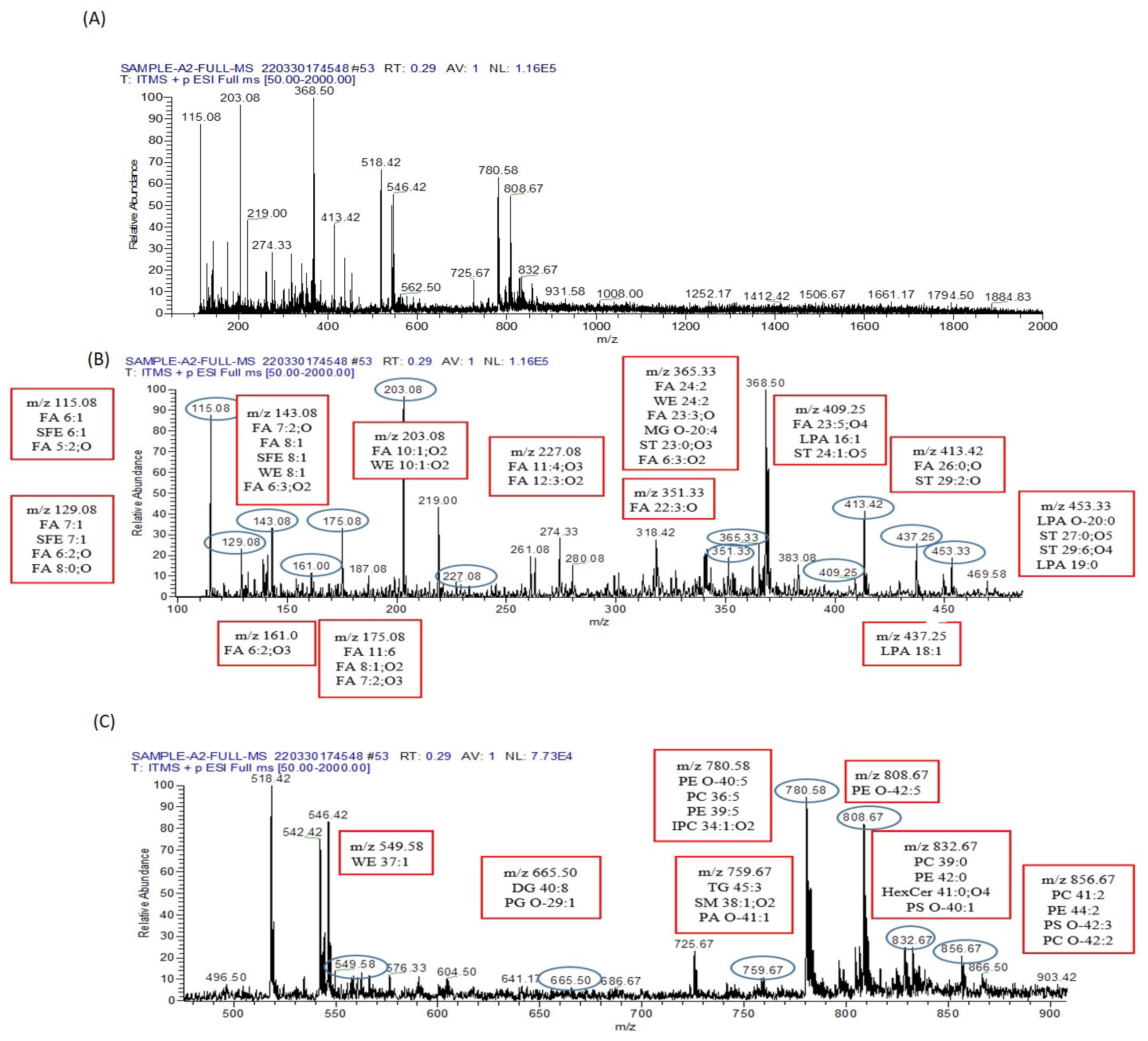
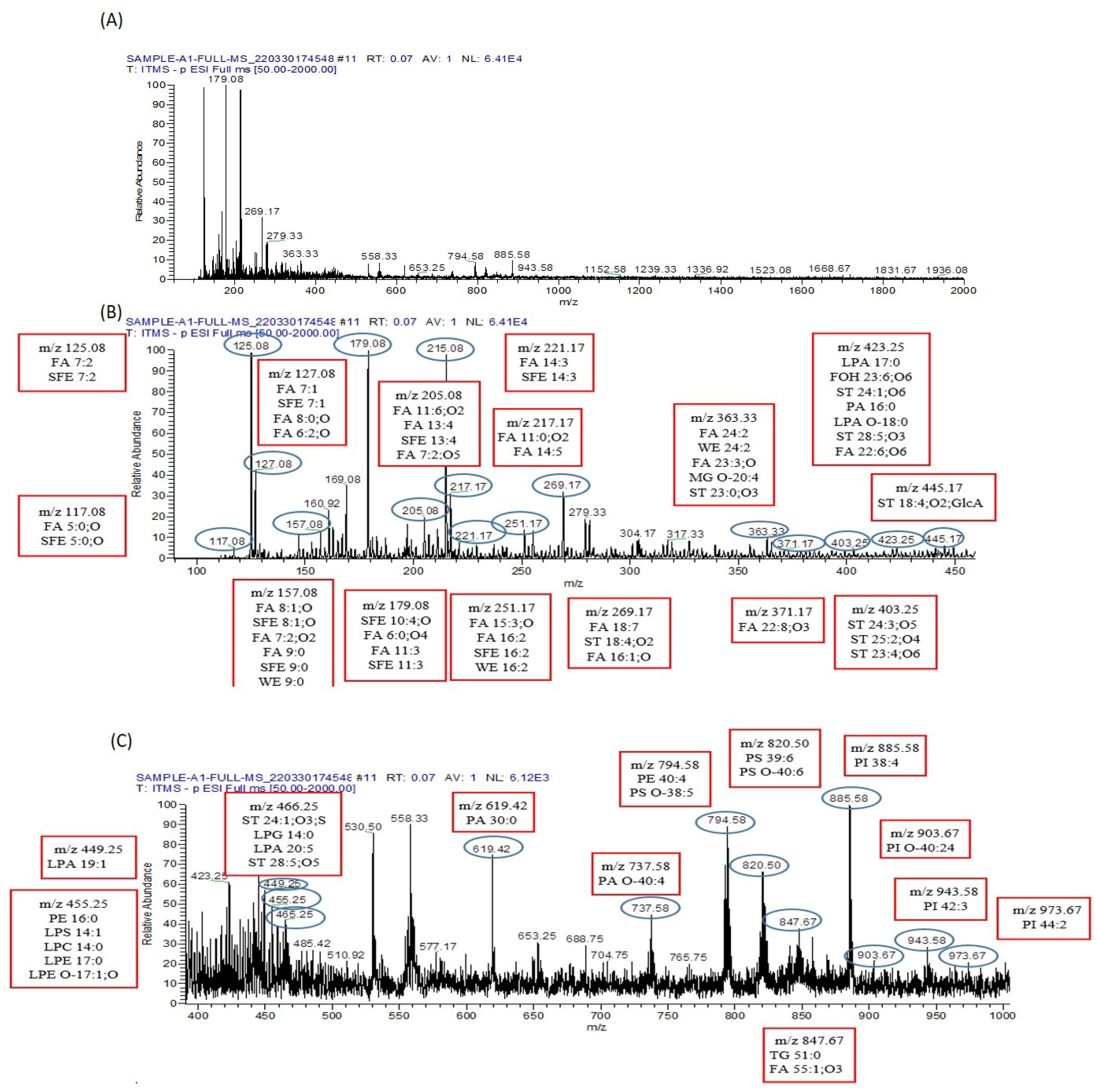
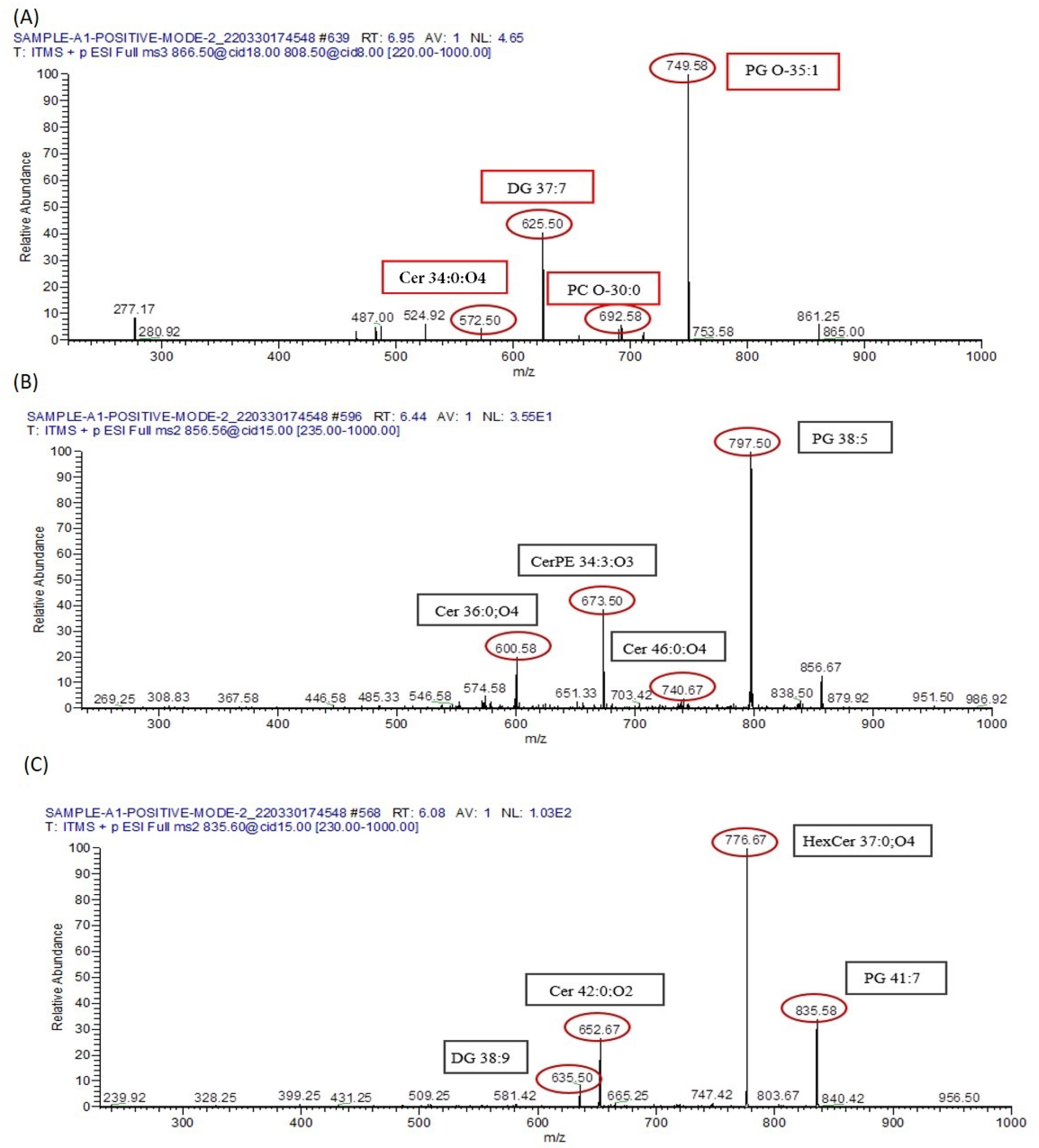
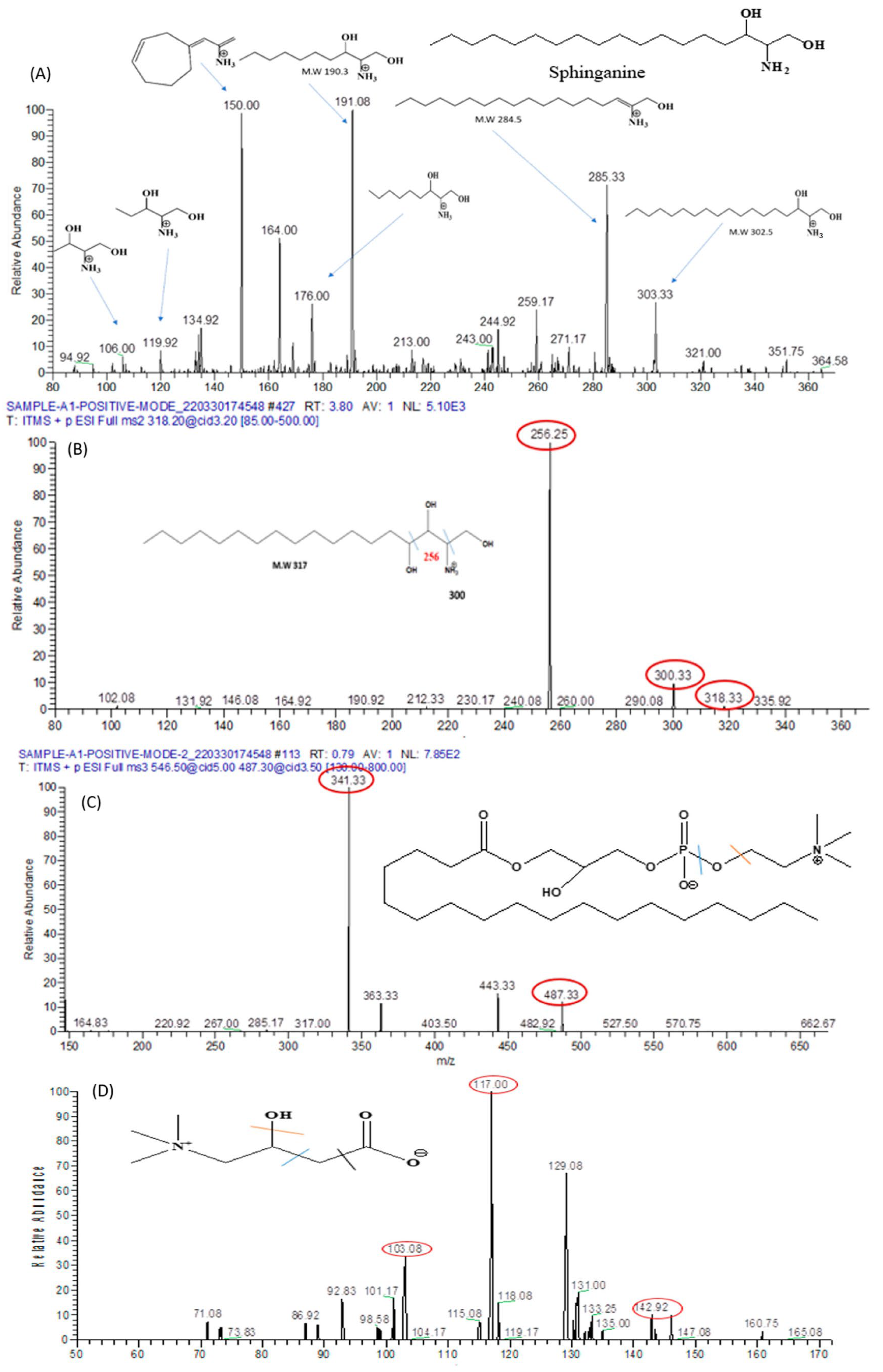
3.9. Metabolomics Analysis
3.9.1. Analysis of Metabolomes by MS/MS
3.9.2. Lipid Metabolomes Based on Peak Analysis from Database
3.9.3. Identification of Metabolomes with Molecular Ion Fragmentation Method
3.9.4. Lipid Metabolomes
Sphinganine
Phyto-Sphinganine
Lysophosphatidyl-Choline
3.9.5. L-Carnitine
3.10. Identified Amino Acids
3.10.1. Arginine
3.10.2. Dimethylarginine
3.10.3. Isoleucine
3.10.4. Miscellaneous Metabolites
3.11. Inorganic Arsenic and Its Metabolites
3.12. Histopathology
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Genchi, G.; Lauria, G.; Catalano, A.; Carocci, A.; Sinicropi, M.S. Arsenic: A review on a great health issue worldwide. Appl. Sci. 2022, 12, 6184. [Google Scholar] [CrossRef]
- Mandal, B.K.; Suzuki, K.T. Arsenic round the world: A review. Talanta 2002, 58, 201–235. [Google Scholar] [CrossRef] [PubMed]
- Samrana, S.; Ali, I.; Azizullah, A.; Daud, M.K.; Gan, Y. Arsenic-Based Pollutio n Status in Pakistan. Ann. Agric. Crop Sci. 2017, 2, 1027. [Google Scholar]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Arsenic, Metals, Fibres, and Dusts; IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; WHO: Geneva, Switzerland, 2012; Volume 100, p. 11. [Google Scholar]
- Wang, S.; Mulligan, C.N. Speciation and surface structure of inorganic arsenic in solid phases: A review. Environ. Int. 2008, 34, 867–879. [Google Scholar] [CrossRef]
- Council, N.R. Distribution of Arsenic in the Environment. In Arsenic: Medical and Biologic Effects of Environmental Pollutants; National Academies Press: Washington, DC, USA, 1977. [Google Scholar]
- Atsdr, U. Toxicological Profile for Arsenic; Agency for Toxic Substances and Disease Registry, Division of Toxicology: Atlanta, GA, USA, 2007. [Google Scholar]
- Smedley, P.L.; Kinniburgh, D.G. United Nations Synthesis Report on Arsenic in Drinking Water; British Geological Survey: Nottingham, UK, 2001; pp. 1–61. [Google Scholar]
- Raju, N.J. Arsenic in the geo-environment: A review of sources, geochemical processes, toxicity and removal technologies. Environ. Res. 2022, 203, 111782. [Google Scholar] [CrossRef] [PubMed]
- Siddique, T.; Dutta, N.K.; Choudhury, N.R. Nanofiltration for arsenic removal: Challenges, recent developments, and perspectives. Nanomaterials 2020, 10, 1323. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.; Hara, T.O.; Tianc, F.; Singh, B. Review of analytical techniques for arsenic detection and determination in drinking water. Environ. Sci. Adv. 2023, 2, 171–195. [Google Scholar] [CrossRef]
- Callegari, A.; Ferronato, N.; Rada, E.C.; Capodaglio, A.G.; Torretta, V. Assessment of arsenic removal efficiency by an iron oxide-coated sand filter process. Environ. Sci. Pollut. Res. 2018, 25, 26135–26143. [Google Scholar] [CrossRef]
- HOwD, R.A. Introduction to drinking water risk assessment. In Risk Assessment for Chemicals in Drinking Water; John Wiley & Sons: Hoboken, NJ, USA, 2007; pp. 1–15. [Google Scholar]
- Chen, C. Environmental health issues in public health. In Oxford Textbook of Public Health, Volume 2: The Methods of Public Health, 5th ed.; Oxford University Press: Oxford, UK, 2009; pp. 825–840. [Google Scholar]
- Saha, J.; Dikshit, A.K.; Bandyopadhyay, M.; Saha, K.C. A review of arsenic poisoning and its effects on human health. Crit. Rev. Environ. Sci. Technol. 1999, 29, 281–313. [Google Scholar] [CrossRef]
- Tseng, C.-H. The potential biological mechanisms of arsenic-induced diabetes mellitus. Toxicol. Appl. Pharmacol. 2004, 197, 67–83. [Google Scholar] [CrossRef]
- Kulshrestha, A.; Jarouliya, U.; Prasad, G.; Flora, S.; Bisen, P.S. Arsenic-induced abnormalities in glucose metabolism: Biochemical basis and potential therapeutic and nutritional interventions. World J. Transl. Med. 2014, 3, 96–111. [Google Scholar] [CrossRef]
- García-Sevillano, M.; Contreras-Acuña, M.; García-Barrera, T.; Navarro, F.; Gómez-Ariza, J.L. Metabolomic study in plasma, liver and kidney of mice exposed to inorganic arsenic based on mass spectrometry. Anal. Bioanal. Chem. 2014, 406, 1455–1469. [Google Scholar] [CrossRef]
- Rivas-Santiago, C.; González-Curiel, I.; Zarazua, S.; Murgu, M.; Cardona, A.R.; Lazalde, B.; Lara-Ramírez, E.E.; Vázquez, E.; Castañeda-Delgado, J.E.; Rivas-Santiago, B.; et al. Lipid metabolism alterations in a rat model of chronic and intergenerational exposure to arsenic. Biomed Res. Int. 2019, 2019, 4978018. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Li, X.; Zhang, Y.; Li, J.; Yang, J.; Jiang, H.; Sun, G.; Huo, T. Metabolic characteristics related to the hazardous effects of environmental arsenic on humans: A metabolomic review. Ecotoxicol. Environ. Saf. 2022, 236, 113459. [Google Scholar] [CrossRef]
- Zhang, J.; Shen, H.; Xu, W.; Xia, Y.; Barr, D.B.; Mu, X.; Wang, X.; Liu, L.; Huang, Q.; Tian, M. Urinary metabolomics revealed arsenic internal dose-related metabolic alterations: A proof-of-concept study in a Chinese male cohort. Environ. Sci. Technol. 2014, 48, 12265–12274. [Google Scholar] [CrossRef]
- Araújo, A.M.; Shen, H.; Xu, W.; Xia, Y.; Barr, D.B.; Mu, X.; Wang, X.; Liu, L.; Huang, Q.; Tian, M. Toxicometabolomics: Small molecules to answer big toxicological questions. Metabolites 2021, 11, 692. [Google Scholar] [CrossRef] [PubMed]
- Association, A.D. Diagnosis and classification of diabetes mellitus. Diabetes Care 2010, 33, S62–S69. [Google Scholar] [CrossRef] [PubMed]
- Holt, R.I. Diagnosis, epidemiology and pathogenesis of diabetes mellitus: An update for psychiatrists. Br. J. Psychiatry 2004, 184, s55–s63. [Google Scholar] [CrossRef]
- Akash, M.S.H.; Sharif, H.; Rehman, K.; Rasheed, S.; Kamal, S. Biochemical Profiling of Arsenic Trioxide-Induced Impaired Carbohydrate Metabolism and its Therapeutic Intervention via Modulation of Metabolic Pathways. Pak. J. Zool. 2022, 54, 1–10. [Google Scholar] [CrossRef]
- Perrone, D.; Fuggetta, M.P.; Ardito, F.; Cottarelli, A.; De Filippis, A.; Ravagnan, G.; De Maria, S.; Lo Muzio, L. Resveratrol (3,5,4′-trihydroxystilbene) and its properties in oral diseases. Exp. Ther. Med. 2017, 14, 3–9. [Google Scholar] [CrossRef]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of type 2 diabetes mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Mahjabeen, W.; Khan, D.A.; Mirza, S.A. Role of resveratrol supplementation in regulation of glucose hemostasis, inflammation and oxidative stress in patients with diabetes mellitus type 2: A randomized, placebo-controlled trial. Complement. Ther. Med. 2022, 66, 102819. [Google Scholar] [CrossRef]
- Fu, Z.; Xi, S. The effects of heavy metals on human metabolism. Toxicol. Mech. Methods 2020, 30, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Sabir, S.; Akash, M.S.H.; Fiayyaz, F.; Saleem, U.; Mehmood, M.H.; Rehman, K. Role of cadmium and arsenic as endocrine disruptors in the metabolism of carbohydrates: Inserting the association into perspectives. Biomed. Pharmacother. 2019, 114, 108802. [Google Scholar] [CrossRef]
- Zhang, H.-N.; Yang, L.; Ling, J.-Y.; Czajkowsky, D.M.; Wang, J.-F.; Zhang, X.-W.; Zhou, Y.-M.; Ge, F.; Yang, M.-K.; Xiong, Q.; et al. Systematic identification of arsenic-binding proteins reveals that hexokinase-2 is inhibited by arsenic. Proc. Natl. Acad. Sci. USA 2015, 112, 15084–15089. [Google Scholar] [CrossRef] [PubMed]
- Huq, M.E.; Fahad, S.; Shao, Z.; Sarven, M.S.; Khan, I.A.; Alam, M.; Saeed, M.; Ullah, H.; Adnan, M.; Saud, S.; et al. Arsenic in a groundwater environment in Bangladesh: Occurrence and mobilization. J. Environ. Manag. 2020, 262, 110318. [Google Scholar] [CrossRef]
- Kashtoh, H.; Baek, K.-H. Recent updates on phytoconstituent alpha-glucosidase inhibitors: An approach towards the treatment of type two diabetes. Plants 2022, 11, 2722. [Google Scholar] [CrossRef]
- Simon, C.; Herling, A.W.; Preibisch, G.; Burger, H.-J. Upregulation of hepatic glucose 6-phosphatase gene expression in rats treated with an inhibitor of glucose-6-phosphate translocase. Arch. Biochem. Biophys. 2000, 373, 418–428. [Google Scholar] [CrossRef]
- Irshad, K.; Rehman, K.; Fiayyaz, F.; Sharif, H.; Murtaza, G.; Kamal, S.; Akash, M.S.H. Role of Heavy Metals in Metabolic Disorders. In Endocrine Disrupting Chemicals-Induced Metabolic Disorders and Treatment Strategies; Springer: Berlin/Heidelberg, Germany, 2021; pp. 203–219. [Google Scholar]
- Singh, A.P.; Goel, R.K.; Kaur, T. Mechanisms pertaining to arsenic toxicity. Toxicol. Int. 2011, 18, 87. [Google Scholar]
- Khan, F.; Hodjat, M.; Rahimifard, M.; Nigjeh, M.N.; Azizi, M.; Baeeri, M.; Bayrami, Z.; Gholami, M.; Hassani, S.; Abdollahi, M. Assessment of arsenic-induced modifications in the DNA methylation of insulin-related genes in rat pancreatic islets. Ecotoxicol. Environ. Saf. 2020, 201, 110802. [Google Scholar] [CrossRef]
- Rubino, F.M. Toxicity of glutathione-binding metals: A review of targets and mechanisms. Toxics 2015, 3, 20–62. [Google Scholar] [CrossRef]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef]
- Hu, Y.; Li, J.; Lou, B.; Wu, R.; Wang, G.; Lu, C.; Wang, H.; Pi, J.; Xu, Y. The role of reactive oxygen species in arsenic toxicity. Biomolecules 2020, 10, 240. [Google Scholar] [CrossRef]
- Ito, F.; Sono, Y.; Ito, T. Measurement and clinical significance of lipid peroxidation as a biomarker of oxidative stress: Oxidative stress in diabetes, atherosclerosis, and chronic inflammation. Antioxidants 2019, 8, 72. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.; Akinloye, O. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Wang, L.; Xu, Z. Effects of arsenic (AsIII) on lipid peroxidation, glutathione content and antioxidant enzymes in growing pigs. Asian-Australas. J. Anim. Sci. 2006, 19, 727–733. [Google Scholar] [CrossRef]
- Dandona, P.; Aljada, A.; Bandyopadhyay, A. Inflammation: The link between insulin resistance, obesity and diabetes. Trends Immunol. 2004, 25, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Rouse, R.L.; Stewart, S.R.; Thompson, K.L.; Zhang, J. Kidney injury biomarkers in hypertensive, diabetic, and nephropathy rat models treated with contrast media. Toxicol. Pathol. 2013, 41, 662–680. [Google Scholar] [CrossRef]
- Council, N.R. Critical Aspects of EPA’s IRIS Assessment of Inorganic Arsenic: Interim Report; National Academies Press: Washington, DC, USA, 2013. [Google Scholar]
- Minatel, B.C.; Sage, A.P.; Anderson, C.; Hubaux, R.; Marshall, E.A.; Lam, W.L.; Martinez, V.D. Environmental arsenic exposure: From genetic susceptibility to pathogenesis. Environ. Int. 2018, 112, 183–197. [Google Scholar] [CrossRef]
- Reichard, J.F.; Puga, A. Effects of arsenic exposure on DNA methylation and epigenetic gene regulation. Epigenomics 2010, 2, 87–104. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.; Fatuzzo, N.; Weng, N.; Michno, W.; Dong, W.; Kienle, M.; Dai, Y.; Pasca, A.; Abu-Remaileh, M.; Rasgon, N.; et al. Carnitine octanoyltransferase is important for the assimilation of exogenous acetyl-L-carnitine into acetyl-CoA in mammalian cells. J. Biol. Chem. 2023, 299, 102848. [Google Scholar] [CrossRef]
- Bonilha, I.; Zimetti, F.; Zanotti, I.; Papotti, B.; Sposito, A.C. Dysfunctional high-density lipoproteins in type 2 diabetes mellitus: Molecular mechanisms and therapeutic implications. J. Clin. Med. 2021, 10, 2233. [Google Scholar] [CrossRef]
- Röschinger, W.; Muntau, A.C.; Duran, M.; Dorland, L.; Ijlst, L.; Wanders, R.J.; A Roscher, A. Carnitine-acylcarnitine translocase deficiency: Metabolic consequences of an impaired mitochondrial carnitine cycle. Clin. Chim. Acta 2000, 298, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, J. Pathways and regulation of homocysteine metabolism in mammals. In Seminars in Thrombosis and Hemostasis; Thieme Medical Publishers, Inc.: New York, NY, USA, 2000. [Google Scholar]
- Wang, X.; Mu, X.; Zhang, J.; Huang, Q.; Alamdar, A.; Tian, M.; Liu, L.; Shen, H. Serum metabolomics reveals that arsenic exposure disrupted lipid and amino acid metabolism in rats: A step forward in understanding chronic arsenic toxicity. Metallomics 2015, 7, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Deng, H.; Wang, D.; Wang, J.; Huang, H.; Qiu, J.; Li, Y.; Zou, T.; Guo, L. Changes in metabolomics and lipidomics in brain tissue and their correlations with the gut microbiome after chronic food-derived arsenic exposure in mice. Ecotoxicol. Environ. Saf. 2021, 228, 112935. [Google Scholar] [CrossRef]
- Sokolowska, E.; Blachnio-Zabielska, A. The role of ceramides in insulin resistance. Front. Endocrinol. 2019, 10, 577. [Google Scholar] [CrossRef]
- Petersen, M.C.; Shulman, G.I. Roles of diacylglycerols and ceramides in hepatic insulin resistance. Trends Pharmacol. Sci. 2017, 38, 649–665. [Google Scholar] [CrossRef]
- Chavez, J.A.; Summers, S.A. A ceramide-centric view of insulin resistance. Cell Metab. 2012, 15, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Konstantynowicz-Nowicka, K.; Harasim, E.; Baranowski, M.; Chabowski, A. New evidence for the role of ceramide in the development of hepatic insulin resistance. PLoS ONE 2015, 10, e0116858. [Google Scholar] [CrossRef]
- Ferrannini, E.; Iervasi, G.; Cobb, J.; Ndreu, R.; Nannipieri, M. Insulin resistance and normal thyroid hormone levels: Prospective study and metabolomic analysis. Am. J. Physiol.-Endocrinol. Metab. 2017, 312, E429–E436. [Google Scholar] [CrossRef] [PubMed]
- Ferrannini, E.; Natali, A.; Camastra, S.; Nannipieri, M.; Mari, A.; Adam, K.-P.; Milburn, M.V.; Kastenmüller, G.; Adamski, J.; Tuomi, T.; et al. Early metabolic markers of the development of dysglycemia and type 2 diabetes and their physiological significance. Diabetes 2013, 62, 1730–1737. [Google Scholar] [CrossRef] [PubMed]
- Kumbhare, V.; Gajbe, U.; Singh, B.R.; Reddy, A.K.; Shukla, S. Histological & histochemical changes in liver of adult rats treated with monosodium glutamate: A light microscopic study. World J. Pharm. Pharm. Sci. 2015, 4, 898–911. [Google Scholar]
- Youssef, S. Light and electron microscopic study of the effect of orlistat on the liver of adult male albino rats and the possible protective role of β-carotene. Forensic Med. Anat. Res. 2018, 6, 20–36. [Google Scholar] [CrossRef]





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolism | Gene Name | Primer | Primer Sequence | Size (bp) |
|---|---|---|---|---|
| Lipid metabolism | Beta-actin | Forward | 5’-CCCATCTATGAGGGTTACGC-3‘ | 150 |
| Reverse | 5’-TTTAATGTCACGCACGATTTC-3‘ | |||
| Carnitinepalmitoyl- transferase I (CPT-I) | Forward | 5′-ATCCACCATTCCACTCTGCT-3′ | 107 | |
| Reverse | 5′-TGTGCCTGCTGTCCTTGATA-3′ | |||
| Carnitinepalmitoyl transferase II (CPT-II) | Forward | 5′-CTGTCCACCAGCACTCTGAA-3′ | 111 | |
| Reverse | 5′-GCAACCTATCCAGTCATCGT-3′ | |||
| Lecithin–cholesterol acyltransferase (LCAT) | Forward | 5′-CTCCTTCTGGCTCCTCAATG-3′ | 171 | |
| Reverse | 5′-TCCTCTGTCTTTCGGTAGCAC-3′ | |||
| Carnitine O-octanoyl6transferase (CROT) | Forward | 5′-AGACGGAAGGGAGATGGAG-3′ | 168 | |
| Reverse | 5′-AAGATGTGAAGGTAGATGCTGCT-3′ | |||
| Mitochondrial carnitine/acylcarnitine carrier protein (CACT) | Forward | 5′-TTCTCCACTGCTGCTCCTG-3′ | 100 | |
| Reverse | 5′-CCTGTCTGCTCCCATTCAG-3′ | |||
| Amino acid metabolism | 5-methyl tetrahydrofolate-homocysteine methyltransferase (MTR) | Forward | 5′-GGTTCGGTTGAAGAAGAGGA-3′ | 112 |
| Reverse | 5′-TATTACAGCCCAGCACCACA-3′ | |||
| Carbohydrate metabolism | Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | Forward | 5′-CCCGTAGACAAAATGGTGAAGGTC-3′ | 215 |
| Reverse | 5′-GCCAAAGTTGTCATGGATGACC-3′ | |||
| Glucose transporter-2 (GLUT-2) | Forward | 5′-TTAGCAACTGGGTCTGCAAT-3′ | 243 | |
| Reverse | 5′-TCTCTGAAGACGCCAGGAAT-3′ | |||
| Glucokinase (GK) | Forward | 5′-CACCCAACTGCGAAATCACC-3′ | 162 | |
| Reverse | 5′-CATTTGTGGGGTGTGGAGTC-3′ | |||
| Glucose 6 Phosphate (G6PH) | Forward | 5′-AAAGAGACTGTCGGCATCAATC-3′ | ||
| Reverse | 5′-AAGAGGCTGGCAAAGGGTGTAG-3′ |
| Parameter | Particulars |
|---|---|
| Instrument with model | Linear Ion Trap Mass spectrometer LTQ XL (Thermo Electron Scientific, Waltham, MA, USA) equipped with electro spray ionization source |
| Software | Xcalibur 2.0.7 |
| Fragmentation (MS/MS) | Various peaks were selected for fragmentation, using collision-induced dissociation (CID) energy ranging from 20–30 |
| Mode of ionization | Both negative and positive scan ion modes |
| Mode of injection | Direct insertion method |
| Scanning mass range | 50–2000 m/z |
| Sheath gas flow rate | 17 units |
| Auxiliary gas flow rate | 6 units |
| Solvent | Methanol |
| Flow rate | 9.8 μL/min |
| Capillary voltage | 4.7 kV |
| Capillary temperature | 278 °C |
| Metabolites | Formula | M.wt. | Ion | Precursor Ion (m/z) | Product Ion (m/z) |
|---|---|---|---|---|---|
| Lipid metabolomes | |||||
| Sphinganine | C18H39NO2 | 302 | [M+H]+ | 303 | 285, 190.3, 176.3, 150, 119.92 and 106. |
| Phytosphingosine | C18H39NO3 | 317 | [M+H]+ | 318 | 300 and 256 |
| Lyso phosphatidylcholine | C26H54NO7P | 523 | [M+H]+ | 546 | 487, 341, 404, and 443 |
| Carnitine | C7H16NO3 | 162 | [M-H]− | 160.75 | 162.92,102.92,84.92 and 60.08 |
| Amino acid | |||||
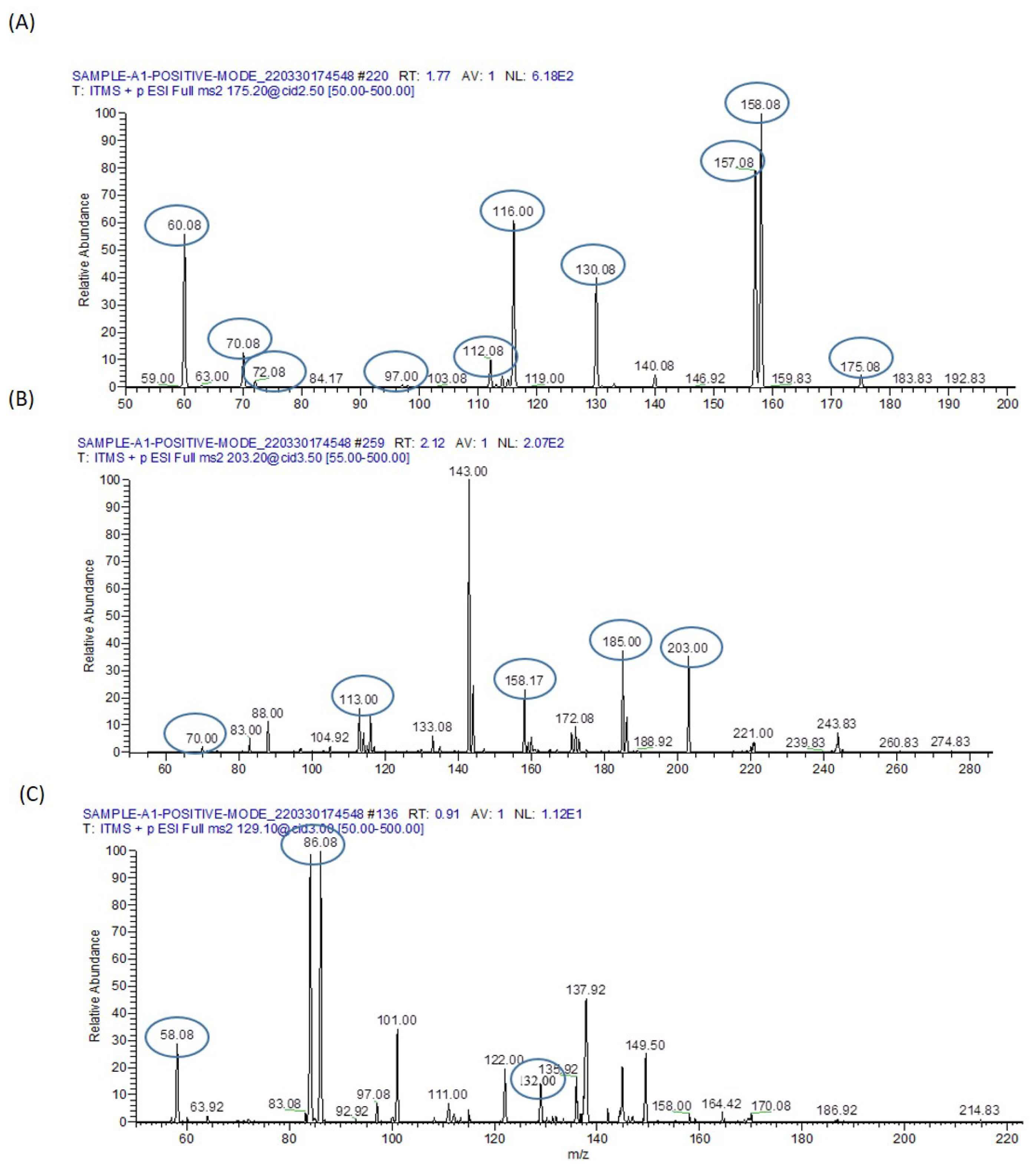
| Arginine | C6H14N4O2 | 174.20 | [M+H]+ | 175.08 | 130.08,116.00,70.08 and 60.08 |
| Dimethyl Arginine | C8H18N4O2 | 202.25 | [M+H]+ | 203.00 | 185.00,158.17,116 and 70.00 |
| Isoleucine | C6H13NO2 | 131.17 | [M+H]+ | 132.00 | 86.08 and 58.08 |
| Valine | C5H11NO2 | 117.14 | [M+H]+ | 118 | 72.08 |
| Thronine | C4H9NO3 | 119.11 | [M+H]+ | 120 | 74.08 |
| Inorganic Arsenic with Its Metabolites | Ion | Molecular Ion Structure | Molecular Ion m/z | Characteristic Fragments | Fragment Ion Structure |
|---|---|---|---|---|---|
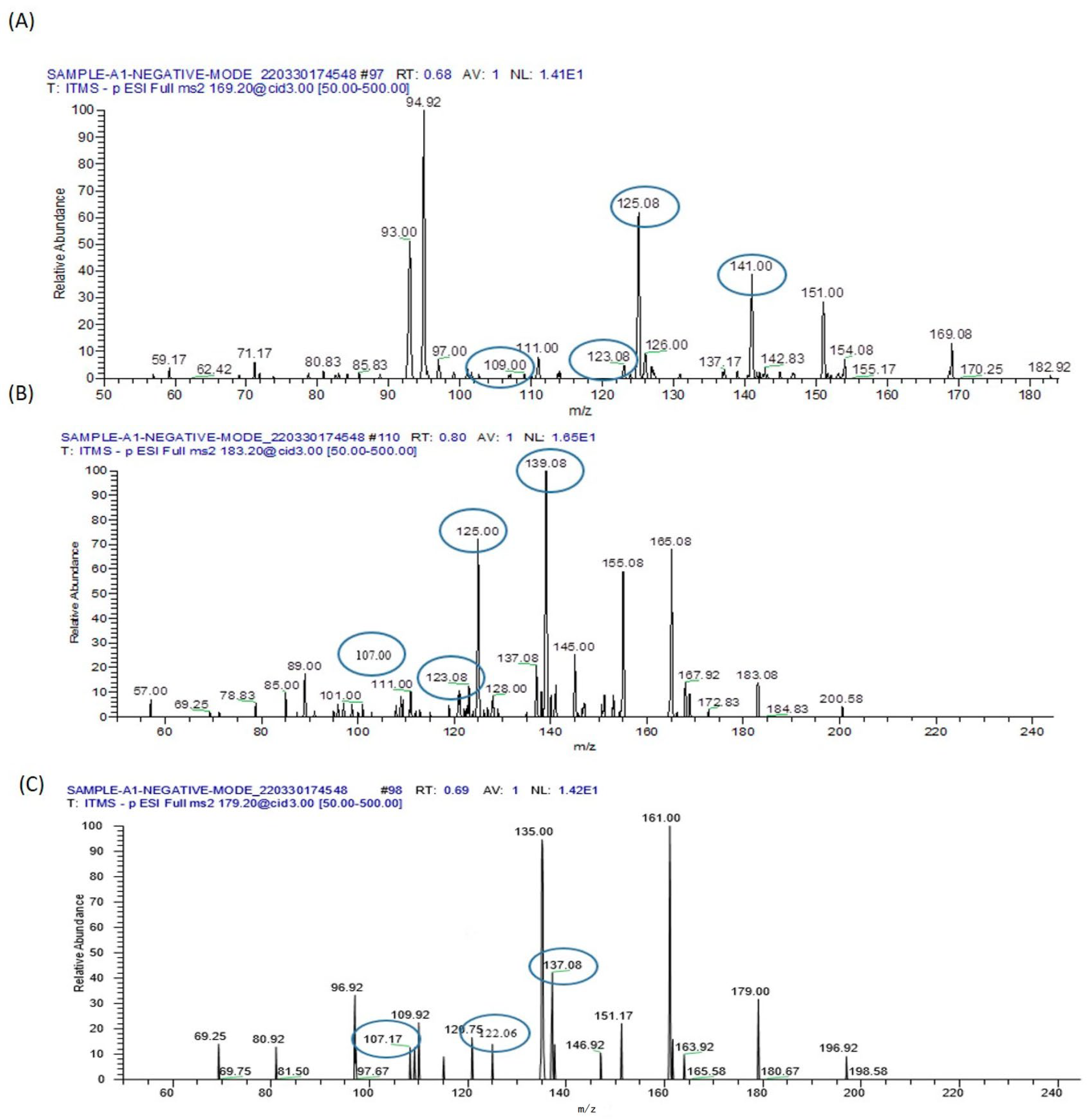
| Arsenate (AsV) | [M-H]− |  | 141 | 123 | AsO3− |
| Arsenite (AsIII) | [M-H]− |  | 125 | 109 | As(OH)2− |
| Monomethyl arsenic acid (MMAV) | [M-H]− |  | 139 | 125 123 121 107 | AsO3H− AsO2CH2− AsO2− |
| Dimethylarsi-nous acid (DMAIII) | [M-H]− |  | 137 | 107, 122 | AsO2− CH3AsO2− |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shoaib, S.M.; Afzal, S.; Feezan, A.; Akash, M.S.H.; Nadeem, A.; Mir, T.M. Metabolomics Analysis and Biochemical Profiling of Arsenic-Induced Metabolic Impairment and Disease Susceptibility. Biomolecules 2023, 13, 1424. https://doi.org/10.3390/biom13091424
Shoaib SM, Afzal S, Feezan A, Akash MSH, Nadeem A, Mir TM. Metabolomics Analysis and Biochemical Profiling of Arsenic-Induced Metabolic Impairment and Disease Susceptibility. Biomolecules. 2023; 13(9):1424. https://doi.org/10.3390/biom13091424
Chicago/Turabian StyleShoaib, Syed Muhammad, Samina Afzal, Ali Feezan, Muhammad Sajid Hamid Akash, Ahmed Nadeem, and Tahir Maqbool Mir. 2023. "Metabolomics Analysis and Biochemical Profiling of Arsenic-Induced Metabolic Impairment and Disease Susceptibility" Biomolecules 13, no. 9: 1424. https://doi.org/10.3390/biom13091424
APA StyleShoaib, S. M., Afzal, S., Feezan, A., Akash, M. S. H., Nadeem, A., & Mir, T. M. (2023). Metabolomics Analysis and Biochemical Profiling of Arsenic-Induced Metabolic Impairment and Disease Susceptibility. Biomolecules, 13(9), 1424. https://doi.org/10.3390/biom13091424